










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.62 n.1 San José Jan./Mar. 2014
Distribution and additive partitioning of diversity in freshwater mollusk communities in Southern Brazilian streams
Distribución y partición aditiva de la diversidad de las comunidades de moluscos de agua dulce en arroyos del Sur de Brasil
Distribución y partición aditiva de la diversidad de las comunidades de moluscos de agua dulce en arroyos del Sur de Brasil
*Dirección para correspondencia:
Abstract
Additive partitioning of species diversity is a promising approach for analyzing patterns of diversity in mollusk communities, especially their spatial distribution. Our aims were to assess the distribution of mollusk communities in Southern Brazilian streams, and to evaluate the partitioning of community diversity at different spatial scales. The study was carried out in the lower course of the Toropi River, one of the main tributaries of the Ibicuí River Basin, in Southern Brazil. Four microbasins were considered: Sertão da Mata, Ribeirão, Tororaipi and Chiniquá, and sampling were undertaken in autumn, April and May 2009. Six sites were sampled in each stream: two in 1st-order segments, two in 2nd-order segments, and two in 3rd-order segments. All species found and the community as a whole, exhibited a clumped distribution. However, the variance-to-mean ratios for the Drepanotrema kermatoides and Heleobia bertoniana were higher than those of other species, suggesting a higher degree of aggregation. The additive partitioning of the species richness showed that the observed richness at smallest scale (α=within streams) represented 20.7%, and among-streams (β1) represented 10.5% of the total richness. The richness and Shannon diversity index observed at the alpha scale, were higher than those observed at the first level of beta diversity scale (β1=among-streams). The interaction between passive dispersal, tolerance to changes in some environmental variables, abiotic factors, and clumped distribution might have determined the spatial distribution of the communities studied. The greatest variation at the larger scales of analysis, involving among-orders and among-microbasins (β2 and β3, respectively) components, was expected, considering that the increase in distance leads to greater differences in richness (higher beta diversity). In conclusion, our results showed that the clumped distribution influenced the partition of the diversity of the mollusk communities in the streams studied. Dispersal methods and tolerance to variations in abiotic factors also have important roles in determining high alpha diversity. The partitioning of diversity showed that to preserve limnetic mollusks, it is important to preserve local areas, i.e., streams. Accordingly, conservation of streams on a local scale will contribute to conservation of the entire basin and maintenance of regional diversity (gamma).
Key words: distribution, Neotropics, spatial scale, beta diversity, clumped.
Resumen
La partición aditiva de la diversidad de especies es un enfoque prometedor para el análisis de los patrones de diversidad en las comunidades de moluscos, especialmente su distribución espacial. Nuestros objetivos fueron evaluar la distribución de las comunidades de moluscos en los arroyos en el Sur de Brasil, y evaluar la partición de diversidad de la comunidad en diferentes escalas espaciales. El muestreo se llevó a cabo en cuatro cuencas del curso inferior del río Toropi, Brasil. Todas las especies encontradas, y también la comunidad en su conjunto mostraron una distribución agregada. La partición aditiva de la riqueza de especies mostró que la riqueza observada en menor escala (α=dentro de los arroyos) representó el 20.7%, y entre los arroyos (β1) representó el 10.5% de la riqueza total. La riqueza y diversidad de Shannon observadas en escala alfa (α=dentro de los arroyos) fueron mayores que las observadas en el primer nivel de escala de la diversidad beta (β1=entre los arroyos). La mayor variación en las grandes escalas de análisis, con la participación de los componentes entre- órdenes y entre-microcuencas (β2 y β3, respectivamente) fue la esperada, ya que el aumento de la distancia conduce a mayores diferencias de riqueza (mayor diversidad beta). En conclusión, nuestros resultados muestran que la dispersión agregada tuvo influencia en la partición de la diversidad de las comunidades de moluscos en los arroyos estudiados. Los métodos de dispersión y tolerancia a las variaciones de los factores abióticos también juegan un papel importante para determinar la diversidad alfa. Así, la conservación de los arroyos a nivel local contribuirá a la conservación de la cuenca y al mantenimiento de la diversidad regional (gamma).
Palabras clave: distribución, Neotrópico, escalas espaciales, diversidad beta, agregación.
Species diversity in an area is important for examining many important questions in ecology as well as for the development of management actions for conserving biodiversity (Engen, Saether, Sverdrup-Thygeson, Grotan & Odegard, 2008). However, spatial patterns of species diversity change over multiple scales (Collier & Clements, 2011). The pattern observed within a local community might be very different from these found over broader areas such as landscapes or regions (Crist, Veech, Gering & Summerville, 2003). Thus, to effectively conserve regional biodiversity, conservationists need to know how diversity is distributed across spatial scales (Jost, DeVries, Walla, Greeney, Chao & Ricotta, 2010).
Partitioning of total species diversity into additive components within and between communities provides a framework by which diversity can be measured at different levels of organization (Lande, 1996). Total taxonomic richness in a region (γ-diversity) consists of two components, a within-assemblage component (α-diversity) and an among-assemblage component (β-diversity) (Whittaker, 1960, 1972). Studies have recently recognized applications for diversity partitioning in survey design (DeVries, Murray & Lande, 1997). An analytical approach based on the analysis of additive partitioning of diversity (Lande, 1996) is very useful for studies of conservation and restoration (Crist et al., 2003). The additive partitioning of diversity decomposes regional differences into components that are expressed in the same units and can be easily compared at different scales of space and time (Crist et al., 2003). For aquatic communities, the additive partitioning approach has been employed for many animal groups, such as fish assemblages (Eros, 2007), aquatic insects (Heino, Louhi & Muotka, 2004; Ligeiro, Melo & Callisto, 2010; Hepp & Melo, 2013) and aquatic nematodes (Flach, Ozorio & Melo, 2012).
River catchment basins are hierarchically organized and nested ecosystems comprising of river systems, stream segments, reach systems, riffle-pool sequences, and microhabitats (Allan & Castillo, 2007). Thus, additive partitioning of species diversity is a promising approach for analyzing patterns of diversity sampled from hierarchically scaled studies (Lande, 1996; Crist et al., 2003; Ligeiro et al., 2010; Hepp, Landeiro & Melo, 2012). It provides an operational method for analyzing species diversity across multiple spatial scales (Crist et al., 2003), and to identify the most important sources of diversity and target conservation efforts appropriately (Stefanescu, Olaru & Boengiu, 2008).
Freshwater environments are one of the most threatened ecosystems in the world. Human activities have altered water characteristics (e.g., Ricciardi & Rasmussen, 1999; Mansur, Heydrich, Pereira, Richnitti, Tarasconi & Rios, 2003), which leads to degradation, and sometimes destruction of these environments (Moreno, Navarro & Las Heras, 2006; Hepp & Santos, 2009; Sensolo, Hepp, Decian & Restello, 2012). In these settings, mollusks are one of the most imperiled animal groups (Ricciardi & Rasmussen, 1999; Brown, 2001; Bogan, 2006). In North America, many mussel species have been extirpated due to damming of major rivers (Bogan, 2006). This problem also occurs in other regions, and with other mollusk taxa, but is less well documented.
Efforts to comprehend the ecological requirements of freshwater mollusks conservation have increased, but have been generally addressed to taxa of Unionoidea (e.g., Strayer, Hunter, Smith & Borg, 1994; McRae, Allan & Burch, 2004; Bogan, 2008). Studies of mollusk communities as a whole, especially those focusing on their spatial distribution, are relatively few (e.g., McRae et al., 2004; Souza, Guilhermino & Antunes, 2005; Lewin, 2006; Pérez-Quintero, 2007; Maltchik, Stenert, Kotzian & Pereira, 2010). Studies about their diversity at different scales are lacking. However, some studies have shown that stream order, substrate, and macrophytes are important drivers (Vannote, Minshal, Cummins, Sedell & Cushing, 1980; Miserendino, 2001; Buss, Baptista, Nessimian & Egler, 2004; Heino & Muotka, 2006; Hepp et al., 2012). In fact, studying freshwater mollusk communities can be a very difficult task (Dorazio, 1999). Clumped distribution (Elliott, 1977; Downing, 1991; Downing & Downing, 1992; Vasconcelos, Espírito-Santo & Barboza, 2009), among other biotic (passive dispersion by means of vertebrate vectors) and intrinsic factors, needs intensive sampling efforts, making it difficult to analyze the structure and distribution of their communities (Dorazio, 1999). However, understanding how mollusks are distributed at different spatial scales could provide important data for defining preservation areas.
The Rio Grande do Sul state, localized in the southernmost Brazil, have a rich hydrographical net. The slope of the plateau is one of the most well preserved region of the state. This ecoregion shows numerous microbasins, formed by streams of small to medium order, characterized by variation in grain size due to surface geology (Vieira, 1984). However, the integrity of these microbasins have been affected by the construction of dams for agricultural activities, which alter the water flow, converting lotic environments in lentic ones. This context shows the urgency in comprehending the diversity and the environmental factors that drive the spatial distribution of mollusk communities inhabiting the streams of the state. In this study we evaluated: i) the spatial distribution of mollusk communities in four micro-basins of Southern Brazilian streams, and ii) the partitioning of diversity of the community at different spatial scales. We believe that the distribution characteristics of mollusks have a direct influence on the partitioning of diversity, in different components of α and β diversity considering different spatial scales. We also believe that α should be higher, or as higher as, β diversity scales due the clumped distribution and passive dispersion of mollusks, and due the landscape homogeneity (small spatial scale of study). This information will allow properly choosing spatial and local environmental factors to preserve the riverine malacofauna of Southernmost Brazil.
Material and Methods
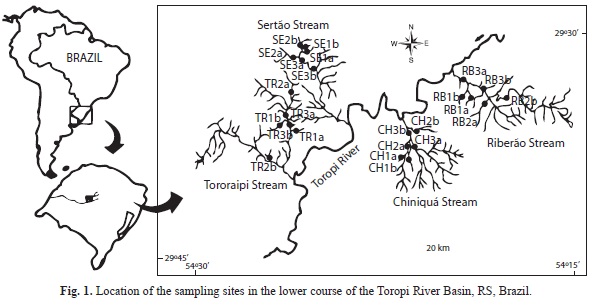
The study was conducted in the lower course of the Toropi River, one of the main tributaries of the Ibicuí River Basin in Southern Brazil (Fig. 1). The river basin is 3 310 km2 in extent (Paiva et al., 2000), and the lower course runs through a transition region, with a low-relief gradient, in the foothills of the Lower Northwest Slope, in the North, and the plain of the Central Depression in the South (Vieira, 1984). In this region, the river flows over the sandstones of the Botucatu and Caturrita formations (Robaina, Trentin, Bazzan, Reckziegel, Verdum & De Nardin, 2010). In general, the river and streambeds have gravel sediment on the slopes, and sand in floodplain areas (Vieira, 1984). The land is used for some fruit orchards in the foothills, but rice plantations and cattle grazing and breeding predominate, especially in the lowlands.
The climate of the region is subtropical humid, with warm summers or Cfa according to Köppen classification (Maluf, 2000). Rain-fall is evenly distributed through the year, with annual totals ranging from 1 500 to 1 750mm; the mean annual temperature varies between 18 and 20ºC, with the lowest temperatures (12°C) in June and July (Pereira, Garcia Netto & Borin, 1989). The vegetation is a transition zone between the Seasonal Deciduous Forest (forest galleries) and the Savanna (grasslands) within the Pampa biome (Marchiori, 2002). The present-day vegetation is highly degraded by conversion to pasture, but riparian vegetation is well preserved on the slopes.
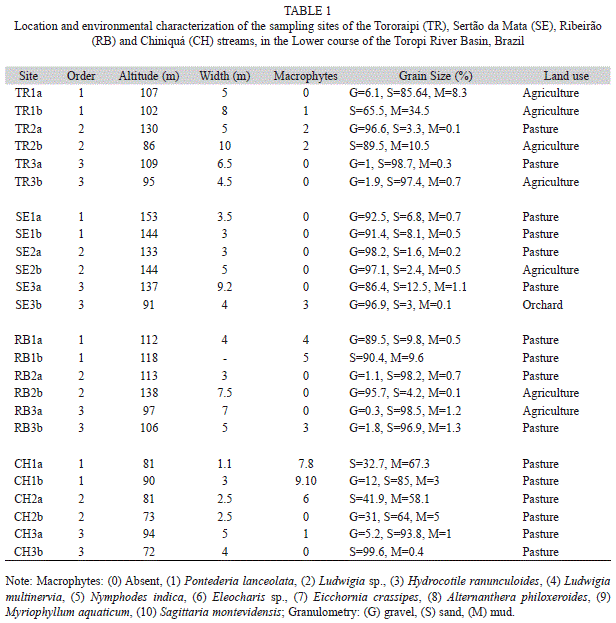
With respect to land use around the streams and sampling sites, pasture predominated at 20 sites (83%). Riparian vegetation was present at 71% of the sites, and 21% of the first-order stretches had no riparian vegetation. Macrophytes were present at 46% of the sites, and sparsest along the Sertão da Mata stream (Table 1). Current velocity (mean=0.19m/s), pH (mean=7.23) and dissolved oxygen (mean=8.29mg/L) varied little among the streams. The streambeds were predominantly sandy, except in the Sertão da Mata, where gravel predominated. Biochemical oxygen demand and water conductivity values were higher in the Sertão da Mata and Tororaipi streams than in the other two.
Sampling was carried out in autumn, in April and May 2009, when floods and dry periods are not common, in four microbasins of the lower course of the Toropi River (Fig. 1). The Sertão da Mata (ca. 71km2) and Ribeirão (ca. 76km2) streams are located closest to the Lower Northwest Slope, their beds have coarse sediment (gravels), and the vegetation cover is well developed. Land use along the Sertão da Mata is mainly fruit orchards, and along the Ribeirão mainly pasture. In the Tororaipi (ca. 153km2) and Chiniquá (ca. 50km2) streams, farther from the slope (Fig. 1), the streambeds are sandy, and only riparian vegetation occurs. Along both streams, land use is devoted to farming, with several rice plantations along the Tororaipi stream. In each stream, six sites were sampled, two in 1st-order segments, two in 2nd-order segments, and two in 3rd-order segments, according to Strahler’s classification (Strahler & Strahler, 1989). At each site, three quadrats of 1x5m were sampled with sieves (mesh=1mm), which were used to dredge the sediment to a depth of 10cm. Gravel was removed and the pieces individually examined. Each quadrat was sampled for 20 minutes, by four similarly trained persons.
Chi-square tests were used to measure patchiness in species populations or in whole communities (i.e., whether the organisms are distributed randomly through the samples, or clumped or uniformly distributed). Variance lower than the mean shows organisms uniformly spaced, whereas variance higher than the mean indicates that organisms are clumped (Krebs, 1999). The analysis was performed with the Beta/Species Distribution test using the BioDiversity Pro 2.0 software (McAleece, Lambshead & Paterson, 1997).
The additive partitioning of species diversity was analyzed by means of a biological matrix, hierarchically organized as follows: within streams (α), among streams (β1), among orders (β2) and among microbasins (β3). Thus, the partitioning diversity model evaluated was: γ (regional iversity) =α+β1+β2+β3. The taxonomic richness, expressed by the number of species identified, and the Shannon index was used as measure of diversity. The significance of the test was obtained from 1 000 randomizations based on sampling units (Crist et al., 2003). The test evaluates how much of the observed diversity is larger or smaller than that expected from the null model. High proportions (e.g., p>0.975) indicate that the observed values were smaller than those expected by chance. On the other hand, low proportions (e.g., p<0.025) indicate that the observed values were larger than those expected. The test was performed in the software R (R Development Core Team, 2012) using the boot package (Canty & Ripley, 2006).
Results
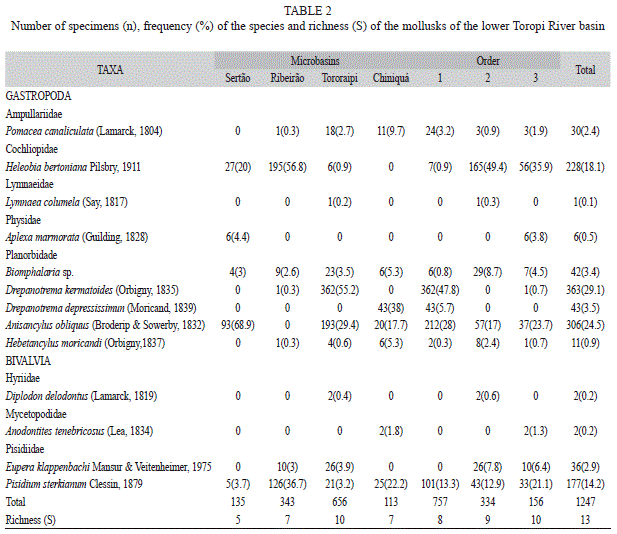
A total of 1 247 specimens were collected and assigned to 13 species (Table 2). Specimens of Biomphalaria were too small and young, preventing identification at species level. Gastropods encompassed 1 030 specimens (82.6%), and 217 bivalves (17.4%). The dominant species were the gastropods Drepanotrema kermatoides (d’Orbigny, 1835) (29.1%) and Anisancylus obliquus (Broderip & Sowerby, 1832) (24.5%). Among the bivalves, Pisidium dorbignyi Clessin, 1879 was the most abundant species (14.2%). Mollusks were not found in 27 quadrats and at four sites. Regarding the occurrence of each species in the 24 sites sampled, the most frequent were Biomphalaria sp., A. obliquus, P. dorbignyi and Heleobia bertoniana Pilsbry, 1911, in descending order. Drepanotrema depressissimum (Moricand, 1839), Lymnaea columella (Say, 1817), Aplexa marmorata (Guilding, 1828) and Diplodon delodontus (Lamarck, 1819) were each one found in only one site.
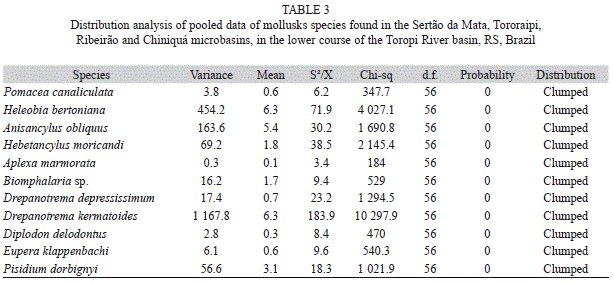
Richness was higher in the Tororaipi Stream and in 1st and 2nd order segments (Table 2), and composition also varied according these spatial factors. For all species found, the chi-square test showed variance values greater than the mean. Thus, all the species found, and also the community as a whole, showed a clumped distribution (Table 3). However, the variance-to-mean ratios for D. kermatoides and H. bertoniana were higher than those of other species, suggesting a greater clumped degree.
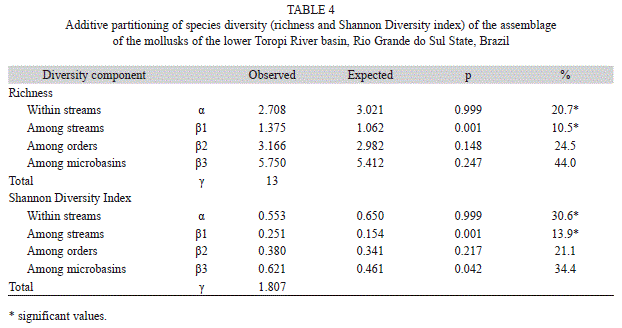
The additive partitioning of the species richness showed that the observed richness at the smallest scale (α=within streams) represented 20.7% of the total richness, and was smaller than that expected by the null model (p=0.999) (Table 4). Richness among streams (β1) comprised 10.5% of the total richness and was higher than that expected by the null model (p<0.001). Although the richness on the among-orders and among-microbasins scales (β2=24.5%, p=0.148; β3=44%, p=0.247, respectively) was high, the observed values were similar to those expected by the null model. The additive partitioning of Shannon diversity index showed that the value observed at the smallest scale was smaller than that expected by the null model (α=30.6%, p=0.999). The among-streams scale showed values of Shan-non diversity higher than those expected by the null model (β1=13.9%, p<0.001). The among-orders and among-microbasins scales showed observed values similar to the expected ones (21.1%, p=0.217; β3=34.4%, p=0.042).
Discussion
Relatively few studies have analyzed and quantified the clumped life habit in freshwater mollusks, but the phenomenon has been mentioned in the literature (Elliott, 1977). Thus, the clumped distribution of all species and of the community as a whole in the streams of the Toropi River basin is not surprising. Several investigators have commented that reproduction is the main cause for freshwater-mollusk clumped condition (Downing & Downing, 1992; Downing, Rochon & Pérusse, 1993; Ansell, 2001; Martell, Tunnicliffe & MacDonald, 2002). Limitations in dispersal capacity can also contribute to the clumped pattern (Thompson & Townsend, 2006) and spatial autocorrelation of organisms’ distribution (Legendre, 1990). Active dispersal in mollusks can be considered limited, i.e., slow (e.g., crawling and rheotactic) (Kappes & Haase, 2012). However, mollusks can compensate for this ‘limitation’ by using passive but efficient means of dispersal, such as current drift and vertebrate vectors (Kappes & Haase, 2012).
Previous studies have shown that geo-graphical distance is one of the main factors affecting variability in stream macroinvertebrate assemblages (Costa & Melo, 2008; Hepp et al., 2012; Hepp & Melo, 2013). If the variation in community composition is random, and accompanied by biotic processes that generate spatial autocorrelation (e.g., reproduction), a gradient in species composition may appear, and beta diversity can be interpreted in terms of the rate of change, or turnover, in species composition along that gradient. Thus, models generated in additive partitioning studies conducted with riverine macroinvertebrates have shown that richness on the smallest scale (α) was lower than that expected by chance (e.g., Ligeiro et al., 2010). In our study, the model generated for the mollusk communities showed a different pattern than those generally obtained for riverine macroinvertebrates. The richness and Shannon diversity index observed at the small scale (α=within-streams) were higher than those observed at the first level of beta diversity scale (β1=among-streams), and the expected values indicated that these differences should be higher.
The distribution pattern observed for mollusk communities studied can be explained by a combination of mollusk intrinsic and biotic factors, including not only the clumped life habit, but also the capability of many species to tolerate variations in abiotic factors, and their dispersal methods. These intrinsic factors must also be influenced by extrinsic ones, i.e., by the environmental features of the sampling sites. Many freshwater mollusks are widely tolerant to environmental conditions, such as desiccation (Jokinen, 1978) and low water oxygen content (McMahon, 1983). In Southern Brazil, freshwater mollusk species are tolerant to variations in some important abiotic factors, such as grain size and presence of macrophytes (see review by Martello, Kotzian & Simões, 2006). In general, the microbasins, as well as many of the studied sites, showed similar values for abiotic factors such as pH and dissolved oxygen. Only macrophytes and substrate grain size varied, especially in the Sertão da Mata microbasin. However, no species was found exclusively in this stream. Members of Heleobia and Hebetancylus inhabit streams and rivers with a variety of substrates (e.g., Lanzer, 1996; Pereira, Veitenheimer-Mendes, Mansur & Silva, 2000).
The role of vertebrates in mollusk dispersal is poorly understood, but the use of fish is a well-known indirect method of dispersal (Kat, 1984). Birds and other terrestrial vertebrates (Brown, 2007; Cadeé, 2011) have also been mentioned as important vectors. In the Ibicuí River basin, which encompasses the Toropi River basin, mollusks with soft parts were found in the final portion of the intestine of some fish, constituting the second most important food item (10.3%; Fagundes, Behr & Kotzian, 2008). Previous studies also suggest that they are able to survive passage through fish and birds’ digestive tracts (Brown, 2007; Cadeé, 2011). In rivers and streams of Southern Brazil, mollusks possibly also use these animals to disperse. Thus, the sites where a freshwater mollusk species can be found could be considered random, because their occurrence must be determined by vertebrate routes of displacement.
In other words, an interaction among passive dispersal, tolerance to changes in some environmental abiotic factors, and clumped distribution may have determined the spatial distribution of the communities studied here. If one or more mollusks abandon their host/vector, settling in a favorable site, they will colonize this place, and their populations could be abundant. In fact, the association between aggregation (variance-to-mean ratio) and abundance may be positive: the more resources available, the greater the abundances and the more clumped the individuals around the patchy resources (Rice & Lambshead,
1994). However, if the mollusks settle in an unfavorable site, they can survive because they are tolerant to abiotic factors variations, but their populations will be low. Thus, species composition and richness can be very variable from site to site, and be random in a particular area, even if environments allow the existence of many species. In any case, populations will be spatially clumped, a condition related to mollusk reproductive characteristics (Downing & Downing, 1992; Downing et al., 1993; Ansell, 2001; Martell et al., 2002). The absence of mollusks from many quadrats and some sites studied here, and the localized occurrence and abundance of many species in the streams of the lower course of the Toropi River basin support the argument for this phenomenon.
The greater variation on some larger scales of analyses (although similarly), involving among-orders and among-microbasins (β2 and β3) components was expected, considering that the increase in distance leads to greater differences in richness (higher beta diversity). Several other studies reported this pattern (Heino et al., 2004, Ligeiro et al., 2010; Hepp et al., 2012). The structure of basins and heterogeneity created by topographical limits demonstrates particularly strong effects on beta diversity, due to the effect of restraint carried by the terrestrial landscape in separating rivers (Kerr, Southwood & Cihlar, 2001; Clarke, Macnally, Bond & Lake, 2008). However, in the region studied, microbasins are separated by the main course of the Toropi River, and the spatial distribution of mollusk communities should be more influenced by the displacement of their vectors than by other factors. On the other hand, grain size and macrophytes exhibited more variation among microbasins than within microbasins, determining that the stream order should have had little influence on mollusk diversity. In other words, although the region studied is homogeneous concerning relief and land use, the small differences in local factors, such as substrate grain size and presence of macrophytes, both very important drivers of mollusk communities (Horsák & Hájek, 2003), should have coupled to clumped distribution and passive dispersion, providing more importance to α and β1 scales.
In conclusion, our results showed that the clumped distribution influenced the partition of the diversity of mollusk communities in the streams studied. Dispersal methods and tolerance to variations in abiotic factors also have important roles in determining high alpha diversity. Our results further showed that variability on larger scales may occur, which is likely to be marked at distances greater than 100km. This conclusion is based on the high variation observed at the larger scales, even if not significant when compared to the null model. The partitioning of diversity showed that to preserve limnetic mollusks, it is important to preserve local areas, i.e., streams. Conservation of streams on a local scale will also contribute to a broader conservation of the basin, thus contributing to the maintenance of regional diversity (gamma).
Acknowledgments
The authors thank Maria Cristina Dreher Mansur (Universidade Federal do Rio Grande do Sul) and Rosane Lanzer (Universidade de Caxias do Sul) for identification of the specimens of Pisidium and Ancylini. Special thanks to Andréa Salvarrey, Bruna Braun, Elisangela Secretti, Elzira Floss, Mateus Pires, Nícolas Figueiredo, Roger de Sá, Sarah Freitas and Vanessa Baptista for helping during fieldwork. This research is part of the doctoral thesis of A. Martello at the UFSM. Financial support for this study was provided by CAPES. L. U. Hepp received financial support from CNPq (proc. #471572/2012-8) and FAPERGS (proc. #12/1354-0).
References
Allan, J. D., & Castillo, M. M. (2007). Stream ecology: structure and function of running waters. New York: Springer. [ Links ]
Ansell, A. D. (2001). Dynamics of aggregation of a gastropod predator/scavenger on a New Zealand harbour beach. Journal of Molluscan Studies, 67(3), 329-341. [ Links ]
Bogan, A. E. (2006). Conservation and extinction of the freshwater molluscan fauna of North America. In C. F. Sturm, & A. Valdés (Eds.), The mollusks: a guide to their study, collection, and preservation (p. 373-383). Pittsburgh: American Malacological Society. [ Links ]
Bogan, A. E. (2008). Global diversity of freshwater mussels (Mollusca, Bivalvia) in freshwater. Hydrobiologia, 198, 139-147. [ Links ]
Brown, M. K. (2001). Mollusca: Gastropoda. In J. H. Thorp, & A. P. Covich (Eds.), Ecology and classification of North American Freshwater Invertebrates (p. 297-329). San Diego: Academic Press. [ Links ]
Brown, R. J. (2007). Freshwater mollusks survive fish gut passage. Arctic, 60(2), 124-128. [ Links ]
Buss, D. F., Baptista, D. F., Nessimian, J. L., & Egler, M. (2004). Substrate specificity, environmental degradation and disturbance structuring macroinvertebrate assemblages in neotropical streams. Hydrobiologia, 518(1), 179-188. [ Links ]
Cadée, G. C. (2011). Hydrobia as “Jonah in the Whale”: shell repair after passing through the digestive tract of Shelducks alive. Palaios, 26(4), 245-249. [ Links ]
Canty, A., & Ripley, B. D. (2006). Boot: BootstrapR (S-Plus) Functions. R package version 1.2-27. [ Links ]
Clarke, A., MaCnally, R., Bond, N. & Lake, P. S. (2008). Macroinvertebrate diversity in headwater streams: a review. Freshwater Biology, 53(9), 1707-1721. [ Links ]
Collier, K. J., & Clements, B. L. (2011). Influences of catchment and corridor imperviousness on urban stream macroinvertebrate communities at multiple spatial scales. Hydrobiologia, 664(1), 35-50. [ Links ]
Costa, S. S., & Melo, A. S. (2008). Beta diversity in stream macroinvertebrate assemblages: among-site and among-microhabitat components. Hydrobiologia, 598(1), 131-138. [ Links ]
Crist, T. O., Veech, J. A., Gering, J. C., & Summerville, K. S. (2003). Partitioning species diversity across landscapes and regions: a hierarchical analysis of α, β, and γ diversity. The American Naturalist, 162(6), 734-743. [ Links ]
DeVries, P. J., Murray, D., & Lande, R. (1997). Species diversity in vertical, horizontal, and temporal dimensions of a fruit-feeding butterfly community in an Ecuadorian Rainforest. Biological Journal of the Linnean Society, 62(3), 343-364. [ Links ]
Dorazio, R. M. (1999). Design-based and model-based inference in surveys of freshwater mollusks. Journal of the North American Benthological Society, 18(1), 118-131. [ Links ]
Downing, J. A. (1991). The effect of habitat structure on the spatial distribution of freshwater invertebrate populations. In S. S. Bell, E. D. McCoy, & H. R. Mushinsky (Eds.), Habitat Structure (p. 87-106). London: Chapman & Hall. [ Links ]
Downing, J. A., & Downing, W. L. (1992). Spatial aggregation, precision, and power in surveys of freshwater mussels populations. Canadian Journal of Fisheries and Aquatic Sciences, 49(5), 985-991. [ Links ]
Downing, J. A., Rochon, Y., & Pérusse, M. (1993). Spatial aggregation, body size, and reproductive success in the freshwater mussel Elliptio complanata. Journal of the North American Benthological Society, 12(2), 148-156. [ Links ]
Elliot, J. M. (1977). Some methods for the statistical analysis of samples of benthic invertebrates. Ambleside: Freshwater Biological Association. [ Links ]
Engen, S., Saether, B., Sverdrup-Thygeson, A., Grotan, V., & Odegaard, F. (2008). Assessment of species diversity from species abundance distributions at different localities. Oikos, 117(5), 738-748. [ Links ]
Eros, T. (2007). Partitioning the diversity of riverine fish: the roles of habitat types and non-native species. Freshwater Biology, 52(7), 1400-1415. [ Links ]
Fagundes, C. K., Behr, E. R., & Kotzian, C. B. (2008). Diet of Iheringichthys labrosus (Siluriformes, Pimelodidae) in the Ibicuí River, Southern Brazil. Iheringia, Sér. Zool., 98(1), 60-65. [ Links ]
Flach, P. Z. S., Ozorio, C. P., & Melo, A. S. (2012). Alpha and beta components of diversity of freshwater nematodes at different spatial scales in subtropical coastal lakes. Fundamental and Applied Limnology, 180(3), 249-258. [ Links ]
Heino, J., & Muotka, T. (2006). Landscape position, local environmental factors, and the structure of molluscan assemblages of lakes. Landscape Ecology, 21(4), 499-507. [ Links ]
Heino, J., Louhi, P., & Muotka, T. (2004). Identifying the scales of variability in stream macroinvertebrate abundance, functional composition and assemblage structure. Freshwater Biology, 49(9), 1230-1239. [ Links ]
Hepp, L. U., & Melo, A. S. (2013). Dissimilarity of stream insect assemblages: effects of multiple scales and spatial distances. Hydrobiologia, 703(1), 239-246. [ Links ]
Hepp, L. U., & Santos, S. (2009). Benthic communities of streams related to different land uses in a hydrographic basin in southern Brazil. Environmental Monitoring and Assessment, 157(1-4), 305-318. [ Links ]
Hepp, L. U., Landeiro, V. L., & Melo, A. S. (2012). Experimental assessment of the effects of environmental factors and longitudinal position on alpha and beta diversities of aquatic insects in a Neotropical stream. International Review of Hydrobiology, 97(2), 157-167. [ Links ]
Horsák, M., & Hájek, M. (2003). Composition and species richness of molluscan communities in relation to vegetation and water chemistry in the western Carpathian Spring Fens: the poor-rich gradient. Journal of Molluscan Studies, 69(4), 349-357. [ Links ]
Jokinen, E. H. (1978). The aestivation pattern of a population of Lymnaea elodes (Say) (Gastropoda: Lymnaeidae). American Midland Naturalist, 100(1), 43-53. [ Links ]
Jost, L., DeVries, P., Walla, T., Greeney, H., Chao, A., & Ricotta, C. (2010). Partitioning diversity for conservation analyses. Diversity and Distributions, 16(1), 65-76. [ Links ]
Kappes, H., & Haase, P. (2012). Slow, but steady: dispersal of freshwater mollusks. Aquatic Sciences, 74(1), 1-14. [ Links ]
Kat, P. W. (1984). Parasitism and the Unionacea (Bivalvia). Biological Review, 59(2), 189-207. [ Links ]
Kerr, J. T., Southwood, T. R. E., & Cihlar, J. (2001). Remotely sensed habitat diversity predicts butterfly species richness and community similarity in Canada. Proceedings of the National Academy of Sciences, 98(20), 11365-11370. [ Links ]
Krebs, C. J. (1999). Ecological Methodology. New York: Harper Collins. [ Links ]
Lande, R. (1996). Statistics and partitioning of species diversity, and similarity among multiple communities. Oikos, 76(1), 5-13. [ Links ]
Lanzer, R. M. (1996). Ancylidade (Gastropoda, Basom-matophora) na América do Sul: sistemática e distribuição. Revista Brasileira de Zoologia, 13(1), 175-210. [ Links ]
Legendre, P. (1990). Quantitative methods and biogeographic analysis. In D. J. Gabary, & R. G. South (Eds.), Evolutionary biogeography of the marine algae of the North Atlantic (p. 9-34). Berlin: Springer Verlag. [ Links ]
Lewin, I. (2006). The gastropod communities in the low-land Rivers of agricultural areas – their biodiversity and bioindicative value in the Ciechanowska upland, Central Poland. Malacologia, 49(1), 7-23. [ Links ]
Ligeiro, R., Melo, A. S., & Callisto, M. (2010). Spatial scale and the diversity of macroinvertebrates in a Neotropical catchment. Freshwater Biology, 55(2), 424-435. [ Links ]
Maltchik, L., Stenert, C., Kotzian, C. B., & Pereira, D. (2010). Responses of freshwater mollusks to environmental factors in Southern Brazil wetlands. Brazilian Journal of Biology, 70(3), 473-482. [ Links ]
Maluf, J. R. T. (2000). Nova classificação climática do Estado do Rio Grande do Sul. Revista Brasileira de Agrometereologia, 8(1),141-150. [ Links ]
Mansur, M. C. D., Heydrich, I., Pereira, D., Richnitti, L. M. Z., Tarasconi, J. C., & Rios, E. C. (2003). Moluscos. In C. S. Fontana, G. A. Bencke, & R. E. Reis (Eds.), Livro vermelho da fauna ameaçada de extinção no Rio Grande do Sul (p. 49-71). Porto Alegre: EDIPUCRS. [ Links ]
Marchiori, J. N. C. (2002). Fitogeografia do Rio Grande do Sul: enfoque histórico e sistemas de classificação. Porto Alegre: EST. [ Links ]
Martell, K. A., Tunnicliffe, V., & MacDonald, I. R. (2002). Biological features of a buccinid whelk (Gastropoda, Neogastropoda) at the Endeavor ventfields of Juan de Fuça Ridge, northeast Pacific. Journal of Molluscan Studies, 68(1), 45-53. [ Links ]
Martello, A. R., Kotzian, C. B., & Simões, M. G. (2006). Quantitative fidelity of Recent freshwater mollusk assemblages from the Touro Passo River, Rio Grande do Sul. Iheringia, Série Zoologia, 96(4), 453-465. [ Links ]
McAleece, N., Lambshead, P. J. D., & Paterson, G. L. J. (1997). Biodiversity Pro (Version 2). London, The Natural History Museum & The Scottish Association for Marine Science. Retrieved from http://www.sams.ac.uk/research/software/bdpro.zip/view [ Links ]
McMahon, R. F. (1983). Physiological ecology of fresh-water pulmonates. In W. D. Russel-Hunter (Ed.), The Mollusca (p. 359-430). Orlando: Academic Press. [ Links ]
McRae, S. E., Allan, J. D., & Burch, J. B. (2004). Reach-and catchment-scale determinants of the distribution of freshwater mussels (Bivalvia: Unionidae) in south-eastern Michigan USA. Freshwater Biology, 49(2), 127-142. [ Links ]
Miserendino, M. L. (2001). Macroinvertebrate assemblages in Andean Patagonian rivers and streams: environmental relationships. Hydrobiologia, 444(1-3), 147-158. [ Links ]
Moreno, J. L., Navarro, C., & Las Heras, J. D. (2006). Abiotic ecotypes in South-central Spanish rivers: reference conditions and pollution. Environmental Pollution, 143(3), 388-396. [ Links ]
Paiva, J. B. D., Paiva, E. M. C. D., Gastaldini, M. C. C., Irion, C. A. O., Dias, J. R., Costa, L. C. M., & Richter, L. A. (2000). Quantificação da demanda de água na bacia do rio Ibicuí – RS. Revista Brasileira de Recursos Hídricos, 5(3), 93-111. [ Links ]
Pereira, D., Veitenheimer-Mendes, I. L., Mansur, M. C. D., & Silva, M. C. P. (2000). Malacofauna límnica do sistema de irrigação da microbacia do arroio Capivara, Triunfo, Brasil. Biociências, 8(1), 137-157. [ Links ]
Pereira, P. R. B., Garcia Netto, L. R., & Borin, C. J. A. (1989). Contribuição a geografia física do município de Santa Maria: unidades de paisagem. Geografia-Ensino e Pesquisa, 3, 37-68. [ Links ]
Pérez-Quintero, J. C. (2007). Diversity, habitat use and conservation of freshwater mollusks in the lower Guadiana River basin (SW Iberian Peninsula). Aquatic Conservation: Marine and Freshwater Ecosystems, 17(5), 485-501. [ Links ]
R Development Core Team (2012). R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna. Retrieved from http://www.R-project.org/ [ Links ]
Ricciardi, A., & Rasmussen, J. B. (1999). Extinction rates of North American freshwater fauna. Conservation Biology, 13(5), 1220-1222. [ Links ]
Rice, A. L., & Lambshead, J. D. (1994). Patch dynamics in the deepsea benthos: the role of a heterogeneous supply of organic matter. In P. S. Giller, A. G. Hildrew & D. G. Rafaelli (Eds.), Scale, pattern and process (p. 469-497). Proceedings British Ecological Society/American Society Limnology and Oceanography Symposium Aquatic Ecology, University College, Cork. [ Links ]
Robaina, L. E. S., Trentin, R., Bazzan, T., Reckziegel, E. W., Verdum, R., & De Nardin, D. (2010). Compartimentação Geomorfológica da Bacia Hidrográfica do Ibicuí, Rio Grande do Sul, Brasil: proposta de classificação. Revista Brasileira de Geomorfologia, 11(2), 11-23. [ Links ]
Sensolo, D., Hepp, L. U., Decian, V., & Restello, R. M. (2012). Influence of landscape on the assemblages of Chironomidae in Neotropical streams. Annales de Limnologie Limnologie – International Journal of Limnology, 48(4), 391-400. [ Links ]
Souza, R., Guilhermino, L., & Antunes, C. (2005). Molluscan fauna in the freshwater tidal área of the River Minho estuary, NW of Iberian Pensinsula. Annales de Limnologie-International Journal of Limnology, 41(2), 141-147. [ Links ]
Stefanescu, M. D., Olaru, M. L., & Boengiu, S. (2008). Partitioning raptor species diversity within the plain area located in the south of Dolj county, Romania. Natura Montenegrina, 7(3), 441-446. [ Links ]
Strahler, A. N., & Strahler, A. H. (1989). Geografia física. Barcelona: Omega. [ Links ]
Strayer, D. L., Hunter, D. C., Smith, L. C., & Borg, C. K. (1994). Distribution, abundance, and roles of freshwater clams (Bivalvia, Unionidae) in the fresh-water tidal Hudson River. Freshwater Biology, 31(2), 239-248. [ Links ]
Thompson, R., & Towsend, C. (2006). A truce with neutral theory: local deterministic factors, species traits and dispersal limitation together determine patterns of diversity in stream invertebrates. Journal of Animal Ecology, 75(2), 476-484. [ Links ]
Vannote, R. L., Minshal, G. W., Cummins, K. W., Sedell, J. R., & Cushing, C. E. (1980). The river continuum concept. Canadian Journal of Fisheries and Aquatic Sciences, 37, 130-137. [ Links ]
Vasconcelos, M. C., Espírito-Santo, M. M., & Barboza, F. A. R. (2009). Depth effects on the abundance, survivorship rate and size of Melanoides tuberculatus (Prosobranchia: Thiaridae) in Dom Helvécio Lake, Minas Gerais, Brazil. Acta Limnologica Brasiliensia, 21, 393-397. [ Links ]
Vieira, E. F. (1984). Rio Grande do Sul: Geografia física e vegetação. Porto Alegre: Sagra. [ Links ]
Whittaker, R. H. (1960). Vegetation of the Siskitou Mountains, Oregon and California. Ecological Monographs, 30, 279-338. [ Links ]
Whittaker, R. H. (1972). Evolution and measurement of species diversity. Taxon, 21, 213-251. [ Links ]
Ansell, A. D. (2001). Dynamics of aggregation of a gastropod predator/scavenger on a New Zealand harbour beach. Journal of Molluscan Studies, 67(3), 329-341. [ Links ]
Bogan, A. E. (2006). Conservation and extinction of the freshwater molluscan fauna of North America. In C. F. Sturm, & A. Valdés (Eds.), The mollusks: a guide to their study, collection, and preservation (p. 373-383). Pittsburgh: American Malacological Society. [ Links ]
Bogan, A. E. (2008). Global diversity of freshwater mussels (Mollusca, Bivalvia) in freshwater. Hydrobiologia, 198, 139-147. [ Links ]
Brown, M. K. (2001). Mollusca: Gastropoda. In J. H. Thorp, & A. P. Covich (Eds.), Ecology and classification of North American Freshwater Invertebrates (p. 297-329). San Diego: Academic Press. [ Links ]
Brown, R. J. (2007). Freshwater mollusks survive fish gut passage. Arctic, 60(2), 124-128. [ Links ]
Buss, D. F., Baptista, D. F., Nessimian, J. L., & Egler, M. (2004). Substrate specificity, environmental degradation and disturbance structuring macroinvertebrate assemblages in neotropical streams. Hydrobiologia, 518(1), 179-188. [ Links ]
Cadée, G. C. (2011). Hydrobia as “Jonah in the Whale”: shell repair after passing through the digestive tract of Shelducks alive. Palaios, 26(4), 245-249. [ Links ]
Canty, A., & Ripley, B. D. (2006). Boot: BootstrapR (S-Plus) Functions. R package version 1.2-27. [ Links ]
Clarke, A., MaCnally, R., Bond, N. & Lake, P. S. (2008). Macroinvertebrate diversity in headwater streams: a review. Freshwater Biology, 53(9), 1707-1721. [ Links ]
Collier, K. J., & Clements, B. L. (2011). Influences of catchment and corridor imperviousness on urban stream macroinvertebrate communities at multiple spatial scales. Hydrobiologia, 664(1), 35-50. [ Links ]
Costa, S. S., & Melo, A. S. (2008). Beta diversity in stream macroinvertebrate assemblages: among-site and among-microhabitat components. Hydrobiologia, 598(1), 131-138. [ Links ]
Crist, T. O., Veech, J. A., Gering, J. C., & Summerville, K. S. (2003). Partitioning species diversity across landscapes and regions: a hierarchical analysis of α, β, and γ diversity. The American Naturalist, 162(6), 734-743. [ Links ]
DeVries, P. J., Murray, D., & Lande, R. (1997). Species diversity in vertical, horizontal, and temporal dimensions of a fruit-feeding butterfly community in an Ecuadorian Rainforest. Biological Journal of the Linnean Society, 62(3), 343-364. [ Links ]
Dorazio, R. M. (1999). Design-based and model-based inference in surveys of freshwater mollusks. Journal of the North American Benthological Society, 18(1), 118-131. [ Links ]
Downing, J. A. (1991). The effect of habitat structure on the spatial distribution of freshwater invertebrate populations. In S. S. Bell, E. D. McCoy, & H. R. Mushinsky (Eds.), Habitat Structure (p. 87-106). London: Chapman & Hall. [ Links ]
Downing, J. A., & Downing, W. L. (1992). Spatial aggregation, precision, and power in surveys of freshwater mussels populations. Canadian Journal of Fisheries and Aquatic Sciences, 49(5), 985-991. [ Links ]
Downing, J. A., Rochon, Y., & Pérusse, M. (1993). Spatial aggregation, body size, and reproductive success in the freshwater mussel Elliptio complanata. Journal of the North American Benthological Society, 12(2), 148-156. [ Links ]
Elliot, J. M. (1977). Some methods for the statistical analysis of samples of benthic invertebrates. Ambleside: Freshwater Biological Association. [ Links ]
Engen, S., Saether, B., Sverdrup-Thygeson, A., Grotan, V., & Odegaard, F. (2008). Assessment of species diversity from species abundance distributions at different localities. Oikos, 117(5), 738-748. [ Links ]
Eros, T. (2007). Partitioning the diversity of riverine fish: the roles of habitat types and non-native species. Freshwater Biology, 52(7), 1400-1415. [ Links ]
Fagundes, C. K., Behr, E. R., & Kotzian, C. B. (2008). Diet of Iheringichthys labrosus (Siluriformes, Pimelodidae) in the Ibicuí River, Southern Brazil. Iheringia, Sér. Zool., 98(1), 60-65. [ Links ]
Flach, P. Z. S., Ozorio, C. P., & Melo, A. S. (2012). Alpha and beta components of diversity of freshwater nematodes at different spatial scales in subtropical coastal lakes. Fundamental and Applied Limnology, 180(3), 249-258. [ Links ]
Heino, J., & Muotka, T. (2006). Landscape position, local environmental factors, and the structure of molluscan assemblages of lakes. Landscape Ecology, 21(4), 499-507. [ Links ]
Heino, J., Louhi, P., & Muotka, T. (2004). Identifying the scales of variability in stream macroinvertebrate abundance, functional composition and assemblage structure. Freshwater Biology, 49(9), 1230-1239. [ Links ]
Hepp, L. U., & Melo, A. S. (2013). Dissimilarity of stream insect assemblages: effects of multiple scales and spatial distances. Hydrobiologia, 703(1), 239-246. [ Links ]
Hepp, L. U., & Santos, S. (2009). Benthic communities of streams related to different land uses in a hydrographic basin in southern Brazil. Environmental Monitoring and Assessment, 157(1-4), 305-318. [ Links ]
Hepp, L. U., Landeiro, V. L., & Melo, A. S. (2012). Experimental assessment of the effects of environmental factors and longitudinal position on alpha and beta diversities of aquatic insects in a Neotropical stream. International Review of Hydrobiology, 97(2), 157-167. [ Links ]
Horsák, M., & Hájek, M. (2003). Composition and species richness of molluscan communities in relation to vegetation and water chemistry in the western Carpathian Spring Fens: the poor-rich gradient. Journal of Molluscan Studies, 69(4), 349-357. [ Links ]
Jokinen, E. H. (1978). The aestivation pattern of a population of Lymnaea elodes (Say) (Gastropoda: Lymnaeidae). American Midland Naturalist, 100(1), 43-53. [ Links ]
Jost, L., DeVries, P., Walla, T., Greeney, H., Chao, A., & Ricotta, C. (2010). Partitioning diversity for conservation analyses. Diversity and Distributions, 16(1), 65-76. [ Links ]
Kappes, H., & Haase, P. (2012). Slow, but steady: dispersal of freshwater mollusks. Aquatic Sciences, 74(1), 1-14. [ Links ]
Kat, P. W. (1984). Parasitism and the Unionacea (Bivalvia). Biological Review, 59(2), 189-207. [ Links ]
Kerr, J. T., Southwood, T. R. E., & Cihlar, J. (2001). Remotely sensed habitat diversity predicts butterfly species richness and community similarity in Canada. Proceedings of the National Academy of Sciences, 98(20), 11365-11370. [ Links ]
Krebs, C. J. (1999). Ecological Methodology. New York: Harper Collins. [ Links ]
Lande, R. (1996). Statistics and partitioning of species diversity, and similarity among multiple communities. Oikos, 76(1), 5-13. [ Links ]
Lanzer, R. M. (1996). Ancylidade (Gastropoda, Basom-matophora) na América do Sul: sistemática e distribuição. Revista Brasileira de Zoologia, 13(1), 175-210. [ Links ]
Legendre, P. (1990). Quantitative methods and biogeographic analysis. In D. J. Gabary, & R. G. South (Eds.), Evolutionary biogeography of the marine algae of the North Atlantic (p. 9-34). Berlin: Springer Verlag. [ Links ]
Lewin, I. (2006). The gastropod communities in the low-land Rivers of agricultural areas – their biodiversity and bioindicative value in the Ciechanowska upland, Central Poland. Malacologia, 49(1), 7-23. [ Links ]
Ligeiro, R., Melo, A. S., & Callisto, M. (2010). Spatial scale and the diversity of macroinvertebrates in a Neotropical catchment. Freshwater Biology, 55(2), 424-435. [ Links ]
Maltchik, L., Stenert, C., Kotzian, C. B., & Pereira, D. (2010). Responses of freshwater mollusks to environmental factors in Southern Brazil wetlands. Brazilian Journal of Biology, 70(3), 473-482. [ Links ]
Maluf, J. R. T. (2000). Nova classificação climática do Estado do Rio Grande do Sul. Revista Brasileira de Agrometereologia, 8(1),141-150. [ Links ]
Mansur, M. C. D., Heydrich, I., Pereira, D., Richnitti, L. M. Z., Tarasconi, J. C., & Rios, E. C. (2003). Moluscos. In C. S. Fontana, G. A. Bencke, & R. E. Reis (Eds.), Livro vermelho da fauna ameaçada de extinção no Rio Grande do Sul (p. 49-71). Porto Alegre: EDIPUCRS. [ Links ]
Marchiori, J. N. C. (2002). Fitogeografia do Rio Grande do Sul: enfoque histórico e sistemas de classificação. Porto Alegre: EST. [ Links ]
Martell, K. A., Tunnicliffe, V., & MacDonald, I. R. (2002). Biological features of a buccinid whelk (Gastropoda, Neogastropoda) at the Endeavor ventfields of Juan de Fuça Ridge, northeast Pacific. Journal of Molluscan Studies, 68(1), 45-53. [ Links ]
Martello, A. R., Kotzian, C. B., & Simões, M. G. (2006). Quantitative fidelity of Recent freshwater mollusk assemblages from the Touro Passo River, Rio Grande do Sul. Iheringia, Série Zoologia, 96(4), 453-465. [ Links ]
McAleece, N., Lambshead, P. J. D., & Paterson, G. L. J. (1997). Biodiversity Pro (Version 2). London, The Natural History Museum & The Scottish Association for Marine Science. Retrieved from http://www.sams.ac.uk/research/software/bdpro.zip/view [ Links ]
McMahon, R. F. (1983). Physiological ecology of fresh-water pulmonates. In W. D. Russel-Hunter (Ed.), The Mollusca (p. 359-430). Orlando: Academic Press. [ Links ]
McRae, S. E., Allan, J. D., & Burch, J. B. (2004). Reach-and catchment-scale determinants of the distribution of freshwater mussels (Bivalvia: Unionidae) in south-eastern Michigan USA. Freshwater Biology, 49(2), 127-142. [ Links ]
Miserendino, M. L. (2001). Macroinvertebrate assemblages in Andean Patagonian rivers and streams: environmental relationships. Hydrobiologia, 444(1-3), 147-158. [ Links ]
Moreno, J. L., Navarro, C., & Las Heras, J. D. (2006). Abiotic ecotypes in South-central Spanish rivers: reference conditions and pollution. Environmental Pollution, 143(3), 388-396. [ Links ]
Paiva, J. B. D., Paiva, E. M. C. D., Gastaldini, M. C. C., Irion, C. A. O., Dias, J. R., Costa, L. C. M., & Richter, L. A. (2000). Quantificação da demanda de água na bacia do rio Ibicuí – RS. Revista Brasileira de Recursos Hídricos, 5(3), 93-111. [ Links ]
Pereira, D., Veitenheimer-Mendes, I. L., Mansur, M. C. D., & Silva, M. C. P. (2000). Malacofauna límnica do sistema de irrigação da microbacia do arroio Capivara, Triunfo, Brasil. Biociências, 8(1), 137-157. [ Links ]
Pereira, P. R. B., Garcia Netto, L. R., & Borin, C. J. A. (1989). Contribuição a geografia física do município de Santa Maria: unidades de paisagem. Geografia-Ensino e Pesquisa, 3, 37-68. [ Links ]
Pérez-Quintero, J. C. (2007). Diversity, habitat use and conservation of freshwater mollusks in the lower Guadiana River basin (SW Iberian Peninsula). Aquatic Conservation: Marine and Freshwater Ecosystems, 17(5), 485-501. [ Links ]
R Development Core Team (2012). R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna. Retrieved from http://www.R-project.org/ [ Links ]
Ricciardi, A., & Rasmussen, J. B. (1999). Extinction rates of North American freshwater fauna. Conservation Biology, 13(5), 1220-1222. [ Links ]
Rice, A. L., & Lambshead, J. D. (1994). Patch dynamics in the deepsea benthos: the role of a heterogeneous supply of organic matter. In P. S. Giller, A. G. Hildrew & D. G. Rafaelli (Eds.), Scale, pattern and process (p. 469-497). Proceedings British Ecological Society/American Society Limnology and Oceanography Symposium Aquatic Ecology, University College, Cork. [ Links ]
Robaina, L. E. S., Trentin, R., Bazzan, T., Reckziegel, E. W., Verdum, R., & De Nardin, D. (2010). Compartimentação Geomorfológica da Bacia Hidrográfica do Ibicuí, Rio Grande do Sul, Brasil: proposta de classificação. Revista Brasileira de Geomorfologia, 11(2), 11-23. [ Links ]
Sensolo, D., Hepp, L. U., Decian, V., & Restello, R. M. (2012). Influence of landscape on the assemblages of Chironomidae in Neotropical streams. Annales de Limnologie Limnologie – International Journal of Limnology, 48(4), 391-400. [ Links ]
Souza, R., Guilhermino, L., & Antunes, C. (2005). Molluscan fauna in the freshwater tidal área of the River Minho estuary, NW of Iberian Pensinsula. Annales de Limnologie-International Journal of Limnology, 41(2), 141-147. [ Links ]
Stefanescu, M. D., Olaru, M. L., & Boengiu, S. (2008). Partitioning raptor species diversity within the plain area located in the south of Dolj county, Romania. Natura Montenegrina, 7(3), 441-446. [ Links ]
Strahler, A. N., & Strahler, A. H. (1989). Geografia física. Barcelona: Omega. [ Links ]
Strayer, D. L., Hunter, D. C., Smith, L. C., & Borg, C. K. (1994). Distribution, abundance, and roles of freshwater clams (Bivalvia, Unionidae) in the fresh-water tidal Hudson River. Freshwater Biology, 31(2), 239-248. [ Links ]
Thompson, R., & Towsend, C. (2006). A truce with neutral theory: local deterministic factors, species traits and dispersal limitation together determine patterns of diversity in stream invertebrates. Journal of Animal Ecology, 75(2), 476-484. [ Links ]
Vannote, R. L., Minshal, G. W., Cummins, K. W., Sedell, J. R., & Cushing, C. E. (1980). The river continuum concept. Canadian Journal of Fisheries and Aquatic Sciences, 37, 130-137. [ Links ]
Vasconcelos, M. C., Espírito-Santo, M. M., & Barboza, F. A. R. (2009). Depth effects on the abundance, survivorship rate and size of Melanoides tuberculatus (Prosobranchia: Thiaridae) in Dom Helvécio Lake, Minas Gerais, Brazil. Acta Limnologica Brasiliensia, 21, 393-397. [ Links ]
Vieira, E. F. (1984). Rio Grande do Sul: Geografia física e vegetação. Porto Alegre: Sagra. [ Links ]
Whittaker, R. H. (1960). Vegetation of the Siskitou Mountains, Oregon and California. Ecological Monographs, 30, 279-338. [ Links ]
Whittaker, R. H. (1972). Evolution and measurement of species diversity. Taxon, 21, 213-251. [ Links ]
*Correspondencia a:
Alcemar R. Martello. Programa de Pós-Graduação em Biodiversidade Animal, Universidade Federal de Santa Maria, CEP 97105-970, Santa Maria, Rio Grande do Sul, Brazil. Correo electrónico armartello@hotmail.com
Luiz U. Hepp. Programa de Pós-Graduação em Ecologia, Universidade Regional Integrada do Alto Uruguai e das Missões – Campus de Erechim, CEP 99700-000, Rio Grande do Sul, Brazil. Correo electrónico lhepp@uricer.edu.br
Carla B. Kotzian. Departamento de Biologia, Universidade Federal de Santa Maria, CEP 97105-970, Santa Maria, Rio Grande do Sul, Brazil. Correo electrónico modrizralok@hotmail.com
1. Programa de Pós-Graduação em Biodiversidade Animal, Universidade Federal de Santa Maria, CEP 97105-970, Santa Maria, Rio Grande do Sul, Brazil. Correo electrónico armartello@hotmail.com
2. Programa de Pós-Graduação em Ecologia, Universidade Regional Integrada do Alto Uruguai e das Missões – Campus de Erechim, CEP 99700-000, Rio Grande do Sul, Brazil. Correo electrónico lhepp@uricer.edu.br
3. Departamento de Biologia, Universidade Federal de Santa Maria, CEP 97105-970, Santa Maria, Rio Grande do Sul, Brazil. Correo electrónico modrizralok@hotmail.com
Received 08-ii-2013. Corrected 10-viii-2013. Accepted 10-IX-2013.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}