










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.62 n.1 San José Jan./Mar. 2014
Efectos de la estructura del paisaje y de la vegetación en la diversidad de murciélagos filostómidos (Chiroptera: Phyllostomidae) de Oaxaca, México
Effects of landscape and vegetation structure on the diversity of phyllostomid bats (Chiroptera: Phyllostomidae) in Oaxaca, México
Effects of landscape and vegetation structure on the diversity of phyllostomid bats (Chiroptera: Phyllostomidae) in Oaxaca, México
*Dirección para correspondencia:
Abstract
The tropical forest fragmentation is known to affect the spatial structure of the landscape and habitat. These alterations can modify the attributes of bat assemblages, however, this phenomenon has been little studied and understood. In this work we evaluated the structure of landscape (i.e. composition and configuration) and vegetation, and its relationship with assemblage- and population-level characteristics of phyllostomid bats in a tropical rainforest of Southeastern Mexico. For this, we previously selected 12 sites located in continuous and fragmented forests, where bats were captured using mist nets during a two years sampling effort (144 nights). Bats relative abundance, species richness (diversity of order 0, 0D), Shannon diversity index (1D) and Simpson index (2D) were evaluated in all sites, and their relationship with seven measures of landscape structure and seven measures of vegetation structure was described using a Hierarchical Partitioning Analysis. A total of 1 840 individuals of 29 species of phyllostomid bats were captured in this period. Differences in the assemblages were manifested only in the relative abundance and not in the richness of the species. The assemblages of fragmented forest exhibited greater variation in species composition and a greater abundance of frugivorous and nectarivorous bats in comparison with the assemblages of continuous forest. The landscape configuration was related to the assemblage- and population-level attributes, contrasting with previous studies where the composition was a key element. At habitat level, tree density and canopy cover determined the abundance of bats. Nectarivorous and frugivorous bats were mostly found in disturbed vegetation landscapes, primarily due to landscape configuration (e.g. edge density). This phenomenon could be a response to the availability of food in primary and intermediate successional stages, which are characterized by an abun-dance of food value.
Key words: Mexico, Neotropical bats, landscape configuration, Los Chimalapas, spatial scale.
Resumen
La fragmentación de bisques tropicales altera la estructura especial del paisaje y del habitat . Estas alteraciones pueden modificar los atributos de las agregaciones de murciélagos, sin embargo este fenómeno ha sido poco estudiado y comprendido. Se evaluó la estructura del paisaje (i.e. composición y configuración) y vegetación, y sus relaciones con características a nivel de agregación (ensamble) y población de murciélagos filostómidos en una selva tropical del sureste de México. Se encontró que las modificaciones en las agregaciones solo se manifiestan en la abundancia relativa y no en la riqueza de especies. La configuración del paisaje fue un elemento relacionado con los atributos a nivel de ensamble y de población, contrastando con estudios previos donde la composición fue un elemento clave. A nivel de hábitat se encontró que la densidad arbórea y cobertura del dosel determinan la abundancia de murciélagos. Los murciélagos nectarívoros y frugívoros prefieren paisajes con vegetación alterada y están relacionados principalmente con la configuración del paisaje. Este fenómeno podría ser una respuesta a la disponibilidad de alimento en ambientes sucesionales primarios e intermedios, que se caracterizan por una alta proliferación de plantas con potencial alimenticio.
Palabras clave: México, murciélagos neotropicales, configuración del paisaje, los chimalapas, escalas espaciales.
Las transformaciones que han sufrido los ecosistemas naturales en el último siglo han sido dramáticas, en gran medida por la expansión de las poblaciones humanas en busca de nuevas zonas habitables y recursos naturales (Erlich & Wilson 1991; Vitousek et al. 1997). Estas transformaciones se han manifestado como pérdida y fragmentación de extensas regiones de vegetación nativa (Vitousek et al. 1997). La fragmentación se define como la transformación de un hábitat de gran tamaño en numerosos fragmentos de área menor, aislados por una matriz de hábitat distinto al original (Wilcove et al. 1986; Andrén 1994) y es considerado un proceso de degradación ambiental humana-inducida a escala de paisaje (McGarigal & Cushman 2002; Haila 2002). Se ha observado que taxa distintos responden de manera diferencial a la fragmentación del hábitat (Fahrig 2003). La descripción de las consecuencias de este fenómeno en murciélagos han sido cualitativos y poco claros (Fenton et al. 1992; Estrada et al. 1993, Cosson et al. 1999; Medellín et al. 2000), en gran medida porque las aproximaciones no han sido adecuadas (Gorresen & Willig 2004). Se ha recomenda-do que evaluaciones que describan elementos explícitos de los paisajes fragmentados como el tamaño, forma, grado de aislamiento, efecto de borde y estructura de la matriz (Ewers & Didham 2006), pudieran ser adecuados para determinar los efectos de la fragmentación en murciélagos (Gorresen & Willig 2004; Meyer & Kalko 2008; Klingbeil & Willig 2009).
Los murciélagos representan un elemento de gran importancia de la biodiversidad de mamíferos en la zona Neotropical (Kalko 1998) y es un grupo que se encuentra amenazado por la fragmentación y destrucción de los bosques tropicales (Racey & Entwistle 2003; Meyer & Kalko 2008). De los murciélagos que se distribuyen en la zona Neotropical, la familia Phyllostomidae constituye el grupo más diverso en número de especies y en abundancia (Timm 1994; Estrada & Coates-Estrada 2002; Meyer & Kalko 2008) y posee una alta diversificación trófica, que les confiere un valioso papel en los ecosistemas como polinizadores y dispersores de semillas, lo que ayuda a mantener la diversidad de plantas, promover la sucesión secundaria y la regeneración de los bosques (Kalko 1998; Patterson et al. 2003). Los murciélagos filostómidos conforman un taxón ideal para probar los efectos de la fragmentación, ya que poseen una alta riqueza de especies, alta diversificación ecológica, variadas adaptaciones tróficas, y una considerable movilidad que les permite cubrir extensas zonas de paisajes fragmentados (Meyer & Kalko 2008).
Existe una respuesta diferencial de vulnerabilidad de las especies murciélagos de distintos gremios tróficos a los disturbios en el hábitat (Meyer & Kalko 2008; Meyer et al. 2008). Así, especies generalistas, como los murciélagos frugívoros de dosel (Artibeus spp.) y sotobosque (Carollia spp. y Sturnira spp.) son menos susceptibles a cambios en la estructura del paisaje, debido en gran medida a su capacidad de explotar recursos alimenticios tanto en bosques continuos como en ambientes perturbados (Laurance 1991; Cos-son et al. 1999, Medellín et al. 2000; Estrada & Coates-Estrada 2002; Evelyn & Stilles 2003; Willig et al. 2007; Loayza & Loiselle, 2008; Klingbeil & Willig 2009), mientras que murciélagos carnívoros y algunos nectarívoros con poca movilidad prefieren forrajear dentro de bosques continuos (Fenton et al. 1992; Medellín et al. 2000; Meyer et al. 2008a; Meyer & Kalko 2008; Klingbeil & Willig 2009).
Estudios recientes sugieren que la fragmentación altera la estructura espacial del paisaje y con ello la abundancia, distribución y la dinámica de las poblaciones animales (Wiens 1989). Así también distintos taxones responden de manera distinta a la composición y configuración del paisaje (McGarigal & McComb 1995; Villard et al. 1999). Los elementos de composición del paisaje tienen mayor poder de predicción que la configuración en la abun-dancia a nivel de ensamble y población en aves y murciélagos en paisajes fragmentados (McGarigal & McComb 1995; Villard et al. 1999; Gorresen & Willig 2004; Meyer & Kalko 2008; Klingbeil & Willig 2009). De manera particular, se ha observado que murciélagos frugívoros responden a las características de la composición del paisaje, mientras que las especies animalívoras son afectadas por cambios en la configuración (Gorresen & Willig 2004; Klingbeil & Willig 2009). Estas respuestas especie-específicas y ensamble-especificas muestran dependencia de la escala espacial de análisis (Wiens 1989; Lyons & Willig 2002; Steffan-Dewenter et al. 2002; Gorresen et al. 2005; Klingbeil & Willig 2009).
A nivel local se ha observado que la fragmentación de hábitat está asociada con cambios drásticos en la composición florística y estructura de la vegetación (Harper et al. 2005; Meyer & Kalko 2008; Medellín et al. 2000; Castro-Luna et al. 2007). Se ha descrito que modificaciones en la estructura de la vegetación original se relacionan con el aumento en la riqueza y abundancia de murciélagos filostómidos en estas áreas perturbadas, principalmente de gremios frugívoros y nectarívoros (Castro-Luna et al. 2007; Willig et al. 2007). Así también elementos como la densidad arbórea y cobertura del dosel están asociados a los cambios en abundancia, actividad y distribución vertical de los murciélagos (Ford et al. 2005; Vargas-Contreras et al. 2009; João et al. 2010, Rex 2011).
Por lo anterior, este estudio tuvo el objetivo de evaluar la relación de la estructura del paisaje (i.e. composición y configuración) y del hábitat (i.e. estructura de la vegetación) con las características a nivel de ensamble (abundancia relativa, riqueza de especies 0D, índice de diversidad de Shannon 1D y dominancia de Simpson 2D) y a nivel de población (abundancia de especie) de murciélagos filostómidos en una selva tropical del sureste de México.
Materiales y Métodos
Área de estudio: Se ubica entre las coordenadas 16º55’-16º47’ N - 94º50’-94º40’ W (Fig. 1), en altitudes menores a los 1 000m sobre el nivel del mar (msnm), en la selva tropical de Los Chimalapas, conformado por los municipios Santa María y San Miguel Chimalapa, en el estado de Oaxaca, México. Esta selva se encuentra dentro de la Región Terrestre prioritaria Selva Zoque-La Sepultura (Anaya & Álvarez 1994; Salas-Morales et al. 2001). El clima es principalmente cálido-húmedo, y la vegetación predominante Selva Alta Perennifolia (SAP) (Salas-Morales et al. 2001) y se caracteriza por árboles de más de 30m de altura y que mantienen sus hojas todo el año; los estratos arbóreo y arbustivo están conformados por especies como Guatteria anomala, Dialium guianense, Terminalia amazonia, Sloanea tuerckheimii, Brosium alicastrum, Luehea spp., Pipper spp., Guarea spp., Psichotria sp., Quararibea sp. y Miconia sp. (Salas-Morales et al. 2001). Datos de precipitación y temperatura de 29 años, indican que la estación seca va de diciembre a mayo, con promedios mensuales de precipitación y temperatura de 84.0mm y 23.8°C, mientras que en la estación lluviosa, de junio a noviembre, los valores son de 302.6mm y 25.5°C, respectivamente (IMTA 1999).
Selección de sitios: Se procesaron imágenes de satélite ortorectificadas del área de estudio (row 23, path 48) del Sensor Landsat 7 ETM+ de mayo de 2007 (USGS, http://glovis.usgs.gov/). Las bandas (3-5 y 8) fueron unidas, y se realizó un realce espectral a la imagen multibanda usando el Índice de Vegetación Diferencial Normalizado (NDVI por su siglas en inglés) (Sellers et al. 1992, Pettorelli et al. 2005). Al mapa resultante se le aplicó una clasificación supervisada, generando dos categorías: zonas con vegetación y zonas sin vegetación aparente con el programa ERDAS IMAGINE v9.2 (ERDAS Inc., Atlanta, GA, USA). De esta forma se generó un mapa base para la selección de sitios.
Se seleccionaron 33 áreas circulares de 4km de diámetro sobre el mapa base, como primer paso para la selección de sitios potenciales. Estas áreas fueron ubicadas por accesibilidad cercana a la carretera principal que comunica al poblado de Santa María Chimalapa (información disponible bajo petición al primer autor). El tamaño de estas áreas circulares representa un compromiso entre áreas pequeñas donde las características biogeográficas no difieran entre los sitios, pero puedan considerarse unidades muestreales estadísticamente independientes (separadas por >1km) (Gorresen & Willig 2004; Gorresen et al. 2005; Meyer & Kalko 2008; Klingbeil & Willig 2009). Se recortaron las áreas circulares en programa ArcGIS 9.3 (ESRI, Redlands, CA, USA) y para cada una de ellas se obtuvieron siete medidas del paisaje aplicadas previamente para describir grupos con variada movilidad como aves y murciélagos (McGarigal & McComb 1995; Gorresen & Willig 2004; Klingbeil & Willig 2009): tres de la composición (cobertura de bosque en %; densidad de fragmentos, fragmentos/100ha; y tamaño medio de los fragmentos medidos en ha) y cuatro de la configuración (densidad de borde, m/ha; forma media de fragmentos, medida adimensional obtenida a través de una forma cuadrada teórica; índice de proximidad media, medida adimensional; y distancia media euclidiana al vecino más cercano, m) (McGarigal & Cushman 2002; Leitão et al. 2006). Las medidas fueron calculadas con el programa FRAGSTATS v3.3 (McGarigal et al. 2002). Las áreas potenciales fueron caracterizadas por las medidas de estructura de paisaje (información disponible bajo petición al primer autor) y fueron ordenados por medio de un Análisis de Componentes Principales (ACP) en el programa NCSS/PASS 2001 (NCSS Statistical Software, Kaysville, Utah) a partir de una matriz de correlación de datos estandarizados (Media=0 y DE=1).
Los dos primeros componentes del ACP explicaron el 64.8% de la variación en los datos (CP1=50.1%, CP2=14.7%). El ACP muestra que las áreas circulares potenciales pueden separarse (principalmente por el CP1) en bosques continuos con mayor proporción de cobertura de bosque (>84.96%), baja densidad de fragmentos (<1.27 fragmentos/100ha) y menor densidad de borde (<38.59m/ha), y en bosques fragmentados con baja cobertura de bosque (<75.87%), alta densidad de fragmentos (>2.46 fragmentos/100ha) y mayor densidad de borde (>62.07m/ha) (Apéndice 1). Considerando los resultados del ACP se eligieron 12 de las 33 áreas circulares iniciales, seis con vegetación continua y seis con vegetación fragmentada. Los sitios de captura de murciélagos se ubica-ron cercanos al centro geométrico de las áreas circulares (Fig. 1).
Captura de murciélagos: El estudio fue conducido desde marzo 2010 a febrero 2012. Se cubrieron cuatro estaciones (dos temporadas secas y dos lluviosas) durante el estudio. Cada uno de los 12 sitios fue muestreado aleatoria-mente durante tres noches en cada estación. El intervalo de tiempo entre cada sesión de captura en cada sitio fue de seis meses. En total se realizaron capturas durante 144 noches (12 sitios*3 noches por estación*4 estaciones). En cada sitio se emplearon ocho redes de niebla de (6x2.5m), separadas aproximadamente por 20m, ubicadas en línea recta sobre los senderos de vegetación. Las redes fueron revisadas cada 30min durante un periodo aproximado de 8hr (18:00hr-02:00hr). Cinco redes se colocaron a nivel del sotobosque (<4m) y tres a nivel superior de dosel (>4m). El esfuerzo de captura promedio en cada sitio fue de 10 420.8m2r•hr (metros cuadrados de red-hora) (DE=1 388.18), en total se aplicó un esfuerzo de 125 049.6m2r•hr.
Cada ejemplar fue identificado siguiendo las claves para murciélagos de Álvarez et al. (1994) y Medellín et al. (1997), y se clasificaron siguiendo la nomenclatura de Simmons (2005). Todos los ejemplares adultos fueron marcados con collares de nylon numerados (Gannon 1994, García-García et al. 2010), para no ser contados más de una ocasión. Los gremios tróficos fueron determinados a partir de la literatura (Freeman 1998; Wetterer et al. 2000; Nogueira & Peracchi 2003; Giannini & Kalko 2004; Tschapka 2004; Kalka & Kalko 2006; da Silva et al. 2008) (Apéndice 2).
Caracterización de la estructura del paisaje: Para evaluar la respuesta de los murciélagos filostómidos a la estructura del paisaje a nivel de ensamble y población, se diseñó un sistema de escala anidada (Gorresen & Willig 2004; Gorresen et al. 2005), estableciendo tres círculos concéntricos (1, 2.5 y 4km de diámetro) al punto de muestreo. Este diseño responde al escaso conocimiento de la escala adecuada de análisis para murciélagos filostómidos y la información incompleta del ámbito de acción y movimiento de este grupo para el área de estudio. El valor máximo fue determinado tomando en cuenta el criterio de la distancia mínima entre los sitios de muestreo y el más pequeño tomando en cuenta el posible ámbito de acción y movimiento nocturno de especies de talla pequeña y media previamente descritas, como Glossophaga spp. (movimiento promedio por noche<1 081m), Artibeus spp. de tamaño pequeño (antes Dermanura spp., ámbito de acción< 9ha) y Platyrrhinus helleri (movimiento promedio por noche<1 000m) (Lemke 1984; Fenton et al. 2001; Albrecht et al. 2007; Chaverri et al. 2007; Medina et al. 2007). En cada círculo anidado se obtuvieron las siete medidas de la estructura del paisaje mencionados en el apartado de selección de sitios.
Caracterización de la estructura de la vegetación: En cada uno de los 12 sitios de muestreo se estableció un transecto de 100m, y se realizó un muestreo de cuadrantes centra-dos en un punto cada 10m (Mueller-Dombois & Ellenberg 1974). En el centro se trazaron dos líneas imaginarias perpendiculares con las que se obtuvieron cuatro cuadrantes. Dentro de cada uno se consideró al arbusto (>1.5m y <3m altura) y árbol (>3m) más cercano al centro y se midió su distancia al centro, la altura (mediante un clinómetro marca Brunton), cobertura y diámetro a la altura de pecho (DAP) (Mueller-Dombois & Ellenberg 1974). Se estimó la densidad (m2) arbórea y herbácea como 100/distancia media de la planta más cercana al centro del cuadrante. La cobertura (m2) con la aproximación de la elipse (d1+d2/4)2*π, donde d1 es el diámetro mayor de la copa y d2 diámetro perpendicular a d1. El área basal (m2) se estimó únicamente para árboles como π*(DAP2/4).
Se midió la riqueza de especies (0D), diversidad de orden 1 (el índice de Shannon-Wiener, 1D) y diversidad de orden 2 (índice de Simpson, 2D) en cada sitio, en bosques continuos, fragmentados y en conjunto. Estas medidas fueron expresadas en términos de número efectivo de especies (Hill 1973, Jost 2006). Los cálculos se realizaron con el paquete Bio-diversityR (Kindt & Coe 2005) ejecutado en la plataforma R v2.14.0 (R Development Core Team 2009). La abundancia relativa se expresó dividiendo el número de ejemplares de cada especie capturado entre el esfuerzo de muestreo (metros cuadrados de red-hora y multiplicados por 100) aplicado en cada sitio (Straube & Bianconi 2002). Se realizaron comparaciones de la abundancia relativa y diversidad entre ambos complejos de vegetación por medio de una prueba de t-Student para dos muestras.
Se evaluó la representatividad de los inventarios en cada sitio a través de curvas de acumulación de especies basadas en muestras (Gotelli & Colwell 2001). Las curvas se obtuvieron mediante la elaboración de matrices de presencia-ausencia que se aleatorizaron 100 ocasiones para eliminar el efecto del orden específico en que ingresan los datos en la construcción de las curvas, en el programa EstimateS versión 8.2.0 (Colwell 2011). Adicionalmente se calculó el número esperado de especies que ocurren en cada sitio mediante la aplicación del estimador no paramétrico Jackknife de primer orden, que tiene un buen desempeño de sesgo, precisión y exactitud (Walther & Moore 2005), y es recomendado para taxones móviles (Brose & Martínez 2004). La completitud de los inventarios se obtuvo midiendo el porcentaje que representan las especies observadas respecto a las esperadas (Moreno & Halffter 2000).
Se empleó un análisis de correspondencia (AC) para evaluar las diferencias en composición de especies entre bosques continuos y fragmentados. El AC permite evaluar la asociación de la abundancia de las especies (filas) y las agregaciones de cada sitio (columnas) y representarlas en un espacio de pocas dimensiones (McCune & Grace 2002; Hair et al. 2009). Esta técnica multivariada es apropiada cuando las respuestas de las especies a los gradientes ambientales subyacentes podrían ser únicas y bien determinadas (forma de campana) (ter Braak 1987; ter Braak & Verdonschot 1995) o bien cuando la interpretación de los elementos de la matriz (filas y columnas) necesita hacerse en conjunto (McCune & Grace 2002).
Se analizó la relación entre las siete variables de la estructura del paisaje y las características de las agregaciones (abundancia relativa y diversidad 0D, 1D y 2D) y a nivel poblacional (únicamente para especies presentes en más de seis sitios y representadas por más de 10 individuos). Se empleó la técnica de Partición Jerárquica (Modelo Lineal Generalizado) (Chevan & Sutherland 1991), que tiene como objetivo generar una base estadística detallada para infe-rir la causalidad en regresiones multivariadas y no para identificar un submodelo óptimo o una ecuación predictiva (Chevan & Sutherland 1991; Watson & Peterson 1999; Mac Nally 2000, 2002; Smith et al. 2009). Basa sus cálculos en la medición del efecto de cada variable de manera independiente y en conjunto con las otras variables (Mac Nally 2000, 2002). Esta técnica es apropiada cuando existen problemas de multicolinearidad de variables y cuando se emplean <10 variables explicativas en los cálculos, como en este caso (Chevan & Sutherland 1991, Olea et al. 2010).
Los cálculos se realizaron con el paquete hier.part v1.0-1 (Mac Nally & Walsh 2004) en la plataforma R v2.14.0. La riqueza de especies (conteos) fue modelada usando una aproximación de errores tipo Poisson y para el resto de las variables de respuesta fueron errores tipos Gaussiano. La significancia estadística (p≤0.05) de las contribuciones independientes de las variables fue probada mediante una rutina de aleatorización (1 000 iteraciones) (hier.part v1.0-1). Se aplicó el mismo procedimiento estadístico empleado en la evaluación de los efectos de la estructura del paisaje para analizar el efecto de cuatro características arbustivas y arbóreas de la vegetación (densidad, altura, cobertura y área basal) en cada sitio y su efecto a nivel de ensamble y población.
Resultados
Diversidad, abundancia y composición de agregaciones: Se capturaron 1 840 ejemplares de 29 especies de murciélagos filostómidos (Cuadro 1). En bosques continuos se capturaron 747 ejemplares de 25 especies (esfuerzo=69 139.2m 2r•hr) y en fragmentados 1 093 ejemplares de 25 especies (esfuerzo=55 910.4m2r•hr). Los bosques continuos (Media=1.08, DE=0.273) y fragmentados (Media=1.96, DE=0.66) difirieron significativamente en la abundancia relativa (t-Student=3.04, p=0.012), pero no en la riqueza (0D) de especies (t-Student=1.40, p=0.189). Los bosques continuos (Media=9.8, DE=1.72) presentaron valores mayores, pero no signi-ficativos de diversidad 1D (t-Student=1.79, p=0.102), respecto a fragmentados (Media=7.9, DE=2.071). Los valores de diversidad 2D de bosques fragmentados (Media=6.6, DE=2.22) y continuos (Media=7.8, DE=2.21) fueron similares (t-Student=0.92, p=0.377). El valor de completitud promedio en bosques continuos (78.4%) fue ligeramente mayor a fragmentados (77.15%). Las especies más abundantes en bosques continuos fueron Carollia sowelli (abundancia relativa 1.6, 24.9%), Artibeus jamaicensis (0.8, 13.1%), Dermanura watsoni (0.7, 11.1%) y Sturnira lilium (0.4, 7.4%), en conjunto representaron el 56.5% de las capturas. Mientras que en fragmentados, S. lilium (2.1, 18%), C. sowelli (1.6, 14%), A. lituratus (1.6, 14%) y Glossophaga morenoi (1.4, 12%) representan el 58% de las capturas.
En bosques continuos se registraron cuatro especies exclusivas, A. aztecus, Glyphonycteris sylvestris, Hylonycteris underwoodi y Vampyrum spectrum, y en fragmentados también se registraron cuatro especies exclusivas, Desmodus rotundus, Lonchorhina aurita, Micronycteris schmidtorum y Uroderma bilobatum. Ambos complejos de vegetación comparten 21 especies (Fig. 2), de los cuales S. lilium (t-Student=4.851, p<0.001) y G. soricina (t-Student=2.210, p=0.051) (marginalmente significativo) fueron más abundante en bosques fragmentados que en continuos. Caso contrario sucede con S. ludovici (t-Stu-dent=2.388, p=0.038) y Phyllostomus discolor (t-Student=2.178, p=0.054) (marginalmente significativo), que son más abundantes en bosques continuos.
Los especímenes capturados representa-ron seis gremios tróficos (Cuadro 1). Los murciélagos frugívoros especialistas de Ficus (FF, S=11 especies, n=682 individuos), frugívoros especialistas de Piper (FP, S=5, n=638) y nectarívoros (Nec, S= 6, n=342) fueron los más importantes en riqueza de especies y abundancia; mientras que los omnívoros (Omn, S=2, n=54), animalívoros (Anm, S=4, n=23) y hematófagos (Hem, S=1, N=58) fueron menos destacados. Los bosques fragmentados muestran mayor abundancia de gremios FF, FP y Nec que los continuos (Fig. 3). Sin embargo la abundancia de todos los gremios no difiere entre ambos complejos (t-Student=0.960, p=0.359). Las diferencias de los gremios tróficos (riqueza y abundancia) entre sitios en cada complejo tampoco son significativas (p>0.05).
El AC indica que las dos primeras dimensiones (ejes) explican el 45.7% de la variación en los datos (inercia total=1.018, X²=1873.8, g.l.=308, p<0.001). A pesar que el CA tiene bajo poder explicativo se emplea para dilucidar los patrones en la composición de las agregaciones. La primera dimensión muestra una separación de agregaciones por su ubicación en bosques continuos o fragmentados (Fig. 4), y la segunda muestra una separación de especies por su afinidad de hábitat. También se distinguen dos grupos, el primero conformado de tres sitios de bosques continuos (C6, C2 y C5) y con predominio de Phyllostomus discolor, Artibeus phaeotis, Glossophaga commisarisi, G. leachii y Centurio senex, especies con mayor presencia y abundancia relativa en bosques continuos. El segundo grupo lo componen los sitios F3, F6 y C4, con un predominio de A. watsoni, C. perspicillata y Plathyrrhinus helleri. Las dos primeras especies presentes en bosques continuos y fragmentados, pero con mayor abundancia en el último. P. helleri también está presente en ambos complejos, pero es más abundante en los continuos. El resto de las agregaciones no muestran una agrupación clara.
Estructura del paisaje: A nivel de ensamble la abundancia relativa, diversidad 1D y 2D responden a características de la configuración del paisaje (Cuadro 2). La diversidad1D y 2D muestran una respuesta negativa a la densidad de borde y forma de los fragmentos a escala grande (4km de diámetro) e intermedia (2.5km). La abundancia relativa responde positivamente a la proximidad de los fragmentos a escala pequeña (1km). A nivel poblacional (abundancia relativa de las especies), se encontró que los murciélagos nectarívoros responden positivamente a características de configuración del paisaje (forma, proximidad y distancia media) y negativamente a las de composición (cobertura de bosque y tamaño medio de fragmento). Los miembros del genero Carollia (FP) responden a la cobertura de bosque (a todas las escalas), proximidad y distancia media de fragmentos, mientras que las especies del género Sturnira responden positivamente a la densidad de borde y forma de fragmentos a escalas grande e intermedia. Los murciélagos FF son afectados principalmente por variables de configuración del paisaje, de manera positiva por la forma, proximidad y distancia de fragmentos, y negativamente por la densidad de borde. En menor medida son afectados por la densidad (Artibeus lituratus) y tamaño medio de fragmentos (Vampyrodes major). Murciélagos omnívoros muestran poco efecto de la estructura del paisaje, solo Centurio senex es afectada negativamente por la densidad de borde a escala grande.
Estructura de la vegetación: La estructura de la vegetación de bosques continuos se caracterizó por mayor densidad arbórea y arbustiva (información disponible bajo petición al primer autor), mayor cobertura arbórea y altura de dosel (arbórea). Los bosques fragmentados se caracterizan por menor densidad arbórea y arbustiva y altura de dosel, pero mayor cobertura arbustiva. La abundancia relativa de murciélagos muestra un efecto de la densidad (positivo) y cobertura arbórea (negativo) a nivel de ensamble (Cuadro 3). Glosshopaga morenoi y G. soricina (Nec) muestran efecto negativo de la densidad arbórea y positivo de la cobertura arbustiva, respectivamente. La densidad (negativo) y altura del dosel (positivo) son características que influyen en la abundancia de Carollia sowelli y Sturnira lilium (FP), respectivamente. S. ludovici es afectada positivamente por la cobertura arbustiva. Los murciélagos FF son afectados por la densidad arbustiva (Artibeus watsoni y Vampyrodes major) y área basal (Plathyrrhinus helleri), mientras que las especies omnívoras no muestran un efecto de la estructura de la vegetación.
Discusión
Contrario a lo encontrado en bosques altamente fragmentados (Estrada et al. 1993; Cosson et al. 1999; Medellín et al. 2000; Meyer & Kalko 2008), los efectos de la fragmentación se manifiestan como modificaciones en la abundancia relativa de las especies. Este patrón concuerda con lo encontrado en paisajes con fragmentación moderada (Gorresen & Willig 2004; Clarke et al. 2005a, b; Willig et al. 2007; Klingbeil & Willig 2009; Presley et al. 2009). Los bosques fragmentados muestran mayor abundancia de murciélagos frugívoros, en particular de especies que se alimentan de frutos de plantas de estados sucesionales tempranos o medios como Sturnira lilium y Artibeus lituratus (Fleming & Heithaus 1986; Fleming 1988; García et al. 2000; Faria 2006; Willig et al. 2007), así como la presencia de nectarívoros tolerantes a áreas perturbadas como Glossophaga morenoi y G. soricina (Arita & Santos-del-Prado 1999; Estrada & Coates-Estrada 2002; Clarke et al. 2005a; Willig et al. 2007).
Entre las especies registradas solo en bosques continuos se encontró a Vampyrum spectrum, un murciélago carnívoro exclusivo de bosques tropicales en buen estado de conservación (López et al. 1998; Vehrencamp et al. 1977), así como Glyphonycteris sylvestris, Hylonycteris underwoodi y Artibeus aztecus, murciélagos localmente poco abundantes con preferencias por ambientes conservados (Estrada & Coates-Estrada 1993; Arita & Santos-del-Prado 1999; Clarke et al. 2005a; Castro-Luna et al. 2007; Willig et al. 2007). En bosques fragmentados, Desmodus rotundus y Lonchorhina aurita fueron especies exclusivas con alta abundancia relativa, la primera asociada por sus hábitos alimenticios a sitios perturbados con ganadería (Medellín et al. 2000; Bernard & Fenton 2002; Estrada & Coates-Estrada 2002) y la segunda, un murciélago perchador de follaje que probablemente prefiere forrajear en áreas abiertas o en el borde de la selva (Klingbeil & Willig 2009).
El AC muestra mayor variación en composición de las agregaciones de bosques fragmentados, formándose un gradiente de composición, desde agregaciones que podrían ser típicos de bosques fragmentados a agregaciones fragmentadas que muestran una composición parecida a los de bosques continuos. Estas variaciones en composición de agregaciones se han documentado en paisajes fragmentados de reciente formación (Gorresen & Willig 2004; Klingbeil & Willig 2009) y se ha propuesto que estos sistemas no han alcanzado cierta estabilidad en sus atributos (e.g. riqueza, abundancia especies), para que las diferencias con las agregaciones de bosques continuos sean evidentes (Ewers & Didham 2006; Meyer & Kalko 2008; Klingbeil & Willig 2009).
La estructura del paisaje y sus relaciones con las características a nivel de ensamble y población muestran efecto de la escala espacial, con mayor poder de predicción en la escala media y grande. Estos resultados refuerzan las evidencias de una respuesta diferencial en murciélagos a distintas escalas de paisaje (Gorresen & Willig 2004; Meyer & Kalko 2008; Klingbeil & Willig 2009). La configuración del paisaje es un elemento determinante en los atributos a nivel de agregaciones y se observa un efecto negativo de la densidad de borde sobre la diversidad 1D de las agregaciones. Este patrón se ha observado en ambientes fragmentados, donde la sensibilidad al borde de las especies está asociado fuertemente con la presencia y abundancia en los fragmentos de vegetación (Meyer et al. 2008). También se observó un efecto significativo sobre la abundancia y diversidad 2D en las agregaciones de otros elementos de configuración, como la proximidad entre fragmentos (positivo) y su forma (negativo).
Los murciélagos nectarívoros presentan mayor abundancia en bosques fragmentados y están afectados por la configuración y composición de éste. Esta observación es ilustrada por dos tendencias, la primera es observada con G. soricina y G. morenoi que muestran mayor preferencia por ambientes fragmentados donde la proximidad y tamaño de los fragmentos son menores y la distancia entre fragmentos es alta. La otra tendencia es una respuesta negativa a paisajes con mayor cobertura de bosque, pero positiva a paisajes con fragmentos irregulares (basados en un forma cuadrangular teórica y con mayor área interior) y mayor distancia entre fragmentos, como en G. comisarissi y Choeroniscus godmani. El efecto conjunto de variables de composición (cobertura de bosque) y configuración (proximidad) del paisaje en murciélagos nectarívoros ha sido observado previamente en otros paisajes fragmentados (Gorresen & Willig 2004; Klingbeil & Willig 2009). La tendencia de los glosofaginos (i.e. Glossophaga spp.) a forrajear en ambientes perturbados es favorecida por la alta disponibilidad de plantas con flores en estados sucesionales primarios (Faria 2006; Willig et al. 2007; Avila-Cabadilla et al. 2009).
Se ha documentado que los murciélagos frugívoros de sotobosque de los géneros Carollia y Sturnira son comunes en zonas abiertas y área de vegetación secundaria, probablemente en respuesta a la mayor disponibilidad de alimento (Dinerstein 1986; Fleming 1988; Thies & Kalko 2004; Willig et al. 2007). En este estudio se encontró que especies del genero Carollia muestran efectos diversos a la cobertura de bosque, pero preferencia por paisajes con alta proximidad y distancia entre fragmentos. Por otra parte, los murciélagos del género Sturnira son más afines a paisajes con alta densidad de borde y fragmentos de forma irregular. Estas evidencias coinciden parcialmente con lo encontrado en Paraguay con S. lilium y C. perspicillata (Gorresen & Willig 2004).
Los murciélagos FF de talla pequeña (Artibeus watsoni, A. phaeotis y Platyrrhinus helleri) muestran una fuerte influencia de la configuración del paisaje, coincidiendo con lo informado previamente (Klingbeil & Willig 2009), en particular, la densidad de borde (negativo), proximidad y forma (positiva) de fragmentos. Atribuimos estos resultados a la preferencia de estas especies por bosques continuos y a la baja movilidad y ámbitos de acción pequeños (<9ha) (Fenton et al. 2001; Albrecht et al. 2007; Chaverri et al. 2007; Medina et al. 2007). Por otro lado, los murciélagos de tamaño grande como A. lituratus y Vampyrodes major son afectados por la densidad y tamaño de los fragmentos. Se ha observado que estos murciélagos poseen alta movilidad y explotan una serie de recursos (Ficus spp.) en todo el paisaje fragmentado (Morrison 1980; Handley et al. 1991; Estrada & Coates-Estrada 2002; Medina et al. 2007), y la configuración del paisaje podría no ser un elemento determinante como lo es la composición (Meyer & Kalko 2008).
Destaca la influencia de la configuración del paisaje en la abundancia a nivel de ensamble y poblaciones. Estos resultados difieren de lo encontrado en bosques fragmentados de Paraguay y el Amazonas (Gorresen & Willig 2004; Klingbeil & Willig 2009), donde se encontró que elementos de composición del paisaje tienen mayor poder predictivo a nivel de ensamble y población. Este fenómeno puede ser el resultado de dos factores: el patrón de deforestación y la naturaleza de la matriz. Las tasas de deforestación en Paraguay son altas, únicamente permanecen el 20% de los bosques continuos (Keel et al. 1993; Gorresen Willig 2004), mientras que en los bosques del Amazonas ocurre de manera intensiva a pequeña escala (27- 99% de permanecía de bosques) (Klingbeil & Willig 2009). En el presente estudio la cobertura promedio de bosque es mayor a 70% (DE>22), por lo que la deforestación es relativamente baja en todas las escalas, a diferencia de los patrones de los estudios anteriores.
Las características de la matriz circundante a los fragmentos de bosques son determinantes en su conectividad funcional (Antongiovanni & Metzger 2005; Ewers & Didham 2006; Meyer & Kalko 2008). La matriz en el paisaje estudiado se caracteriza por un predominio de pastizales inducidos para la ganadería semi-intensiva y zonas de cultivo itinerante (Salas-Morales et al. 2001). Esto produce un sistema de alto contraste matriz-fragmentos y posiblemente sea poco permeable para el movimiento de especies de murciélagos sensibles a la perturbación. Con estas características de la matriz, la configuración del paisaje es un elemento fundamental para explicar las características de las agregaciones y poblaciones, al contrario de lo que ocurre en los bosques donde la explotación forestal es selectiva y no intensiva o extensiva (Klingbeil & Willig 2009). Cuando la deforestación es extensiva, se presenta una rápida conversión de bosques a tierras agrícolas, se crea un paisaje de islas pequeñas y aisladas de vegetación original (Keel et al. 1993; Gorresen & Willig 2004), donde el comportamiento de las especies es más parecido al de zonas con disturbios antropogénicos a gran escala.
La abundancia de murciélagos filostómidos a nivel de ensamble es afectada por la densidad arbórea y cobertura del dosel. Esta respuesta se expresa en un aumento de la abundancia de murciélagos en función del aumento de la cobertura del dosel y disminución de la densidad arbórea. Previamente se ha enconrado que la cobertura de dosel es un elemento determinante en la abundancia y actividad de murciélagos (Ford et al. 2005; Castro-Luna et al. 2007) y se ha interpretado como una res-puesta al riesgo de depredación o la disponibilidad de alimento (Estrada et al. 1993; Hecker & Brigham 1999; Medellín et al. 2000; Kalko & Handley 2001; Castro-Luna et al. 2007). Posiblemente la abundancia sea el resultado de la interacción de estos dos elementos, sin embargo consideramos que la disponibilidad alimenticia podría ser la causa más importante, ya que la densidad y cobertura del dosel son elementos asociados a ella (Castro-Luna et al. 2007), principalmente por la abundancia de plantas típicas de vegetación secundaria, un recurso alimenticio potencial para murciélagos filostómidos (Charles-Dominique 1986; Fleming 1988; Tschapka, 2004; Vargas-Contreras et al. 2009).
A nivel poblacional se encontraron dos respuestas a la estructura de la vegetación, que determinan la abundancia de las especies de murciélagos. Primero, algunas especies de nectarívoros y FP prefieren hábitats de vegetación perturbada con baja densidad arbórea (G. morenoi y C sowelli) y arbustiva (A. watsoni), baja altura de dosel (S. lilium), pero con alta cobertura arbustiva (G. soricina). Segundo, especies que prefieren hábitats de bosques cerrados (frugívoros especialistas de Ficus), es decir, con alta cobertura (S. ludovici) y baja densidad arbustiva (V. major) y árboles con mayor área basal (P. helleri). La distribución de especies, podría ser el resultado de una combinación de factores, como las características de forrajeo, condiciones microambientales, abundancia y calidad de recursos y la distribución espacial (estratificación espacial) (Kalko & Handley 2001; Bernard 2001; Vargas-Contreras et al. 2009; João et al. 2010; Rex 2011). Se ha documentado que murciélagos nectarívoros y frugívoros especialistas de Piper prefieren forrajear en el sotobosque (Bernard 2001; João et al. 2010), donde el alimento es más abundante (Fleming & Heithaus 1986; Marinho-Filho 1991; Thies & Kalko 2004) y disminuye el riesgo de depredación (Kalko & Handley 2001). Lo contrario sucede con murciélagos FF que están asociados al dosel, donde el alimento se encuentra en mayor cantidad y calidad (Kalko & Handley 2001; Bernard 2001; Giannini & Kalko 2004; Vargas-Contreras et al. 2009).
Destaca la importancia de la configuración del paisaje en las características a nivel de ensamble y población. Contraponiéndose a lo encontrado previamente en murciélagos filostómidos y en otros grupos de animales donde la composición del paisaje es determinante (McGarigal & McCom 1995; Trzcinski et al. 1999; Gorresen & Willig 2004; Heikkinen et al. 2004; Klingbeil & Willig 2009). Se ha documentado que alteraciones en la configuración del paisaje pueden alterar las habilidades de dispersión y disminuir las probabilidades de persistencia de las poblaciones animales (Bélisle 2005), por lo que determinar los efectos de la configuración del paisaje puede ser más importante para contribuir al entendimiento de la conectividad funcional y la dinámica meta-poblacional de los murciélagos. Sin embargo también se ha planteado que la importancia de los elementos del paisaje (composición y configuración) es controversial y pueden variar dependiendo de las características de las especies o el gremio trófico (Villard et al 1999).
Se resalta la alta diversidad y abundancia de murciélagos filostómidos registrados, solo comparable con lo descrito en algunas regiones del sureste de México (Medellín et al. 2000; Estrada & Coates-Estrada 2002; Montiel et al. 2006; Castro-Luna et al. 2007; García-García & Santos-Moreno 2008). En conjunto estas regiones representan el último reservorio de la distribución más norteña de muchos mur-ciélagos filostómidos de origen Neotropical. Por lo que es necesario el establecimiento de estrategias de conservación y reservas ecológicas que aseguren la conservación de los murciélagos en la región. Particularmente en la selva de Los Chimalapas que ha sido centro de graves conflictos sociales por la tenencia de la tierra, además de ser una zona de incendios forestales, elevada actividad ganadera y extracción de árboles maderables (Asbjornsen & Gallardo-Hernández 2004; Asbjornsen et al. 2005), que posiblemente ha derivado en una pérdida aun no cuantificada de la cobertura vegetal y en general de la biodiversidad, en un periodo de tiempo relativamente corto, a partir de la década de 1950 (Ceballos et al. 1998; Watson & Peterson 1999; Salas-Morales et al. 2001; Asbjornsen & Gallardo-Hernández 2004). En el diseño de estas reservas se deben considerar una perspectiva a escalas espaciales múltiples que tomen en cuenta la estructura del paisaje (i.e. composición y configuración) y de la vegetación (hábitat) y su influencia en las agregaciones y poblaciones de murciélagos.
Agradecimientos
El Consejo Nacional de Ciencia y Tecno-logía de México otorgo una beca de estudios (No. 228955) de posgrado al primer autor. Esta investigación tuvo apoyo económico de la Secretaría de Investigación y Posgrado del Instituto Politécnico Nacional de México a través de los proyectos SIP-20100377, SIP-20110395 y SIP- 20120962, y del Programa Institucional de Formación de Investigadores (PIFI). Se agradece las observaciones para la mejora del presente documento de D. Mondragón, G. Ramos, R. del Castillo y R. Solano. Se agradece a las autoridades del municipio de Santa María Chimalapa y a las personas que dieron invaluable asistencia en el trabajo de campo.
Referencias
Albrecht, L., Meyer, C. F. J., & Kalko, E. K. V. (2007). Differential mobility in two small phyllostomid bats, Artibeus watsoni and Micronycteris microtis, in a fragmented neotropical landscape. Acta. Theriologica, 52(2), 141-149. [ Links ]
Álvarez, T., Álvarez-Castañeda, S. T., & López-Vidal, J. C. (1994). Claves para murciélagos mexicanos. México: Instituto Politécnico Nacional. [ Links ]
Anaya, A. L. & Álvarez, M. (1994). Plan de Desarrollo y Conservación de una Reserva Campesina en los Chimalapas. México: Secretaría de Desarrollo Social. Instituto Nacional de Ecología. Programa de Acción Forestal Tropical. [ Links ]
Andrén, H. (1994). Effects of habitat fragmentation on birds and mammals in landscapes with different proportions of suitable habitat: a review. Oikos, 71(3), 55-366. [ Links ]
Antongiovanni, M., & Metzger, J. (2005). Influence of matrix habitats on the occurrence of insectivorous bird species in Amazonian forest fragments. Biological Conservation, 122(3), 441-451. [ Links ]
Arita, H. T., & Santos-del-Prado, K. (1999). Conservation biology of nectar-feeding bats in Mexico. Journal of Mammalogy, 80(1), 31-41. [ Links ]
Asbjornsen, H., & Gallardo-Hernández, C. (2004). Effects of the 1998 fires on tropical montane cloud forests; Chimalapas, Oaxaca. In L. Villers-Ruíz & G. Lopez-Blanco (Eds.), Forest Fires in Mexico: Methods and Evaluation (pp. 125-146). México: Universidad Nacional Autónoma de México. [ Links ]
Asbjornsen, H., Velázquez-Rosas, N., García-Soriano, R., & Gallardo-Hernández, C. (2005). Deep ground fires cause massive above- and below-ground biomass losses in tropical montane cloud forests in Oaxaca, Mexico. Journal of Tropical Ecology, 21(4), 427-434. [ Links ]
Avila-Cabadilla, L. D., Stoner, K. E., Henry, M., & Añorve, M. Y. A. (2009). Composition, structure and diversity of phyllostomid bat assemblages in different successional stages of a tropical dry forest. Forest Ecology and Management, 258(6), 986-996. [ Links ]
Bélisle, M. (2005). Measuring landscape connectivity: the challenge of behavioral landscape ecology. Ecology, 86(8), 1988-1995. [ Links ]
Bernard, E. (2001). Vertical stratification of bat communities in primary forests of Central Amazon, Brazil. Journal of Tropical Ecology, 17(1), 115-126. [ Links ]
Bernard, E., & Fenton, M. B. (2002). Species diversity of bats (Mammalia: Chiroptera) in forest fragments, primary forests, and savannas in central Amazonia, Brazil. Canadian Journal of Zoology, 80(6), 1124-1140. [ Links ]
Brose, U., & Martínez, N. D. (2004). Estimating the rich-ness of species with variable mobility. Oikos, 105(2): 292-300. [ Links ]
Castro-Luna, A. A., Sosa, V. J., & Castillo-Campos, G. (2007). Bat diversity and abundance associated with the degree of secondary succession in a tropical forest mosaic in south-eastern Mexico. Animal Conservation, 10(2), 219-228. [ Links ]
Ceballos, G., Rodríguez, P., & Medellín, R. (1998). Assessing conservation priorities in megadiverse Mexico: mammalian diversity, endemicity, and endangerment. Ecological Applications, 8(1), 8-17. [ Links ]
Charles-Dominique, P. (1986). Inter-relations between frugivorous vertebrates and pioneer plants: Cecropia, birds and bats in French Guyana. In A. Estrada & T. H. Fleming (Eds.), Frugivores and seed dispersal (pp. 119-135). Holanda: Dr. W. Junk Publishers, Dordrecht. [ Links ]
Chaverri, G., Quirós, O. E., & Kunz, T. H. (2007). Ecological correlates of range size in the tent-making bat Artibeus watsoni. Journal of mammalogy, 88(2), 477-486. [ Links ]
Chevan, A., & Sutherland, M. (1991). Hierarchical Partitioning. American Statistician, 45(2), 90-96. [ Links ]
Clarke, F. M., Pio, D. V., & Racey, P. A. (2005). A Comparison of Logging Systems and Bat Diversity in the Neotropics. Conservation Biology, 19(4), 1194-1204. [ Links ]
Clarke, F. M., Rostant, L. V., & Racey, P. A. (2005). Life after logging: post-logging recovery of a neotropical bat community. Journal of Applied Ecology, 42(2), 409-420. [ Links ]
Colwell, R. K. (2011). EstimateS, Version 8.2: Statistical Estimation of Species Richness and Shared Species from Samples (Software and User’s Guide). [ Links ]
Cosson, J. -F., Pons, J. -M., & Masson, D. (1999). Effects of forest fragmentation on frugivorous and nectarivorous bats in French Guiana. Journal of Tropical Ecology, 15(4), 515-534. [ Links ]
Da Silva, A. G., Gaona, O., & Medellín, R. (2008). Diet and trophic structure in a community of fruit-eating bats in Lacandon Forest, Mexico. Journal of Mamma-logy, 89(1), 43-49. [ Links ]
Dinerstein, E. (1986). Reproductive ecology of fruit bats and the seasonality of fruit production in a Costa Rican cloud forest. Biotropica, 18(4), 307-318. [ Links ]
Erlich, P. R., & Wilson, E. O. (1991). Biodiversity studies: science and policy. Science, 253(5021), 758-72. [ Links ]
Estrada, A., & Coates-Estrada, R. (2002). Bats in continuous forest, forest fragments and in an agricultural mosaic habitat-island at Los Tuxtlas, Mexico. Biolo-gical Conservation, 103(2), 237-245. [ Links ]
Estrada, A., Coates-Estrada, R., & Meritt, D. (1993). Bat species richness and abundance in tropical rain forest fragments and in agricultural habitats at Los Tuxtlas, Mexico. Ecography, 16(4), 309-318. [ Links ]
Evelyn, M. J., & Stiles, D. A. (2003). Roosting Require-ments of Two Frugivorous Bats (Sturnira lilium and Arbiteus intermedius) in Fragmented Neotropical Forest. Biotropica, 35(3), 405-418. [ Links ]
Ewers, R. M., & Didham, R. K. (2006). Confounding factors in the detection of species responses to habitat fragmentation. Biological Reviews, 81(1), 117-142. [ Links ]
Fahrig, L. (2003). Effects of Habitat Fragmentation on Biodiversity. Annual Review of Ecology Evolution and Systematics, 34(1), 487-515. [ Links ]
Faria, D. (2006). Phyllostomid bats of a fragmented lands-cape in the north-eastern Atlantic forest, Brazil. Journal of Tropical Ecology, 22(5), 531-542. [ Links ]
Fenton, M., Bernard, E., & Bouchard, S. (2001). The bat fauna of Lamanai, Belize: roosts and trophic roles. Journal of Tropical, 17(4), 511-524. [ Links ]
Fenton, M. B., Acharya, L., Audet, D., Hickey, M. B. C., Merriman, C., Obrist, M. K., Syme, D.M., & Adkins, B. (1992). Phyllostomid bats (Chiroptera: Phyllostomidae) as indicators of habitat disruption in the Neotropics. Biotropica, 24(3), 440-446. [ Links ]
Fleming, T.H. (1988). The Short-tailed Fruit Bat: a Study in Plant-Animal Interactions. Chicago, Illinois, EEUU: University of Chicago Press. [ Links ]
Fleming, T., & Heithaus, E. (1986). Seasonal foraging behavior of the frugivorous bat Carollia perspicillata. Journal of Mammalogy, 67(4), 660-671. [ Links ]
Ford, W., Menzel, M., Rodrigue, J., Menzel, J., & Johnson, J. (2005). Relating bat species presence to simple habitat measures in a central Appalachian forest. Biological Conservation, 126(4), 528-539. [ Links ]
Freeman, P. (1988). Frugivorous and animalivorous bats (Microchiroptera): dental and cranial adaptations. Biological Journal of the Linnean Society, 33(3), 249-272. [ Links ]
Gannon, M. R. (1993). A new technique for marking bats. Bat Research News. 34(4), 88-89. [ Links ]
García-García, J. L., & Santos-Moreno, A. (2008). Diversidad de cuatro agregaciones de murciélagos en San Miguel Chimalapa, Oaxaca, México. In C. Lorenzo, E. Espinoza & J. Ortega (Eds.), Avances en el estudio de los mamíferos de México (pp. 411-326), Publicaciones especiales, Vol. II. México: Asociación Mexicana de Mastozoología, A. C. [ Links ]
García-García, J. L., Santos-Moreno, A., & Rodríguez-Alamilla, A. (2010). Population dynamics of the bat Dermanura tolteca (Chiroptera: Phyllostomidae) in a tropical forest in Mexico. Revista de Biología Tropical, 58(4), 1323-1334. [ Links ]
Garcia, Q. S., Rezende, J. L., & Aguiar, L. (2000). Seed dispersal by bats in a disturbed area of Southeastern Brazil. Revista de Biologia Tropical, 48(1), 125-128. [ Links ]
Giannini, N. P., & Kalko, E. K. V. (2004). Trophic structure in a large assemblage of phyllostomid bats in Panama. Oikos, 105(2), 209-220. [ Links ]
Gorresen, P. M., & Willig, M. R. (2004). Landscape Responses of Bats To Habitat Fragmentation in Atlantic Forest of Paraguay. Journal of Mammalogy, 85(4), 688-697. [ Links ]
Gorresen, P. M. & Willig, M. R. (2005). Multivariate analysis of scale-dependent associations between bats and landscape structure. Ecological Applications, 15(6), 2126-2136. [ Links ]
Gotelli, N. J., & Colwell, R. K. (2001). Quantifying biodiversity: procedures and pitfalls in the measurement and comparison of species richness. Ecology Letters, 4(4), 379-391. [ Links ]
Haila, Y. (2002). A conceptual genealogy of fragmentation research: from island biogeography to landscape ecology. Ecological applications, 12(2), 321-334. [ Links ]
Hair, J. F., Black, W. C., Babin, J. B., & Anderson, R. E. (2009). Multivariate Data Analysis, 7 edition. EEUU: Prentice Hall. [ Links ]
Handley, C., Wilson, D. E., & Gardner, A. L. (1991). Demography and natural history of the common fruit bat, Artibeus jamaicensis, on Barro Colorado Island, Panama. Washington, D.C. EEUU: Smithsonian Institution Press. [ Links ]
Harper, K. A., MacDonald, S. E., Burton, P. E., Chen, J., Brosofske, K. D., Saunders, S. C., Euskirchen, E. S., Roberts, D., Maiteh, M. S., & Esseen, P. A. (2005). Edge influence on forest structure & composition in fragmented landscapes. Conservation Biology, 19(3), 768-782. [ Links ]
Hecker, K. R., & Brigham, R. M. (1999). Does moonlight change vertical stratification of activity by forest-dwelling insectivorous bats? Journal of Mammalogy, 80(4), 1196-1201. [ Links ]
Heikkinen, R. K., Luoto, M., Virkkala, R., & Rainio, K. (2004). Effects of habitat cover, landscape structure and spatial variables on the abundance of birds in an agricultural-forest mosaic. Journal of Applied Ecology, 41(5), 824-835. [ Links ]
Hill, M. O. (1973). Diversity and evenness: a unifying notation and its consequences. Ecology, 54(2), 427-432. [ Links ]
IMTA (Instituto Mexicano de Tecnología del Agua). (1999). Extractor Rápido de Información Climática ERIC II, programa ejecutable. Morelos, México. [ Links ]
João, M., Pereira, R., Marques, J. T., & Palmeirim, J. M. (2010). Vertical stratification of bat assemblages in flooded and unflooded Amazonian forests. Current Zoology, 56(4), 469-478. [ Links ]
Jost, L. (2006). Entropy and diversity. Oikos, 113(2), 363-375. [ Links ]
Kalka, M., & Kalko, E. K. V. (2005). Gleaning bats as underestimated predators of herbivorous insects: diet of Micronycteris microtis (Phyllostomidae) in Panama. Journal of Tropical Ecology, 22(1), 1-10
Links ]">
Kalko, E. K. V. (1998). Organisation and diversity of tropical bat communities through space and time. Zoology, 101(4), 281-297. [ Links ]
Kalko, E. K. V., & Handley, C. O. (2001). Neotropical bats in the canopy: diversity, community structure, and implications for conservation. Plant Ecology, 153(1), 319-333. [ Links ]
Keel, S., Gentry, A. H., & Spinzi, L. (1993). Using vegetation analysis to facilitate the selection of conservation sites in eastern Paraguay. Conservation Biology, 7(1), 66-75. [ Links ]
Kindt, R., & Coe, R. (2005). Tree diversity analysis: A manual and software for common statistical methods for ecological and biodiversity studies. Nairobi, Kenya: World Agroforestry Centre (ICRAF). [ Links ]
Klingbeil, B. T., & Willig, M. R. (2009). Guild-specific responses of bats to landscape composition and configuration in fragmented Amazonian rainforest. Journal of Applied Ecology, 46(1), 203-213. [ Links ]
Laurance, W. F. (1991). Ecological Correlates of Extinction Proneness in Australian Tropical Rain Forest Mammals. Conservation Biology, 5(1), 79-89. [ Links ]
Leitão, A. B., J. Miller, J., Ahern, J., & McGarigal, K. (2006). Measuring Landscapes: A Planners Hand-book. Washington, D.C., EEUU: Island Press. [ Links ]
Lemke, T. O. (1984). Foraging Ecology of the Long-Nosed Bat, Glossophaga Soricina, With Respect to Resource Availability. Ecology, 65(2), 538–548. [ Links ]
Loayza, A. P., & Loiselle, B. A. (2008). Preliminary Information on the Home Range and Movement Patterns of Sturnira lilium (Phyllostomidae) in a Naturally Fragmented Landscape in Bolivia. Biotropica, 40(5), 630-635. [ Links ]
López, T., Medellín, R., & Yanes, G. 1998. Vampyrum spectrum en Chiapas, México. Revista Mexicana de Mastozoología, 3, 135-136. [ Links ]
Lyons, S., & Willig, M. R. (2002). Species richness, latitude, and scale-sensitivity. Ecology, 83(1), 47-58. [ Links ]
Mac Nally, R. (2000). Regression and model-building in conservation biology, biogeography and ecology: the distinction between–and reconciliation of– ’predictive’and “explanatory”models. Biodiversity & Conservation, 9(5), 655-671. [ Links ]
Mac Nally, R. (2002). Multiple regression and inference in ecology and conservation biology: further comments on identifying important predictor variables. Biodiversity & Conservation, 11(8), 1397-1401. [ Links ]
Mac Nally, R., & Walsh, C. J. (2004). Hierarchical Par-titioning Public-domain Software. Biodiversity & Conservation, 13(3), 659-660. [ Links ]
Marinho-Filho, J. S. (1991). The Coexistence of Two Frugivorous Bat Species and the Phenology of Their Food Plants in Brazil The coexistence of two frugivorous bat species and the phenology of their food plants in Brazil. Journal of Tropical Ecology, 7(1), 59-67. [ Links ]
McCune, B., & Grace, J. B. (2002). Analysis of ecological communities. Oregon, EEUU: MJM Software Design, Gleneden Beach. [ Links ]
McGarigal, K., & McComb, W. C. (1995). Relationships between landscape structure and breeding birds in the Oregon Coast Range. Ecological Monographs, 65(3), 235-260. [ Links ]
McGarigal, K., & Cushman, S. (2002). Comparative evaluation of experimental approaches to the study of habitat fragmentation effects. Ecological Applications, 12(2), 335-345. [ Links ]
McGarigal, K., Cushman, S. A., Neel, M. C., & Ene, E. (2002). FRAGSTATS: Spatial Pattern Analysis Program for Categorical Maps. Analysis. University of Massachusetts. Disponible en: http://www.umass.edu/landeco/research/fragstats/fragstats.html
Links ]">
Medellín, R. A., Arita, H. T., & Sánchez, O. (1997). Identificación de los murciélagos de México, clave de campo. México: Asociación Mexicana de Mastozoología, A. C. [ Links ]
Medellín, R. A., Equihua, M., & Amin, M. A. (2000). Bat Diversity and Abundance as Indicators Disturbance in Neotropical Rainforests. Conservation Biology, 14(6), 1666-1675. [ Links ]
Medina, A., Harvey, C., & Merlo, D. (2007). Bat Diversity and Movement in an Agricultural Landscape in Matiguas, Nicaragua. Biotropica, 39(1), 120-128. [ Links ]
Meyer, C. F. J., & Kalko, E. K. V. (2008). Assemblage-level responses of phyllostomid bats to tropical forest fragmentation: land-bridge islands as a model system. Journal of Biogeography, 35(9), 1711–1726. [ Links ]
Meyer, C. F. J., Fründ, J., Lizano, W. P., & Kalko, E. K. V. (2007). Ecological correlates of vulnerability to fragmentation in Neotropical bats. Journal of Applied Ecology, 45(1), 381-391. [ Links ]
Montiel, S., Estrada, A., & León, P. (2006). Bat assemblages in a naturally fragmented ecosystem in the Yucatan Peninsula, Mexico: species richness, diversity and spatio-temporal dynamics. Journal of Tropical Ecology, 22(3), 267-276. [ Links ]
Moreno, C. E., & Halffter, G. (2000). Assessing the completeness of bat biodiversity inventories using species accumulation curves. Journal of Applied Ecology, 37(1), 149-158. [ Links ]
Morrison, D. (1980). Foraging and day-roosting dynamics of canopy fruit bats in Panama. Journal of Mammalogy, 61(1), 20-29. [ Links ]
Mueller-Dombois, D., & Ellenberg, H. (1974). Aims and methods in vegetation ecology. Nueva York, EEUU: John Wiley & Sons. [ Links ]
Nogueira, M., & Peracchi, A. (2003). Figseed predation by 2 species of Chiroderma: discovery of a new feeding strategy in bats. Journal of Mammalogy, 84(1), 225-233. [ Links ]
Olea, P. P., Mateo-Tomás, P., & de Frutos, A. (2010). Estimating and modelling bias of the hierarchical partitioning public-domain software: implications in environmental management and conservation. PLoS One, 5(7), e11698. [ Links ]
Patterson, B. D., Willig, M. R., & Stevens, R. D. (2003). Trophic strategies, niche partitioning, and patterns of ecological organization. In T. H. Kunz & M. B. Fenton (Eds.), Bat Ecology (pp. 536-579). EEUU: University of Chicago Press. [ Links ]
Pettorelli, N., Vik, J. O., Mysterud, A., Gaillard, J. -M., Tucker, C. J., & Stenseth, N. C. (2005). Using the satellite-derived NDVI to assess ecological responses to environmental change. Trends in ecology & evolution, 20(9), 503-510. [ Links ]
Presley, S., Willig, M. R., & Castro-Arellano, I. (2009). Effects of habitat conversion on temporal activity patterns of phyllostomid bats in lowland Amazonian rain forest. Journal of Mammalogy, 90(1), 210-221. [ Links ]
R Development Core Team. (2009). R: A Language and Environment for Statistical Computing. Vienna, Aus-tria: R Foundation for Statistical Computing. [ Links ]
Racey, P. A., & Entwistle, A. C. (2003). Conservation ecology of bats. In T. H. Kunz & M. B. Fenton (Eds.), Bat ecology (pp. 680-743). EEUU: University of Chicago Press. [ Links ]
Rex, K., Michener, R., Kunz, T. H., & Voigt, C.C. (2011). Vertical stratification of Neotropical leaf-nosed bats (Chiroptera: Phyllostomidae) revealed by stable car-bon isotopes. Journal of Tropical Ecology, 27(3), 211-222. [ Links ]
Salas-Morales, S. H., Schibli, L., & Torres-Bahena, E. (2001). La importancia Ecológica y Biológica. In World Wildlife Found Program & Secretaria de Medio Ambiente, Recurso Naturales y Pesca (Eds.), Chimalapas: la última oportunidad (pp. 27-47). Oaxaca, México: Taller el Castor. [ Links ]
Sellers, P. J., Berry, J. A., Collatz, G. J., Field, C. B., & Hall, F. G. (1992). Canopy reflectance, photosynthesis, and transpiration. III. A reanalysis using improved leaf models and a new canopy integration scheme. Remote sensing of environment, 42(3), 187-216. [ Links ]
Simmons, N. B. (2005). Order Chiroptera. In D. E. Wilson & D. M. Reeder (Eds.), Mammal species of the world: A taxonomic and geographic reference, 2nd edition (pp. 312-529). Baltimore. EEUU: Johns Hopkins University. [ Links ]
Smith, A. C., Koper, N., Francis, C. M. & Fahrig, L. (2009). Confronting collinearity: comparing methods for disentangling the effects of habitat loss and fragmentation. Landscape Ecology, 24(10), 1271-1285. [ Links ]
Steffan-Dewenter, I., Münzenberg, U., Bürger, C., Thies, C., & Tscharntke, T. (2002). Scale-Dependent Effects of Landscape Context on Three Pollinator Guilds. Ecology, 83(5), 1421-1432. [ Links ]
Straube, F. C., & Bianconi, G. V. (2002). Sobre a grandeza e a unidade utilizada para estimar esforço de captura com utilização de redes-de-neblina. Chiroptera Neo-tropical, 8(1-2), 150-152. [ Links ]
ter Braak, C. J. F. (1987). Ordination. In R. H. G. Jongman, C. J. E ter Braak & O. E. R. van Tongeren (Eds.), Data analysis in communityy and landscape ecology (pp. 91-173). Cambridge, Reino Unido: Cambridge University Press. [ Links ]
ter Braak, C. J. F., & Verdonschot, P. F. M. (1995). Canonical correspondence analysis and related multivariate methods in aquatic ecology. Aquatic Sciences, 57(3), 255-289. [ Links ]
Thies, W., & Kalko, E. K. V. (2004). Phenology of neotro-pical pepper plants (Piperaceae) and their association with their main dispersers two short-tailed fruit bats, Carollia perspicillata and C. castanea (Phyllostomidae). Oikos, 104(2), 362-376. [ Links ]
Timm, R. M. (1994). The mammal fauna. In L. A. McDade, K. S. Bawa, H. A. Hespenheide & G. S. Hartshorn (Eds.), La Selva: ecology and natural history of a neotropical rainforest (pp. 229-237). Chicago, Illinois, EEUU: University of Chicago Press. [ Links ]
Trzcinski, M. K., Fahrig, L., & Merriam, G. (1999). Independent effects of forest cover and fragmentation on the distribution of forest breeding birds. Ecological Applications, 9(2), 586-593. [ Links ]
Tschapka, M. (2004). Energy density patterns of nectar resources permit coexistence within a guild of Neo-tropical flower-visiting bats. Journal of Zoology, 263(1), 7-21. [ Links ]
Vargas-Contreras, J. A., Medellín, R. A., Escalona-Segura, G., & Interián-Sosa, L. (2009). Vegetation complexity and bat-plant dispersal in Calakmul, Mexico. Journal of Natural History, 43(3-4), 219-243. [ Links ]
Vehrencamp, S. L., Stiles, F. G., & Bradbury, J. W. (1977). Observations on the foraging behavior and avian prey of the neotropical carnivorous bat, Vampyrum spectrum. Journal of Mammalogy, 58(4), 469-478. [ Links ]
Villard, M., Trzcinski, M. K., & Merriam, G. (1999). Frag-mentation Effects on Forest Birds: Relative Influence of Woodland Cover and Configuration on Landscape Occupancy. Conservation Biology, 13(4), 774-783. [ Links ]
Vitousek, P., Mooney, H., Lubchenco, J., & Melillo, J. (1997). Human domination of Earth’s ecosystems. Science, 277(5325), 494-499. [ Links ]
Walther, B. A., & Moore, J. L. (2005). The concepts of bias, precision and accuracy, and their use in testing the performance of species richness estimators, with a literature review of estimator performance. Ecography, 28(6), 815-829. [ Links ]
Watson, M. D., & Peterson, T. A. (1999). Determinants of diversity in a naturally fragmented landscape: humid montane forest avifaunas of Mesoamerica. Ecography, 22(5), 582-589. [ Links ]
Wetterer, A. L., Rockman, M. V., & Simmons, N. B. (2000). Phylogeny of Phyllostomid Bats (Mammalia: Chiroptera): Data From Diverse Morphological Systems, Sex Chromosomes, and Restriction Sites. Bulletin of the American Museum of Natural History, 248(1), 1-200. [ Links ]
Wiens, J. (1989). Spatial scaling in ecology. Functional ecology, 3(4), 385-397. [ Links ]
Wilcove, D. S., McLellan, C. H., & Dobson, A. P. (1986). Habitat fragmentation in the temperate zone. In M. E. Soulé (Ed.), Conservation biology: the science of scarcity and diversity (pp. 237-256). Massachusetts, EEUU: Sinauer Associates, Inc., Sutherland. [ Links ]
Willig, M. R., Presley, S., Bloch, C., & Hice, C. (2007). Phyllostomid bats of lowland Amazonia: effects of habitat alteration on abundance. Biotropica, 39(6), 737-746. [ Links ]
Álvarez, T., Álvarez-Castañeda, S. T., & López-Vidal, J. C. (1994). Claves para murciélagos mexicanos. México: Instituto Politécnico Nacional. [ Links ]
Anaya, A. L. & Álvarez, M. (1994). Plan de Desarrollo y Conservación de una Reserva Campesina en los Chimalapas. México: Secretaría de Desarrollo Social. Instituto Nacional de Ecología. Programa de Acción Forestal Tropical. [ Links ]
Andrén, H. (1994). Effects of habitat fragmentation on birds and mammals in landscapes with different proportions of suitable habitat: a review. Oikos, 71(3), 55-366. [ Links ]
Antongiovanni, M., & Metzger, J. (2005). Influence of matrix habitats on the occurrence of insectivorous bird species in Amazonian forest fragments. Biological Conservation, 122(3), 441-451. [ Links ]
Arita, H. T., & Santos-del-Prado, K. (1999). Conservation biology of nectar-feeding bats in Mexico. Journal of Mammalogy, 80(1), 31-41. [ Links ]
Asbjornsen, H., & Gallardo-Hernández, C. (2004). Effects of the 1998 fires on tropical montane cloud forests; Chimalapas, Oaxaca. In L. Villers-Ruíz & G. Lopez-Blanco (Eds.), Forest Fires in Mexico: Methods and Evaluation (pp. 125-146). México: Universidad Nacional Autónoma de México. [ Links ]
Asbjornsen, H., Velázquez-Rosas, N., García-Soriano, R., & Gallardo-Hernández, C. (2005). Deep ground fires cause massive above- and below-ground biomass losses in tropical montane cloud forests in Oaxaca, Mexico. Journal of Tropical Ecology, 21(4), 427-434. [ Links ]
Avila-Cabadilla, L. D., Stoner, K. E., Henry, M., & Añorve, M. Y. A. (2009). Composition, structure and diversity of phyllostomid bat assemblages in different successional stages of a tropical dry forest. Forest Ecology and Management, 258(6), 986-996. [ Links ]
Bélisle, M. (2005). Measuring landscape connectivity: the challenge of behavioral landscape ecology. Ecology, 86(8), 1988-1995. [ Links ]
Bernard, E. (2001). Vertical stratification of bat communities in primary forests of Central Amazon, Brazil. Journal of Tropical Ecology, 17(1), 115-126. [ Links ]
Bernard, E., & Fenton, M. B. (2002). Species diversity of bats (Mammalia: Chiroptera) in forest fragments, primary forests, and savannas in central Amazonia, Brazil. Canadian Journal of Zoology, 80(6), 1124-1140. [ Links ]
Brose, U., & Martínez, N. D. (2004). Estimating the rich-ness of species with variable mobility. Oikos, 105(2): 292-300. [ Links ]
Castro-Luna, A. A., Sosa, V. J., & Castillo-Campos, G. (2007). Bat diversity and abundance associated with the degree of secondary succession in a tropical forest mosaic in south-eastern Mexico. Animal Conservation, 10(2), 219-228. [ Links ]
Ceballos, G., Rodríguez, P., & Medellín, R. (1998). Assessing conservation priorities in megadiverse Mexico: mammalian diversity, endemicity, and endangerment. Ecological Applications, 8(1), 8-17. [ Links ]
Charles-Dominique, P. (1986). Inter-relations between frugivorous vertebrates and pioneer plants: Cecropia, birds and bats in French Guyana. In A. Estrada & T. H. Fleming (Eds.), Frugivores and seed dispersal (pp. 119-135). Holanda: Dr. W. Junk Publishers, Dordrecht. [ Links ]
Chaverri, G., Quirós, O. E., & Kunz, T. H. (2007). Ecological correlates of range size in the tent-making bat Artibeus watsoni. Journal of mammalogy, 88(2), 477-486. [ Links ]
Chevan, A., & Sutherland, M. (1991). Hierarchical Partitioning. American Statistician, 45(2), 90-96. [ Links ]
Clarke, F. M., Pio, D. V., & Racey, P. A. (2005). A Comparison of Logging Systems and Bat Diversity in the Neotropics. Conservation Biology, 19(4), 1194-1204. [ Links ]
Clarke, F. M., Rostant, L. V., & Racey, P. A. (2005). Life after logging: post-logging recovery of a neotropical bat community. Journal of Applied Ecology, 42(2), 409-420. [ Links ]
Colwell, R. K. (2011). EstimateS, Version 8.2: Statistical Estimation of Species Richness and Shared Species from Samples (Software and User’s Guide). [ Links ]
Cosson, J. -F., Pons, J. -M., & Masson, D. (1999). Effects of forest fragmentation on frugivorous and nectarivorous bats in French Guiana. Journal of Tropical Ecology, 15(4), 515-534. [ Links ]
Da Silva, A. G., Gaona, O., & Medellín, R. (2008). Diet and trophic structure in a community of fruit-eating bats in Lacandon Forest, Mexico. Journal of Mamma-logy, 89(1), 43-49. [ Links ]
Dinerstein, E. (1986). Reproductive ecology of fruit bats and the seasonality of fruit production in a Costa Rican cloud forest. Biotropica, 18(4), 307-318. [ Links ]
Erlich, P. R., & Wilson, E. O. (1991). Biodiversity studies: science and policy. Science, 253(5021), 758-72. [ Links ]
Estrada, A., & Coates-Estrada, R. (2002). Bats in continuous forest, forest fragments and in an agricultural mosaic habitat-island at Los Tuxtlas, Mexico. Biolo-gical Conservation, 103(2), 237-245. [ Links ]
Estrada, A., Coates-Estrada, R., & Meritt, D. (1993). Bat species richness and abundance in tropical rain forest fragments and in agricultural habitats at Los Tuxtlas, Mexico. Ecography, 16(4), 309-318. [ Links ]
Evelyn, M. J., & Stiles, D. A. (2003). Roosting Require-ments of Two Frugivorous Bats (Sturnira lilium and Arbiteus intermedius) in Fragmented Neotropical Forest. Biotropica, 35(3), 405-418. [ Links ]
Ewers, R. M., & Didham, R. K. (2006). Confounding factors in the detection of species responses to habitat fragmentation. Biological Reviews, 81(1), 117-142. [ Links ]
Fahrig, L. (2003). Effects of Habitat Fragmentation on Biodiversity. Annual Review of Ecology Evolution and Systematics, 34(1), 487-515. [ Links ]
Faria, D. (2006). Phyllostomid bats of a fragmented lands-cape in the north-eastern Atlantic forest, Brazil. Journal of Tropical Ecology, 22(5), 531-542. [ Links ]
Fenton, M., Bernard, E., & Bouchard, S. (2001). The bat fauna of Lamanai, Belize: roosts and trophic roles. Journal of Tropical, 17(4), 511-524. [ Links ]
Fenton, M. B., Acharya, L., Audet, D., Hickey, M. B. C., Merriman, C., Obrist, M. K., Syme, D.M., & Adkins, B. (1992). Phyllostomid bats (Chiroptera: Phyllostomidae) as indicators of habitat disruption in the Neotropics. Biotropica, 24(3), 440-446. [ Links ]
Fleming, T.H. (1988). The Short-tailed Fruit Bat: a Study in Plant-Animal Interactions. Chicago, Illinois, EEUU: University of Chicago Press. [ Links ]
Fleming, T., & Heithaus, E. (1986). Seasonal foraging behavior of the frugivorous bat Carollia perspicillata. Journal of Mammalogy, 67(4), 660-671. [ Links ]
Ford, W., Menzel, M., Rodrigue, J., Menzel, J., & Johnson, J. (2005). Relating bat species presence to simple habitat measures in a central Appalachian forest. Biological Conservation, 126(4), 528-539. [ Links ]
Freeman, P. (1988). Frugivorous and animalivorous bats (Microchiroptera): dental and cranial adaptations. Biological Journal of the Linnean Society, 33(3), 249-272. [ Links ]
Gannon, M. R. (1993). A new technique for marking bats. Bat Research News. 34(4), 88-89. [ Links ]
García-García, J. L., & Santos-Moreno, A. (2008). Diversidad de cuatro agregaciones de murciélagos en San Miguel Chimalapa, Oaxaca, México. In C. Lorenzo, E. Espinoza & J. Ortega (Eds.), Avances en el estudio de los mamíferos de México (pp. 411-326), Publicaciones especiales, Vol. II. México: Asociación Mexicana de Mastozoología, A. C. [ Links ]
García-García, J. L., Santos-Moreno, A., & Rodríguez-Alamilla, A. (2010). Population dynamics of the bat Dermanura tolteca (Chiroptera: Phyllostomidae) in a tropical forest in Mexico. Revista de Biología Tropical, 58(4), 1323-1334. [ Links ]
Garcia, Q. S., Rezende, J. L., & Aguiar, L. (2000). Seed dispersal by bats in a disturbed area of Southeastern Brazil. Revista de Biologia Tropical, 48(1), 125-128. [ Links ]
Giannini, N. P., & Kalko, E. K. V. (2004). Trophic structure in a large assemblage of phyllostomid bats in Panama. Oikos, 105(2), 209-220. [ Links ]
Gorresen, P. M., & Willig, M. R. (2004). Landscape Responses of Bats To Habitat Fragmentation in Atlantic Forest of Paraguay. Journal of Mammalogy, 85(4), 688-697. [ Links ]
Gorresen, P. M. & Willig, M. R. (2005). Multivariate analysis of scale-dependent associations between bats and landscape structure. Ecological Applications, 15(6), 2126-2136. [ Links ]
Gotelli, N. J., & Colwell, R. K. (2001). Quantifying biodiversity: procedures and pitfalls in the measurement and comparison of species richness. Ecology Letters, 4(4), 379-391. [ Links ]
Haila, Y. (2002). A conceptual genealogy of fragmentation research: from island biogeography to landscape ecology. Ecological applications, 12(2), 321-334. [ Links ]
Hair, J. F., Black, W. C., Babin, J. B., & Anderson, R. E. (2009). Multivariate Data Analysis, 7 edition. EEUU: Prentice Hall. [ Links ]
Handley, C., Wilson, D. E., & Gardner, A. L. (1991). Demography and natural history of the common fruit bat, Artibeus jamaicensis, on Barro Colorado Island, Panama. Washington, D.C. EEUU: Smithsonian Institution Press. [ Links ]
Harper, K. A., MacDonald, S. E., Burton, P. E., Chen, J., Brosofske, K. D., Saunders, S. C., Euskirchen, E. S., Roberts, D., Maiteh, M. S., & Esseen, P. A. (2005). Edge influence on forest structure & composition in fragmented landscapes. Conservation Biology, 19(3), 768-782. [ Links ]
Hecker, K. R., & Brigham, R. M. (1999). Does moonlight change vertical stratification of activity by forest-dwelling insectivorous bats? Journal of Mammalogy, 80(4), 1196-1201. [ Links ]
Heikkinen, R. K., Luoto, M., Virkkala, R., & Rainio, K. (2004). Effects of habitat cover, landscape structure and spatial variables on the abundance of birds in an agricultural-forest mosaic. Journal of Applied Ecology, 41(5), 824-835. [ Links ]
Hill, M. O. (1973). Diversity and evenness: a unifying notation and its consequences. Ecology, 54(2), 427-432. [ Links ]
IMTA (Instituto Mexicano de Tecnología del Agua). (1999). Extractor Rápido de Información Climática ERIC II, programa ejecutable. Morelos, México. [ Links ]
João, M., Pereira, R., Marques, J. T., & Palmeirim, J. M. (2010). Vertical stratification of bat assemblages in flooded and unflooded Amazonian forests. Current Zoology, 56(4), 469-478. [ Links ]
Jost, L. (2006). Entropy and diversity. Oikos, 113(2), 363-375. [ Links ]
Kalka, M., & Kalko, E. K. V. (2005). Gleaning bats as underestimated predators of herbivorous insects: diet of Micronycteris microtis (Phyllostomidae) in Panama. Journal of Tropical Ecology, 22(1), 1-10
Links ]">
Kalko, E. K. V. (1998). Organisation and diversity of tropical bat communities through space and time. Zoology, 101(4), 281-297. [ Links ]
Kalko, E. K. V., & Handley, C. O. (2001). Neotropical bats in the canopy: diversity, community structure, and implications for conservation. Plant Ecology, 153(1), 319-333. [ Links ]
Keel, S., Gentry, A. H., & Spinzi, L. (1993). Using vegetation analysis to facilitate the selection of conservation sites in eastern Paraguay. Conservation Biology, 7(1), 66-75. [ Links ]
Kindt, R., & Coe, R. (2005). Tree diversity analysis: A manual and software for common statistical methods for ecological and biodiversity studies. Nairobi, Kenya: World Agroforestry Centre (ICRAF). [ Links ]
Klingbeil, B. T., & Willig, M. R. (2009). Guild-specific responses of bats to landscape composition and configuration in fragmented Amazonian rainforest. Journal of Applied Ecology, 46(1), 203-213. [ Links ]
Laurance, W. F. (1991). Ecological Correlates of Extinction Proneness in Australian Tropical Rain Forest Mammals. Conservation Biology, 5(1), 79-89. [ Links ]
Leitão, A. B., J. Miller, J., Ahern, J., & McGarigal, K. (2006). Measuring Landscapes: A Planners Hand-book. Washington, D.C., EEUU: Island Press. [ Links ]
Lemke, T. O. (1984). Foraging Ecology of the Long-Nosed Bat, Glossophaga Soricina, With Respect to Resource Availability. Ecology, 65(2), 538–548. [ Links ]
Loayza, A. P., & Loiselle, B. A. (2008). Preliminary Information on the Home Range and Movement Patterns of Sturnira lilium (Phyllostomidae) in a Naturally Fragmented Landscape in Bolivia. Biotropica, 40(5), 630-635. [ Links ]
López, T., Medellín, R., & Yanes, G. 1998. Vampyrum spectrum en Chiapas, México. Revista Mexicana de Mastozoología, 3, 135-136. [ Links ]
Lyons, S., & Willig, M. R. (2002). Species richness, latitude, and scale-sensitivity. Ecology, 83(1), 47-58. [ Links ]
Mac Nally, R. (2000). Regression and model-building in conservation biology, biogeography and ecology: the distinction between–and reconciliation of– ’predictive’and “explanatory”models. Biodiversity & Conservation, 9(5), 655-671. [ Links ]
Mac Nally, R. (2002). Multiple regression and inference in ecology and conservation biology: further comments on identifying important predictor variables. Biodiversity & Conservation, 11(8), 1397-1401. [ Links ]
Mac Nally, R., & Walsh, C. J. (2004). Hierarchical Par-titioning Public-domain Software. Biodiversity & Conservation, 13(3), 659-660. [ Links ]
Marinho-Filho, J. S. (1991). The Coexistence of Two Frugivorous Bat Species and the Phenology of Their Food Plants in Brazil The coexistence of two frugivorous bat species and the phenology of their food plants in Brazil. Journal of Tropical Ecology, 7(1), 59-67. [ Links ]
McCune, B., & Grace, J. B. (2002). Analysis of ecological communities. Oregon, EEUU: MJM Software Design, Gleneden Beach. [ Links ]
McGarigal, K., & McComb, W. C. (1995). Relationships between landscape structure and breeding birds in the Oregon Coast Range. Ecological Monographs, 65(3), 235-260. [ Links ]
McGarigal, K., & Cushman, S. (2002). Comparative evaluation of experimental approaches to the study of habitat fragmentation effects. Ecological Applications, 12(2), 335-345. [ Links ]
McGarigal, K., Cushman, S. A., Neel, M. C., & Ene, E. (2002). FRAGSTATS: Spatial Pattern Analysis Program for Categorical Maps. Analysis. University of Massachusetts. Disponible en: http://www.umass.edu/landeco/research/fragstats/fragstats.html
Links ]">
Medellín, R. A., Arita, H. T., & Sánchez, O. (1997). Identificación de los murciélagos de México, clave de campo. México: Asociación Mexicana de Mastozoología, A. C. [ Links ]
Medellín, R. A., Equihua, M., & Amin, M. A. (2000). Bat Diversity and Abundance as Indicators Disturbance in Neotropical Rainforests. Conservation Biology, 14(6), 1666-1675. [ Links ]
Medina, A., Harvey, C., & Merlo, D. (2007). Bat Diversity and Movement in an Agricultural Landscape in Matiguas, Nicaragua. Biotropica, 39(1), 120-128. [ Links ]
Meyer, C. F. J., & Kalko, E. K. V. (2008). Assemblage-level responses of phyllostomid bats to tropical forest fragmentation: land-bridge islands as a model system. Journal of Biogeography, 35(9), 1711–1726. [ Links ]
Meyer, C. F. J., Fründ, J., Lizano, W. P., & Kalko, E. K. V. (2007). Ecological correlates of vulnerability to fragmentation in Neotropical bats. Journal of Applied Ecology, 45(1), 381-391. [ Links ]
Montiel, S., Estrada, A., & León, P. (2006). Bat assemblages in a naturally fragmented ecosystem in the Yucatan Peninsula, Mexico: species richness, diversity and spatio-temporal dynamics. Journal of Tropical Ecology, 22(3), 267-276. [ Links ]
Moreno, C. E., & Halffter, G. (2000). Assessing the completeness of bat biodiversity inventories using species accumulation curves. Journal of Applied Ecology, 37(1), 149-158. [ Links ]
Morrison, D. (1980). Foraging and day-roosting dynamics of canopy fruit bats in Panama. Journal of Mammalogy, 61(1), 20-29. [ Links ]
Mueller-Dombois, D., & Ellenberg, H. (1974). Aims and methods in vegetation ecology. Nueva York, EEUU: John Wiley & Sons. [ Links ]
Nogueira, M., & Peracchi, A. (2003). Figseed predation by 2 species of Chiroderma: discovery of a new feeding strategy in bats. Journal of Mammalogy, 84(1), 225-233. [ Links ]
Olea, P. P., Mateo-Tomás, P., & de Frutos, A. (2010). Estimating and modelling bias of the hierarchical partitioning public-domain software: implications in environmental management and conservation. PLoS One, 5(7), e11698. [ Links ]
Patterson, B. D., Willig, M. R., & Stevens, R. D. (2003). Trophic strategies, niche partitioning, and patterns of ecological organization. In T. H. Kunz & M. B. Fenton (Eds.), Bat Ecology (pp. 536-579). EEUU: University of Chicago Press. [ Links ]
Pettorelli, N., Vik, J. O., Mysterud, A., Gaillard, J. -M., Tucker, C. J., & Stenseth, N. C. (2005). Using the satellite-derived NDVI to assess ecological responses to environmental change. Trends in ecology & evolution, 20(9), 503-510. [ Links ]
Presley, S., Willig, M. R., & Castro-Arellano, I. (2009). Effects of habitat conversion on temporal activity patterns of phyllostomid bats in lowland Amazonian rain forest. Journal of Mammalogy, 90(1), 210-221. [ Links ]
R Development Core Team. (2009). R: A Language and Environment for Statistical Computing. Vienna, Aus-tria: R Foundation for Statistical Computing. [ Links ]
Racey, P. A., & Entwistle, A. C. (2003). Conservation ecology of bats. In T. H. Kunz & M. B. Fenton (Eds.), Bat ecology (pp. 680-743). EEUU: University of Chicago Press. [ Links ]
Rex, K., Michener, R., Kunz, T. H., & Voigt, C.C. (2011). Vertical stratification of Neotropical leaf-nosed bats (Chiroptera: Phyllostomidae) revealed by stable car-bon isotopes. Journal of Tropical Ecology, 27(3), 211-222. [ Links ]
Salas-Morales, S. H., Schibli, L., & Torres-Bahena, E. (2001). La importancia Ecológica y Biológica. In World Wildlife Found Program & Secretaria de Medio Ambiente, Recurso Naturales y Pesca (Eds.), Chimalapas: la última oportunidad (pp. 27-47). Oaxaca, México: Taller el Castor. [ Links ]
Sellers, P. J., Berry, J. A., Collatz, G. J., Field, C. B., & Hall, F. G. (1992). Canopy reflectance, photosynthesis, and transpiration. III. A reanalysis using improved leaf models and a new canopy integration scheme. Remote sensing of environment, 42(3), 187-216. [ Links ]
Simmons, N. B. (2005). Order Chiroptera. In D. E. Wilson & D. M. Reeder (Eds.), Mammal species of the world: A taxonomic and geographic reference, 2nd edition (pp. 312-529). Baltimore. EEUU: Johns Hopkins University. [ Links ]
Smith, A. C., Koper, N., Francis, C. M. & Fahrig, L. (2009). Confronting collinearity: comparing methods for disentangling the effects of habitat loss and fragmentation. Landscape Ecology, 24(10), 1271-1285. [ Links ]
Steffan-Dewenter, I., Münzenberg, U., Bürger, C., Thies, C., & Tscharntke, T. (2002). Scale-Dependent Effects of Landscape Context on Three Pollinator Guilds. Ecology, 83(5), 1421-1432. [ Links ]
Straube, F. C., & Bianconi, G. V. (2002). Sobre a grandeza e a unidade utilizada para estimar esforço de captura com utilização de redes-de-neblina. Chiroptera Neo-tropical, 8(1-2), 150-152. [ Links ]
ter Braak, C. J. F. (1987). Ordination. In R. H. G. Jongman, C. J. E ter Braak & O. E. R. van Tongeren (Eds.), Data analysis in communityy and landscape ecology (pp. 91-173). Cambridge, Reino Unido: Cambridge University Press. [ Links ]
ter Braak, C. J. F., & Verdonschot, P. F. M. (1995). Canonical correspondence analysis and related multivariate methods in aquatic ecology. Aquatic Sciences, 57(3), 255-289. [ Links ]
Thies, W., & Kalko, E. K. V. (2004). Phenology of neotro-pical pepper plants (Piperaceae) and their association with their main dispersers two short-tailed fruit bats, Carollia perspicillata and C. castanea (Phyllostomidae). Oikos, 104(2), 362-376. [ Links ]
Timm, R. M. (1994). The mammal fauna. In L. A. McDade, K. S. Bawa, H. A. Hespenheide & G. S. Hartshorn (Eds.), La Selva: ecology and natural history of a neotropical rainforest (pp. 229-237). Chicago, Illinois, EEUU: University of Chicago Press. [ Links ]
Trzcinski, M. K., Fahrig, L., & Merriam, G. (1999). Independent effects of forest cover and fragmentation on the distribution of forest breeding birds. Ecological Applications, 9(2), 586-593. [ Links ]
Tschapka, M. (2004). Energy density patterns of nectar resources permit coexistence within a guild of Neo-tropical flower-visiting bats. Journal of Zoology, 263(1), 7-21. [ Links ]
Vargas-Contreras, J. A., Medellín, R. A., Escalona-Segura, G., & Interián-Sosa, L. (2009). Vegetation complexity and bat-plant dispersal in Calakmul, Mexico. Journal of Natural History, 43(3-4), 219-243. [ Links ]
Vehrencamp, S. L., Stiles, F. G., & Bradbury, J. W. (1977). Observations on the foraging behavior and avian prey of the neotropical carnivorous bat, Vampyrum spectrum. Journal of Mammalogy, 58(4), 469-478. [ Links ]
Villard, M., Trzcinski, M. K., & Merriam, G. (1999). Frag-mentation Effects on Forest Birds: Relative Influence of Woodland Cover and Configuration on Landscape Occupancy. Conservation Biology, 13(4), 774-783. [ Links ]
Vitousek, P., Mooney, H., Lubchenco, J., & Melillo, J. (1997). Human domination of Earth’s ecosystems. Science, 277(5325), 494-499. [ Links ]
Walther, B. A., & Moore, J. L. (2005). The concepts of bias, precision and accuracy, and their use in testing the performance of species richness estimators, with a literature review of estimator performance. Ecography, 28(6), 815-829. [ Links ]
Watson, M. D., & Peterson, T. A. (1999). Determinants of diversity in a naturally fragmented landscape: humid montane forest avifaunas of Mesoamerica. Ecography, 22(5), 582-589. [ Links ]
Wetterer, A. L., Rockman, M. V., & Simmons, N. B. (2000). Phylogeny of Phyllostomid Bats (Mammalia: Chiroptera): Data From Diverse Morphological Systems, Sex Chromosomes, and Restriction Sites. Bulletin of the American Museum of Natural History, 248(1), 1-200. [ Links ]
Wiens, J. (1989). Spatial scaling in ecology. Functional ecology, 3(4), 385-397. [ Links ]
Wilcove, D. S., McLellan, C. H., & Dobson, A. P. (1986). Habitat fragmentation in the temperate zone. In M. E. Soulé (Ed.), Conservation biology: the science of scarcity and diversity (pp. 237-256). Massachusetts, EEUU: Sinauer Associates, Inc., Sutherland. [ Links ]
Willig, M. R., Presley, S., Bloch, C., & Hice, C. (2007). Phyllostomid bats of lowland Amazonia: effects of habitat alteration on abundance. Biotropica, 39(6), 737-746. [ Links ]
*Correspondencia: José Luis García-García: Laboratorio de Ecología Animal, Centro Interdisciplinario de Investigación para el Desarrollo Integral Regional Unidad Oaxaca, Instituto Politécnico Nacional. Hornos No. 1003, Col. Noche Buena, Santa Cruz Xoxocotlán C.P. 71230. Oaxaca, México; jgarciag0800@ipn.mx
Antonio Santos-Moreno: Laboratorio de Ecología Animal, Centro Interdisciplinario de Investigación para el Desarrollo Integral Regional Unidad Oaxaca, Instituto Politécnico Nacional. Hornos No. 1003, Col. Noche Buena, Santa Cruz Xoxocotlán C.P. 71230. Oaxaca, México; asantom90@hotmail.com
1. Laboratorio de Ecología Animal, Centro Interdisciplinario de Investigación para el Desarrollo Integral Regional Unidad Oaxaca, Instituto Politécnico Nacional. Hornos No. 1003, Col. Noche Buena, Santa Cruz Xoxocotlán C.P. 71230. Oaxaca, México; jgarciag0800@ipn.mx, asantom90@hotmail.com
Recibido 19-II-2013. Corregido 06-IX-2013. Aceptado 10-X-2013.


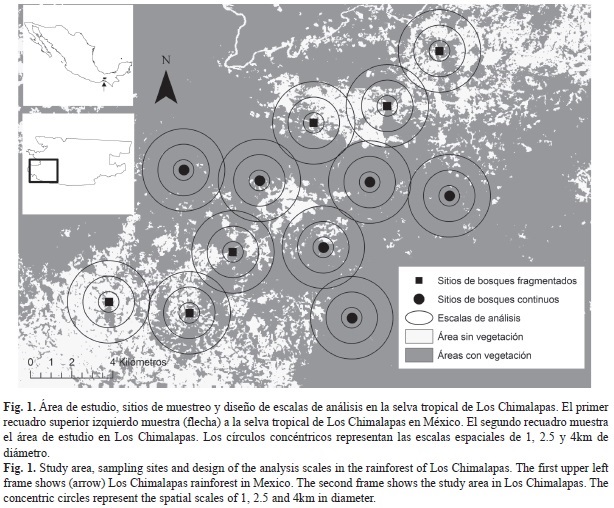{kind=link}
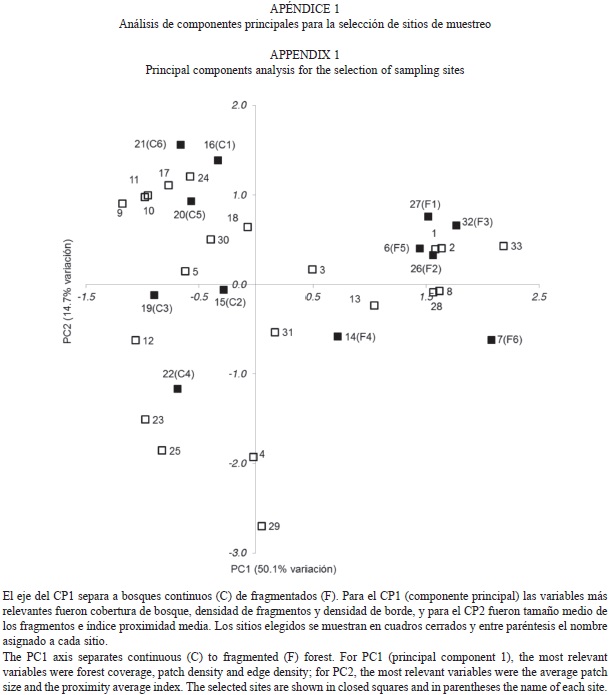{kind=link}
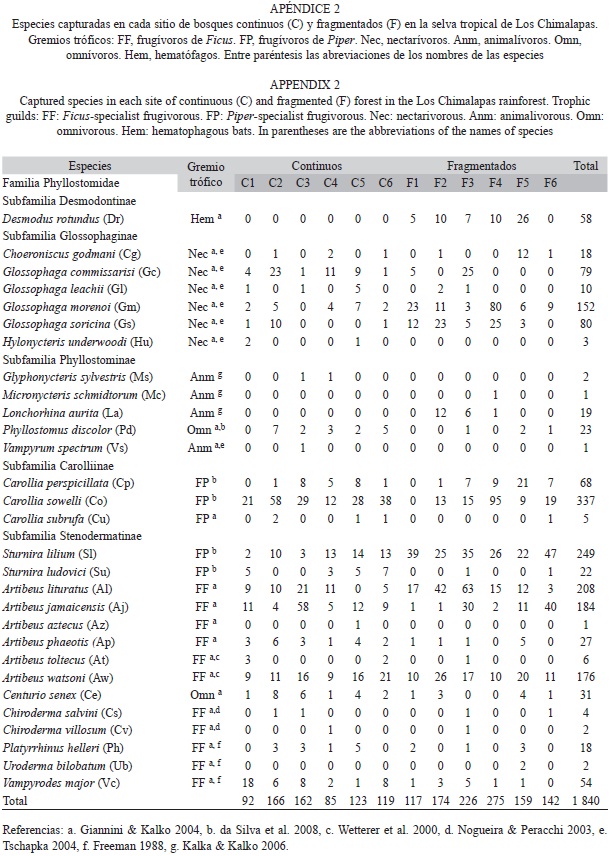{kind=link}
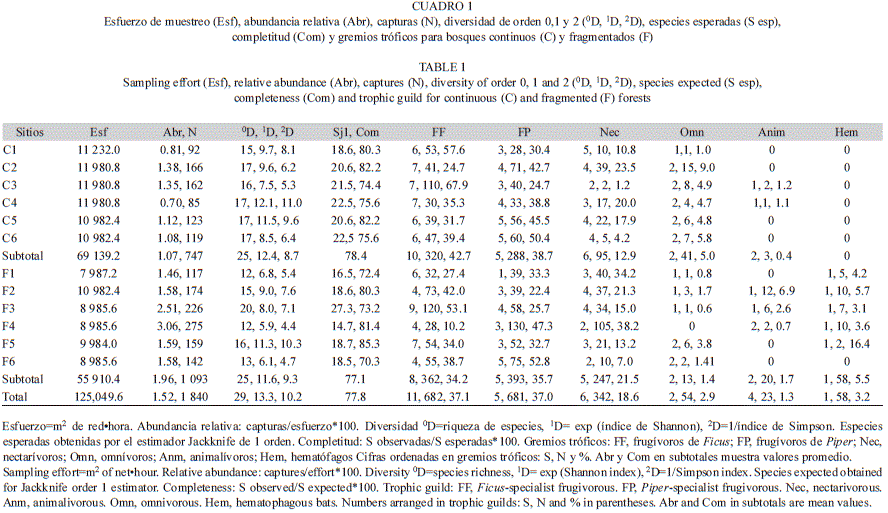{kind=link}
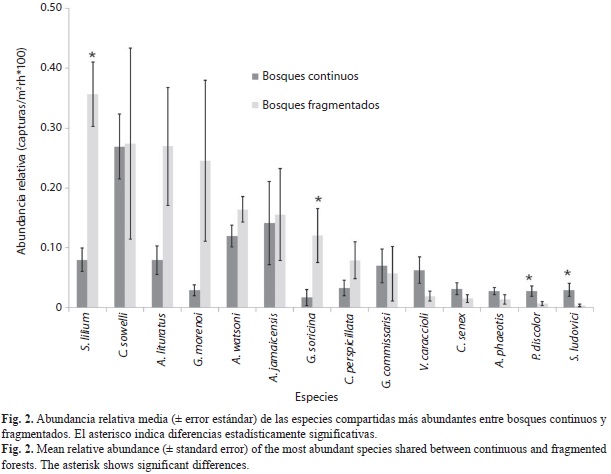{kind=link}
{kind=link}
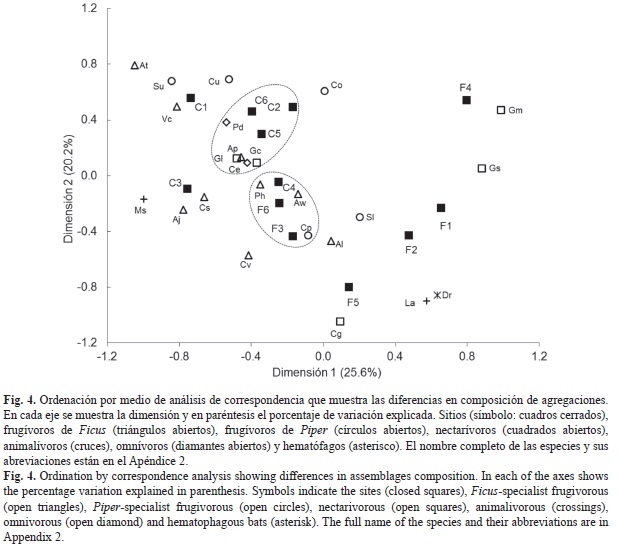{kind=link}
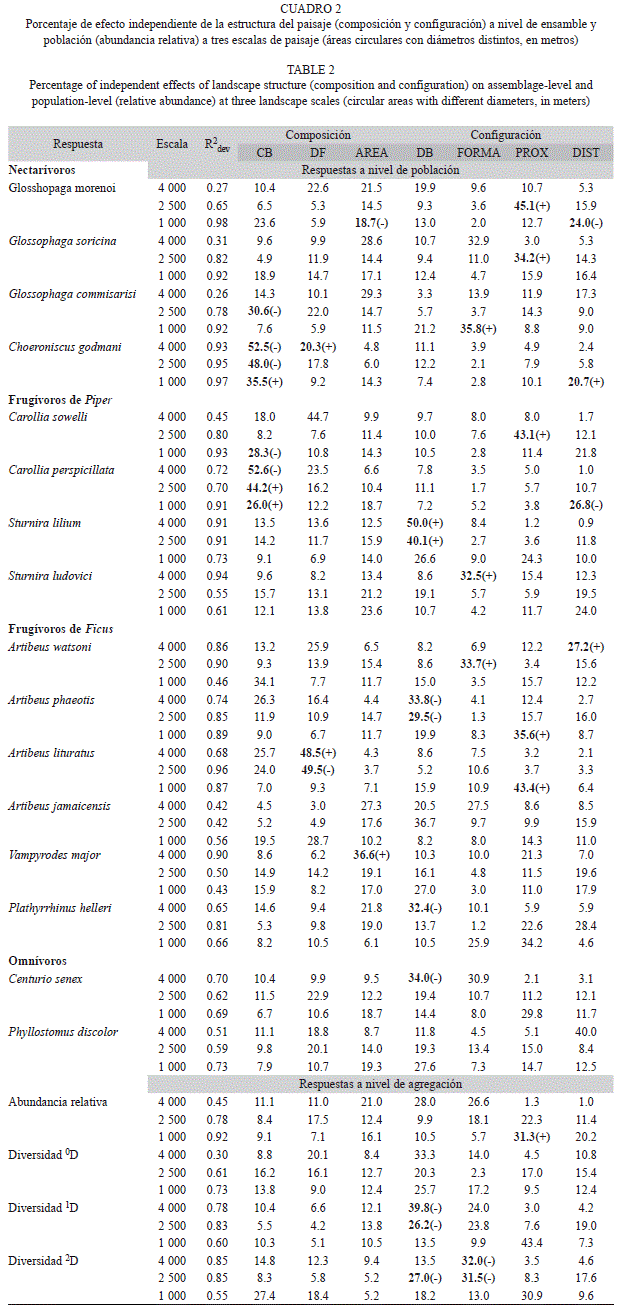{kind=link}
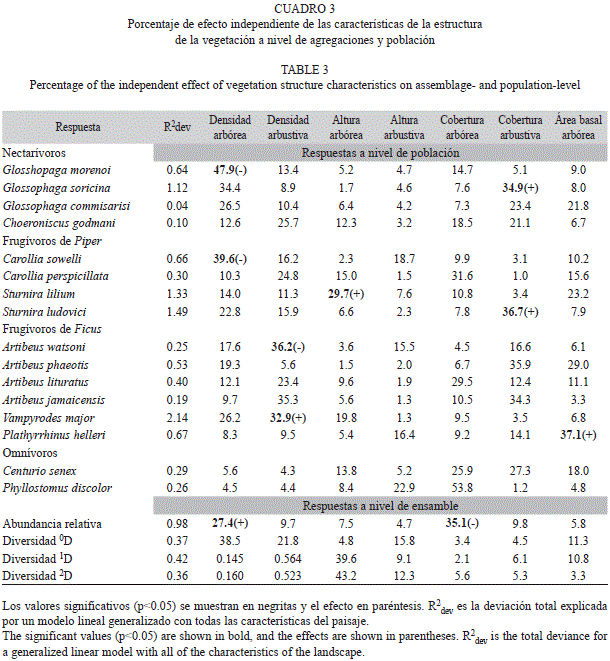{kind=link}