










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.62 n.1 San José Jan./Mar. 2014
Reproductive biology of Pleurodema guayapae (Anura: Leptodactylidae: Leiuperinae)
Biología reproductiva de Pleurodema guayapae (Anura: Leptodactylidae: Leiuperinae)
Biología reproductiva de Pleurodema guayapae (Anura: Leptodactylidae: Leiuperinae)
*Dirección para correspondencia:
Abstract
Pleurodema guayapae is a species that inhabits saline environments and semidesert zones from central Argentina. To date, the knowledge about the reproductive biology of this species is very poor, and our aim is to contribute to its knowledge with the description of some important reproductive aspects. For this, field work was undertaken in an area near to Patquía, La Rioja province. Sampling was undertaken during three summer periods (2006-2007; 2007-2008; 2008-2009) in Chamical-Patquía area, where we could find reproductively active populations. We observed and described breeding sites, type of clutch, process of foam nest construction, clutch and egg number and sizes, and hatching time and stage. Behaviour observations were performed from the time that males began to call until the pairs ended up the foam nests building, and layed the eggs. Additionally, one amplected pair was observed and filmed in the process of foam nest construction, and four amplectant pairs were collected and separatelly placed in plastic containers, for nests observations in the laboratory. Hatching time was based on three different foam nests of known age. We found that P. guayapae populations were acoustically active only after a rainfall. Its breeding sites were represented by ephemeral ponds of fresh water, product of rains. The males emitted their calls inside or outside these ponds. A detailed description of the foam nest construction process by both females and males was made. The clutches were in dome-shaped foam nest type of 6-9cm in diameter and 1-3cm in height, some of which were in communal nests. The nests had an average of 1 137 pigmented eggs. This species showed a short hatching time. Our results allow us to conclude that this species should be considered an extreme explosive breeder. Our results are discussed with others obtained for related species.
Key words: communal nest, ephemeral environment, explosive breeder, foam nest construction.
Resumen
Pleurodema guayapae es una especie que habita en zonas semidesérticas y salinas del área central de Argentina. Los conocimientos actuales sobre la biología reproductiva de esta especie son muy pobres, por lo que nuestro objetivo fue contribuir a su conocimiento con la descripción de algunos aspectos reproductivos importantes. Para ello se realizaron tres campañas de muestreo durante los veranos (2006-2007, 2007-2008 y 2008-2009) en el área de Chamical-Patquía, provincia de La Rioja, donde se encontraron poblaciones reproductivamente activas. Se observaron y describieron los sitios reproductivos, tipo de puesta, proceso de construcción del nido de espuma, tamaño de la puesta y de los huevos, número de huevos por puesta y tiempo y estado de eclosión de la larva. Las observaciones de comportamiento se realizaron desde el momento en que los machos comenzaron a emitir los cantos hasta que la pareja en amplexo terminó de construir el nido de espuma con los huevos en su interior. Además, se observó y filmó una pareja en amplexo, en el proceso de construcción del nido de espuma y se recolectaron cuatro parejas en amplexo y se colocaron separadamente en recipientes plásticos para que sus puestas pudieran ser observadas en el laboratorio. El tiempo de eclosión se determinó en tres nidos de espuma de edad conocida. Se encontró que las poblaciones de P. guayapae solo estuvieron acústicamente activas luego de una lluvia torrencial. Los sitios de reproducción fueron charcas efímeras de agua de lluvia. Los machos emitieron sus cantos de advertencia desde adentro o fuera de esas charcas. Se realizó una descripción detallada del proceso de construcción del nido de espuma por parte de la hembra y el macho de P. guayapae. El tipo de la puesta en esta especie es un nido de espuma flotante con forma de domo de 6 a 9cm de diámetro y de 1 a 3cm de alto, algunos de los cuales estaban unidos entre sí formando puestas comunales. Los nidos de espuma poseen en promedio 1 137 huevos pigmentados. Esta especie mostró un tiempo de eclosión corto. Nuestros resultados nos permiten concluir que P. guayapae debe ser considerada como reproductor explosivo extremo. Nuestros resultados son discutidos con los obtenidos en otras especies afines.
Palabras claves: nidos comunales, ambiente efímero, reproductor explosivo, construcción de nido de espuma.
The reproductive mode in amphibians was defined by Salthe & Duellman (1973) as “a combination of traits that includes oviposition site, ovum and clutch characteristics, rate and duration of development, stage and size of hatchling, and type of parental care, if any”. Thirtynine reproductive modes have been recorded for anurans around the world (Haddad & Prado, 2005). Altig & McDiarmind (2007) provide a generalized framework for diversity of amphibian eggs and ovipositional modes and introduce a standardized terminology for clutch structures.
The genus Pleurodema Tschudi (Leptodactylidae: Leiuperinae) is distributed from Panama throughout South America to Southern Chile and Argentina, and currently is represented by 15 species (Cei, 1980; Valetti, Salas & Martino, 2009; Maciel & Nunez, 2010; Kolenc et al., 2011; Faivovich et al., 2012). This genus has very variable ovipositional modes. The egg-clutch structure in the Southern Chilean populations of P. thaul and P. bufoninum is strings in water (Weigandt, Úbeda & Díaz, 2004). This egg-laying mode is accompanied by inguinal amplexus (Duellman & Veloso, 1977). P. thaul of central and Northern Chile, P. bibroni, P. kriegi and P. cordobae lay eggs in semisub-merged gelatinous eggs-masses (Valetti et al. 2009; Valetti, Otero, Grenat & Martino, 2011; Faivovich et al., 2012). P. tucumanum lays eggs in plate-like floating masses (Martori, Aún & Vignolo, 1994). All other known Pleurodema species lay eggs in aquatic foam nest (Barrio, 1964; Hulse, 1979; Cei, 1980; Peixoto, 1982; Hödl, 1992; Faivovich et al., 2012).
Pleurodema guayapae inhabits saline and dry Chacoan environments from Central Argentina (Barrio, 1964; Cei, 1980; Ferraro & Casagranda, 2009). In addition, De la Riva and Gonzales (1998) reported a population tentatively assigned to P. guayapae from the dry Chacoan areas of Santa Cruz, Bolivia. To date, the knowledge about the reproductive biology of P. guayapae was only briefly described by Barrio (1964). Consequently, the aims of this investigation are: (1) to identify the breeding sites of the species; (2) to describe the process of foam nest construction; (3) to determine clutch and egg sizes, and (4) to identify the time and stage of hatching.
Materials and Methods
Field work was undertaken in an area near to Patquía, La Rioja province, Argentina (23°26’S - 45°04’W) at 434m in elevation. This arid environment was well-described by Barrio (1964). After three summers of sampling (2006-2007; 2007-2008; 2008-2009) in Chamical-Patquía area (La Rioja province), we could find reproductively active populations of Pleurodema guayapae. Field observations of breeding aggregations were made at dawn of 18 January 2009 in two ephemeral ponds of 60m2 and 160m2, respectively. Behaviour observations were performed from the time that male individuals began to emit the advertisement calls until the pairs ended up building the foam nests and lay the eggs. One amplected pair was observed and filmed in the process of the foam nest construction with a Canon PowerShot S5 IS camera (at 30 frames per second), and this behaviour is described following Hödl (1990, 1992).
To determine the number of eggs per foam nest, four amplectant pairs were collected and each pair was put into a plastic container so that nests could be obtained in the laboratory. All eggs of these foam nests were counted. Hatching time was based on three different foam nests of known age. The tadpoles were staged according to Gosner (1960).
Additionally, on the morning of 18 January 2009, data on foam nests were collected in the field to describe the general aspect of the nests and eggs. The number of foam nests in each pond was registered and height and diameter of the nests were measured. One foam nest was placed in 10% formalin solution for morphometric description of its eggs. Egg diameters without the gelatinous capsule were measured using a Zeiss SR stereomicroscope with a micrometer to nearest 0.01mm. Eggs, tadpoles and adult specimens are housed at the herpetological collection of the Department of Natural Sciences, National University of Río Cuarto, Argentina.
Results
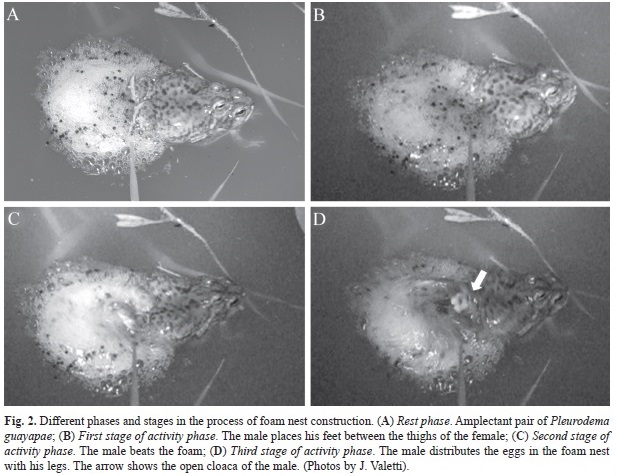
Breeding sites of P. guayapae: Calling and spawning activities were limited only to one night after ca. 30mm of rainfall during a very short time. The calling activity began at 4.00am (immediately after the rain when the ponds were formed) and ended at 7.00am (sunrise). Two ponds were formed in sites where water was stored after the rain. The sizes of the two rain ponds were 15x4 and 40x4 meters (Fig. 1A) and a depth less than 25cm, with approximately 50 males calling in each pond. The males emitted their calls on stones in the pond, inside the pond near the edge or outside the pond (Fig. 1B). The amplexus in P. guayapae is axillary and the pairs were observed out of the water, near the edge of the pond or floating on the surface of the water (Fig. 2A). Once the amplexus is formed and the pair is into the pond, starts the construction of the foam nest.
Process of foam nest construction: The construction of the foam nest has alternating phases of activity and rest (sensu Hödl, 1990). In the rest phase, the pair floats immobile. The male is in axillary amplexus on the female with his feet retracted and the cloaca above that of the female (Fig. 2A). These rest phases are alternated by activity phases that can be described in three stages. The first stage is when the male immerses the feet in the water between the thighs of the female and then the cloacae of both individuals are juxtaposed (Fig. 2B). In this moment, the female spawns and the male lifts the eggs with his tarsal tubercles by retracting the feet above his cloaca returning to the starting position (1 260ms). In the second stage, the male begins a rotational movement of the legs beating the foam. The femur and the tibia rotate perpendicular to the longitudinal axis of the body and the foot is almost parallel to it. After 12 of these cyclical movements (1 500ms, Fig. 2C), the third stage starts and the male opens the legs and his open cloaca can be observed (see Fig. 2D). The sperm release is probably produced here, but this event could not be appreciated clearly. Immediately, the male performs approximately six lateral movements of his extended legs pushing the eggs toward the centre of the foam (430ms, Fig. 2D). Once these movements have been completed, the pair begins another rest phase. This behaviour is repeated until the construction of the foam nest and the simultaneous egg-laying have been completed.
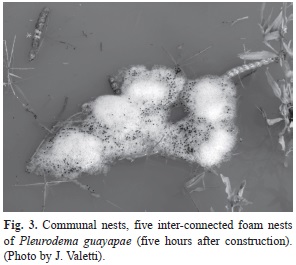
Clutch and egg sizes. Time and stage of hatching: Eleven foam nests were counted in the smaller pond and 25 in the bigger one. The dome-shaped foam nests have diameter and height that varied from 6 to 9cm and 1 to 3cm, respectively. Most of the foam nests were floating alone, although some of them were attached to vegetation. A small number of foam nests were also observed in groups of 2 to 5 interconnected nests (Communal nests; Fig. 3).
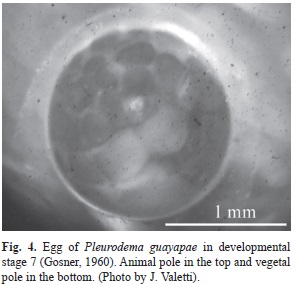
The pigmented eggs (see Fig. 3) have the animal pole darker than the vegetal one (Fig. 4). All eggs are covered by one gelatinous capsule, and the average egg diameter (stage 6-7; Gosner, 1960) without the gelatinous capsule is 1.44mm (SD=0.07, range=1.29-1.61, n=83). The snout-vent length for females ranged from 39.1 to 44.2mm (41.7mm±1.7; n=17) and the average number of eggs per foam nest was 1 137 (SD=275, range=900-1 500, n=4).
The hatching under laboratory conditions of the P. guayapae took between 26 to 30 hours (Stage 19).
Discussion
The reproductive sites where P. guayapae was found the night of January 18 were formed after the rain (ca. 30mm). P. guayapae males have called only at that night and they were not detected in the subsequent nights. The species-specific breeding phenologies are adaptive and often correlate with breeding habitat, seasonal changes in climate, or the presence of potential competitors or predators (Wilbur & Alford, 1985; Duellman & Trueb, 1986). In this sense, Wells (1977), considering the temporal patterns of anuran reproduction, classified species into two broad types: explosive breeders and pro-longed breeders. Most tropical anurans breed in every month of the year and therefore they are classified as prolonged breeders. By contrast, temperate species are more likely to breed explosively and this is common in many species that use seasonally ephemeral habitats such as rain ponds, and they may involve a single annual bout (Heyer, McDiarmid & Weigmann, 1975; Crump, 1983). According to our results, P. guayapae fits the characteristics of the explosive breeder species.
Although individuals of P. guayapae breed in ephemeral ponds, individuals of both sexes were observed outside of permanent and semipermanent ponds, but never inside the ponds. Such situations were detected in many occasions during the field season 2007-08 (January-February, J. Valetti, personal observation). In addition, it was not possible to verify the formation of breeding aggregations in such opportunities. A possible explanation may be the abundance of predators inside of these ponds. These ponds were densely inhabited by Lepidobatrachus llanensis (Anura: Ceratophryidae) individuals (Approximately 1 individual/m2), a natural predator of P. guayapae (Hulse, 1978; J. Valetti, personal observation). At these sites, adults of P. guayapae were distant from the water bodies (between 5 and 50m) and L. llanensis individuals were not observed outside of the ponds. In addition, as Hulse (1978) established, L. llanensis individuals were not observed inside of ephemeral ponds where P. guayapae breeding. Nevertheless, an appro-priate experimental design should be done to evaluate this alleged relationship.
The amplexus observed in P. guayapae is axillary and the eggs are deposited in foam nests floating on water, as the syntopic congeneric species P. nebulosum (Barrio, 1964). The calling sites of P. guayapae males differ from that of P. diplolister. Males of P. diplolister call floating on the surface of puddles (Hödl, 1992) whereas those of P. guayapae always call attached to vegetation into the ponds, on stones or even outside the ponds. Observations in nearby ponds allowed us to determine that the sympatric congeneric species P. tucumanum and P. nebulosum also showed diverse acoustic behaviour. The males of these species emitted the calls floating in ponds or from outside them. In P. guayapae, several amplectant pairs were observed floating on the surface of ponds and others were out of water bodies. Regarding the latter, Hödl (1990) indicated that Physalaemus ephippifer performs the amplexus into the water, but the pair usually leaves the amplexus site and returns to the water several minutes before oviposition. Respect to the breeding site of P. guayapae, there is an important difference with regard to the report by Barrio (1964). This author indicated that the breeding sites are represented by temporary ponds of brackish water, while we find that P. guayapae breed in ephemeral ponds of fresh water, product of a recent rain. In fact, the first foam nests were constructed four hours after the pond formation.
Nine species of Pleurodema lay eggs in aquatic foam nest (Barrio, 1964; Duellman & Veloso, 1977; Hulse, 1979; Cei, 1980; Peixoto, 1982; Hödl, 1992). Halloy & Fiaño (2000) indicate that P. borellii males use their hind legs to whip up a foam nest that prevents the eggs from desiccation. This behaviour is similar to that described for P. diplolister (Hödl, 1992). Our results indicate that P. guayapae lays eggs in aquatic foam nest and males also use their hind legs to construct the nest, as was described for P. borellii and P. diplolister.
According to the terminology used by Hödl (1990, 1992), P. guayapae constructs the foam nest by alternating rest and activity phases. The activity phases composed by three stages or phases were also described for Physalaemus ephippifer (Hödl, 1990) and Pleurodema diplolister (Hödl, 1992). The activity phase of these species is similar to that described here for P. guayapae. Moreover, the second stage of the activity phase of P. guayapae (foam beating phase of Hödl, 1992) is the longest, a feature shared with P. diplolister. Furthermore, both species show series of cyclic movements of the legs at this stage (12 in P. guayapae, and 10 in P. diplolister: Hödl, 1992). However, the foam beating phase of P. diplolister was characterized by Hödl (1992) as rotational movements of the tibia and tarsus perpendicular to the body axis, whereas the movements of legs in P. guayapae differ slightly. In P. guayapae, the femur and the tibia rotate perpendicular to the body axis, while the foot is almost parallel to it. Despite that signs of ejaculation were not observed by Hödl (1992) during the process of foam nest construction, the “female lordosis and male basket formation phase” is suggested for the reason of the juxtaposition of their cloacae. We suggest that in P. guayapae the sperm release probably occurs in the foam beating phase, after the eggs are released by the female. This is based on the observation of the male cloaca that opens at this phase. However, a more detailed analysis over this process well be needed to determine the exact moment of ejaculation.
Pleurodema guayapae lays eggs in foam nests and demonstrated a reproductive strategy adapted to the unpredictable rainfall patterns of the region. Thus, the reproductive mode that corresponds to P. guayapae is 8 (sensu Duellman & Trueb, 1986) or 11 (sensu Haddad & Prado, 2005): “Foam nests floating on the water in ponds; exotrophic tadpoles in ponds”. It has already been demonstrated that species inhabiting environments where water resources are restricted to only short periods during the year, generally have generalized reproductive modes more resistant to the desiccation (Haddad & Prado, 2005; Vieira, Santana & Arzabe, 2009). Several authors suggested that foam nest construction is an adaptation to avoid desiccation of eggs and early larval stages (Dobkin & Gettinger, 1985; Downie, 1988; Cardoso & Arzabe, 1993; Prado, Uetanabaro & Haddad, 2002). Eggs and larvae that develop within foam nests are more protected from predators and against desiccation (Crump, 1974; Haddad, Pombal & Gordo, 1990; Silva, Giaretta & Facure, 2005; Dalgetty & Kennedy, 2010) and they are exposed to adequate supplies of oxygen and to temperatures that favour their development (Petranka, Hopey, Jennings, Baird & Boone, 1994; Haddad & Hödl, 1997). Moreover, Hödl (1992) suggested that the foam nest of Pleurodema diplolister reduced egg predation by conspecific larvae. Also, several inter-connected communal nests of P. guayapae were observed. Zina (2006), in a work about communal nests in Engystomops pustulosus, reported that the foam protects eggs and tadpoles from desiccation for at least four days after oviposition, and nesting communally can extend this protection even further. Therefore, communal nesting in species that deposits eggs in foam may be an important adaptation enhancing survival of these species inhabiting environments of unpredictable rainfall patterns (Downie, 1988; Hödl, 1990; Zina, 2006). In this regard, it has already been confirmed the presence of communal nests in some species of Leiuperinae (Barreto & Andrade, 1995; Tárano, 1998; Giaretta & Menin, 2003; Dalgetty & Kennedy, 2010) and also within the genus Pleurodema in P. diplolister (Höld, 1992; Car-doso & Arzabe, 1993) and P. cinereum (Agos-tini, Cajade & Roesler, 1997).
Faivovich et al. (2012) suggested that eggs laid in a foam nest built by the mating pair during the amplexus is a synapomorphy for the clade composed by Edalorhina, Engystomops, Physalaemus and Pleurodema, but their results also showed that Pleurodema has experienced subsequent evolutive transformations from foam nest clutches to eggs laid in gelatinous structures, being subspherical masses, ovoid and plate-like, or egg strings.
The size and form of P. guayapae foam nests were similar to those declared for P. diplolister (Höld, 1992; Cardoso & Arzabe, 1993). However, the number of eggs per foam nest was greater in P. guayapae (900-1 500 against 528-748 in P. diplolister: Höld 1992). The number of eggs per clutches remains unknown for most species of Pleurodema. In this regard, the foam nests of P. borellii produce, on average, about 1 300 hatchlings (Halloy & Fiaño, 2000) and the semi-submerged gelatinous eggs-masses of P. cordobae (Valetti et al., 2009) have between 74 and 215 eggs (Valetti et al., 2011). The egg diameter of P. guayapae without the gelatinous capsule was somewhat smaller (between 1.29 and 1.61mm) that the reported for P. bufoni-num (1.26mm to 2.94mm), a species that lays eggs in solid and cylindrical strings (Weigandt et al., 2004).
The hatching time of the P. guayapae tadpoles was short (26 to 30 hours at the developmental Stage 19), inferring a rapid larval development. This pattern of development is typical of species that breed explosively in temporary or ephemeral environments, so the larvae metamorphose in a short period, before desiccation of the water bodies. Rapid embryonic and larval development in P. diplolister was interpreted by Peixoto (1982) as an adaptation to highly xeric environments. We corroborated that 48 hours after the start of foam nest construction, the water level of ponds decreased considerably. In this sense, some foam nests stayed out of the water and all eggs and small tadpoles recently hatching were found dead.
In summary, our results indicated that the reproduction period of P. guayapae is associated to the summer rainfalls and ephemeral environments generated after these rains. Moreover, the short time at which the populations of P. guayapae are reproductively active, the type of clutches in foam nest, the presence of communal nests, and the rapid embryonic development, allow us to classify this species as an extreme explosive breeder.
Acknowledgments
The Secretary of Research and Technology of National University of Río Cuarto (SECY-TUNRC) provided funds by Grant PPI 18/ C350. We thank Pablo Brandolin and Miguel Avalos for help in field work. We also acknowledge four anonymous reviews and an editor for their careful critiques, which helped in strengthening the paper and Susan Vilor for her assistance with the English review. JAV, PRG and MB thank CONICET – Argentina (Consejo Nacional de Investigaciones Científicas y Tecnológicas) for fellowship granted.
References
Agostini, M. G., Cajade, R., & Roesler, I. (2007). Commu-nal oviposition. Pleurodema cinereum. Herpetological Review, 38, 441. [ Links ]
Altig, R., & McDiarmid, R. W. (2007). Morphological diversity and evolution of egg and clutch structure in amphibians. Herpetological Monographs, 21, 1-32. [ Links ]
Barreto, L., & Andrade, G. V. (1995). Aspects of the reproductive biology of Physalaemus cuvieri (Anura: Leptodactylidae) in Northeastern Brazil. Amphibia-Reptilia, 16, 67-76. [ Links ]
Barrio, A. (1964). Especies crípticas del género Pleurodema que conviven en una misma área, identificadas por el canto nupcial (Anura, Leptodactylidae). Physis, 24(68), 471-489. [ Links ]
Cardoso, A. J., & Arzabe, C. (1993). Corte e desenvolvimento larvário de Pleurodema diplolistris (Anura: Leptodactylidae). Revista Brasileira de Biologia, 53, 561-570. [ Links ]
Cei, J. M. (1980). Amphibians of Argentina. Monitore Zoologico Italiano (NS) Monografia 2. [ Links ]
Crump, M. L. (1974). Reproductive strategies in a tropical anuran community. University of Kansas Museum of Natural History Miscellaneous Publications, 61, 1-68. [ Links ]
Crump, M. L. (1983). Opportunistic cannibalism by amphibian larvae in temporary aquatic environments. American Naturalist, 121, 281-287. [ Links ]
Dalgetty, L., & Kennedy, M. W. (2010). Building a home from foam túngara frog foam nest architecture and three-phase construction process. Biology Letters, 6, 293-6. [ Links ]
De la Riva, I., & Gonzales, L. (1998). Sobre la presencia de Pleurodema guayapae Barrio, 1964 (Anura, Leptodactylidae) en Bolivia. Alytes, 16, 68-76. [ Links ]
Dobkin, D. S., & Gettinger, R. D. (1985). Thermal aspects of anuran foam nests. Journal of Herpetology, 19, 271-273. [ Links ]
Downie, J. R. (1988). Functions of the foam in the foam-nesting leptodactylid Physalaemus pustulosus. Herpetological Journal, 1, 302-307. [ Links ]
Duellman, W. E., & Trueb, L. (1986). Biology of Amphibians. New York: McGrawHill Book Co. [ Links ]
Duellman, W. E., & Veloso, A. (1977). Phylogeny of Pleurodema (Anura: Leptodactylidae): A biogeographic model. Occasional Papers, Museum of Natural History, University of Kansas, Lawrance, Kansas, 64, 1-46. [ Links ]
Faivovich, J., Ferraro, D. P., Basso, N. G., Haddad, C. F. D., Rodrigues, M. T., Wheeler, W. C., & Lavilla, E. O. (2012). A phylogenetic analysis of Pleurodema (Anura: Leptodactylidae: Leiuperinae) based on mitochondrial and nuclear gene sequences, with comments on the evolution of anuran foam nests. Cladistics, 28, 460-482. [ Links ]
Ferraro, D. P., & Casagranda, M. D. (2009). Geographic distribution of the genus Pleurodema in Argentina (Anura: Leiuperidae). Zootaxa, 2024, 33-55. [ Links ]
Giaretta, A. A., & Menin, M. (2003). Reproduction, phenology and mortality source of a species of Physalaemus (Anura, Leptodactylidae). Journal of Natural History, 38, 1711-1722. [ Links ]
Gosner, K. L. (1960). A simplified table for staging anuran embryos and larvae with notes on identification. Herpetologica, 16, 183-190. [ Links ]
Haddad, C. F. B., & Hödl, W. (1997). New reproductive mode in anurans: bubble nest in Chiasmocleis leucosticta (Mycrohylidae). Copeia, 3, 585-588. [ Links ]
Haddad, C. F. B., & Prado, C. P. A. (2005). Reproductive modes in frogs and their unexpected diversity in the Atlantic Forest of Brazil. BioScience, 55, 207-217. [ Links ]
Haddad, C. F. B., Pombal, J. P., & Gordo, M. (1990). Foam nesting in a hylid frog. Journal of Herpetology, 24, 225-226. [ Links ]
Halloy, M., & Fiaño, J. M. (2000). Oviposition site selection in Pleurodema borellii (Anura: Leptodactylidae) may be influenced by tadpole presence. Copeia, 2000(2), 606-609. [ Links ]
Heyer, W. R., McDiarmid, R. W., & Weigmann, D. L. (1975). Tadpoles, predation, and pond habitats in the tropics. Biotropica, 7, 100-111. [ Links ]
Hödl, W. (1990). An analysis of foam nest construction in the Neotropical frog Physalaemus ephippifer (Leptodactylidae). Copeia, 1990, 547-554. [ Links ]
Hödl, W. (1992). Reproductive behavior in the neotropical foamnesting frog Pleurodema diplolistris (Leptodac-tylidae). Amphibia-Reptilia, 13, 263-274. [ Links ]
Hulse, A. C. (1978). Food Habits of the Frog Lepidobatrachus llanensis (Amphibia, Anura, Leptodactylidae) in Argentina. Journal of Herpetology, 12(2), 258-260. [ Links ]
Hulse, A. C. (1979). Notes on the Biology of Pleurodema cinerea (Amphibia, Anura, Leptodactylidae) in Northwestern Argentina. Journal of Herpetology, 13(2), 153-156. [ Links ]
Kolenc, F., Baldo, D., Borteiro, C., Marangoni, F., Ferraro, D. P., & Faivovich, J. (2011). The Identity of Eupemphix fuscomaculatus Steindachner, 1864 (Amphibia: Anura) Copeia, 4, 513-522. [ Links ]
Maciel, D. B., & Nunes, I. (2010). A new species of four-eyed frog genus Pleurodema Tschudi, 1838 (Anura: Leiuperidae) from the rock meadows of Espinhaço range, Brazil. Zootaxa, 2640, 53-61. [ Links ]
Martori, R., Aún, L., & Vignolo, P. E. (1994). Aporte al conocimiento de la biología y distribución de Pleurodema tucumana (Anura, Leptodactylidae). Boletín de la Asociación Herpetológica Argentina, 10, 18-19. [ Links ]
Peixoto, O. L. (1982). Observações sobre a larva de Pleurodema diplolistris (Peters 1870) (Amphibia, Anura, Leptodactylidae). Revista Brasileira de Biologia, 42, 631-633. [ Links ]
Petranka, J. W., Hopey, M. E., Jennings, B. T., Baird, S. D., & Boone, S. J. (1994). Breeding habitat of wood frogs and American toads: the role of interespecic tadpole predation and adult choice. Copeia, 3, 691-697. [ Links ]
Prado, C. P. A., Uetanabaro, M., & Haddad, C. F. B. (2002). Description of a new reproductive mode in Leptodactylus (Anura, Leptodactylidae), with a review of the reproductive specialization towards terrestriality in the genus. Copeia, 2002, 1128-1133. [ Links ]
Salthe, S. N., & Duellman, W. E. (1973). Quantitative constraints associated with reproductive mode in anurans. In J. L. Vial (Ed.). Evolutionary biology of the anurans. Contemporary research on major problems (pp. 229-249). Columbia: University of Missouri Press. [ Links ]
Silva, W. R., Giaretta, A. A., & Facure, K. G. (2005). On the natural history of the South American pep-per frog, Leptodactylus labyrinthicus (Spix, 1824) (Anura: Leptodactylidae). Journal of Natural History, 39(7), 555-566. [ Links ]
Tárano, Z. (1998). Cover and ambient light influence nesting preferences in the Túngara frog Physalaemus pustulosus. Copeia, 1998, 250-251. [ Links ]
Valetti, J. A., Salas, N. E., & Martino, A. L. (2009). A new polyploid species of Pleurodema (Anura: Leiu-peridae) from Sierra de Comechingones, Córdoba, Argentina and redescription of Pleurodema kriegi (Müller, 1926). Zootaxa, 2073, 1-21. [ Links ]
Valetti, J. A., Otero, M. A., Grenat, P. R., & Martino, A. L. (2011). Clutch size. Pleurodema cordobae (Octoploid Pleurodema). Herpetological Review, 42(3), 413. [ Links ]
Vieira, W. L. S., Santana, G. G., & Arzabe, C. (2009). Diversity of reproductive modes in anurans communities in the Caatinga (dryland) of Northeastern Brazil. Biodiversity and Conservation, 18, 55-66. [ Links ]
Weigandt, M., Úbeda, C. A., & Díaz, M. (2004). The larva of Pleurodema bufoninum Bell, 1843, with comments on its biology and on the egg strings (Anura, Leptodactylidae). Amphibia-Reptilia, 25, 429-437. [ Links ]
Wells, K. D. (1977). The social behavior of anuran amphibians. Animal Behaviour, 25, 666-693. [ Links ]
Wilbur, H. M., & Alford, R. A. (1985). Priority effects in experimental pond communities, responses of Hyla to Bufo and Rana. Ecology, 66, 1106-l114. [ Links ]
Zina, J. (2006). Communal nests in Physalaemus pustulosus (Amphibia: Leptodactylidae): experimental evidence for female oviposition preferences and protection against desiccation. Amphibia-Reptilia, 27, 148-150. [ Links ]
Altig, R., & McDiarmid, R. W. (2007). Morphological diversity and evolution of egg and clutch structure in amphibians. Herpetological Monographs, 21, 1-32. [ Links ]
Barreto, L., & Andrade, G. V. (1995). Aspects of the reproductive biology of Physalaemus cuvieri (Anura: Leptodactylidae) in Northeastern Brazil. Amphibia-Reptilia, 16, 67-76. [ Links ]
Barrio, A. (1964). Especies crípticas del género Pleurodema que conviven en una misma área, identificadas por el canto nupcial (Anura, Leptodactylidae). Physis, 24(68), 471-489. [ Links ]
Cardoso, A. J., & Arzabe, C. (1993). Corte e desenvolvimento larvário de Pleurodema diplolistris (Anura: Leptodactylidae). Revista Brasileira de Biologia, 53, 561-570. [ Links ]
Cei, J. M. (1980). Amphibians of Argentina. Monitore Zoologico Italiano (NS) Monografia 2. [ Links ]
Crump, M. L. (1974). Reproductive strategies in a tropical anuran community. University of Kansas Museum of Natural History Miscellaneous Publications, 61, 1-68. [ Links ]
Crump, M. L. (1983). Opportunistic cannibalism by amphibian larvae in temporary aquatic environments. American Naturalist, 121, 281-287. [ Links ]
Dalgetty, L., & Kennedy, M. W. (2010). Building a home from foam túngara frog foam nest architecture and three-phase construction process. Biology Letters, 6, 293-6. [ Links ]
De la Riva, I., & Gonzales, L. (1998). Sobre la presencia de Pleurodema guayapae Barrio, 1964 (Anura, Leptodactylidae) en Bolivia. Alytes, 16, 68-76. [ Links ]
Dobkin, D. S., & Gettinger, R. D. (1985). Thermal aspects of anuran foam nests. Journal of Herpetology, 19, 271-273. [ Links ]
Downie, J. R. (1988). Functions of the foam in the foam-nesting leptodactylid Physalaemus pustulosus. Herpetological Journal, 1, 302-307. [ Links ]
Duellman, W. E., & Trueb, L. (1986). Biology of Amphibians. New York: McGrawHill Book Co. [ Links ]
Duellman, W. E., & Veloso, A. (1977). Phylogeny of Pleurodema (Anura: Leptodactylidae): A biogeographic model. Occasional Papers, Museum of Natural History, University of Kansas, Lawrance, Kansas, 64, 1-46. [ Links ]
Faivovich, J., Ferraro, D. P., Basso, N. G., Haddad, C. F. D., Rodrigues, M. T., Wheeler, W. C., & Lavilla, E. O. (2012). A phylogenetic analysis of Pleurodema (Anura: Leptodactylidae: Leiuperinae) based on mitochondrial and nuclear gene sequences, with comments on the evolution of anuran foam nests. Cladistics, 28, 460-482. [ Links ]
Ferraro, D. P., & Casagranda, M. D. (2009). Geographic distribution of the genus Pleurodema in Argentina (Anura: Leiuperidae). Zootaxa, 2024, 33-55. [ Links ]
Giaretta, A. A., & Menin, M. (2003). Reproduction, phenology and mortality source of a species of Physalaemus (Anura, Leptodactylidae). Journal of Natural History, 38, 1711-1722. [ Links ]
Gosner, K. L. (1960). A simplified table for staging anuran embryos and larvae with notes on identification. Herpetologica, 16, 183-190. [ Links ]
Haddad, C. F. B., & Hödl, W. (1997). New reproductive mode in anurans: bubble nest in Chiasmocleis leucosticta (Mycrohylidae). Copeia, 3, 585-588. [ Links ]
Haddad, C. F. B., & Prado, C. P. A. (2005). Reproductive modes in frogs and their unexpected diversity in the Atlantic Forest of Brazil. BioScience, 55, 207-217. [ Links ]
Haddad, C. F. B., Pombal, J. P., & Gordo, M. (1990). Foam nesting in a hylid frog. Journal of Herpetology, 24, 225-226. [ Links ]
Halloy, M., & Fiaño, J. M. (2000). Oviposition site selection in Pleurodema borellii (Anura: Leptodactylidae) may be influenced by tadpole presence. Copeia, 2000(2), 606-609. [ Links ]
Heyer, W. R., McDiarmid, R. W., & Weigmann, D. L. (1975). Tadpoles, predation, and pond habitats in the tropics. Biotropica, 7, 100-111. [ Links ]
Hödl, W. (1990). An analysis of foam nest construction in the Neotropical frog Physalaemus ephippifer (Leptodactylidae). Copeia, 1990, 547-554. [ Links ]
Hödl, W. (1992). Reproductive behavior in the neotropical foamnesting frog Pleurodema diplolistris (Leptodac-tylidae). Amphibia-Reptilia, 13, 263-274. [ Links ]
Hulse, A. C. (1978). Food Habits of the Frog Lepidobatrachus llanensis (Amphibia, Anura, Leptodactylidae) in Argentina. Journal of Herpetology, 12(2), 258-260. [ Links ]
Hulse, A. C. (1979). Notes on the Biology of Pleurodema cinerea (Amphibia, Anura, Leptodactylidae) in Northwestern Argentina. Journal of Herpetology, 13(2), 153-156. [ Links ]
Kolenc, F., Baldo, D., Borteiro, C., Marangoni, F., Ferraro, D. P., & Faivovich, J. (2011). The Identity of Eupemphix fuscomaculatus Steindachner, 1864 (Amphibia: Anura) Copeia, 4, 513-522. [ Links ]
Maciel, D. B., & Nunes, I. (2010). A new species of four-eyed frog genus Pleurodema Tschudi, 1838 (Anura: Leiuperidae) from the rock meadows of Espinhaço range, Brazil. Zootaxa, 2640, 53-61. [ Links ]
Martori, R., Aún, L., & Vignolo, P. E. (1994). Aporte al conocimiento de la biología y distribución de Pleurodema tucumana (Anura, Leptodactylidae). Boletín de la Asociación Herpetológica Argentina, 10, 18-19. [ Links ]
Peixoto, O. L. (1982). Observações sobre a larva de Pleurodema diplolistris (Peters 1870) (Amphibia, Anura, Leptodactylidae). Revista Brasileira de Biologia, 42, 631-633. [ Links ]
Petranka, J. W., Hopey, M. E., Jennings, B. T., Baird, S. D., & Boone, S. J. (1994). Breeding habitat of wood frogs and American toads: the role of interespecic tadpole predation and adult choice. Copeia, 3, 691-697. [ Links ]
Prado, C. P. A., Uetanabaro, M., & Haddad, C. F. B. (2002). Description of a new reproductive mode in Leptodactylus (Anura, Leptodactylidae), with a review of the reproductive specialization towards terrestriality in the genus. Copeia, 2002, 1128-1133. [ Links ]
Salthe, S. N., & Duellman, W. E. (1973). Quantitative constraints associated with reproductive mode in anurans. In J. L. Vial (Ed.). Evolutionary biology of the anurans. Contemporary research on major problems (pp. 229-249). Columbia: University of Missouri Press. [ Links ]
Silva, W. R., Giaretta, A. A., & Facure, K. G. (2005). On the natural history of the South American pep-per frog, Leptodactylus labyrinthicus (Spix, 1824) (Anura: Leptodactylidae). Journal of Natural History, 39(7), 555-566. [ Links ]
Tárano, Z. (1998). Cover and ambient light influence nesting preferences in the Túngara frog Physalaemus pustulosus. Copeia, 1998, 250-251. [ Links ]
Valetti, J. A., Salas, N. E., & Martino, A. L. (2009). A new polyploid species of Pleurodema (Anura: Leiu-peridae) from Sierra de Comechingones, Córdoba, Argentina and redescription of Pleurodema kriegi (Müller, 1926). Zootaxa, 2073, 1-21. [ Links ]
Valetti, J. A., Otero, M. A., Grenat, P. R., & Martino, A. L. (2011). Clutch size. Pleurodema cordobae (Octoploid Pleurodema). Herpetological Review, 42(3), 413. [ Links ]
Vieira, W. L. S., Santana, G. G., & Arzabe, C. (2009). Diversity of reproductive modes in anurans communities in the Caatinga (dryland) of Northeastern Brazil. Biodiversity and Conservation, 18, 55-66. [ Links ]
Weigandt, M., Úbeda, C. A., & Díaz, M. (2004). The larva of Pleurodema bufoninum Bell, 1843, with comments on its biology and on the egg strings (Anura, Leptodactylidae). Amphibia-Reptilia, 25, 429-437. [ Links ]
Wells, K. D. (1977). The social behavior of anuran amphibians. Animal Behaviour, 25, 666-693. [ Links ]
Wilbur, H. M., & Alford, R. A. (1985). Priority effects in experimental pond communities, responses of Hyla to Bufo and Rana. Ecology, 66, 1106-l114. [ Links ]
Zina, J. (2006). Communal nests in Physalaemus pustulosus (Amphibia: Leptodactylidae): experimental evidence for female oviposition preferences and protection against desiccation. Amphibia-Reptilia, 27, 148-150. [ Links ]
*Correspondencia: Julián Alonso Valetti: Ecología, Departamento de Ciencias Naturales, Facultad de Ciencias Exactas, Físico-Químicas y Naturales, Universidad Nacional de Río Cuarto, Ruta Nacional N° 8 - km 601, (X5804BYA) Río Cuarto, Argentina; jvaletti@exa.unrc.edu.ar
Pablo Raúl Grenat: Ecología, Departamento de Ciencias Naturales, Facultad de Ciencias Exactas, Físico-Químicas y Naturales, Universidad Nacional de Río Cuarto, Ruta Nacional N° 8 - km 601, (X5804BYA) Río Cuarto, Argentina; pgrenat@exa.unrc.edu.ar
Mariana Baraquet: Ecología, Departamento de Ciencias Naturales, Facultad de Ciencias Exactas, Físico-Químicas y Naturales, Universidad Nacional de Río Cuarto, Ruta Nacional N° 8 - km 601, (X5804BYA) Río Cuarto, Argentina; mbaraquet@exa.unrc.edu.ar
Adolfo Ludovico Martino: Ecología, Departamento de Ciencias Naturales, Facultad de Ciencias Exactas, Físico-Químicas y Naturales, Universidad Nacional de Río Cuarto, Ruta Nacional N° 8 - km 601, (X5804BYA) Río Cuarto, Argentina; amartino@exa.unrc.edu.ar
1. Ecología, Departamento de Ciencias Naturales, Facultad de Ciencias Exactas, Físico-Químicas y Naturales, Universidad Nacional de Río Cuarto, Ruta Nacional N° 8 - km 601, (X5804BYA) Río Cuarto, Argentina; jvaletti@exa.unrc.edu.ar, pgrenat@exa.unrc.edu.ar, mbaraquet@exa.unrc.edu.ar, amartino@exa.unrc.edu.ar
Received 11-II-2013. Corrected 20-VII-2013. Accepted 21-VIII-2013.


{kind=link}
{kind=link}
{kind=link}
{kind=link}