










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.62 n.1 San José Jan./Mar. 2014
Reproductive strategies of two Neotropical killifish, Austrolebias vandenbergi and Neofundulus ornatipinnis (Cyprinodontiformes: Rivulidae) in the Bolivian Gran Chaco
Estrategias reproductivas de dos especies de rivulidos Neotropicales Austrolebias vandenbergi y Neofundulus ornatipinnis (Cyprinodontiformes: Rivulidae) en el Gran Chaco Boliviano
Estrategias reproductivas de dos especies de rivulidos Neotropicales Austrolebias vandenbergi y Neofundulus ornatipinnis (Cyprinodontiformes: Rivulidae) en el Gran Chaco Boliviano
*Dirección para correspondencia:
Abstract
The dry Chaco, a semiarid thorn forest, is experiencing some of the highest deforestation rates globally, coupled with the fact that small-bodied fish are at the highest risk of extinction, the killifish inhabiting this region may be some of the most threatened taxa. Yet, aspects of ecology and life history for Neotropical killifishes in the Bolivian Gran Chaco region are completely lacking, and basic life-history data is of critical importance for the design and implementation of conservation measures. Collections were conducted during the early (January 2011) and late (March-April 2011) rainy season using an area-based sampler and dip net surveys. Fish standard length and body depth were measured as well as the number of oocytes per size class, mean oocyte diameter per size class, and total fecundity for the females of each species. A total of 490 specimens of rivulids were captured; Austrolebias vandenbergi: 85 females, 105 males and 39 juveniles, and N. ornatipinnis: 62 females, 113 males, 86 juveniles. Sexual size dimorphism, absolute fecundity, oocyte developmental stages, oocyte diameter, and population sex ratios were determined for each species. Both species exhibited sexual size dimorphism. Male A. vandenbergi exhibited longer standard length (mean±SD; males: 27.07±3.89mm, females: 23.6±2.02mm) and body depth (males: 8.9±1.7mm, females: 7.2±1.1mm) as compared to females. Male N. ornatipinnis had a similar pattern for both standard length (males: 26.0±7.1mm, females: 19.1±5.83mm) and body depth (males: 5.6±1.9mm, females: 4.7±1.0mm). Austrolebias vandenbergi had fewer and smaller oocytes per female (47±31.6) than N. ornatipinnis (206±131.2). There was a positive relationship between fecundity and female body size in both species. The presence of multiple developmental stages of oocytes (immature, maturing, and mature) suggest that both species of rivulids exhibit fractional spawning, a reproductive strategy that enhances reproductive success in these extreme habitats. Neofundulus ornatipinnis exhibited a higher mean oocyte diameter for all three developmental stages (immature, maturing, and mature) as compared to A. vandenbergi. Austrolebias vandenbergi exhibited an equal adult sex ratio (males:females, 1:1), but there was a slight female biased ratio for N. ornatipinnis (males:females, 1:1.8). These results provide fundamental and valuable information for ensuring rivulid conservation in tropical regions, and also improve the knowledge on the biology and ecology of these poorly known species. To our knowledge, this study represents the first contribution on the reproductive biology of two Neotropical annual rivulid fishes (Austrolebias vandenbergi and Neofundulus ornatipinnis) inhabiting semi-permanent and ephemeral ponds in the Gran Chaco of Southeastern Bolivia.
Key words: annual fish, ephemeral ponds, fecundity, life history, sexual dimorphism, tropical dry forest.
Resumen
Información sobre la ecología e historia de vida de varias especies de rivulidos no ha sido estudiada, por lo tanto se analiza la biología reproductiva de dos especies neotropicales (Austrolebias vandenbergi y Neofundulus ornatipinnis) que habitan pozos semipermanentes y temporales en el Gran Chaco Boliviano. Las recolectas se realizaron al inicio y al final de la estación lluviosa. Dimorfismo sexual, fecundidad absoluta, diámetro de los ovocitos y proporción sexual fue determinada para cada especie de rivulido. Machos en las dos especies presentaron mayor tamaño de longitud estándar y altura del cuerpo cuando fue comparado con las hembras. Austrolebias vandenbergi tuvo menor fecundidad por hembra y huevos más pequeños que N. ornatipinnis, pero hembras en ambas especies presentaron una relación estrecha y positiva entre fecundidad y tamaño cuerpo. La presencia de varios estadios gonadales sugiere un desove fraccionado, lo cual es una estrategia reproductiva que promueve la viabilidad y sobrevivencia de estos peces en hábitats de extremas condiciones. La proporción sexual no varió entre machos y hembras de A. vandenbergi (1:1), pero en N. ornatipinnis la proporción sexual fue dominada por las hembras (1:1.8). Este estudio representa uno de los primeros que investiga aspectos de la biología reproductiva de A. vandenbergi y N. ornatipinnis, lo cual es importante para la conservación de rivulidos en regiones tropicales.
Palabras clave: bosque seco tropical, dimorfismo sexual, fecundidad, historia de vida, peces anuales, pozos efímeros.
The family Rivulidae (Cyprinodonti-formes) is a species-rich group of freshwater fishes with more than 350 valid species that extend from Southern Florida to Northeastern Argentina (Costa, 2011). Many of these species are able to complete their life cycles in temporary aquatic habitats, and thus are often referred to as annual killifishes. These species possess a number of adaptations to deal with the constraints of ephemeral habitats. Populations can persist in these habitats because their eggs are deposited in the substrate of the pond where they undergo diapause until the next rainy season (Wourms, 1972). Annual rivulids possess an opportunistic life-history strategy, characterized by having a small body and undergoing rapid maturation (Winemiller & Rose, 1992). Also characteristic of species possessing an opportunistic life-history strategy is sexual selection (Winemiller, 1992). Rivulids are often sexually dimorphic; males are brightly colored and conduct elaborated courtship displays (Belote & Costa, 2004; Garcia, Loureiro & Tassino, 2008).
Globally, small-bodied freshwater fish are amongst the highest fish taxa at risk of extinction, which has been attributed to their vulnerability to habitat alterations and degradation (Olden, Hogan & Zanden, 2007). Killifish are no exception; they often possess a limited geo-graphical range and are amongst some of the most threatened fish in Brazil (Rosa & Lima, 2008; Volcan, Lanés & Cheffe, 2010; Costa, 2012). Understanding life-history traits that are fundamental to population regulation, like a species reproductive biology, are essential in the design and implementation of conservation strategies (Winemiller, 2005).
While the phylogeny of rivulids has been the subject of much study (Costa, 1988, 1998, 2002, 2006), information on the ecology and life-history on a number of these species is lacking. The genus Neofundulus is comprised of five species, but two of the five species of Neofundulus are only known from their holo-types (Costa, 1998). There are 39 valid species recognized belonging to the genus Austrolebias (Costa, 2006; Loureiro, Duarte & Zarucki, 2011), and aspects of the ecology, behavior, and reproduction of some species have been the subject of study (Belote & Costa 2004; Calviño, 2005; Garcia et al., 2008; Volcan, Fonseca & Robaldo, 2011). In this study, we investigated the reproductive biology of two species of killifish that were recently found in the Bolivian Gran Chaco; Van den Berg’s pearlfish ( Austrolebias vandenbergi; Huber, 1995) and the ornate killifish (Neofundulus ornatipinnis; Myers, 1935) (Montaña, Schalk, & Taphorn, 2012). Preliminary details on the reproductive biology and diet of A. vandenbergi from this site have been reported (Montaña et al., 2012), but nothing is known about N. ornatipinnis in the region. Here we present a more extensive analysis on the reproductive biology of both species of killifish from the Bolivian Gran Chaco.
Material and methods
Study area: The Gran Chaco is a large xeric floodplain that contains vast expanses of semiarid thorn forest and palm savannahs distributed across the countries of Argentina, Brazil, Bolivia, and Paraguay. The Chaco has a warm, wet summer (November-March) and a cool, dry winter (April-October). In the study area in the Bolivian Chaco, average rainfall and temperature are 513mm and 24.6ºC, respectively (Navarro & Maldonado, 2002). The surrounding forest is xerophytic thorn forest, with Schinopsis lorentzii (Engler, 1880) and Aspidosperma quebracho-blanco (Schlectendal, 1861) being common tree species and cacti (e.g. Opuntia sp., Cleistocactus baumannii [Lemaire, 1861], and Eriocereus guelichii [A. Berger, 1929]) and bromeliads constituting the common understory plants (Navarro & Maldonado, 2002). We sampled fish from both semi-permanent (N=1) and temporary (N=3) ponds around the Isoceño community of Kuaridenda, Provincia Cordillera, Departamento Santa Cruz, Bolivia (19°10’24’’ S -62°35’16’’ W and 19°10’34’’S - 62°35’21’’ W).
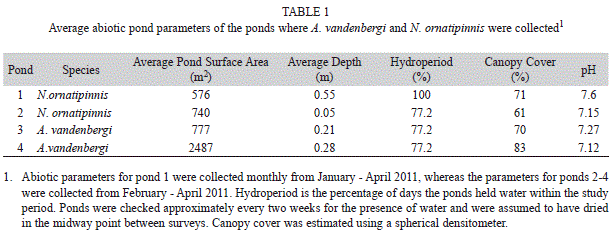
Fish surveys: Austrolebias vandenbergi were collected only at the end of the rainy season (April 2011), whereas individuals of N. ornatipinnis were collected towards the beginning (January 2011) and the end (March-April 2011) of rainy season using a combination of methods including an area-based sampler (area: 1.3m2; height: 0.70m) that was cleared with a dipnet (mesh: 2mm) and dipnet surveys. In the instance when the depth of the pond exceeded the height of the area-based sampler, we conducted five seine net sweeps (1m length x 1m height, 0.5mm mesh) (Werner, Skelly, Relyea & Yurewicz, 2007). The pipe-sampler was placed every three meters along a pond’s longest axis in ponds <100m in length or every ten meters in ponds >100m in length. The abiotic characteristics of the ponds surveyed in this study are listed in Table 1. All individuals captured were immediately fixed in 10% formalin and later sorted and stored in 70% ethanol. Specimens collected and analyzed were deposited in the Biodiversity Research and Teaching Collections (TCWC 15182.01, 15183.01, 15184.01) at Texas A&M University (TAMU), College Station Texas, U.S.A. Additional specimens not examined in this study were deposited in the Museo Noel Kempff Mercado in Santa Cruz de la Sierra, Bolivia.
Fish measurements and reproductive analysis: Specimens were identified to species level using taxonomic keys (Costa, 1988; Huber, 1995a-b; Costa, 2006) and measured with a dial caliper (to nearest 0.01mm). Morphological measurements including standard length (SL) and body depth (BD) were measured for each individual. Standard length was used to classify the specimens into size groups. Specimens were identified as juveniles, males and females, and female gonadal state of maturation was determined following the methodology by Vazzoler (1996), which allows the inference of the degree of development of the ovaries. For each female ovary, oocytes were separated and classified into three different levels of maturity using a dissecting microscope: immature (homogeneous and light colored oocytes and without follicle differentiation), maturing (oocytes in intermediate stages with yellow coloration and follicle in different stages), and mature (oocytes in final stage of maturation, totally developed ovaries with large follicle yellow to orange color). Fifteen oocytes were randomly chosen from each ovary and measured with an ocular micrometer in a dissecting microscope to estimate oocyte diameter (Gonçalves, Souza & Volcan, 2011). Absolute fecundity was estimated by counting the total eggs in the ovary of each female (Vazzoler, 1996). Finally, sex ratio was determined using the absolute frequencies of males and females collected during the sampling period. The Gonadosomatic Index (GSI) was calculated only for female N. ornatipinnis, for which there were samples collected during the early rainy season (only three mature females were examined because the majority of individuals were juveniles), March (11 females examined), end of rainy season (48 females examined). The GSI (GSI = Gw = weight of the gonad /Wt = weight of the individual * 100) allows inferences to be made about species spawning periods. For example, an increase in the GSI suggests an approach to spawning season (Vazzoler, 1996).
The relationship between fecundity (F) and standard length was estimated for females of each species using the ordinary least squares regression model given by F = a+b (x), where F represents fecundity (dependent variable), x represents length (independent variable), a is the numerical variable of intercept, and b is the slope. Length classes were determined to verify size differences between sexes (Sparre & Venema, 1997). Due to the variation in individual size classes and non-normality of the data, we used the Kolmogorov-Smirnov test (K-S), a non-parametric t-test, to test for differences in the standard lengths and body depths between species of killifish. A one-way analysis of variance (ANOVA) was performed to compare the means of the oocyte diameter among size classes between both species of killifish. Finally, chi-square (χ2, p<0.05) was performed in the sexes of each species to determine whether or not they differ from the expectation (1:1). All analyses were conducted using the software JMP 7 (SAS Institute Inc. 2007).
Results
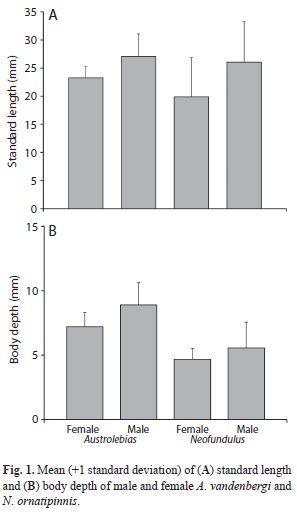
A total of 490 specimens of rivulids were captured. Of those, 229 were A. vandenbergi (85 females, 105 males and 39 juveniles) and 261 were N. ornatipinnis (62 females, 113 males, 86 juveniles). Austrolebias vandenbergi were only collected from the two temporary ponds, whereas N. ornatipinnis were collected from both a temporary and the semi-permanent pond in the area. These species did not co-occur with one another (Table 1). Standard length varied between 19.0 and 24.9mm (23.6 mean±2.02 standard deviation) for females and between 18.3 and 28.8mm (27.07±3.8) for males (Fig. 1A). Body depth was greater for males (8.9±1.7) than for females (7.2±1.1) (Fig. 1B). Standard length in N. ornatipinnis ranged from 10.6 to 34.7mm (19.1±5.8) for females and from 15.6 to 50.6mm (26.0±7.1) for males (Fig. 1A). Body depth was also greater for males (5.6±1.9) than for females (4.7±1.0) (Fig. 1B). The K-S test indicated significant differences in the standard length of males and females of A. vandenbergi (D=0.51, p<0.001) and N. ornatipinnis (D=0.66, p<0.009), with males showing larger sizes. Significant differences also were found in the body depth of males and females of A. vandenbergi (D=0.60, p<0.008) and N. ornatipinnis (D=0.33, p<0.0006), respectively.
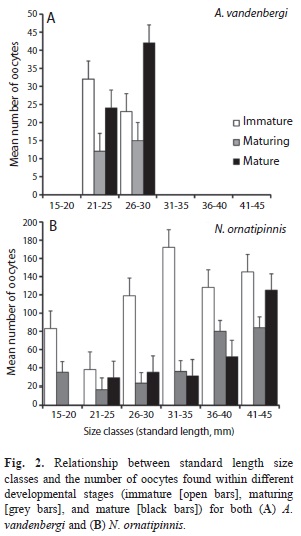
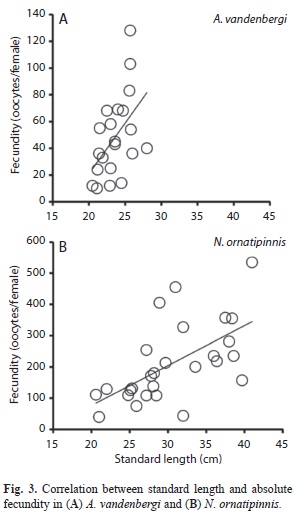
There were 85 ovaries of A. vandenbergi and 62 ovaries of N. ornatipinnis examined to determine absolute fecundity and ovarian developmental stages. The mean fecundity of A. vandenbergi (mean 47 oocytes) was lower than N. ornatipinnis (mean 206 oocytes) (Table 2). Examination of oocyte diameter revealed that both species of rivulids demonstrated fractional spawning, as determined by the presence of at least three oocyte developmental stages (Fig. 2, Table 2). Neofundulus ornatipinnis exhibited a higher mean oocyte diameter for all three developmental stages (immature, maturing, and mature) as compared to A. vandenbergi (Table 2). The ANOVA revealed significant differences among oocytes diameter size for each development stage in both species, A. vandenbergi (df=82, F=61.7, p<0.02) and N. ornatipinnis (df=59, F=29.9, p<0.002). Austrolebias vandenbergi exhibited an equal adult sex ratio (male:female 1:1; χ2=45.7, p=0.92), and there was a slight female bias was observed for N. ornatipinnis (1:1.8), but it was not statistically significant (χ2=30.0, p=0.20). The relationship between fecundity and standard length is represented by the equations: y=-130.6 + 7.6x (r=0.48) for A. vandenbergi and y=-208.3 + 13.6x (r=0.62) for N. ornatipinnis. There was a significant trend for increasing fecundity with increasing standard length in females for both A. vandenbergi (R2=0.24, p<0.0001) and N. ornatipinnis (R2=0.39, p<0.0008) (Fig. 3). The GSI estimates in females of N. ornatipinnis did not exhibit significant differences between early (January, GSI=7.24±0.42) and later (April, GIS=7.55±1.37) rainy season samples. However, there was a slight peak in March (GIS=8.08±1.44).
Discussion
In the four ponds surveyed, A. vandenbergi and N. ornatipinnis did not co-occur together. A.vandenbergi were captured only in ephemeral ponds, whereas N. ornatipinnis were collected in both an ephemeral and a semi-permanent pond. All ponds had high canopy cover with a habitat structure in the form of leaf litter input and woody debris. Structural heterogeneous substrates in these ephemeral ponds may offer shelter to these annual fishes or egg deposition sites.
Most rivulids are sexually dimorphic, with males exhibiting bright coloration and possessing relatively larger body sizes than females (Garcia et al., 2008; Gonçalves et al., 2011), and we observed a similar pattern in both species in this study. Overall, A. vandenbergi had shorter standard length but greater body depth than N. ornatipinnis, which reached a maximum standard length of 50.6mm. Males of both species exhibited a larger body size than females in regards to both standard length and body depth. This trend has been observed in other species of tropical annual rivulids including Cynopoecilus melanotaenia (Regan, 1912) and several species of Austrolebias (Laufer, Arim, Loureiro, Piñeiro-Guerra, Clavijo-Baquet & Fagúndez, 2009).
These differences in body size play an important role in sexual selection (Garcia et al., 2008). In fish, body size of females is favored by increased fecundity, whereas in males, size is affected by sexual selection. Female mating preferences for large males can be attributed to superior genes inherited by offspring or a variety of direct benefits including increased sperm quality and better paternal care (Shine, 1989).
In this study, the A. vandengerbi had an equal adult sex ratio (i.e., 1:1), but there was a slight non significant female biased ratio for N. ornatipinnis. These values were similar to Laufer et al. (2009) who found a ratio of 1:1 (males:females) for Austrolebias spp. and C. melanotaenia in ephemeral ponds in Uruguay; but differed from Gonçalves et al. (2011) whom observed a ratio of 2:1 for C. melanotaenia in ephemeral ponds Southern Brazil, and from Calviño (2005) who reported a sex ratio of 1:4 for N. ornatipinnis from Argentina. Sex ratio data provide important information to understand the population dynamics in fish, but whether these sex ratios may be caused by physiological and/or environmental factors that affect sexes (Vazzoler, 1996) still remains unknown at this time.
According to Winemiller & Rose (1992), the opportunistic strategy of annual rivulids which is characterized by short life, with fast sexual maturation, and continual spawning, is related to an increase in offspring survivorship and fitness in environments subjected to frequent disturbance (in this system pond drying). In this study, most gonads examined did not show fat in the visceral cavity indicating that the short life cycle of these species may require the use of energy reserves for feeding and reproductive activities (Vazzoler, 1996). Results from this study showed that both species of rivulids are fractional spawners, continuously reproducing as evidence by the presence of three different oocyte stages (e.g., immature, maturing and mature). Females of both species appear to begin reproducing when reaching a size of 20mm SL. Neofundulus ornatipinnis has a significantly larger egg diameter than A. vandenbergi, but the egg diameters of both species in this study ranged within the sizes reported for Austrolebias nigrofasciatus Costa & Cheffe, 2001 (Volcan et al., 2011), Leptolebias spp. (Costa & Leal, 2009), Simpsonichthys spp. (Fava & Toledo-Piza, 2007) and C. melanotaenia (Arenzon, Peret & Bohrer, 1999). Based on the oocyte diameter and developmental stages, both species in this study likely lay eggs over relatively short periods (e.g., days). Volcan et al. (2011) found that in Southern Brazil, A. nigrofasciatus spawned an average 21.5 eggs/female/week, and similar results have been reported for Austrolebias belloti (Steindachner, 1881) in Argentina (Calviño, 2005). No information is known on the spawning rate of the fishes within the genus Neofundulus, though we would expect it to be higher than that of Austrolebias due to the higher absolute fecundity of N. ornatipinnis as compared to A. vandenbergi. Fractional spawning is a common trait in fishes possessing an opportunistic life history (e.g., annual fishes), but also a consequence of physiological variation due to environmental alterations, and possibly the result of exposure to predation from snakes (Schalk & Montaña, 2012), macroinvertebrates (Schalk, 2010), or carnivorous tadpoles of Ceratophrys cranwelli (Anura: Ceratophryidae) (Barrio, 1980) (Schalk, unpublished data) in these shallow and temporal habitats.
Fecundity is an important variable in fish life history, because it is a major component in fitness (Endler, 1986). Fecundity may increase with increased body size or differ with species in a population and/or between populations of the same species or seasonally in a population (Wootton, 1990). Fecundity of A. vandenbergi and N. ornatipinnis was positively correlated with body length, though N. ornatipinnis had a higher absolute fecundity than A. vandenbergi. This relationship also has been observed in other annual fishes such as Cynopeocilus melanotaenia (Gonçalves et al., 2011), in live-bearing poeciliid fishes (Reznick & Miles, 1989), and in other freshwater fish (Lowe-McConnell, 1975).
Mean of gonadosomatic index (GSI) estimated in females of N. ornatipinnis, did not indicate significant variation in the gonadal status in relation to gonad weight at the beginning nor at the end of rainy season, probably because of the continuously developmental stages observed in all females examined. However, higher values of GSI in March indicate a pronounced ovary weight towards the end of the rainy season. The GSI is an important indicator of the reproductive cycle and type of spawn of fishes. To better understand this index, more detailed studies are required to investigate the cues (e.g., feeding ecology and abiotic variables) that trigger these species to spawn and correlate them to GSI values across the entire rainy season.
The Gran Chaco is experiencing some of the highest deforestation rates compared with other ecoregions across the globe (Zak, Cabido & Hodgson, 2004). The restricted distribution of many species of rivulids in addition to their small body size and lack of information on their life history make these species particularly vulnerable to anthropogenic alterations to the landscape. Austrolebias vandenbergi and N. ornatipinnis are no exception; outside of the report by Montaña et al. (2012), virtually nothing is known about the ecology of either of these species. This study represents the first contribution of the reproductive biology of A. vandenbergi and N. ornatipinnis; and attempts to advance our understanding of the natural history of these species in extreme environments of the Bolivian Gran Chaco. In this region, there is a need for further studies in the biology and ecology of other species that inhabit these ponds (e.g. Austrolebias monstrosus Huber, 1995a; Osinaga, 2006). Our findings represent a basic tool for conservation of these species and their environments that are constantly threatened by anthropogenic activities.
Acknowledgments
We thank Florencio Mendoza and Doris Ticona for their assistance with the field work. We thank the Capitania del Alto y Bajo Isoso (CABI) for permission to conduct research in Isoso and Kathia Rivero at the Museo de Historia Natural de Noel Kempff Mercado for permit support. Rosa Leny Cuellar assisted with the logistic support while conducting research in Bolivia. Donald Taphorn, Wade A. Ryberg, and three anonymous reviewers provided constructive comments on the manuscript. Support was provided by the National Science Foundation’s Graduate Research Fellowship Program and the Applied Biodiversity Science NSF-IGERT Program at Texas A&M University (NSF-IGERT Award # 0654377). This is publication number 1455 of the Biodiversity Research and Teaching Collections at Texas A&M University.
References
Arenzon, A. A., Peret, A. C., & Bohrer, M. B. (1999). Reproduction of the annual fish Cynopoecilus maelanotaenia (Regan, 1912) based on temporary water body population in Rio Grande do Sul State, Brazil. Hydrobiologia, 411, 65-70. [ Links ]
Belote, D. F., & Costa, W. J. E. M. (2004). Reproductive behavior patterns in three species of the South American annual fish genus Austrolebias Costa, 1998 (Cyprinodontiformes, Rivulidae). Boletim do Museu Nacional, Nova Série, Rio de Janeiro, 514, 1-7. [ Links ]
Calviño, P. (2005). Austrolebias toba (Cyprinodontiformes: Rivulidae), una especie nueva de pez anual de la Argentina. Revista del Museo Argentino de Ciencias Naturales, 7(2), 188-190. [ Links ]
Costa, W. J. E. M. (1988). Sistemática e distribuição do gênero Neofundulus (Cyprinodontiformes, Rivulidae). Revista Brasileira de Biologia, 48(2), 103-111. [ Links ]
Costa, W. J. E. M. (1998). Phylogeny and classification of Rivulidae revisited: origin and evolution of annualism and miniaturization in rivulid fishes (Cyprinodontiformes: Aplocheiloidei). Journal of Comparative Biology, 3(1), 33-92. [ Links ]
Costa, W. J. E. M. (2002) Monophyly and Phylogenetic Relationships of the Neotropical Annual Fish Genera Austrolebias and Megalebias (Cyprinodontiformes: Rivulidae). Copeia, 2002(4), 916-927. [ Links ]
Costa, W. J. E. M. (2006). The South American annual killifish genus Austrolebias (Teleostei: Cyprinodontiformes: Rivulidae): phylogenetic relationships, descriptive morphology and taxonomic revision. Zootaxa, 1213, 1-162. [ Links ]
Costa, W. J. E. M. (2011). Phylogenetic position and taxonomic status of Anablepsoides, Atlantirivulus, Cynodonichthys, Laimosemion and Melanorivulus (Cyprinodontiformes: Rivulidae). Ichthyological Exploration of Freshwaters, 22(3), 233-249. [ Links ]
Costa, W. J. E. M. (2012). Delimiting priorities while biodiversity is lost: Rio’s seasonal killifishes on the edge of survival. Biodiversity and Conservation, 21(10), 1-10. [ Links ]
Costa, W. J. E. M., & Leal, F. (2009). Egg surface morphology in the Neotropical seasonal killifish genus Leptolebias (Telostei: Cyprinodontiformes: Rivulidae). Vertebrate Zoology 59(1), 25-29. [ Links ]
Endler, J. A. (1986). Natural selection in the wild. Princeton, NJ: Princeton University Press. [ Links ]
Fava, D., & Toledo-Piza, M. (2007). Egg surface structure in the annual fishes Simpsonichthys (subgenera Ophthalmolebias and Xenurolebias) and Nematolebias (Teleostei: Cyprinodontiformes: Rivulidae): variability and phylogenetic significance. Journal of Fish Biology, 71(3), 889-907. [ Links ]
Garcia, D., Loureiro, M., & Tassino, B. (2008). Reproductive behavior in the annual fish Austrolebiasreicherti Loureiro & García, 2004 (Cyprinodontiformes: Rivulidae). Neotropical Ichthyology, 6(2), 243-248. [ Links ]
Gonçalves, C., Souza, U. P., & Volcan, M. V. (2011). The opportunistic feeding and reproduction strategies of the annual fish Cynopoecilus melanotaenia (Cyprinodontiformes: Rivulidae) inhabiting ephemeral habitats on southern Brazil. Neotropical Ichthyology, 9(1), 191-200. [ Links ]
Huber, J. H. (1995a). Nouvelles collections de cyprinodontes paraguayens, avec description de 4 especes rivulinesi nedites et redecourvete dune espece a la localite typique jusqualors indeterminee. Killi-Contact, 23, 1-24. [ Links ]
Huber, J. H. (1995b). Synthetic description of four new annual Rivulin Cyprinodonts from Paraguay. Freshwater & Marine Aquarium, 18(11), 104-124. [ Links ]
Laufer, G., Arim, M., Loureiro, M., Piñeiro-Guerra, J. M., Clavijo-Baquet, S., & Fagúndez, C. (2009). Diet of four annual killifishes: an intra-interspecific comparison. Neotropical Ichthyology, 7(1), 77-86. [ Links ]
Loureiro, M., Duarte, A., & Zarucki, M. (2011). A new species of Austrolebias Costa (Cyprinodontiformes: Rivulidae) from northeastern Uruguay, with comments on distribution patterns. Neotropical Ichthyology, 9(2), 335-342. [ Links ]
Lowe-McConnell, R. H. (1975). Fish communities in tropical freshwaters. London: Longmans. [ Links ]
Montaña, C. G., Schalk, C. M., & Taphorn, D. C. (2012). First record of Van den Berg’s Pearlfish, Austrolebias vandenbergi Huber, 1995 (Cyprinodontiformes: Rivulidae) in Bolivia with comments on its diet and reproductive biology. Check List, 8(3), 589-591. [ Links ]
Navarro, G., & Maldonado, M. (2002). Geografía ecológica de Bolivia: vegetación y ambientes acuáticos. Santa Cruz de la Sierra, Boliva: Centro de Ecología Difusión Simon I Patino. [ Links ]
Olden, J. D., Hogan, Z. S., & Zanden, M. (2007). Small fish, big fish, red fish, blue fish: size-biased extinction risk of the world’s freshwater and marine fishes. Global Ecology and Biogeography, 16(6), 694-701. [ Links ]
Osinaga, K. (2006). Nuevo registro para Bolivia de Austrolebias monstrosus (Huber, 1995 Rivulidae). Kempffiana, 2(1), 60-62. [ Links ]
Reznick, R. N., & Miles, D. B. (1989). A review of life history patterns in Poeciliidfishes. In G. K. Meffe & F. F. Snelson (Eds.), Evolution and ecology of Poeciliid fishes (pp. 125-147). Englewood Cliffs, NJ: Prentice Hall. [ Links ]
Rosa, R. S., & Lima, F. C. T. (2008). Peixes. In A. B. M. Machado, G. M. Drummond & A. P. Paglia (Eds.), Livrovermelho da fauna brasileira ameaçada de extinção (pp. 8-258). Ministério do Meio Ambiente, Brasília. [ Links ]
Schalk, C. M. (2010). Physalaemus biligonigerus (NCN) Predation. Herpetological Review, 41(2), 202. [ Links ]
Schalk, C. M., & Montaña, C. G. (2012). Liophispoecilogyrus (Yellow-bellied Liophis) Diet and Foraging Behavior. Herpetological Review, 43(1), 148. [ Links ]
Shine, R. (1989). Ecological causes for the evolution of sexual dimorphism: review of the evidence. Quarterly Review of Biology, 64(4), 419-461. [ Links ]
Sparre, P., & Venema, S. C. (1997). Introduçãoà avaliação de mananciais de peixestropicais. Parte 1. Roma, Italia: FAO. [ Links ]
Vazzoler, A. E. (1996). Biologia da reprodução de peixes teleósteos: teoria e práctica. Maringá, Brasil: Eduem/SBI. [ Links ]
Volcan, M. V., Lanés, L. E. K., & Cheffe, M. M. (2010). Distribuição e conservação de peixes anuais (Cyprinodontiformes: Rivulidae) no município do Chuí, sul do Brasil. Biotemas, 23(4), 51-58. [ Links ]
Volcan, M. V., Fonseca, A. P., & Robaldo, R. B. (2011). Reproduction of the threatened annual killifish Austrolebiasnigrofasciatus (Cyprinodontiformes: Rivulidae), confined to a natural environment. Journal of Threatented Taxa, 3(6), 1868-1867. [ Links ]
Werner, E. E., Skelly, D. K., Relyea, R. A., & Yurewicz, K. L. (2007). Amphibian species richness across environmental gradients. Oikos, 116(10), 1697-1712. [ Links ]
Winemiller, K. O. (1992). Life-history strategies and the effectiveness of sexual selection. Oikos, 63(2), 318-327. [ Links ]
Winemiller, K. O. (2005). Life history strategies, population regulation, and implications for fisheries management. Canadian Journal of Fisheries and Aquatic Sciences, 62(4), 872-885. [ Links ]
Winemiller, K. O., & Rose, K. A. (1992). Patterns of life-history diversification in North American fishes: implications for population regulation. Canadian Journal of Fisheries and Aquatic Sciences, 49(10), 2196-2218. [ Links ]
Wootton, J. R. (1990). Ecology of teleost fishes. New York: Kluwer Academic Publisher. [ Links ]
Wourms, J. P. (1972). The developmental biology of annual fishes. III. Pre-embryonic and embryonic diapause of variable duration in the eggs of annual fishes. Journal of Experimental Zoology, 182(3), 389-414. [ Links ]
Zak, M. R., Cabido, M., & Hodgson, J. G. (2004). Do subtropical seasonal forests in the Gran Chaco, Argentina, have a future? Biological Conservation, 120(4), 589-598. [ Links ]
Belote, D. F., & Costa, W. J. E. M. (2004). Reproductive behavior patterns in three species of the South American annual fish genus Austrolebias Costa, 1998 (Cyprinodontiformes, Rivulidae). Boletim do Museu Nacional, Nova Série, Rio de Janeiro, 514, 1-7. [ Links ]
Calviño, P. (2005). Austrolebias toba (Cyprinodontiformes: Rivulidae), una especie nueva de pez anual de la Argentina. Revista del Museo Argentino de Ciencias Naturales, 7(2), 188-190. [ Links ]
Costa, W. J. E. M. (1988). Sistemática e distribuição do gênero Neofundulus (Cyprinodontiformes, Rivulidae). Revista Brasileira de Biologia, 48(2), 103-111. [ Links ]
Costa, W. J. E. M. (1998). Phylogeny and classification of Rivulidae revisited: origin and evolution of annualism and miniaturization in rivulid fishes (Cyprinodontiformes: Aplocheiloidei). Journal of Comparative Biology, 3(1), 33-92. [ Links ]
Costa, W. J. E. M. (2002) Monophyly and Phylogenetic Relationships of the Neotropical Annual Fish Genera Austrolebias and Megalebias (Cyprinodontiformes: Rivulidae). Copeia, 2002(4), 916-927. [ Links ]
Costa, W. J. E. M. (2006). The South American annual killifish genus Austrolebias (Teleostei: Cyprinodontiformes: Rivulidae): phylogenetic relationships, descriptive morphology and taxonomic revision. Zootaxa, 1213, 1-162. [ Links ]
Costa, W. J. E. M. (2011). Phylogenetic position and taxonomic status of Anablepsoides, Atlantirivulus, Cynodonichthys, Laimosemion and Melanorivulus (Cyprinodontiformes: Rivulidae). Ichthyological Exploration of Freshwaters, 22(3), 233-249. [ Links ]
Costa, W. J. E. M. (2012). Delimiting priorities while biodiversity is lost: Rio’s seasonal killifishes on the edge of survival. Biodiversity and Conservation, 21(10), 1-10. [ Links ]
Costa, W. J. E. M., & Leal, F. (2009). Egg surface morphology in the Neotropical seasonal killifish genus Leptolebias (Telostei: Cyprinodontiformes: Rivulidae). Vertebrate Zoology 59(1), 25-29. [ Links ]
Endler, J. A. (1986). Natural selection in the wild. Princeton, NJ: Princeton University Press. [ Links ]
Fava, D., & Toledo-Piza, M. (2007). Egg surface structure in the annual fishes Simpsonichthys (subgenera Ophthalmolebias and Xenurolebias) and Nematolebias (Teleostei: Cyprinodontiformes: Rivulidae): variability and phylogenetic significance. Journal of Fish Biology, 71(3), 889-907. [ Links ]
Garcia, D., Loureiro, M., & Tassino, B. (2008). Reproductive behavior in the annual fish Austrolebiasreicherti Loureiro & García, 2004 (Cyprinodontiformes: Rivulidae). Neotropical Ichthyology, 6(2), 243-248. [ Links ]
Gonçalves, C., Souza, U. P., & Volcan, M. V. (2011). The opportunistic feeding and reproduction strategies of the annual fish Cynopoecilus melanotaenia (Cyprinodontiformes: Rivulidae) inhabiting ephemeral habitats on southern Brazil. Neotropical Ichthyology, 9(1), 191-200. [ Links ]
Huber, J. H. (1995a). Nouvelles collections de cyprinodontes paraguayens, avec description de 4 especes rivulinesi nedites et redecourvete dune espece a la localite typique jusqualors indeterminee. Killi-Contact, 23, 1-24. [ Links ]
Huber, J. H. (1995b). Synthetic description of four new annual Rivulin Cyprinodonts from Paraguay. Freshwater & Marine Aquarium, 18(11), 104-124. [ Links ]
Laufer, G., Arim, M., Loureiro, M., Piñeiro-Guerra, J. M., Clavijo-Baquet, S., & Fagúndez, C. (2009). Diet of four annual killifishes: an intra-interspecific comparison. Neotropical Ichthyology, 7(1), 77-86. [ Links ]
Loureiro, M., Duarte, A., & Zarucki, M. (2011). A new species of Austrolebias Costa (Cyprinodontiformes: Rivulidae) from northeastern Uruguay, with comments on distribution patterns. Neotropical Ichthyology, 9(2), 335-342. [ Links ]
Lowe-McConnell, R. H. (1975). Fish communities in tropical freshwaters. London: Longmans. [ Links ]
Montaña, C. G., Schalk, C. M., & Taphorn, D. C. (2012). First record of Van den Berg’s Pearlfish, Austrolebias vandenbergi Huber, 1995 (Cyprinodontiformes: Rivulidae) in Bolivia with comments on its diet and reproductive biology. Check List, 8(3), 589-591. [ Links ]
Navarro, G., & Maldonado, M. (2002). Geografía ecológica de Bolivia: vegetación y ambientes acuáticos. Santa Cruz de la Sierra, Boliva: Centro de Ecología Difusión Simon I Patino. [ Links ]
Olden, J. D., Hogan, Z. S., & Zanden, M. (2007). Small fish, big fish, red fish, blue fish: size-biased extinction risk of the world’s freshwater and marine fishes. Global Ecology and Biogeography, 16(6), 694-701. [ Links ]
Osinaga, K. (2006). Nuevo registro para Bolivia de Austrolebias monstrosus (Huber, 1995 Rivulidae). Kempffiana, 2(1), 60-62. [ Links ]
Reznick, R. N., & Miles, D. B. (1989). A review of life history patterns in Poeciliidfishes. In G. K. Meffe & F. F. Snelson (Eds.), Evolution and ecology of Poeciliid fishes (pp. 125-147). Englewood Cliffs, NJ: Prentice Hall. [ Links ]
Rosa, R. S., & Lima, F. C. T. (2008). Peixes. In A. B. M. Machado, G. M. Drummond & A. P. Paglia (Eds.), Livrovermelho da fauna brasileira ameaçada de extinção (pp. 8-258). Ministério do Meio Ambiente, Brasília. [ Links ]
Schalk, C. M. (2010). Physalaemus biligonigerus (NCN) Predation. Herpetological Review, 41(2), 202. [ Links ]
Schalk, C. M., & Montaña, C. G. (2012). Liophispoecilogyrus (Yellow-bellied Liophis) Diet and Foraging Behavior. Herpetological Review, 43(1), 148. [ Links ]
Shine, R. (1989). Ecological causes for the evolution of sexual dimorphism: review of the evidence. Quarterly Review of Biology, 64(4), 419-461. [ Links ]
Sparre, P., & Venema, S. C. (1997). Introduçãoà avaliação de mananciais de peixestropicais. Parte 1. Roma, Italia: FAO. [ Links ]
Vazzoler, A. E. (1996). Biologia da reprodução de peixes teleósteos: teoria e práctica. Maringá, Brasil: Eduem/SBI. [ Links ]
Volcan, M. V., Lanés, L. E. K., & Cheffe, M. M. (2010). Distribuição e conservação de peixes anuais (Cyprinodontiformes: Rivulidae) no município do Chuí, sul do Brasil. Biotemas, 23(4), 51-58. [ Links ]
Volcan, M. V., Fonseca, A. P., & Robaldo, R. B. (2011). Reproduction of the threatened annual killifish Austrolebiasnigrofasciatus (Cyprinodontiformes: Rivulidae), confined to a natural environment. Journal of Threatented Taxa, 3(6), 1868-1867. [ Links ]
Werner, E. E., Skelly, D. K., Relyea, R. A., & Yurewicz, K. L. (2007). Amphibian species richness across environmental gradients. Oikos, 116(10), 1697-1712. [ Links ]
Winemiller, K. O. (1992). Life-history strategies and the effectiveness of sexual selection. Oikos, 63(2), 318-327. [ Links ]
Winemiller, K. O. (2005). Life history strategies, population regulation, and implications for fisheries management. Canadian Journal of Fisheries and Aquatic Sciences, 62(4), 872-885. [ Links ]
Winemiller, K. O., & Rose, K. A. (1992). Patterns of life-history diversification in North American fishes: implications for population regulation. Canadian Journal of Fisheries and Aquatic Sciences, 49(10), 2196-2218. [ Links ]
Wootton, J. R. (1990). Ecology of teleost fishes. New York: Kluwer Academic Publisher. [ Links ]
Wourms, J. P. (1972). The developmental biology of annual fishes. III. Pre-embryonic and embryonic diapause of variable duration in the eggs of annual fishes. Journal of Experimental Zoology, 182(3), 389-414. [ Links ]
Zak, M. R., Cabido, M., & Hodgson, J. G. (2004). Do subtropical seasonal forests in the Gran Chaco, Argentina, have a future? Biological Conservation, 120(4), 589-598. [ Links ]
*Correspondencia a:
Christopher M. Schalk. Ecology, and Evolutionary Biology Program, Department of Wildlife & Fisheries Sciences and Biodiversity Research and Teaching Collections, 210 Nagle Hall, Texas A&M University, College Station, TX 77843-2258, USA. Correo electrónico cschalk@tamu.edu
Carmen G. Montaña. Ecology, and Evolutionary Biology Program, Department of Wildlife & Fisheries Sciences and Biodiversity Research and Teaching Collections, 210 Nagle Hall, Texas A&M University, College Station, TX 77843-2258, USA. Correo electrónico car1607@tamu.edu
Monika E. Libson. Ecology, and Evolutionary Biology Program, Department of Wildlife & Fisheries Sciences and Biodiversity Research and Teaching Collections, 210 Nagle Hall, Texas A&M University, College Station, TX 77843-2258, USA. Correo electrónico monikaeloisa@hotmail.com
1. Ecology, and Evolutionary Biology Program, Department of Wildlife & Fisheries Sciences and Biodiversity Research and Teaching Collections, 210 Nagle Hall, Texas A&M University, College Station, TX 77843-2258, USA. Correo electrónico cschalk@tamu.edu, car1607@tamu.edu, monikaeloisa@hotmail.com
Received 14-ii-2013. Corrected 12-vii-2013. Accepted 21-viii-2013.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}