










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.62 n.1 San José Jan./Mar. 2014
Composición por especies y tallas de los peces en la laguna Barra de Navidad, Pacífico central mexicano
Species and size composition of fishes in Barva de Navidad lagoon, Mexican central Pacific
Species and size composition of fishes in Barva de Navidad lagoon, Mexican central Pacific
Gaspar González-Sansón1*, Consuelo Aguilar-Betancourt1, Daniel Kosonoy-Aceves1, Gabriela Lucano-Ramírez1, Salvador Ruiz-Ramírez1, Juan Ramón Flores-Ortega1, Ángel Hinojosa-Larios1 & Francisco de Asís Silva-Bátiz1
*Dirección para correspondencia:
Abstract
Coastal lagoons are considered important nursery areas for many coastal fishes. Barra de Navidad coastal lagoon (3.76km2) is important for local economy as it supports tourism development and artisanal fisheries. However, the role of this lagoon in the dynamics of coastal fish populations is scarcely known. Thus, the objectives of this research were: to characterize the water of the lagoon and related weather conditions, to develop a systematic list of the ichthyofauna, and to estimate the proportion of juveniles in the total number of individuals captured of most abundant species. Water and fish samples were collected between March 2011 and February 2012. Physical and chemical variables were measured in rainy and dry seasons. Several fishing gears were used including a cast net, beach purse seine and gillnets of four different mesh sizes. Our results showed that the lagoon is most of the time euhaline (salinity 30-40ups), although it can be mixopolyhaline (salinity 18-30ups) during short periods. Chlorophyll and nutrients concentrations suggested eutrophication in the lagoon. Mean water temperature changed seasonally from 24.9C (April, high tide) to 31.4C (October, low tide). Considering ichthyofauna species, a total of 36 448 individuals of 92 species were collected, 31 of them adding up to 95% of the total of individuals caught. Dominant species were Anchoa spp. (44.6%), Diapterus peruvianus (10.5%), Eucinostomus currani (8.1%), Cetengraulis mysticetus (7.8%), Mugil curema (5.2%) and Opisthonema libertate (4.5%). The lagoon is an important juvenile habitat for 22 of the 31 most abundant species. These included several species of commercial importance such as snappers (Lutjanus argentiventris, L. colorado and L. novemfasciatus), snook (Centropomus nigrescens) and white mullet (Mugil curema). Other four species seem to use the lagoon mainly as adults. This paper is the first contribution on the composition of estuarine ichthyofauna in Jalisco State, and it also constitutes the more complete reference on the detailed size composition of a set of dominant species in a coastal lagoon in the central Mexican Pacific.
Key words: estuarine ichthyofauna, coastal lagoon, nursery area, Central Mexican Pacific.
Resumen
Las lagunas costeras son consideradas áreas de crianza importantes para muchas especies de peces costeros. La laguna costera Barra de Navidad (3.76km2) es importante para la economía local y soporta un desarrollo turístico y pesquerías artesanales. Sin embargo, el rol de esta laguna en la dinámica de las poblaciones de peces costeros es poco conocido. Los objetivos de la investigación fueron: caracterizar el agua de la laguna y las condiciones climáticas relacionadas, elaborar el elenco sistemático de la ictiofauna y estimar la proporción de juveniles en el total de individuos capturados de las especies más abundantes. Las operaciones de recolecta de peces se realizaron entre marzo 2011 y febrero 2012. Se utilizaron varios artes de pesca diferentes que incluyeron atarraya, chinchorro playero y redes de agalla con cuatro tamaños de malla diferentes. Se midieron las variables físicas y químicas en épocas de lluvias y de secas. La laguna es euhalina (salinidad 30-40 ups) la mayor parte del tiempo, aunque en determinados periodos cortos puede tener características mixopolihalinas (salinidad 18-30 ups). Las concentraciones de clorofila y nutrientes indican que la laguna está eutrofizada. La temperatura media del agua varió estacionalmente de 24.9C (abril, pleamar) a 31.4C (octubre, bajamar). Se recolectaron en total 36 448 individuos, pertenecientes a 92 especies, de las cuales 31 tienen una relevancia ecológica con base en el número de individuos capturados. Las especies dominantes fueron: Anchoa spp. (44.6%), Diapterus peruvianus (10.5%), Eucinostomus currani (8.1%), Cetengraulis mysticetus (7.8%), Mugil curema (5.2%) y Opisthonema libertate (4.5%). La laguna es un hábitat de juveniles importante para 22 de las 31 especies más abundantes. Estas incluyeron algunas especies de importancia comercial como los pargos (Lutjanus argentiventris, L. colorado y L. novemfasciatus), el robalo (Centropomus nigrescens) y la lisa (Mugil curema). Otras cuatro especies parecen utilizar la laguna principalmente como adultos. El presente trabajo es la primera contribución sobre la composición de la ictiofauna estuarina en el Estado de Jalisco y es también la referencia más completa hasta el momento sobre la composición por tallas del conjunto de especies dominantes en una laguna costera del Pacífico central mexicano.
Palabras clave: ictiofauna estuarina, laguna costera, área de crianza, Pacífico central mexicano.
La laguna de Barra de Navidad, declarada sitio RAMSAR el 2 de febrero 2008, es el principal cuerpo de agua litoral (humedal costero) de la Bahía de Navidad y en general, de la costa sur de Jalisco en el Pacífico mexicano. Por su extensión (376 hectáreas), es el tercer cuerpo de agua natural más grande de la costa del Estado de Jalisco. Una de las características de mayor valor de la laguna, además de los bienes y servicios que brinda para el desarrollo de las actividades de pesca artesanal, turísticas y recreacionales, es la presencia de bosques de mangle en buen estado de conservación. La extensión total de su cobertura es de 571 hectáreas, siendo el segundo manglar más extenso del Estado de Jalisco (Silva-Bátiz, González-Sansón & Godínez-Domínguez, 2010).
Algunos aspectos aislados de la estructura y la dinámica del ecosistema en esta laguna han sido estudiados por Sandoval, Flores, Zaragoza, Day & Estrada (1988), Meyer, Velázquez-González & Patiño-Barragán (2006), Méndez-Linares, López-Portillo, Hernández-Santana, Ortiz-Pérez & Oropeza-Orozco (2007) y Hernández, Serrano, Hernández & Robles (2012). La información existente, sin embargo, es muy fragmentaria, no aborda los procesos ecológicos más importantes y no está actualizada. Por estos motivos, su utilidad es muy limitada en las condiciones actuales de la laguna, que ha sufrido modificaciones notables en los últimos años. Los cambios más importantes han sido la modificación de los márgenes y el relleno parcial, con la consecuente alteración de los patrones de circulación y un azolvamiento acelerado (Silva-Bátiz et al., 2012). Desafortunadamente, no existen trabajos publicados sobre la composición de la ictiofauna y los factores que provocan sus variaciones espaciales y temporales en esta importante laguna costera.
Las lagunas costeras son importantes hábitats para juveniles de numerosas especies de organismos marinos y en particular los peces. La definición de tales ecosistemas como áreas de crianza esenciales, depende en gran medida de la contribución que hacen a las poblaciones de adultos que viven en la plataforma adyacente (Beck et al., 2001; Dahlgren et al., 2006). La situación particular de cada cuerpo de agua estuarino con respecto a este importante aspecto del ciclo de vida de los peces, requiere de una investigación específica sobre las características físicas y químicas del cuerpo de agua, las especies que aparecen en tales ecosistemas y los estadios de desarrollo predominantes de cada especie dentro del ambiente estuarino. En la revisión de la literatura, que abarcó todo el Pacífico central mexicano, solo se encontraron dos trabajos que incluyen datos limitados sobre la composición por tallas de los peces en sistemas lagunares costeros (Amezcua-Linares, 1977; Mendoza, Castillo-Rivera, Zárate-Hernández & Ortiz-Burgos, 2009). Esto indica que existe un gran vacío de información cuantitativa sobre la existencia de juveniles en los sistemas estuarinos de la región.
Los objetivos de esta investigación se orientaron a: a) caracterizar el agua de la laguna y algunos factores climáticos, b) elaborar el elenco sistemático de la ictiofauna en la laguna Barra de Navidad y c) definir la proporción de juveniles en el total de individuos capturados de las especies más abundantes.
Materiales y Métodos
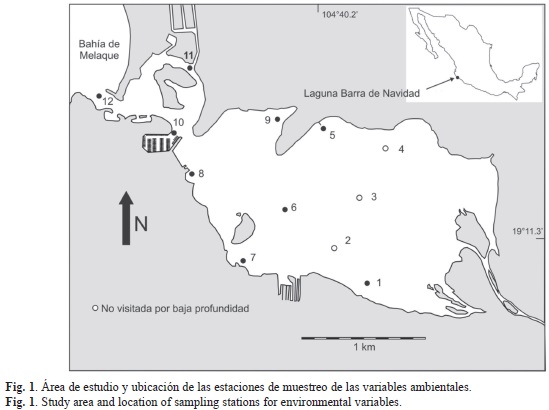
Área de estudio: La Laguna Barra de Navidad se ubica en el litoral centro occidental mexicano del Océano Pacífico (19°11’25’’ N - 104°39’53’’ W), en el extremo de la costa sur del Estado de Jalisco, municipio Cihuatlán, al sureste de la localidad denominada Barra de Navidad (Fig. 1). Fue clasificada por Lankford (1976), como tipo III-A (III-B), lo que indica un dominio de las olas y corrientes costeras en su formación. La laguna tiene comunicación permanente con el mar mediante una boca de 98m de ancho, la cual no se modifica estacionalmente debido a que sus orillas están cubiertas con material rocoso. Las profundidades típicas no exceden los dos metros.
Determinación de variables ambientales: Los muestreos para determinar las características generales del agua se realizaron en nueve estaciones fijas, dos veces al día (pleamar y bajamar), en tres fechas representativas de las estaciones climáticas (viajes de muestreo de un día en abril, julio y octubre 2011). Tres estaciones definidas al comenzar la investigación no pudieron ser visitadas debido a la baja profundidad, que impidió la llegada de la embarcación (Fig. 1). Se utilizó una sonda YSI 556MPS para la medición in situ de salinidad, temperatura, pH y oxígeno disuelto. Se tomaron muestras de agua para la medición de las concentraciones de nitratos, nitritos, amonio, fosfatos y clorofila a en el laboratorio, siguiendo la metodología descrita en Parsons, Maita & Lalli (1984). Los datos mensuales de precipitación y temperatura media del aire para cada año del periodo 1934-2011, se tomaron de los registros de la estación meteorológica de Cihuatlán (1914'17” N-10434' 09” W) ubicada a 11.4km de la laguna. Se consideró que los valores mensuales de precipitación y temperatura media del aire del año 2011 eran significativamente diferentes del promedio histórico, si los valores del año quedaban por fuera de los intervalos de confianza calculados para la serie histórica.
Muestreo de la ictiofauna: Para determinar la diversidad de especies y la estructura por tallas, se realizaron 42 operaciones de recolecta de peces, espaciadas regularmente cada 7-9 días, entre marzo 2011 y febrero 2012. Las capturas se distribuyeron por toda la laguna, para garantizar un muestreo representativo de todo el cuerpo de agua. Se emplearon tres artes de pesca diferentes: a) atarraya de 3m de largo y 2.5cm de luz de malla; b) redes de enmalle de 60m de largo y de 7.0, 7.6, 8.9 y 10.2cm de luz de malla; c) chinchorro playero experimental de 10m de largo y 1cm de luz de malla en el copo. Este amplio espectro de métodos de captura buscó reducir el sesgo introducido por la selectividad de cada arte de pesca e incrementar los hábitats incluidos en el muestreo. La atarraya se utilizó en sitios seleccionados aleatoriamente en cada viaje de pesca en toda la laguna; se realizaron entre 40 y 50 lances de atarraya entre las 4:00 y las 7:30pm de cada día de muestreo. Las redes se colocaron en sitios seleccionados aleatoriamente en cada viaje de muestreo, los cuales fueron ubicados siempre en la porción centro-occidental de la laguna, que es la única zona donde este tipo de arte puede operar de forma efectiva; las redes se colocaron a las 7pm y se dejaron en el agua hasta las 7am del siguiente día en que fueron recogidas. El chinchorro se operó en dos sitios con zonas de playa en el norte y en el sur de la laguna; se realizaron cinco lances en cada sitio entre las 4:00 y las 7:30pm de cada día de muestreo. Este diseño de muestreo permitió obtener una composición por tallas de los individuos capturados, que refleja de forma más adecuada el espectro de tamaños de los individuos de cada especie presente en la laguna.
Los peces capturados por tipo de arte de pesca, fueron preservados en recipientes con hielo y fueron transportados al laboratorio. El análisis biológico incluyó su identificación, el conteo de los organismos por artes de pesca y la medición de la longitud total (mm) con un ictiómetro. La identificación de las especies se basó en Fischer et al. (1995) y Amezcua-Linares (2008). Los nombres válidos de las especies se presentan según Eschmeyer (2013) y los taxones superiores según Nelson (2006). La información sobre la longitud total de primera maduración para las especies incluidas en este estudio se obtuvo de la base de datos en línea FishBase (Froese & Pauly, 2013). Para comparar la composición por especies entre las diferentes artes de pesca, se utilizó el coeficiente de correlación por rangos de Spearman (rs) según Zar (2010). Se consideró que el orden de importancia según el número total de individuos capturados por las artes no difiere, si el valor de rs es significativamente mayor que 0. En caso contrario se considera que la diferencia es significativa. Las distribuciones de tallas en peces, raramente cumplen con una distribución normal, por lo cual, estas se representaron mediante diagramas de caja que permiten determinar gráficamente la asimetría de las distribuciones con base en la posición de la mediana y la ubicación de los cuartiles 25%-75% respecto al rango de valores no extremos. Para evaluar la significancia estadística de las diferencias en la composición por tallas por especie, cuando esta estaba representada en las tres artes de pesca, se realizó una prueba de Kruskal-Wallis (H). Cuando la prueba indicó diferencia significativa, se utilizó la prueba de comparaciones múltiples recomendada por Siegel & Castellan (1988) para determinar diferencias entre pares de artes. Cuando la especie estaba representada en sólo dos artes de pesca, se utilizó la prueba no paramétrica U de Mann y Whitney (Zar, 2010). En todos los análisis, el nivel de significación fue 0.05. Los cálculos fueron realizados mediante el programa Statistica 7.1 (StatSoft, 2006).
Resultados
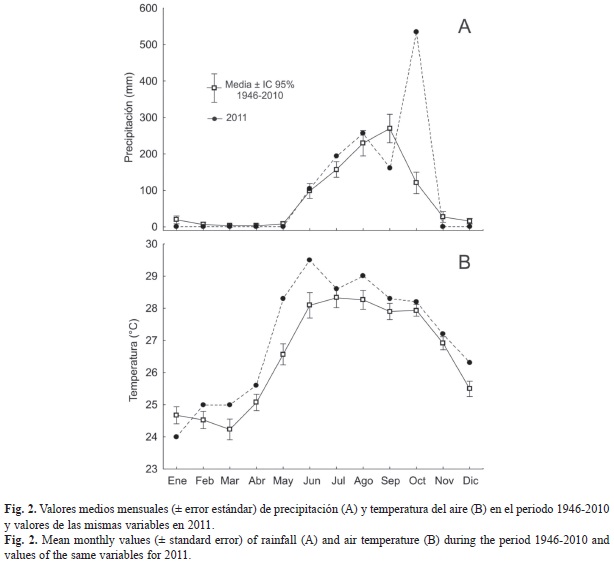
Características ambientales: Los promedios de los valores de precipitación mensual calculados para el periodo 1943-2010 indican que existe una estacionalidad muy definida en el régimen de precipitaciones que afecta la laguna, con una estación de lluvias que abarca desde junio hasta octubre y una estación de secas el resto del año. En 2011 se comportó según este patrón hasta el mes de agosto, pero septiembre resultó significativamente más seco que la media histórica, y octubre presentó el registro más alto de precipitación para todo el periodo analizado (1943-2010). Esto último se debió a las fuertes lluvias asociadas al huracán Jova (categoría tres en la escala de Saffir-Simpson), que afectó esta zona entre el 11 y el 13 de octubre 2011. Los restantes meses no difieren significativamente de la media histórica (Fig. 2A).
La temperatura del aire media mensual calculada para el periodo 1946-2010 (no hay datos para 1943-1945) refleja la estacionalidad moderada de esta variable, la cual presenta un cambio de sólo cuatro grados, desde 24.2C en el mes más frío (marzo) hasta 28.3C en el mes más cálido (julio). El 2011 se puede considerar ligeramente cálido con respecto a la tendencia media, pues en 11 meses presentó temperaturas medias significativamente superiores al valor medio de la serie analizada (Fig. 2B).
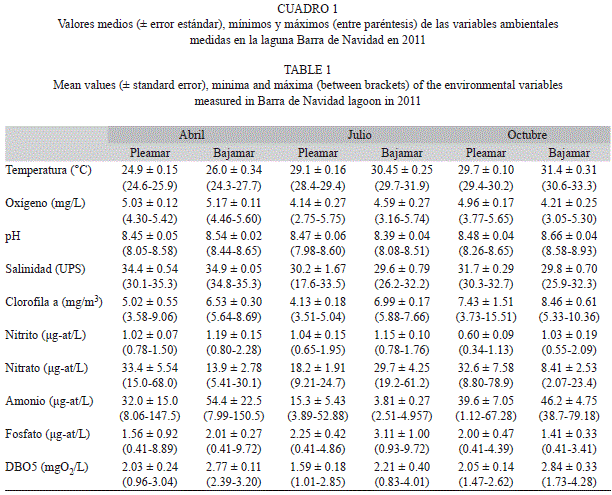
Los valores medios de la temperatura del agua de la laguna, presentaron cambios moderados a lo largo del año, y mostraron una variación de entre cuatro y cinco grados Celsius entre las mediciones del mes más frío y el más cálido. La salinidad también varió poco entre la época de seca y la de lluvia, exhibiendo cambios no superiores a cinco UPS entre los valores medios respectivos. Menos variables aún, resultaron los promedios de la concentración de oxígeno disuelto y el pH. En todos los casos, los valores relativamente bajos del error estándar (menores al 5% de la media en la mayor parte de las estimaciones) en las cuatro variables descritas, indican poca variación espacial (entre estaciones), aunque en algunas de ellas se notaron valores extremos. Las otras variables medidas en el agua de la laguna mostraron mayor variación temporal y espacial, pero no resulta claro un patrón que se pueda relacionar con la ubicación de las estaciones o las épocas del año (Cuadro 1).
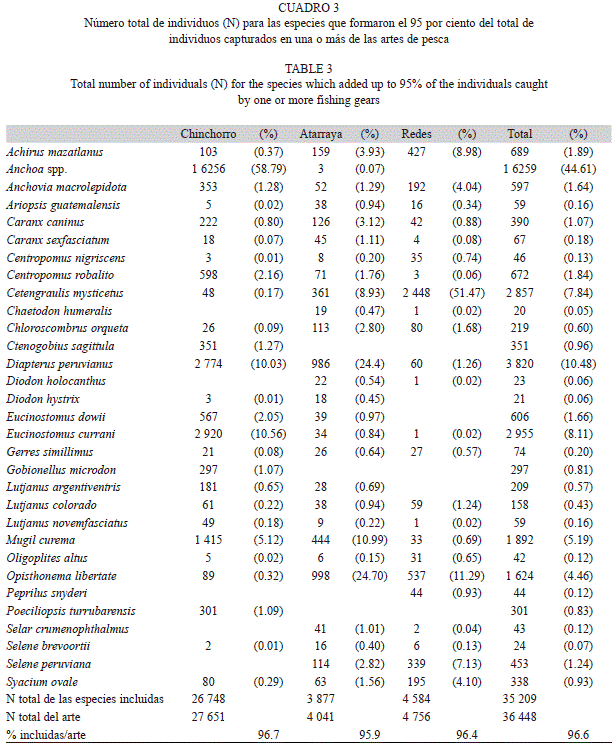
Ictiofauna: Se recolectaron en total 36 448 individuos, pertenecientes a 92 especies, 45 familias y 15 órdenes (Cuadro 2). Según el arte de pesca utilizado, las especies se ordenaron según el número total de individuos capturados y se calculó el número acumulado de individuos. Se seleccionaron como especies típicas de cada arte de recolecta, aquellas que conformaron 95% del total de individuos capturados por ese arte de pesca. La composición por especies (en términos del orden de abundancia) del chinchorro difirió significativamente de la obtenida con atarraya (rs=0.38, p=0.070) y de la obtenida con las redes de agalla (rs=0.13, p=0.591). Por su parte, la composición por especies no fue significativamente diferente entre la atarraya y las redes de agalla (rs=0.66, p<0.001). Al combinar los resultados para las tres artes de pesca, se identificaron 31 especies que conforman alrededor del 95% de todos los individuos capturados con todas las artes de pesca (Cuadro 3). Más de la mitad de los individuos (54%) pertenecieron a la familia Engraulidae y estuvieron dominados por varias especies del género Anchoa, y las especies Cetengraulis mysticetus y Anchovia macrolepidota. Las especies del género Anchoa, se determinaron con base en individuos seleccionados, pero resultó prácticamente imposible hacer una identificación masiva confiable, por lo que este grupo se excluye de los análisis por especies que se presentan más adelante. En segundo lugar se ubicó la familia Gerreidae (21%) con un amplio dominio de las especies Diapterus peruvianus y Eucinostomus currani. Con mucha menor representación numérica, se ubicaron, en orden decreciente, las familias Mugilidae (5.3%, principalmente Mugil curema), Clupeidae (4.6%, dominada por Opisthonema libertate), Carangidae (3.8%, principalmente Selene peruviana y Caranxcaninus), Centropomidae (2.2% con amplio dominio de Centropomus robalito y presencia ocasional de Centropomus nigrescens) y Achiridae (1.9%, representada por Achirus mazatlanus). Otros taxones tuvieron una representación numérica muy baja (<1%).
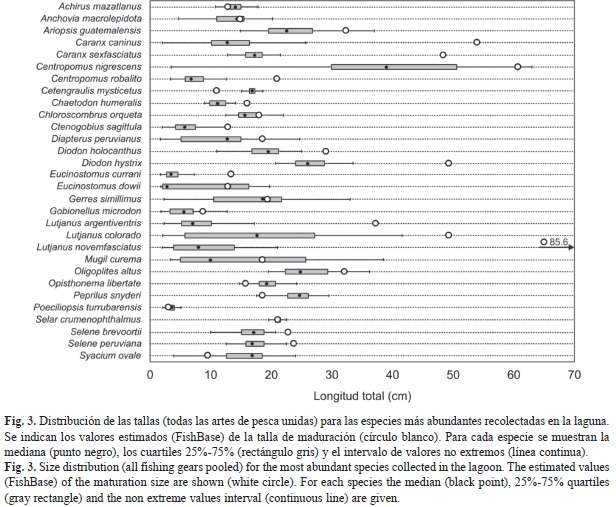
El análisis de la composición por tallas general de las especies más abundantes permitió identificar aquellas que se presentan predominantemente como juveniles y las que aparecen en la laguna como adultos (Fig. 3). En el caso de las especies Caranx caninus, C. sexfasciatus, Centropomus robalito, Chaetodon humeralis, Diodon holocanthus, D. hystrix, Eucinostomus currani, Lutjanus argentiventris, L. colorado, L. novemfasciatus, Selene brevoorti y S. peruviana, los individuos captura-dos estuvieron siempre, con independencia del arte de pesca, por debajo de la talla de primera maduración estimado para cada especie. Otras especies presentaron tallas por debajo y por encima de la talla de primera maduración, variando los porcentajes de acuerdo a cada especie, aunque en todos los casos, más de la mitad de los individuos se incluye dentro de la categoría de juveniles. Esta situación se encontró en siguientes especies, el porcentaje de individuos por debajo de la talla de maduración se presenta entre paréntesis: Anchovia macrolepidota (52.5), Ariopsis guatemalensis (85.5), Centropomus nigrescens (91.6), Chloroscombrus orqueta (72.4), Diapterus peruvianus (96.0), Eucinostomus dowii (63.8), Gerres simillimus (52.1), Gobionellus micro-don (80.4), M. curema (54.4) y Oligoplites altus (82.1). Por otra parte, en varias especies (A. mazatlanus, C. mysticetus, O. libertate, Peprilus snyderi, Poeciliopsis turrubarensis y Syacium ovale) más del 90% de los ejemplares capturados, presentaron longitudes mayores que la talla de primera maduración.
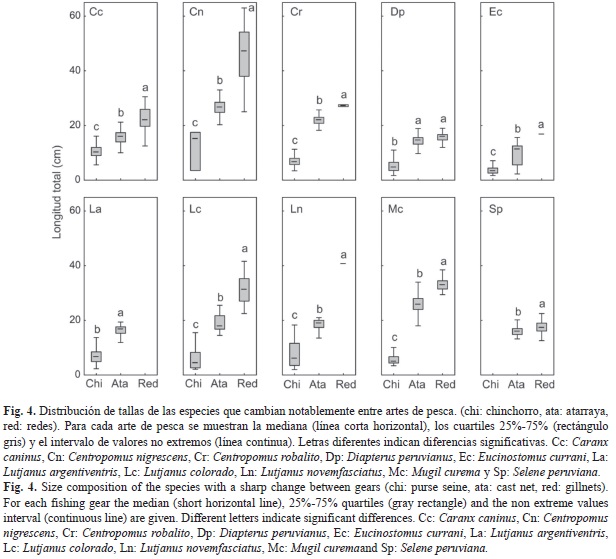
La representación diferencial de las tallas capturadas por cada arte de pesca, indica que existen especies en la laguna que presentan un amplio intervalo de tallas y que los valores de las longitudes totales difieren significativamente entre artes de pesca (Fig. 4). En este grupo se incluyen Caranx caninus (H=248.6, p<0.001), Centropomus nigrescens (H=26.3, p<0.001), C. robalito (H=204.2, p<0.001), Diapterus peruvianus (H=1 136.9, p<0.001), Eucinostomus currani (U=7 586.5, p<0.001), Lutjanus argentiventris (U=75.5, p<0.001), L. colorado (H=124.7, p<0.001), L. novemfasciatus (U=10.0, p<0.001), Mugil curema (H=737.6, p<0.001) y Selene peruviana (U=10951, p<0.001). Las pruebas de comparaciones por pares indicaron en la mayoría de los casos, que las tres artes de pesca presentan diferencias significativas entre sí, siendo el orden siempre el mismo (chinchorro<atarraya<redes). Solamente en el caso de D. peruvianus, se encontró que la composición por tallas no presentaba diferencias significativas entre atarraya y redes. En la mayoría de estos casos, las tallas están siempre por debajo del largo de primera maduración reportado en la literatura. Sólo en la especie M. curema, las tallas capturadas por la atarraya y las redes son superiores al largo de maduración reportado para la especie.
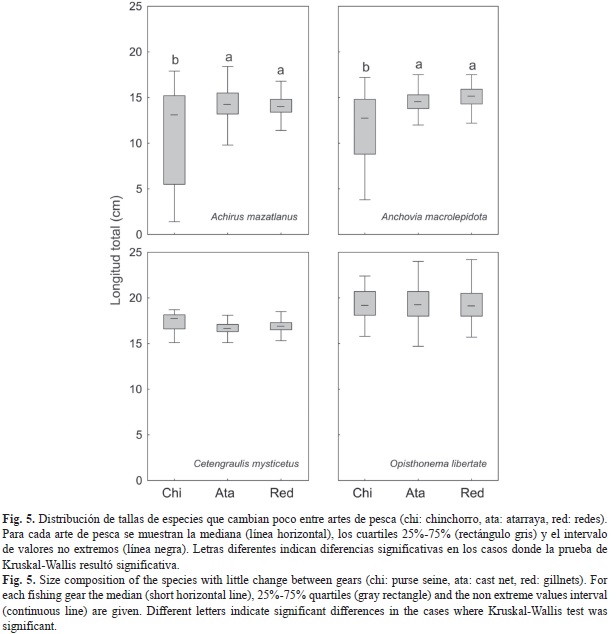
Otro grupo de especies presentó distribuciones por tallas con valores de la mediana similares entre artes de pesca (Fig. 5). Cetengraulis mysticetus (H=1.325, p=0.515) y Opisthonema libertate (H=0.030, p=0.985) presentaron una homogeneidad notable en las tallas, las cuales son superiores, en todo los casos, a la talla de maduración reportada para estas especies. Achirus mazatlanus y Anchovia macrolepidota presentaron valores de la mediana muy similares entre artes de pesca, pero la distribución de tallas en el chinchorro, presentó una asimetría importante debido a la represen-tación de tallas pequeñas en este arte de pesca. En el caso de A. mazatlanus, la prueba de Kruskal-Wallis resultó significativa (H=17.79, p<0.001), pero las comparaciones por pares indicaron que no hay diferencias significativas entre las tallas capturadas por la atarraya y las redes, pero sí con el chichorro. En el caso de esta especie, el 98% de los individuos presentó tallas superiores a la de primera maduración (Fig. 3).En el caso de A. macrolepidota las pruebas dieron un resultado idéntico. En este caso la prueba de Kruskal-Wallis también resultó significativa (H=107.09, p<0.001), pero las comparaciones por pares indicaron que no hay diferencias significativas entre las tallas capturadas por la atarraya y la redes. La mayoría de los peces recolectados presentó tallas menores a la de primera maduración (Fig. 3).
Discusión
Según los valores de salinidad encontrados en esta investigación y tomando como referencia las categorías identificadas en el Sistema de Venecia (Anónimo, 1958), la laguna se comporta la mayor parte del tiempo como un sistema euhalino (salinidad 30-40) que puede pasar a mixopolihalino (salinidad 18-30) en época de lluvias. Sólo en determinadas zonas al Este y durante un tiempo muy corto (días) el sistema puede llegar a ser mixomesohalino o incluso mixooligohalino. Resultados similares fueron presentados para esta laguna por Sandoval et al. (1988) y Meyer et al. (2006). Se puede concluir que la laguna es un sistema estuarino bien mezclado por las mareas (Mann & Lazier, 1996), las cuales dominan sobre el flujo de agua dulce.
Las concentraciones de nitritos, amonio y fosfato medidas en esta investigación son notablemente mayores que las registradas diez años atrás para esta misma laguna por Meyer et al. (2006), mientras que los valores de nitrato son semejantes a los informados por esos autores, los cuales consideraron que la laguna se encontraba en proceso de eutrofización. Los valores más altos de algunos compuestos nitrogenados y de fósforo en la actualidad indican que el proceso de eutrofización de la laguna ha seguido avanzando. Este proceso no ha afectado severamente la concentración de oxígeno disuelto ni los valores de pH, que se encuentran dentro de los valores normales para este tipo de ambiente, mientras que los valores de clorofila son bajos en comparación con otras lagunas mexicanas (Contreras & Warner, 2004). No es posible determinar el impacto que este proceso de eutrofización ha tenido en la ictiofauna, ya que no existen antecedentes de investigaciones sobre la misma en esta laguna. No obstante, la información obtenida fundamenta la necesidad urgente de investigar este tema.
No se encontró ningún trabajo previo publicado sobre la ictiofauna de la laguna Barra de Navidad o de cualquier otra laguna costera en el estado de Jalisco, por lo que el presente trabajo constituye una contribu-ción novedosa al conocimiento de los peces estuarinos en esta región costera de México. Se localizaron artículos publicados sobre la ictiofauna estuarina en otras partes del Pacífico central mexicano que incluyen la península de Baja California (Danemann & Cruz-Agüero, 1993; Arellano, Cruz-Agüero & Cota, 1996; Acevedo, 1997; Cruz-Agüero & Cota, 1998; Rosales, 2004; González, Cruz-Agüero, Cruz-Agüero& Ruiz, 2005; Rodríguez-Romero et al., 2011) y los estados de Sonora (Grijalva, Núñez- Quevedo & Castro-Longoria,1996), Sinaloa (Amezcua-Linares, 1977; Amezcua-Linares,Madrid & H. Aguirre 2006), Nayarit (Álvarez-Rubio,Amezcua-Linares & Yáñez-Arancibia, 1986; Flores Verdugo et al., 1990; Benítez et al., 2007), Colima (Cabral-Solís & Espino-Barr, 2004), Michoacán (Raz-Guzman & Huidobro, 2002), Guerrero (Yáñez-Arancibia., 1978), Oaxaca (Tapia-García, Suárez-Núñez, Cerdenares, Macuitl-Montes & García-Abad, 1998; Mendoza et al., 2009) y Chiapas (Díaz, Aguirre & Cano, 2006). Estos trabajos constituyen en su conjunto un aporte importante al conocimiento de la ictiofauna estuarina en la región. Al cotejar las especies que han sido encontradas por esos autores en los sistemas estuarinos, se constata que las más frecuentes y características de estos ambientes coinciden en gran medida con las encontradas en el presente trabajo como dominantes. Solamente dos especies encontradas con mucha frecuencia por otros autores no fueron recolectadas en nuestras muestras. Se trata de Lile stolifera, que apareció en 12 lagunas de varios estados (Amezcua-Linares, 1977; Yáñez-Arancibia, 1978; Álvarez-Rubio et al., 1986; Flores-Verdugo et al., 1990; Tapia-García et al., 1998) y Cichlasoma trimaculatum, que fue una especie muy frecuente en nueve lagunas de Gue-rrero (Yáñez-Arancibia, 1978). Ninguno de los trabajos citados, incluye un análisis detallado de las longitudes de los ejemplares capturados. En dos trabajos se dan datos parciales sobre composición por tallas de la ictiofauna estuarina. Mendoza et al. (2009) solo incluyen los rangos de tallas de los ejemplares capturados y Amezcua-Linares (1977) da las composiciones por tallas mensuales de dos especies.
El análisis de la composición por tallas, combinado con los estimados de talla de prime-ra maduración obtenidos de FishBase, permitió definir cuáles especies se presentan principal-mente como juveniles en la laguna y cuáles utilizan el ecosistema principalmente como adultos. Según nuestros resultados, la laguna es un hábitat de juveniles importante para, al menos, 22 especies. De éstas, 12 se presentan con tallas pequeñas bien distantes de la talla de primera maduración, por lo que no se descarta que con tallas mayores, aún siendo juveniles, se muevan fuera de la laguna a otros hábitats costeros que también sirvan como hábitats de juveniles. Este uso progresivo de hábitats diferentes en la fase juvenil ha sido consignado por otros autores (Mumby et al., 2004; Nage-lkerken, 2009). Otras diez especies aparecen principalmente como juveniles, pero también presentan individuos que por su talla, pueden ser subadultos e incluso adultos jóvenes que están madurando. En este caso puede suponerse que estas especies usan la laguna como su hábitat de crianza principal.
Otro grupo de especies parece utilizar la laguna principalmente como adultos. En este caso se encuentran dos especies pelágicas (Opisthonema libertate y Cetengraulis mysticetus) que entran periódicamente a la laguna buscando refugio y posiblemente alimento. También se encuentran en este caso los dos lenguados más abundantes de la laguna. Todas son especies marinas que visitan el ambiente estuarino. Por otra parte, en la laguna se encuentran con frecuencia, ejemplares adultos de un poecílido (P. turrubarensis) que es una especie dulceacuícola secundaria.
El presente trabajo permite concluir que la laguna Barra de Navidad es un área de crianza para numerosas especies. Este resultado es de importancia para las pesquerías locales, las cuales explotan directamente los peces de la laguna o las fases adultas en las áreas costeras cercanas. En los planes de manejo pesquero que se implementen en el futuro, debe tenerse en cuenta que muchas especies de peces cumplen parte de su ciclo de vida en la laguna. Esto refuerza también la necesidad de realizar planes de conservación integrales de este ecosistema. Es importante resaltar que la definición de la laguna como zona de crianza esencial o hábitat efectivo de juveniles, depende de la proporción de individuos que aporta este cuerpo de agua a las poblaciones adultas, ya sea por unidad de área (Beck et al., 2001) o como porcentaje del total aportado por todos los hábitats de juveniles (Dahlgren et al., 2006). Esta información no está disponible actualmente y tendrá que ser objetivo de investigaciones posteriores.
Agradecimientos
Esta investigación fue financiada parcial-mente, mediante el proyecto 05-2010-1-746 del fondo COECYTJAL-UDG 2010 y el apoyo a Nuevos Profesores de Tiempo Completo del PROMEP (Oficio: 103.5/12/3418). Agradecemos a Albert Maurits Van der Heiden Joris, por su decisivo apoyo para la identificación de la especie Centropomus robalito. Los datos de lluvia y temperatura del aire fueron suministrados por el Servicio Meteorológico Nacional de México. Nuestro agradecimiento también a los estudiantes de la carrera de Licenciado en Biología Marina de la Universidad de Guadalajara que participaron como voluntarios en la investigación.
Referencias
Acevedo, A. (1997). Caracterización ecológica de la comunidad íctica de la laguna Ojo de Liebre, B.C.S. México. (Tesis inédita de maestría). CICIMAR, Instituto Politécnico Nacional, La Paz, B.C.S. [ Links ]
Álvarez-Rubio, M., Amezcua-Linares, F., & Yáñez-Arancibia, A. (1986). Ecología y estructura de la comunidad de peces en el sistema Lagunar Teacapán-Agua Brava. Nayarit, Pacífico central de México. Anales del Instituto de Ciencias del Mar y Limnología, Universidad Nacional Autónoma de México, 13, 185-242. [ Links ]
Amezcua-Linares, F. (1977). Generalidades ictiológicas del sistema lagunar costero de Huizache-Caimanero, Sinaloa, México. Anales del Instituto de Ciencias del Mar y Limnología, Universidad Nacional Autónoma de México, 4, 1-26. [ Links ]
Amezcua-Linares, F., Madrid, J., & Aguirre, H. (2006). Efecto de la pesca artesanal de camarón sobre la ictiofauna en el sistema lagunar de Santa María la Reforma, suroeste del Golfo de California. Ciencias Marinas, 32(1B), 97-109. [ Links ]
Amezcua-Linares, F. (2008). Peces demersales del Pacífico de México. México: Instituto de Ciencias del Mar y Limnología, Universidad Nacional Autónoma de México. [ Links ]
Anónimo. (1958). Symposium on the classification of brackish waters. Venice, April 8-14, 1958. Archives for Limnology and Oceanography, 11(suppl.), 1-248. [ Links ]
Arellano, M., Cruz-Agüero, J., & Cota, V. M. (1996). Lista sistemática de los peces marinos de las lagunas Ojo de Liebre y Guerrero Negro, BCS y BC, México. Ciencias Marinas, 22, 111-128. [ Links ]
Beck, M. W., Heck, K. L., Jr., Able, K. W., Childers, D. L., Eggleston, D. B., Gillanders, B. M., Halpern, B., Hays, C. G., Hoshino, K., Minello, T. J., Orth, R. J., Sheridan, P. F., & Weinstein, M. P. (2001). The identification, conservation, and management of estuarine and marine nurseries for fish and invertebrates. BioScience, 51(8), 633-641. [ Links ]
Benítez, C., Ruiz, J. M., Peña, E., Blanco, M., López, C., López, P., & Castañeda, A. (2007). Diversidad y abundancia de la comunidad de peces del estero “El Custodio”, Municipio de Compostela, Nayarit, México. REDVET, 8, 1-13. [ Links ]
Cabral-Solís, E. G., & Espino-Barr, E. (2004). Distribución y abundancia espacio-temporal de los peces en la laguna de Cuyutlán, Colima, México. Oceánides, 19, 19-27. [ Links ]
Contreras, F., & Warner, B. G. (2004). Ecosystem characteristics and management considerations for coastal wetlands in Mexico. Hydrobiologia, 511, 233-245. [ Links ]
Cruz-Agüero, J., & Cota, V. M. (1998). Ictiofauna de la laguna de San Ignacio, Baja California Sur, México: Nuevos registros y ampliaciones de ámbito. Ciencias Marinas, 24, 353-358. [ Links ]
Dahlgren, C. P., Kellison, G. T., Adams, A. J., Gillanders, B. M., Kendall, M. S., Layman, C. A., Ley, J. A., Nagelkerken, I., & Serafy, J. E. (2006). Marine nurseries and effective juvenile habitats: concepts and applications. Marine Ecology Progress Series, 312, 291-295. [ Links ]
Danemann, G., & Cruz-Agüero, J. (1993). Ictiofauna de laguna San Ignacio, Baja California Sur, México. Ciencias Marinas, 19(3), 333-341. [ Links ]
Díaz, S., Aguirre, A., & Cano, E. (2006). Evaluación de las comunidades de peces en dos sistemas lagunares estuarinos del sur de Chiapas, México. Hidrobiológica, 16(2), 197-210. [ Links ]
Eschmeyer, W. N. (Ed.). (2013). Catalog of Fishes. [Versión electrónica de California Academy of Sciences] Recuperado de: http://research.calacademy.org/ research/ichthyology/catalog/fishcatmain.asp
Links ]">
Fischer, W., Krupp, F., Schneider, W., Sommer, C., Carpenter, K. E., & Niem, V. (1995). Guía FAO para identificación de especies para los fines de la pesca Pacífico centro-oriental. Volumen III. Vertebrados, Parte 2, Roma, FAO. [ Links ]
Flores-Verdugo, F., González-Farías, F., Ramírez-Flores, O., Amezcua-Linares, F., Yáñez-Arancibia, A., Álvarez-Rubio, M., & Day, J. W., Jr. (1990). Mangrove ecology, aquatic primary productivity and fish community dynamics in the Teacapán-Agua Brava lagoon-estuarine system (Mexican Pacific). Estuaries, 13(2), 219-230. [ Links ]
Froese, R., & Pauly, D. (Eds.). (2013). FishBase. [World Wide Web electronic publication]. Recuperado de www.fishbase.org, versión (02/2013). [ Links ]
González, A. F., Cruz-Agüero, G., Cruz-Agüero, J., & Ruiz, G. (2005). Seasonal pattern of the fish assemblage of El Conchalito mangrove swamp, La Paz bay, Baja California Sur, Mexico. Hidrobiológica, 15(2 especial), 205-214. [ Links ]
Grijalva, J. M., Núñez-Quevedo, S., & Castro-Longoria, R. (1996). Ictiofauna de la laguna costera La Cruz, Sonora, México. Ciencias Marinas, 22(2), 129-150. [ Links ]
Hernández, S., Serrano, S., Hernández, X. A., & Robles, M. I. (2012). Variación temporal y espacial de aves playeras en la laguna Barra de Navidad, Jalisco, en tres temporadas no reproductivas. Revista de Biología Tropical, 60(3), 1317-1326. [ Links ]
Lankford, R. R. (1976). Coastal lagoons of Mexico: their origin and classification. In Wiley, M. (Ed.), Estuarine Processes. Vol. II (182-215). New York: Academic Press. [ Links ]
Mann, K. H., & Lazier, J. R. N. (1996). Dynamics of marine ecosystems: biological–physical interactions in the oceans. Oxford: Blackwell, 2nd Ed. [ Links ]
Méndez-Linares, A. P., López-Portillo, J., Hernández-Santana, J. R., Ortiz Pérez, M. A., & Oropeza-Orozco, O. (2007). The mangrove communities in the Arroyo Seco deltaic fan, Jalisco, Mexico, and their relation with the geomorphic and physical–geographic zonation. Catena, 70, 127-142. doi:10.1016/j. catena.2006.05.010
Links ]">
Mendoza, E., Castillo-Rivera, M., Zárate-Hernández, R., & Ortiz-Burgos, S. (2009). Seasonal variations in the diversity, abundance, and composition of species in an estuarine fish community in the tropical eastern Pacific, Mexico. Ichthyological Research, 56, 330-339. doi: 10.1007/s10228-009-0102-5
Links ]">
Meyer, A., Velázquez-González, O., & Patiño-Barragán, M. (2006). Ciclo anual de las variables hidrobiológicas en el estuario Barra de Navidad, México. In M. C. Jiménez-Quiróz, & E. Espino-Barr (Eds.). Los recursos pesqueros y acuícolas de Jalisco, Colima y Michoacán (81-100). Sagarpa /Instituto Nacional de la Pesca. [ Links ]
Mumby, P. J., Edwards, A. J., Arias-Gonzalez, J. E., Lin-deman, K. C., Blackwell, P. G., Gall, A., Gorczynska, M. I., Harborne, A. R., Pescod, C. L., Renken, H., Wabnitz, C. C. C., & Llewellyn, G. (2004). Mangroves enhance the biomass of coral reef fish communities in the Caribbean. Nature, 427, 533-536. doi:10.1038/nature02286. [ Links ]
Nagelkerken, I. (2009). Evaluation of nursery function of mangroves and seagrass beds for tropical decapods and reef fishes: Patterns and underlying mechanisms. In I. Nagelkerken (Ed.), Ecological Connectivity among Tropical Coastal Ecosystems (357-399). Dordrecht: Springer Science+Business Media B.V. [ Links ]
Nelson, J. S. (2006). Fishes of the world. New Jersey: Wiley, 4ta Ed. [ Links ]
Parsons, T. R., Maita, Y., & Lalli, C. M. (1984). A manual of chemical and biological methods for seawater analysis. Oxford: Pergamon. [ Links ]
Raz-Guzman, A., & Huidobro, L. (2002). Fish communities in two environmentally different estuarine systems of Mexico. Journal of Fish Biology, 61(supplement A), 182-195. doi:10.1006/jfbi.2002.2076
Links ]">
Rodríguez-Romero, J., López-González, L. del C., Galván-Magaña, F., Sánchez-Gutiérrez, F. J., Inohuye-Rivera, R. B., & Pérez-Urbiola, J. C. (2011). Seasonal chan-ges in a fish assemblage associated with mangroves in a coastal lagoon of Baja California Sur, Mexico. Latin American Journal of Aquatic Research, 39(2), 250-260. doi: 10.3856/vol39-issue2-fulltext-6
Links ]">
Rosales, J. A. (2004). Composición, importancia y movimiento de los peces de bahía de San Quintín, Baja California, México. Ciencias Marinas, 30(1A), 109-117. [ Links ]
Sandoval, L., Flores, F., Zaragoza, U., Day, J. W., & Estrada, A. (1988). Phytoplankton productivity in the Barra de Navidad coastal Lagoon on the Pacific coast of Mexico. Revue d’hydrobiologie tropicale, 21(2), 101-108. [ Links ]
Siegel, S., & Castellan, N. J. (1988). Nonparametric statistics for the behavioral sciences. New York: McGraw-Hill. [ Links ]
Silva-Bátiz, F. de A., González-Sansón, G., & Godínez-Domínguez, E. (2010). Los humedales costeros de Jalisco: Bases conceptuales para su conservación y uso racional. In E. Godínez-Domínguez, C. Franco-Gordo, J. A. Rojo-Vázquez, F. de A. Silva-Bátiz, & G. González-Sansón (Eds.), Ecosistemas marinos de la costa sur de Jalisco y Colima (213-248). Autlán de Navarro: Universidad de Guadalajara. [ Links ]
Silva-Bátiz, F. de A., González-Sansón, G., Nené, A., Godínez, E., Franco, M. del C., Corgos, A., Hernández, S., Hinojosa, J. A., Galván, V. H., & Rojo, J. (2012). Bases para el manejo y conservación de la laguna de Barra de Navidad, Jalisco. Guadalajara: Editorial Página Seis. [ Links ]
StatSoft (2006). STATISTICA (data analysis software system) version 7.1. www.statsoft.com
Links ]">
Tapia-García, M., Suárez-Núñez, C., Cerdenares, G., Macuitl-Montes, M. C., & García-Abad, M. C. (1998). Composición y distribución de la ictiofauna en la Laguna del Mar Muerto, Pacífico mexicano. Revista de Biología Tropical, 46(2), 277-284. [ Links ]
Yáñez-Arancibia, A. (1978). Taxonomía, ecología, y estructura de las comunidades de peces en lagunas costeras con bocas efímeras del Pacífico de México. Publicaciones Especiales del Instituto de Ciencias del Mar y Limnología, Universidad Nacional Autónoma de México, 2, 1-306. [ Links ]
Zar, J. H. (2010). Biostatistical Analysis. New Jersey: Prentice-Hall. [ Links ]
Álvarez-Rubio, M., Amezcua-Linares, F., & Yáñez-Arancibia, A. (1986). Ecología y estructura de la comunidad de peces en el sistema Lagunar Teacapán-Agua Brava. Nayarit, Pacífico central de México. Anales del Instituto de Ciencias del Mar y Limnología, Universidad Nacional Autónoma de México, 13, 185-242. [ Links ]
Amezcua-Linares, F. (1977). Generalidades ictiológicas del sistema lagunar costero de Huizache-Caimanero, Sinaloa, México. Anales del Instituto de Ciencias del Mar y Limnología, Universidad Nacional Autónoma de México, 4, 1-26. [ Links ]
Amezcua-Linares, F., Madrid, J., & Aguirre, H. (2006). Efecto de la pesca artesanal de camarón sobre la ictiofauna en el sistema lagunar de Santa María la Reforma, suroeste del Golfo de California. Ciencias Marinas, 32(1B), 97-109. [ Links ]
Amezcua-Linares, F. (2008). Peces demersales del Pacífico de México. México: Instituto de Ciencias del Mar y Limnología, Universidad Nacional Autónoma de México. [ Links ]
Anónimo. (1958). Symposium on the classification of brackish waters. Venice, April 8-14, 1958. Archives for Limnology and Oceanography, 11(suppl.), 1-248. [ Links ]
Arellano, M., Cruz-Agüero, J., & Cota, V. M. (1996). Lista sistemática de los peces marinos de las lagunas Ojo de Liebre y Guerrero Negro, BCS y BC, México. Ciencias Marinas, 22, 111-128. [ Links ]
Beck, M. W., Heck, K. L., Jr., Able, K. W., Childers, D. L., Eggleston, D. B., Gillanders, B. M., Halpern, B., Hays, C. G., Hoshino, K., Minello, T. J., Orth, R. J., Sheridan, P. F., & Weinstein, M. P. (2001). The identification, conservation, and management of estuarine and marine nurseries for fish and invertebrates. BioScience, 51(8), 633-641. [ Links ]
Benítez, C., Ruiz, J. M., Peña, E., Blanco, M., López, C., López, P., & Castañeda, A. (2007). Diversidad y abundancia de la comunidad de peces del estero “El Custodio”, Municipio de Compostela, Nayarit, México. REDVET, 8, 1-13. [ Links ]
Cabral-Solís, E. G., & Espino-Barr, E. (2004). Distribución y abundancia espacio-temporal de los peces en la laguna de Cuyutlán, Colima, México. Oceánides, 19, 19-27. [ Links ]
Contreras, F., & Warner, B. G. (2004). Ecosystem characteristics and management considerations for coastal wetlands in Mexico. Hydrobiologia, 511, 233-245. [ Links ]
Cruz-Agüero, J., & Cota, V. M. (1998). Ictiofauna de la laguna de San Ignacio, Baja California Sur, México: Nuevos registros y ampliaciones de ámbito. Ciencias Marinas, 24, 353-358. [ Links ]
Dahlgren, C. P., Kellison, G. T., Adams, A. J., Gillanders, B. M., Kendall, M. S., Layman, C. A., Ley, J. A., Nagelkerken, I., & Serafy, J. E. (2006). Marine nurseries and effective juvenile habitats: concepts and applications. Marine Ecology Progress Series, 312, 291-295. [ Links ]
Danemann, G., & Cruz-Agüero, J. (1993). Ictiofauna de laguna San Ignacio, Baja California Sur, México. Ciencias Marinas, 19(3), 333-341. [ Links ]
Díaz, S., Aguirre, A., & Cano, E. (2006). Evaluación de las comunidades de peces en dos sistemas lagunares estuarinos del sur de Chiapas, México. Hidrobiológica, 16(2), 197-210. [ Links ]
Eschmeyer, W. N. (Ed.). (2013). Catalog of Fishes. [Versión electrónica de California Academy of Sciences] Recuperado de: http://research.calacademy.org/ research/ichthyology/catalog/fishcatmain.asp
Links ]">
Fischer, W., Krupp, F., Schneider, W., Sommer, C., Carpenter, K. E., & Niem, V. (1995). Guía FAO para identificación de especies para los fines de la pesca Pacífico centro-oriental. Volumen III. Vertebrados, Parte 2, Roma, FAO. [ Links ]
Flores-Verdugo, F., González-Farías, F., Ramírez-Flores, O., Amezcua-Linares, F., Yáñez-Arancibia, A., Álvarez-Rubio, M., & Day, J. W., Jr. (1990). Mangrove ecology, aquatic primary productivity and fish community dynamics in the Teacapán-Agua Brava lagoon-estuarine system (Mexican Pacific). Estuaries, 13(2), 219-230. [ Links ]
Froese, R., & Pauly, D. (Eds.). (2013). FishBase. [World Wide Web electronic publication]. Recuperado de www.fishbase.org, versión (02/2013). [ Links ]
González, A. F., Cruz-Agüero, G., Cruz-Agüero, J., & Ruiz, G. (2005). Seasonal pattern of the fish assemblage of El Conchalito mangrove swamp, La Paz bay, Baja California Sur, Mexico. Hidrobiológica, 15(2 especial), 205-214. [ Links ]
Grijalva, J. M., Núñez-Quevedo, S., & Castro-Longoria, R. (1996). Ictiofauna de la laguna costera La Cruz, Sonora, México. Ciencias Marinas, 22(2), 129-150. [ Links ]
Hernández, S., Serrano, S., Hernández, X. A., & Robles, M. I. (2012). Variación temporal y espacial de aves playeras en la laguna Barra de Navidad, Jalisco, en tres temporadas no reproductivas. Revista de Biología Tropical, 60(3), 1317-1326. [ Links ]
Lankford, R. R. (1976). Coastal lagoons of Mexico: their origin and classification. In Wiley, M. (Ed.), Estuarine Processes. Vol. II (182-215). New York: Academic Press. [ Links ]
Mann, K. H., & Lazier, J. R. N. (1996). Dynamics of marine ecosystems: biological–physical interactions in the oceans. Oxford: Blackwell, 2nd Ed. [ Links ]
Méndez-Linares, A. P., López-Portillo, J., Hernández-Santana, J. R., Ortiz Pérez, M. A., & Oropeza-Orozco, O. (2007). The mangrove communities in the Arroyo Seco deltaic fan, Jalisco, Mexico, and their relation with the geomorphic and physical–geographic zonation. Catena, 70, 127-142. doi:10.1016/j. catena.2006.05.010
Links ]">
Mendoza, E., Castillo-Rivera, M., Zárate-Hernández, R., & Ortiz-Burgos, S. (2009). Seasonal variations in the diversity, abundance, and composition of species in an estuarine fish community in the tropical eastern Pacific, Mexico. Ichthyological Research, 56, 330-339. doi: 10.1007/s10228-009-0102-5
Links ]">
Meyer, A., Velázquez-González, O., & Patiño-Barragán, M. (2006). Ciclo anual de las variables hidrobiológicas en el estuario Barra de Navidad, México. In M. C. Jiménez-Quiróz, & E. Espino-Barr (Eds.). Los recursos pesqueros y acuícolas de Jalisco, Colima y Michoacán (81-100). Sagarpa /Instituto Nacional de la Pesca. [ Links ]
Mumby, P. J., Edwards, A. J., Arias-Gonzalez, J. E., Lin-deman, K. C., Blackwell, P. G., Gall, A., Gorczynska, M. I., Harborne, A. R., Pescod, C. L., Renken, H., Wabnitz, C. C. C., & Llewellyn, G. (2004). Mangroves enhance the biomass of coral reef fish communities in the Caribbean. Nature, 427, 533-536. doi:10.1038/nature02286. [ Links ]
Nagelkerken, I. (2009). Evaluation of nursery function of mangroves and seagrass beds for tropical decapods and reef fishes: Patterns and underlying mechanisms. In I. Nagelkerken (Ed.), Ecological Connectivity among Tropical Coastal Ecosystems (357-399). Dordrecht: Springer Science+Business Media B.V. [ Links ]
Nelson, J. S. (2006). Fishes of the world. New Jersey: Wiley, 4ta Ed. [ Links ]
Parsons, T. R., Maita, Y., & Lalli, C. M. (1984). A manual of chemical and biological methods for seawater analysis. Oxford: Pergamon. [ Links ]
Raz-Guzman, A., & Huidobro, L. (2002). Fish communities in two environmentally different estuarine systems of Mexico. Journal of Fish Biology, 61(supplement A), 182-195. doi:10.1006/jfbi.2002.2076
Links ]">
Rodríguez-Romero, J., López-González, L. del C., Galván-Magaña, F., Sánchez-Gutiérrez, F. J., Inohuye-Rivera, R. B., & Pérez-Urbiola, J. C. (2011). Seasonal chan-ges in a fish assemblage associated with mangroves in a coastal lagoon of Baja California Sur, Mexico. Latin American Journal of Aquatic Research, 39(2), 250-260. doi: 10.3856/vol39-issue2-fulltext-6
Links ]">
Rosales, J. A. (2004). Composición, importancia y movimiento de los peces de bahía de San Quintín, Baja California, México. Ciencias Marinas, 30(1A), 109-117. [ Links ]
Sandoval, L., Flores, F., Zaragoza, U., Day, J. W., & Estrada, A. (1988). Phytoplankton productivity in the Barra de Navidad coastal Lagoon on the Pacific coast of Mexico. Revue d’hydrobiologie tropicale, 21(2), 101-108. [ Links ]
Siegel, S., & Castellan, N. J. (1988). Nonparametric statistics for the behavioral sciences. New York: McGraw-Hill. [ Links ]
Silva-Bátiz, F. de A., González-Sansón, G., & Godínez-Domínguez, E. (2010). Los humedales costeros de Jalisco: Bases conceptuales para su conservación y uso racional. In E. Godínez-Domínguez, C. Franco-Gordo, J. A. Rojo-Vázquez, F. de A. Silva-Bátiz, & G. González-Sansón (Eds.), Ecosistemas marinos de la costa sur de Jalisco y Colima (213-248). Autlán de Navarro: Universidad de Guadalajara. [ Links ]
Silva-Bátiz, F. de A., González-Sansón, G., Nené, A., Godínez, E., Franco, M. del C., Corgos, A., Hernández, S., Hinojosa, J. A., Galván, V. H., & Rojo, J. (2012). Bases para el manejo y conservación de la laguna de Barra de Navidad, Jalisco. Guadalajara: Editorial Página Seis. [ Links ]
StatSoft (2006). STATISTICA (data analysis software system) version 7.1. www.statsoft.com
Links ]">
Tapia-García, M., Suárez-Núñez, C., Cerdenares, G., Macuitl-Montes, M. C., & García-Abad, M. C. (1998). Composición y distribución de la ictiofauna en la Laguna del Mar Muerto, Pacífico mexicano. Revista de Biología Tropical, 46(2), 277-284. [ Links ]
Yáñez-Arancibia, A. (1978). Taxonomía, ecología, y estructura de las comunidades de peces en lagunas costeras con bocas efímeras del Pacífico de México. Publicaciones Especiales del Instituto de Ciencias del Mar y Limnología, Universidad Nacional Autónoma de México, 2, 1-306. [ Links ]
Zar, J. H. (2010). Biostatistical Analysis. New Jersey: Prentice-Hall. [ Links ]
*Correspondencia: Gaspar González-Sansón: Departamento de Estudios para el Desarrollo Sustentable de la Zona Costera, Gómez Farías 82, San Patricio-Melaque, Cihuatlán, Jalisco, CP 48980; gaspargonzalez2001@yahoo.es
Consuelo Aguilar-Betancourt: Departamento de Estudios para el Desarrollo Sustentable de la Zona Costera, Gómez Farías 82, San Patricio-Melaque, Cihuatlán, Jalisco, CP 48980; coquiaguilar06@yahoo.es
Daniel Kosonoy-Aceves: Departamento de Estudios para el Desarrollo Sustentable de la Zona Costera, Gómez Farías 82, San Patricio-Melaque, Cihuatlán, Jalisco, CP 48980; dkosonoy@yahoo.com.mx
Gabriela Lucano-Ramírez: Departamento de Estudios para el Desarrollo Sustentable de la Zona Costera, Gómez Farías 82, San Patricio-Melaque, Cihuatlán, Jalisco, CP 48980; lucanorg@costera.melaque.mx
Salvador Ruiz-Ramírez: Departamento de Estudios para el Desarrollo Sustentable de la Zona Costera, Gómez Farías 82, San Patricio-Melaque, Cihuatlán, Jalisco, CP 48980; sruizram@costera.melaque.mx
Juan Ramón Flores-Ortega: Departamento de Estudios para el Desarrollo Sustentable de la Zona Costera, Gómez Farías 82, San Patricio-Melaque, Cihuatlán, Jalisco, CP 48980;
Ángel Hinojosa-Larios: Departamento de Estudios para el Desarrollo Sustentable de la Zona Costera, Gómez Farías 82, San Patricio-Melaque, Cihuatlán, Jalisco, CP 48980; hinojosalarios@gmail.com
Francisco de Asís Silva-Bátiz: Departamento de Estudios para el Desarrollo Sustentable de la Zona Costera, Gómez Farías 82, San Patricio-Melaque, Cihuatlán, Jalisco, CP 48980; fasilva@costera.melaque.mx
1. Departamento de Estudios para el Desarrollo Sustentable de la Zona Costera, Gómez Farías 82, San Patricio-Melaque, Cihuatlán, Jalisco, CP 48980; gaspargonzalez2001@yahoo.es, coquiaguilar06@yahoo.es, dkosonoy@yahoo.com.mx, lucanorg@costera.melaque.mx, sruizram@costera.melaque.mx, hinojosalarios@gmail.com, fasilva@costera.melaque.mx
Recibido 05-vi-2013. Corregido 13-IX-2013. Aceptado 15-X-2013.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}