










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.62 n.1 San José Jan./Mar. 2014
On Limnocytherina axalapasco, a new freshwater ostracod (Podocopida: Limnocytheridae) from Mexican crater lakes
*Dirrección para correspondencia:
Abstract
Limnocytherina is a genus conformed by 12 species; its distribution in the American continent is known to be exclusively on the North (neartics), but little is reported about its distribution from Mexico (transition zone) and Central America (Neotropics). Different sampling campaigns were undertaken in three crater lakes from the Axalapascos region in east-central Mexico, during 2008, 2009 and 2011. As a product of these campaings, the new species of Limnocytherina axalapasco was found, which displays some intraspecific variability among populations. In this study, we described the taxonomy, the habitat, the ecological preferences and the larval development of this new species. A total of 10 sediment samples (8 littoral, 2 deepest point) were collected from lakes Alchichica, La Preciosa and Quechulac. We found that L. axalapasco is closely related to two North American species: L. posterolimba and L. itasca as well as one Central American species L. royi comb. nov. With the inclusion of L. axalapasco and L. royi to the genus, the distribution of Limnocytherina is extended to Central America. The four most important distinguishing characters of this new species are: 1) valve surface and margins covered with small, spine-like projections; 2) most of the A1 setae with a highly developed setule at distal part, producing a bifurcate appearance; 3) the upper ramus on the hemipenis is elongated, and by far overpasses dorsal/distal margins, distal lobe is triangular and short, while the hook-like process is prominent, outward orientated, and overpassing the tip of the distal lobe; 4) the UR is moderately developed with seta f3 elongated and setae f1 and f2 short. Considering its ecological characteristics and larval development, L. axalapasco was preferably found in alkaline waters dominated by Cl- or HCO3- and Na+ or Mg2+, temperatures ranging between 19.1 to 20.3°C, and dissolved oxygen concentrations from 5 to 6.5mg/L. This species was abundant in deeper (~64m) areas of the saline Alchichica lake, where surface water displayed conductivity values of up to 2 250μS/cm, and the sand with low percentage of silt resulted the preferred substrate. Along with the description of L. axalapasco, we provide additional information on the hemipenis of L. itasca, L. royi and L. sanctipatricii, and we discuss on the Limnocytherina-type of hemipenis.
Key words: Ostracoda, taxonomy, ecology, Limnocytherina axalapasco, hemipenis.
Resumen
Limnocytherina axalapasco, un nuevo ostrácodo dulceacuícola (Podocopida: Limnocytheridae) de lagos de cráter mexicanos. Limnocytherina es un género conformado por 12 especies; su distribución en el continente Americano se conoce exclusivamente en el norte (zona neártica), pero se sabe poco de su distribución en México (zona de transición) y Centro América (zona neotropical). Luego de diferentes campañas de muestreo en tres lagos cráter de la región de los Axalapascos en la zona centroeste de México, durante 2008, 2009 y 2011, se encontró a Limnocytherina axalapasco, una nueva especie, que mostró variabilidad intraespecífica entre poblaciones. Un total de 10 muestras de sedimentos (8 del litoral, 2 del punto más profundo) fueron recolectadas de los lagos Alchichica, La Preciosa y Quechulac. L. axalapasco esta estrechamente relacionada con dos especies norteamericanas: L. posterolimba y L. itasca, así como con una especie centroamericana Limnocytherina royi comb. nov. Con la inclusión de L. axalapasco y L. royi al género, la distribución de Limnocytherina se extiende hasta Centro América. Los cuatro caracteres distintivos más importantes de esta nueva especie son: 1) superficie y márgenes de valvas cubiertos por pequeñas proyecciones parecidas a espinas; 2) La mayoría de las setas de A1 tienen una setula muy desarrollada en la zona distal, con apariencia de estar bifurcadas; 3) El ramo superior del hemipene es alargado y por mucho sobrepasa los márgenes dorsal/distal; el lóbulo distal es corto y triangular, mientras que el proceso prensil es prominente, orientado externamente y sobrepasa la longitud del lóbulo distal; 4) el UR esta moderadamente desarrollado con la seta f3 alargada y las setas f1 y f2 cortas. Para esta especie se describe también su hábitat, preferencias ecológicas y desarrollo larvario. L. axalapasco prefiere aguas alcalinas dominadas por Cl- o HCO3 - y Na+ o Mg2+, temperaturas entre 19.1 a 20.3°C y concentraciones de oxígeno disuelto de 5 a 6.5 mg/L. Esta especie es más abundante en las zonas profundas (~64m) del lago salino Alchichica, donde la conductividad del agua es de hasta 2 250μS/cm. Arenas con bajos porcentajes de limo parecen ser el sustrato preferido. Aunado a la descripción de L. axalapasco, proveemos información adicional para los hemipenes de L. itasca, L. royi y L. sanctipatricii, y discutimos el tipo de hemipene en Limnocytherina.
Palabras clave: Ostracoda, taxonomía, ecología, Limnocytherina axalapasco, hemipene.
Ostracods from Mexico and North Central America are poorly known, and so far the majority of studies ocused on paleolimnology (Bridgwater, Heaton, & O’Hara, 1999a; Bridgwater, Holmes, & O’Hara, 1999b; Palacios-Fest, Carreño, Ortega-Ramírez, & Alvarado-Valdez, 2002; Chávez-Lara, Priyadarsi, Caballero, Carreño, & Lakshumanan, 2012), only few studies are dealing with taxonomy and ecology (Furtos, 1936; Tressler, 1954). Despite this, 22 species has been described in the region so far, with most of the species (18) known from the Yucatán peninsula (Southern Mexico) and North Guatemala, several of such species seem to be endemic (Furtos, 1936; Furtos, 1938; Brehm, 1939; Pérez, Lorenschat, Brenner, Scharf & Schwalb, 2010a; Pérez et al., 2010b; Pérez et al., 2011; Pérez et al., 2012; Cohuo-Durán, Elías-Gutiérrez & Karanovic, 2013). From the Family Limnocytheridae, only one species has been described in the region, Limnocythere bradbury Forester, 1985, but nine species have been reported here. Limnocythere bradbury, L. friabilis Benson, & McDonald, 1963, L. staplini Gutentag & Benson, 1962, L. sappaensis Staplin, 1963, Limnocytherina itasca (Cole, 1969) and L. sanctipatricii (Brady & Robertson, 1869) are known from Northcentral Mexico (McKenzie & Swain, 1967; Swain 1967; Forester 1985; Bridgwater et al., 1999a; Maddocks, Machain-Castillo & Gío-Argáez, 2009), while Cytheridella Americana (Furtos, 1936), C. ilosvayi Daday, 1905, and Limnocythere opesta Brehm, 1939 are known from Southern Mexico and Northern Guatemala (Furtos, 1936; Furtos, 1938; Brehm, 1939; Pérez et al., 2010a). Recent surveys of the crater lakes in central Mexico revealed one new species from the genus Limnocytherina Negadaev-Nikonov, 1967. This genus was originally described as a subgenus of Limnocythere Brady, 1868, but Martens (1996, 2000) subsequently raised its systematic position to a separate genus. He mainly focused on some distinct characters of the hemipenis (only clearly described in the type species, Limnocytherina sanctipatricii but not in the other species of the genus). According to Martens & Savatenalinton (2011) Limnocytherina currently comprises 12 species, 11 being restricted to North America (Delorme, 1971), and only Limnocytherina sanctipatricii has been found in Europe as well (Delorme, 1971; Meisch, 2000). The objective of this work was to provide an accurate morphological description of this new species, and also include information on its distribution, habitat, cological preferences and larval development.
Materials and Methods
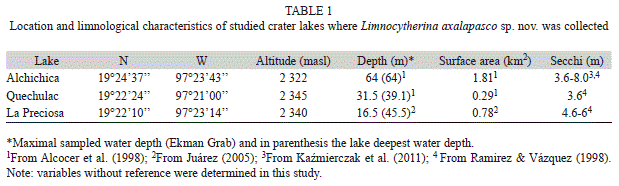
The ostracods were collected from the crater lakes Alchichica, Quechulac and La Preciosa, located in Puebla state, Mexico (Tables 1, 2). Sampling campaigns were carried out during the dry seasons of December 2008 and March 2009, and during the rainy season of June 2011. Sampling took approximately one day per study site. The samples were taken from the limnetic and littoral zones, using plankton net (50μm mesh size, 30cm in diameter). Surface sediments containing ostracods were also collected from Alchichica and Quechulac lake’s deepest point (63m and 31.5m) using Ekman grab. Sampling protocol follows Pérez et al. (2010a,b, 2011, 2012). All together, we collected 10 sediment samples (8 littoral, 2 lake deepest point samples; ca. 100mL wet sediment each) that were preserved in 96% ethanol, and stored in 100mL PE-bottles for ostracod analysis in the laboratory.
Environmental variables of lake waters (temperature, pH, dissolved oxygen, conductivity, total dissolved solids (TDS) were measured with a Hydrolab Hach Quanta QD03594. Secchi disc transparency at each lake was determined at the lake deepest point. Parallel water samples were collected for water chemistry analysis (CO32-, HCO3-, Cl-, SO42-, Na+, K+, Mg2+, Ca2+) which was carried out in the laboratory at the Universidad Nacional Autónoma de México (UNAM). Stable isotopes (
 18O, 13C, D) in lake waters were measured at the Department of Geological Sciences, University of Florida. The detailed methodology is in Pérez et al. (2010b). All samples were kept under refrigeration until previous analysis in the laboratory.
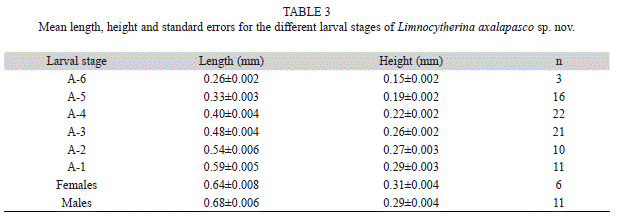
18O, 13C, D) in lake waters were measured at the Department of Geological Sciences, University of Florida. The detailed methodology is in Pérez et al. (2010b). All samples were kept under refrigeration until previous analysis in the laboratory. At least 300 ostracod valves (equivalent to 150 specimen carapaces) were extracted under a stereomicroscope Leica EZ4. Ten specimens with well-preserved soft parts were dissected, and soft parts mounted on icroscope slides in Faure’s medium following the procedure proposed by Stock & Von Vaupel Klein (1996). Drawings were made using a drawing tube attached to an Olympus BX51 bright-field compound microscope. Non- dissected material was preserved in plastic tubes with 70% ethanol. Ostracod valve measurements (n=100) and light microscope photographs were taken with a Canon Powershot A640 digital camera attached to a Zeiss Axiostar-plus light microscope. Length and height were determined using the software AxioVision Release 4.6.3. The mean values and standard errors are in table 3. Scanning Electron Microscope (SEM) photographs were taken with a Jeol JSM-5600LV LCM scanning microscope from the Central Microscopy Laboratory, Institute of Physics, Universidad Nacional Autónoma de México. The examined material was deposited in El Colegio de la Frontera Sur, Reference Zooplankton Collection (ECO-CH-Z, Accession numbers are provided). Hard parts were also deposited at the Museo María del Carmen Perrilliat from Instituto de Geología de la Universidad Nacional Autónoma de México (IGM, accession numbers are provided). Eleven Limnocytherina itasca specimens were collected from the littoral zone of a Tundra pond, Canada (58°37’33” N - 94°13’42” W), in August 2006. The analysis of COI gene for species comparison is not presented here, because it was not possible to amplify the gene with the available primers (Folmer, Black, Hoeh, Lutz, & Vrijenhoek, 1994; Schön, 2001; Martens, Rossetti, Butlin, & Schön, 2005) and because the methodology has to be improved for Cytherids. Therefore, the design of new specific primers for this group will be necessary and they will be published elsewhere.
Thoracopod terminology follows Meisch (2000), while the hemipenis terminology follows Danielopol, artens & Casale (1989) and Martens (1990, 1998). Here we also follow the setal classification proposed by Garm (2004). Abbreviations used in the text and figures (in alphabetical order): Al –antennula; A2 –antenna; Ex -Exopodite; f(1-3) –setae 1-3 of Uropodal Ramus; LV –left valve; masl –meters above sea level; Md –Mandibula; Mdp –Mandibular palp; Mx –Maxilla; Mxp –maxilular palp; Pr –Protopodite; RV –right valve;
Tl –first leg; T2 –second leg, T3 –hird leg; UR –Uropodal Ramus.
Results
Systematics (according to Martens, 2000 and Martens & Savatenalinton, 2011):
Order Podocopida Sars, 1866
Superfamily Cytheroidea Baird, 1850
Family Limnocytheridae Klie, 1938
Subfamily Limnocytherinae Klie, 1938
Tribe Limnocytherini Klie, 1938
Genus Limnocytherina Negadaev-Nikonov, 1967
Abbreviated diagnosis: Carapace subrectangular to subtrapezoidal, elongated in lateral view, weakly or strongly calcified. LV and RV subequal in shape and structures. Surface reticulated, smooth or noded, sometimes ornamented with tubercles, pustules and with a well or poorly developed allae. Marginal pore canals always present and unbranched. Marginal area often forming vestibule. Four adductor muscles arranged in a vertical row. Valves displaying sexual dimorphism: males being longer than females and often expanded posteriorly to accommodate a large hemipenis. A1 with the last segment fairly elongated with flagellated claws, A2 terminating with three strong claws. Mxp strong and 2-segmented. Thoracic legs increasing in size (T1<T2<T3) and with setae of variable length. Hemipenis with large upper ramus and sometimes with a tentacle-like structure. Lower ramus elongated and with a hook-like process. Uropodal ramus moderately developed, with f1 and f2 setae normally developed and f3 transformed into a broad and sclerified rami. Copulatory process with variable morphology.
Type species: Limnocytherina sanctipatricii (Brady & Robertson, 1869).
Limnocytherina axalapasco sp. nov.
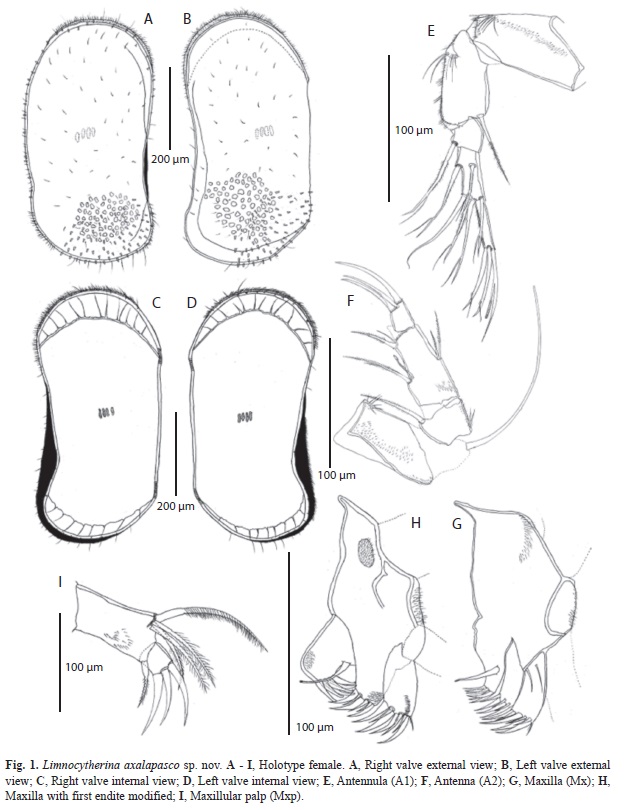
(Fig. 1-3, 5)
Type material: Collected by Ana L. Martínez, December 15, 2008. Holotype: female dissected on one slide (ECO-CH-Z-07548) from Lake Alchichica (Table 1). Allotype: Male dissected on one slide (ECO-CH-Z- 07547) from the same locality. Paratypes: 10 specimens (5 males and 5 females) from the same locality, all of them preserved in 70% ethanol (ECO-CH-Z-07556). One male and female carapace preserved in separated paleontological slides (IGM-1497-Mi, IGM-1502-Mi).
Other material: Collected by Ana L. Martínez, December 16, 2008. Two females from Lake La Preciosa (Table 1), dissected on separate two slides (ECO-CH-Z-07553; ECOCH-Z-07554), one male from the same locality, dissected on one slide (ECO-CH-Z-07555), one female carapace preserved in one paleontological slide (IGM-1509-Mi). One female from Lake Quechulac (Table 1) dissected on one slide (ECO-CH-Z-07550) and two males from the same locality dissected in separated slides (ECO-CH-Z-07551; ECO-CH-Z-07552).
Etymology: The species is named after the region of Axalapascos where the studied crater lakes (Alchichica, Quechulac, La Preciosa) are located and where the ostracod species were collected in high abundance (<240valves/ml in surface sediments). The name should to be treated as a noun in apposition.
Differential diagnosis: In both sexes surface valves reticulated, covered with small spine-like projections. In females the valves are slightly downward projected posteriorly. A1 with the majority of setae with a long setule distally. A2 with spinneret (exopodite) not reaching distal end of last segment. T3 sexually dimorphic, in females 5-segmented and with dp–seta reduced; in males 4-segmented (last segment and claw fused) and with dp–seta swollen and strong. Hemipenis with distal lobe triangular and short; upper ramus elongated by far overpassing lateral/distal lobe; hook-like process exceeding tip of distal lobe. UR moderately developed with claw-like f3. Posterior part of the body of females with the uropod bearing two seta; genital lobe cone shaped ending with a rounded tip.
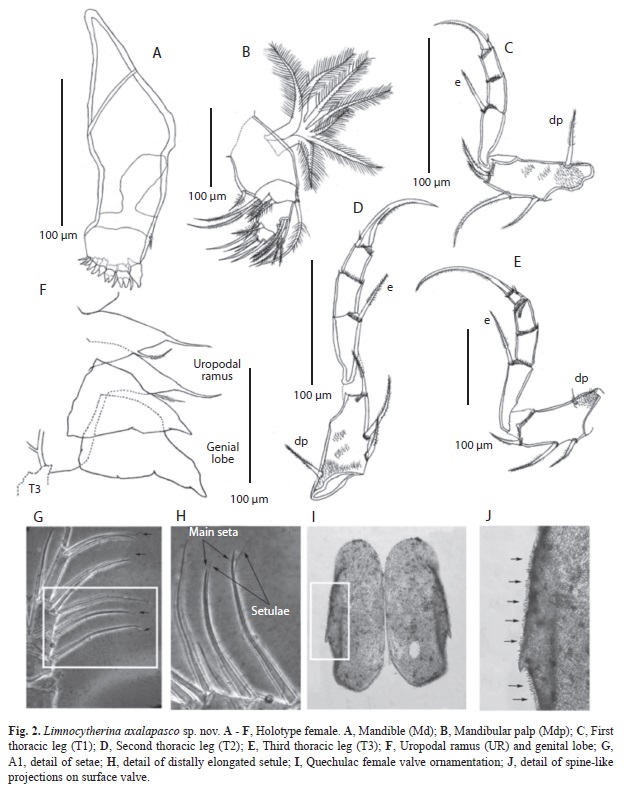
Female (Holotype)
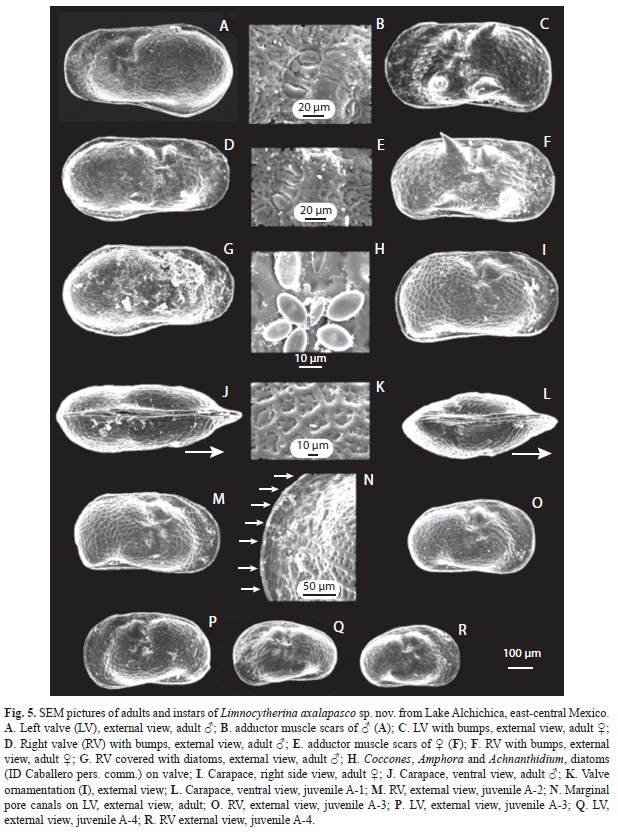
Hard parts: Carapace subrectangular. Valves ornamented with nodes, concentrated medially, in some specimens the nodes are modified into poor or well developed alar prolongations (Fig. 5C, F). Surface reticulate and covered with few, short, simple hair-like setae, as well as few small spine-like projections (Fig.1A, B; 2J), which are also present along margins. Marginal pore canal unbranched. Valves slightly asymmetrical with LV overlapping RV, anteriorly. LV (Fig. 1B, D), length 0.62mm and height 0.27mm. Dorsal margin straight and covered with short setae. Ventral margin slightly concave medially and postero-distally slightly downward projected. Anterior margin rounded with short setae. Selvage peripheral. Posterior margin downward projected with few setae. RV (Fig. 1A, C), length 0.61mm and height 0.25mm. Dorsal margin almost straight, covered with few setae. Ventral margin slightly concave medially. Anterior margin rounded with selvage peripheral and abundant setae. Posterior margin projected downward (Fig. 5I) with few long setae. Four muscular imprints on shell (Fig. 1C, D; 5B, E).
Soft parts: A1 (Fig. 1E). 5-segmented. First segment with no setae, but with fringe of short and delicate setules on anterior margin, ending with six long setules antero-distally. Second segment bearing one apical serrulate seta not reaching distal end of penultimate segment; anterior margin with several long and unequal setules. Third segment with one apical anterior serrulate seta (with last setule elongated) reaching half length of last segment (Fig. 2G, H). Fourth segment with two serrulate setae situated medially, each one with a long setule distally (Fig. 2G, H). Same segment distally with four distinct setae. The most anterior one simple seta clearly exceeding distal end of terminal segment. Following three setae with a long setule almost at the end, producing a bifurcate appearance (Fig. 2G, H). Fifth segment hirsute anteriorly; distally carrying three setae, longer one fused with aesthetasc; other two setae subequally long and with a long setule distally. Length ratios of four distal segments 1.5:0.6:1.1:1.
A2 (Fig. 1F). 4-segmented. First segment with strong setules antero-proximally and short setules along posterior margin; exopodite not reaching distal end of last segment. Second segment with one antero-apical serrulate seta, not reaching distal end of penultimate segment; postero-proximally with group of setules. Third segment with two unequal and serrulate setae postero-medially, longer one, just reaching distal end of terminal segment. Antero-medially with the Y–aesthetasc and two unequal serrulate setae, longer one just exceeding distal end of third segment. Antero-distally on the same segment, two setae, one of them, long and strong, reaching distal end of terminal segment; second one smooth, half as long as previous seta. Fourth segment with three claws, two of them equally long, 2.2 times longer than terminal segment; third claw 2.6 times longer than terminal segment. Length ratios of last three segment 3.2:1.15:1.
Md (Fig. 2A). Mandibular coxa armed with ten teeth and two teeth-like setae, short simple seta present proximally on coxa.
Mdp (Fig. 2B). 4-segmented. First segment bearing exopodite composed of seven long plumose rays. Same segment anteriorly bearing short serrulate seta not reaching distal end of segment. Second segment with five serrulate setae at distal end, most anterior one by far exceeding distal end of terminal segment. Most posterior ones subequally long overpassing distal end of terminal segment. Two medial setae equally long and not reaching distal end of penultimate segment. Third segment anteromedially with two groups of serrulate setae, proximal one with three equally long setae, distal one with two subequal setae exceeding distal end of terminal segment. Postero-distally three apical setae present, two of them short, just exceeding distal end of last segment; third seta smooth, 4.8 times longer than terminal segment. Fourth segment with four unequally
long claw-like setae.
Mx endites (Fig. 1G). Third endite ending with six short and strong teeth-like setae; a swollen and long seta situated medially. Second and first endites ending with six and seven teeth-like setae respectively; on proximal side of first endite two subequally long and simple setae present.
Mxp (Fig. 1I). 2-segmented. First segment with short and sparsely distributed setules; antero-apically one short and strong serrulate seta present, which exceeds distal end of terminal segment. Posteriorly, four pappose setae present, longer one 6.2 times as long as terminal segment. The two subequally long setae are 3/4 as long as longer seta. Smaller seta slightly exceeds half length of longest seta. Second segment with three claw-like setae, subequally long, 3.4 times longer than segment.
T1 (Fig. 2C). 4-segmented. Protopodite hirsute and posteriorly armed with four unequally long serrulate setae. Seta dp short and strong, situated more proximal on segment. Second segment with e–seta not reaching distal end of terminal segment. Fourth segment ending with a claw, 2.5 times longer than segment. Basally this claw is bearing one short seta. Length ratios of last three segments 2.1:1.1:1.
T2. (Fig. 2D). 4-segmented. Protopodite with three serrulate setae posteriorly. Most proximal seta, short and strong. Medial seta strong, exceeding distal end of the segment; apical seta short not reaching distal end of following segment. Seta dp relatively short, thickened at its base. Second segment with e–seta, exceeding distal end of penultimate segment. Fourth segment bearing one long claw, which basally carries a small seta. Length ratios of last three segments 2.2:1.2:1.
T3. (Fig. 2E). 5-segmented. First segment posteriorly with three serrulate setae, thickened at the base and unequally long. Seta dp tiny and smooth. Second segment with long and serrulate e–seta, just reaching distal end of penultimate segment. Fifth segment bearing short seta medially and ending with one elongated claw, 8.3 times longer than terminal segment. Length ratios of four last segments 6.2:3:2.4:1.
Posterior part of body (Uropod) consisting of two unequal setae. Genital lobe cone-shaped ending in a round tip (Fig. 2F).
Male (Allotype)
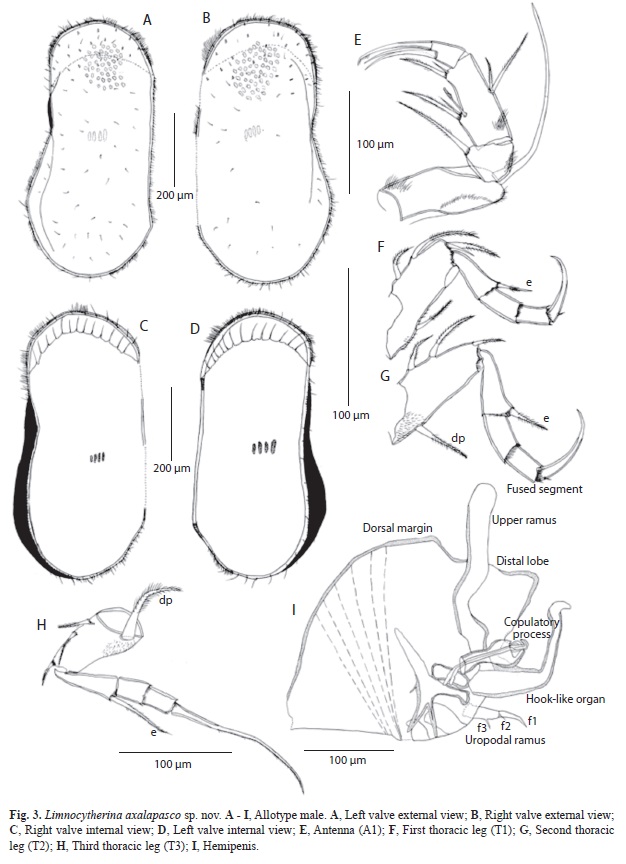
Hard parts: Carapace sexually dimorphic, subrectangular anteriorly and widened posteriorly; marginal pore canals straight. Valves with few nodes, sometimes modified into poor developed alar projections (Fig. 5D). Surface reticulated and covered with setae, and few small, spine-like projections covering surface and margins. Valves slightly asymmetrical, LV is overlapping RV anteriorly. LV (Fig. 3A, D), length 0.66mm, height 0.36mm. Dorsal margin almost straight and covered with short setae. Ventral margin anteriorly straight with a poorly developed flange near medial region, posteriorly widened (maximum wide at 3/4 of valve). Anterior margin rounded with abundant short setae. Selvage peripheral. Posterior margin broadly rounded covered by short setae. RV (Fig. 3B, C), length 0.65mm, height 0.34mm. Dorsal margin straight and covered with setae. Ventral margin straight anteriorly and slightly concave around middle; posteriorly widened. Anterior margin with peripheral selvage and covered with setae. Posterior margin broadly rounded and covered with short setae. Four muscular imprints on shell (Fig. 3 C, D).
Soft parts: A1 and A2 (Fig. 3E), without sexual dimorphism, but on A2 the setae of third segment and the claws of terminal segment seem to be slightly more elongated than in female. Terminal claws length as follows: two subequally long claws 2.6 times longer than terminal segment; third claw is 2.8 times longer than terminal segment. T1 (Fig. 3F) similar to female, but one of setae situated on protopodite being simple, instead of serrulate as in females. Terminal claw strong and longer than in females, 2.7 times longer than terminal segment.T2 (Fig. 3G) similar to female but dp–seta slightly shorter. Distal seta on protopodite and e–seta shorter than in female. T3 (Fig. 3H) 4-segmented and sexually dimorphic, dp–seta swollen, pappose and long; e–seta more elongated than in female and exceeding distal end of terminal segment. Terminal claw elongated, 7.1 times longer than terminal segment and basally not carrying any seta.
Hemipenis (Fig. 3I). Body not very muscular, with distal lobe triangular with blunt tip; not prominent and not articulated. Dorsal margin convex. Ventral margin square-shaped. Hook-like process strong, well sclerotized and elongated, overpassing tip of distal lobe. Upper ramus elongated (tentacle-like appearance), sinuated anteriorly and rectangular medially and distally, tip rounded. Caudal ramus moderately developed with f3 claw-like, setae f2 and f1 short and thin. Copulatory process well sclerotized and distally rounded.
Juveniles and adult variability: On earlier instars (<A-3), anterior part of valves are wider than posterior one. On subsequent instars, anterior and posterior ends becomes subequally wide, and the ventral margin is slightly concave, while dorsal margin slightly convex medially (Fig. 5O-R). In last instars, valves are subrectangular with reticulate surface, often ornamented with bumps and sometimes with poor-developed alar prolongations. Measurements of 100 adult (A) and juveniles (A-6 to A-1) valves contained in the >63μm fraction of surface sediments collected from the littoral zone of lake Alchichica, are shown in figure 4. Besides, table 3 shows the mean length and height, and the standard errors for the different larval stages. Some intraspecific variability was observed in adult specimens: In Alchichica lake, one organism shows the third endite of Mxp inwardly displaced, almost with a hook-like appearance (Fig. 1H), this is probably a deformation of the specimen, but due the highly specialized taxa encountered in this lake, we provided this characteristic for further analysis. Surface valves of specimens from Quechulac lakeare covered by a large number of spine-like projections (Fig. 2I, J), such projections on specimens from the other lakes are scarce and sparsely distributed. Specimens from Quechulac were also bigger than the ones observed in the other two lakes.
Adult measurements:
Female
• Quechulac: Length 0.71±0.004mm and height0.35±0.004mm (n=10).
• La Preciosa: Length 0.67±0.005mm and height0.37±0.003mm (n=9).
• Alchichica: Length 0.64±0.008mm and height0.27±0.004mm (n=6).
Male
• Quechulac: Length 0.75±0.006mm, height0.49±0.003mm (n=8).
• La Preciosa: Length 0.71±0.005mm, height0.40±0.004mm (n=7).
• Alchichica: Length 0.68±0.006mm, height0.36±0.004mm (n=11).
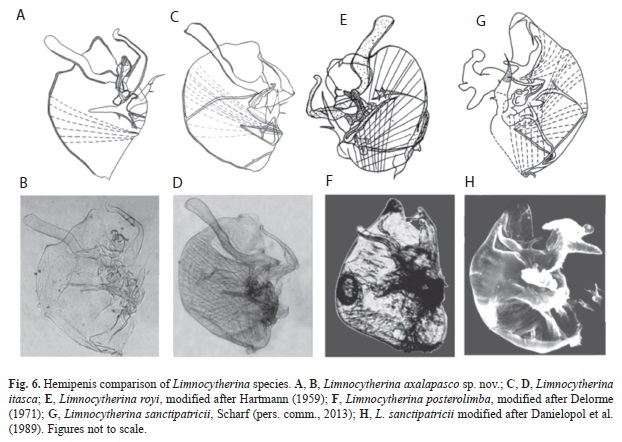
Remarks and affinities: Limnocytherina axalapasco sp. nov. is most similar to the following species: L. posterolimba Delorme, 1967, L. royi (Hartmann, 1959) comb. nov. and L. itasca. All four species have an elongated upper ramus of the hemipenis (Fig. 6), which exceeds lateral and/or distal lobe and they also have a well-developed hook-like process. Their UR is moderately elongated (Cole, 1949; Hartmann, 1959).
Limnocytherina posterolimba (Fig. 6F) differs from L. axalapasco sp. nov. (Fig. 6A, B) mainly in some details of hemipenis. Namely, the upper ramus in the former species is just overpassing the dorsal margin, while in L.axalapasco sp. nov.it exceeds by far the same margin. In addition, the hook-like process is in L. posterolimba inwardly orientated and not reaching tip of the distal lobe, while in L. axalapasco sp. nov. this part is outwardly orientated and overpassing the distal lobe. The valves of L. posterolimba have a depression around the posterior region and a well-developed flange postero-ventrally. Both characteristics absent in L. axalapasco sp. nov.
Limnocytherina itasca (Fig. 6C, D) and L. axalapasco sp. nov. (Fig. 6A, B) also differ in details of the hemipenis morphology. In L. itasca the upper ramus has at its base a small projection, which is more developed than in L. axalapasco sp. nov. The distal lobe in L. Itasca is square-shaped. The hook-like process seems to be articulated and does not reach the tip of the distal lobe. In L. axalapasco sp. nov. the distal lobe is triangular-shaped and the hooklike process (which is not articulated) overpasses the tip of the distal lobe. The f2 seta on the UR is more elongated in L. itasca than in L. axalapasco sp. nov.
Limnocytherina royi comb. nov. (Fig. 6E) and L. axalapasco sp. nov. (Fig. 6A, B) mainly differ in the hemipenis morphology. In the case of L. royi the body of the hemipenis is compressed with the distal lobe almost rounded and with the hook-like process not reaching the tip of this lobe. This species also has an elongated UR, almost reaching the tip of the hook-like process.
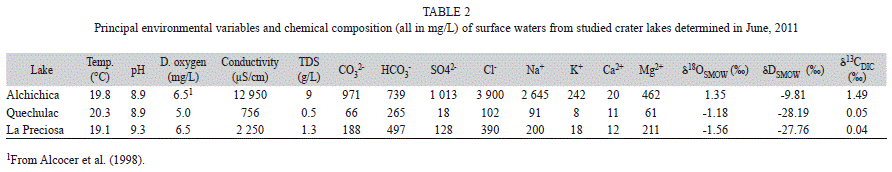
Species habitat description: The location and principal limnological characteristics and environmental variables of the three crater lakes and species habitat are included in tables 1 and 2. In general, the three lakes displayed in June similar temperatures of the surface water (19.1-20.3°C), pH (8.9-9.3) and dissolved oxygen concentrations (5.0-6.5mg/L). Lake Alchichica (characterized by a semi-arid climate and a low annual precipitation <400mm) is the deepest (~64m) and the largest (1.8km2) among the three studied lakes. It is a saline lake, displaying a high conductivity (12950μS/cm) and TDS (9g/L); the water is dominated by chloride, sulfate, carbonate and sodium, magnesium and potassium. Calcium is also present, but in smaller concentrations. Water transparency ranges from 3.6 to 8.0m. Lake sediments are dominated by sand; Ramírez & Vázquez (1988) reported a high percentage (80.5%) of it in the sediments of Alchichica. Gravel, carbonate
and organic matter are also present but in lower percentage. Typical aquatic vegetation includes Ruppia maritima L., diatoms, filamentous chlorophytes and cyanophytes. The last two are particularly abundant and they partially cover the lake bottom (Alcocer, Escobar, Lugo & Peralta, 1998). Lakes Quechulac and La Preciosa are smaller water bodies (≤0.78km2) and are considered freshwater lakes. Their conductivities are lower than those in Alchichica (<2250μS/cm). Quechulac surface waters are dominated by bicarbonate, chloride, carbonate, sodium, magnesium and calcium (Table 2). La Preciosa is dominated by the same anions, however magnesium, sodium and potassium are the dominant cations in surface waters (Table 2). Oxygen, deuterium and carbon isotope values in surface waters of Alchichica are more positive than those from Quechulac and La Preciosa. Stable isotopic composition of surface water is very similar in Quechulac and La Preciosa. The sediments of these lakes are mainly composed of sands (75.6%, 64.9%), less percentage of silts (22.5%, 26.5%) and low concentration of organic matter (3.5%, 5.6%). Aquatic vegetation present in both lakes includes Potamogeton pectinatus L. and Ranunculus cymbalaria Pursh (Ramírez & Vázquez, 1988).
Species ecological preferences: Data indicates that this species inhabits in waters with a broad range of salinity and therefore conductivity (756-12 950μS/cm), as well as the concentrations of CO32-, HCO3-, SO42-, Cl-, Na+, K+ and Mg2+. Calcium content of surface waters where the species was collected ranged from 11 and 20mg/L. The species seems to prefer waters with higher concentration of Cl- or HCO32- and Na+ or Mg2+. The preferred temperature ranges seems to be between 19.1 and 20.3°C, the pH is alkaline (8.9-9.3), and the dissolved oxygen concentration was between 5.0 and 6.5mg/L. Limnocytherina axalapasco sp. nov. was more abundant in the lake Alchichica (239valves/mL in surface sediments) than in lakes Quechulac and La Preciosa. This suggests that the environmental conditions of Alchichica, i.e. more saline, are favorable for this species. More specimens were found in the littoral zone (~0.5m) than in deeper water (Quechulac, 31.5m). This species is benthic and it shows a preference for sandy sediments (≤80.5%). We also found abundant adult carapaces with well-preserved soft parts (15carapaces/mL) in surface sediments from Lake Alchichica during our sampling in June 2011, suggesting that in this lake species reaches adulthood during the summer period. Limnocytherina axalapasco sp. nov. was collected together with cladocerans, chironomids, trichopterans, amphipods, platyhelminthes and thecamoebians. Other ostracod species found with L. axalapasco sp. nov. include Candona sp. and Darwinula stevensoni (Brady & Robertson, 1870).
Discussion
Danielopol et al. (1989) divided the Subfamily Limnocytherinae into four tribes, Limnocytherini, Dinarocytherini, Cytheridellini and Leucocytherini. Martens (1996, 2000) revised the classification of Limnocytherini and suggested that there are in fact three different lineages: Limnocythere-lineage, Paralimnocythere-lineage and Neolimnocythere- lineage. The same author preliminary assigned the following three genera to the Limnocythere-lineage: Limnocythere, Galolimnocythere Schornikov, 1973 and Limnocytherina. Karanovic (2012) highlighted the necessity for a revision of these three genera, because important morphological characters are still unclear in several species (poorly described hemipenis and species known only after females) causing problems in the systematic of these genera. However, Limnocytherina can be distinguished from Limnocythere and Galolimnocythere based on morphology of the hemipenis, in which the characters are well described only in the type species, Limnocytherina sanctipatricii, while poorly known or only inferred in others. After this study, it became clear that on Limnocytherina the hemipenis morphology is varied. One specific Limnocytherina-type of hemipenis, is the one observed in L. axalapasco sp. nov., L. Itasca and L. royi comb. nov. It is characterized by an elongated upper ramus, which also carries a large tentacle-like structure; lower ramus distally composed of lobes, forming a long and thin hook-like process distally outward displaced; and UR moderately enlarged bearing three setae. In comparison with the hemipenis of Limnocytherina sanctipatricii (type species of Limnocytherina), the UR is the only character clearly shared among them. According to Meisch (2000), the enlargement of the UR is one of the most important distinct characters of Limnocytherina, but it needs to be pointed out, that different levels of UR enlargements can also be found in species of closely related genus such as Limnocythere opesta and L. elongata Delachaux, 1928, in which the UR is also moderately developed (unusual feature in Limnocythere). Although the morphology of UR is very important in the determination of species in Limnocytherina, it has to be taken with caution and should be accompanied by other indicators of phylogenetic relationships. Therefore, L. opesta and L. elongate could not be transferred into Limnocytherina because they have some other characteristics on the hemipenis related to Limnocythere such as lower ramus triangular-shaped, strong and short and upper ramus reduced (Delachaux, 1928; Brehem, 1939). Limnocythere neotropica Klie, 1934, could be considered another example of intermediate characters (strongly developed hook-like process, lower and upper reduced and UR developed as a separate structure), which deserves a revision. Furthermore, distinct levels of enlargement of UR could also be observed in the other lineages of the tribe such as Neolimnocythere where the UR is large to very large (Delachaux, 1928) and Paralimnocythere in which the UR is relatively large (Martens, 2000). The entire Limnocytherinae deserve a revision and reevaluation of characters for an accurate classification of the species and also for a clear understanding of its phylogeny.
Based on the current diagnosis of Limnocytherina, here we propose a new combination for Limnocytherina royi (Hartmann, 1959) previously described in Limnocythere. This species is closely related to L. itasca and L. axalapasco sp. nov. (as pointed out above). It has an elongated upper ramus on the hemipenis with a large tentacle-like structure, lower ramus transformed into a hook-like process, and the UR is moderately developed with f3 seta strongly developed. With the description of L. axalapasco sp. nov. and the inclusion of L. royi, the current distribution of the is much wider than considered in the past, because this genus was previously known only from North America. LimnocytherinaLimnocytherina axalapasco sp. nov. is from the transition zone of the continent (Neartic and Neotropical) and also potentially endemic to this region (previous studies of nearby lakes, as well as other water bodies outside this region have not reported it) while Limnocytherina royi is a tropical species known from the Great lake of Nicaragua (Hartmann, 1959). It is expected that intensive sampling of the transition zone and Central America will reveal a number of new Limnocytherina species, hopefully providing important information for understanding the evolution of the Limnocytherinae (Martens, 2000).
Based on the ecological information obtained from the field, L. axalapasco sp. nov. may be considered as a good tool in paleo environmental reconstructions, because this species was collected in a narrow temperature(~19- 20°C), pH (8.9-9.3), and dissolved oxygen (5.0-6.5mg/L) ranges. It also has great potential as a bioindicator for chemical composition of water because it prefers water dominated by Cl- or HCO32- and Na+ or Mg2+. Future studies should focus on collecting ostracod samples at different seasons of the same year, together with the measurement of the environmental variables. Experiments in the laboratory are as well needed to determine the species tolerance to different ecological variables. This will refine our observations and improve the use of this species as a paleo environmental indicator. The analysis of the communities of L. axalapasco sp. nov. from the three crater lakes showed that adult specimens from the saline lake Alchichica are smaller than those from the lake La Preciosa. The largest adults were collected from the lake Quechulac, which also have a large number of small spine-like structures covering the entire shell surface (such number not observed in other specimens). These variability could be preliminary attributed to adaptive responses to specific ecological condition in each ecosystems, i.e. different water chemistry of the lakes and the presence of highly specialized fauna composed of endemic predators such as Poblana alchichica de Buen, 1945, Caecidotea williamsi Escobar-Briones & Alcocer, 2002, Krizousacorixa tolteca Jansson, 1979 (Alcocer & Hammer, 1998; Escobar-Briones & Alcocer, 2002). However, Montiel-Martínez, Cirios-Pérez, Ortega-Mayagoitia & Elías-Gutiérrez (2008) demonstrated that in the saline lake Alchichica, a speciation process has been taking place in copepods; the anterior was highlighted by small morphological variability among populations, as well as, reproductive behavior and molecular data. It is possible that similar speciation process could be occurring in L. axalapasco sp. nov., expressed in the variability observed among populations, but this should be careful further analyzed.
During our sampling in June 2011, one potential reproductive period was observed, only male and female adults were collected in high abundance in this month. Sampling campaigns developed during the other months (December, March), reveal higher counts of A-6 to A-1 instars than adult specimens suggesting, a second reproductive period and it is possible that juvenile specimens live at the sampling location.
Acknowledgments
We thank those people who participated in our field trips, laboratory analyses, and comments: Manuel Elías (ECOSUR, Chetumal Unit), Margarita Caballero, Edyta Zawisza (Instituto de Geofísica, UNAM), Socorro Lozano, Alexander Correa-Metrio, Esperanza Torres (Instituto de Geología, UNAM), María Aurora Armienta (Instituto de Geofísica), Jason Curtis (University of Florida, Gainesville) and Ana Lucía Martínez (Benemérita Universidad de Puebla). We also thank the agencies that provided financial support: CONACYT (project number 167621), National Science Foundation (NSF award number 0902864). We appreciate the constructive comments and drawings provided by Burkhard Scharf, which helped in the improvement of this manuscript. We also appreciate the help of the institution involved in this research: El Colegio de la Frontera Sur, Chetumal Unit, Hanyang University and Universidad Nacional Autónoma de México.
References
Alcocer, J., Escobar, E., Lugo, A., & Peralta, L. (1998). Littoral benthos of the saline crater lakes of the basin of Oriental, Mexico. International Journal of Salt Lake Research, 7, 87-108. [ Links ]
Alcocer, J., & Hammer, U. (1998). Saline lake ecosystems of Mexico. Aquatic Ecosystem Health and Management, 1, 291-315. [ Links ]
Brehm, V. (1939). La fauna microscópica del Lago Petén, Guatemala. Anales de la Escuela Nacional de Ciencias Biológicas, 1, 173-203. [ Links ]
Bridgwater, N., Heaton, T., & O’Hara, S. (1999a). A late Holocene paleolimnological record from central Mexico, based on faunal and stable-isotope analysis of ostracod shells. Journal of Paleolimnology, 22, 383-397. [ Links ]
Bridgwater, N., Holmes, J. A., & O’Hara, S. (1999b). Complex controls on the trace-element chemistry of nonmarine ostracods: an example from Lake Pátzcuaro, central Mexico. Palaeogeography, Palaeoclimatology, Palaeoecology, 148, 117-131. [ Links ]
Chávez-Lara, C., Priyadarsi, R., Caballero, M., Carreño, A. L., & Lakshumanan, C. (2012). Lacustrine ostracodes from the Chihuahuan Desert of Mexico and inferred Late Quaternary paleoecological conditions. Revista Mexicana de Ciencias Geológicas, 29, 422-431. [ Links ]
Cohuo-Durán, S., Elías-Gutiérrez, M., & Karanovic, I. (2013). On three new species of Cypretta Vávra, 1895 (Crustacea: Ostracoda) from the Yucatan Peninsula, Mexico. Zootaxa, 3636, 501-524. [ Links ]
Cole, G. (1949). A New Cytherid Ostracod from Minnesota. Transactions of the American Microscopical Society, 68, 350-354. [ Links ]
Danielopol, D. L., Martens, K., & Casale, L. M. (1989). Revision of the genus Leucocythere Kaufmann, 1892
(Crustacea, Ostracoda, Limnocytheridae), with the description of a new species and two new tribes. Bulletin de L’Institute Royal des Sciences Naturelles de Belgique, 59, 63-94. [ Links ]
Delachaux, T. (1928). Faune invertébrée d’eau douce des hauts plateaux du Pérou. Extrait du Bulletin de la Société Neucháteloise des Sciences Naturelles, Nouv Sér, 1, 45-77. [ Links ]
Delorme, L. (1971). Freshwater ostracodes of Canada. Part V: Families Limnocytheridae, Loxoconchidae. Canadian Journal of Zoology, 49, 43-64. [ Links ]
Escobar-Briones, E., & Alcocer, J. (2002). Caecidotea williamsi (Crustacea: Isopoda: Asellidae), a new species from a saline crater-lake in the eastern Mexican Plateau. Hydrobiologia, 477, 93-105. [ Links ]
Folmer, O., Black, M., Hoeh, W., Lutz, R., & Vrijenhoek, R. (1994). DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Molecular Marine Biology and Biotechnology, 3, 294-299. [ Links ]
Forester, R. (1985). Limnocythere bradburyi n. sp.: A Modern Ostracode from Central Mexico and a Possible Quaternary Paleoclimatic Indicator. Journal of Paleontology, 59, 8-20. [ Links ]
Furtos, N. (1936). On the Ostracoda from the cenotes of Yucatan and vicinity. Publ. Carnegie Institution of Washington, 457, 89-115. [ Links ]
Furtos, N. (1938). A new species of Cypridopsis from Yucatán. Publ. Carnegie Institution of Washington, 491, 155-157. [ Links ]
Garm, A. (2004). Revising the definition of the crustacean seta and setal classification systems based on examinations of the mouthpart setae of seven species of decapods. Zoological Journal of the Linnean Society, 142, 233-252. [ Links ]
Hartmann, G. (1959). Beitrag zur Kenntnis des Nicaragua- Sees unter besonderer Berücksichtigung seiner Ostracoden (mit Beschreibung von 5 neuen Arten). Zoologischer Anzeiger, 162, 269-294. [ Links ]
Juárez, D. (2005). Registro de cambios paleoambientales en sedimentos del lago “La Preciosa”, Puebla, con base en el estudio de ostrácodos. (M.Sc. unpublished thesis). Universidad Nacional Autónoma de México, México. [ Links ]
Karanovic, I. (2012). Recent freshwater Ostracods of the World. Crustacea, Ostracoda, Podocopida. Germany: Springer-Verlag Berlin and Heidelberg. [ Links ]
Kaźmierczak, J., Kempe, S., Kremer, B., López-García, P., Moreira, D., & Taver, R. (2011). Hydrochemistry and microbialites of the alkaline crater lake Alchichica, Mexico. Facies, 57 (4), 543-570. [ Links ]
Klie, W. (1934). Zur Kenntnis der Ostracoden-Gattung Limnocythere. Arch Naturgesch, Z Wiss Zool, Abt B, 3, 534-544. [ Links ]
Maddocks, R., Machain-Castillo, M., & Gío-Argáez, F. (2009). Podocopan Ostracoda (Crustacea) of the Gulf of Mexico. In D. Felder & D. K. Camp (Eds.). Gulf of Mexico–Origins, Waters, and Biota. Biodiversity (p. 877-894). Texas A & M Press, College. [ Links ]
Martens, K. (1990). Revision of African Limnocythere s.s. Brady, 1867 (Crustacea,Ostracoda), with special reference to the Rift Valley Lakes: morphology, taxonomy, evolution and (paleo-) ecology. Archiv für Hydrobiologie, Supplement 83, 453-524. [ Links ]
Martens, K. (1996). On Korannocythere gen. nov. (Crustacea, Ostracoda), a new genus of temporary pool limnocytherids from southern Africa, with the description of three new species and a generic reassessment of the Limnocytherinae. Bulletin de L’Institut Royal Des Sciences Naturelles de Belgique, 66, 51-72. [ Links ]
Martens, K. (1998). General morphology of non-marine Ostracoda. In K. Martens (Ed.). Sex and Parthenogenesis. Evolutionary Ecology of Reproductive Modes in Non-marine Ostracods (p. 57-75). Leiden,
Backhuys Publ. [ Links ]
Martens, K. (2000). Factors affecting the divergence of mate recognition systems in the Limnocytherinae (Crustacea, Ostracoda). Hydrobiologia, 419, 83-101. [ Links ]
Martens, K., Rossetti, G., Butlin, R., & Schön, I. (2005). Molecular and morphological phylogeny of the ancient asexual Darwinulidae (Crustacea, Ostracoda). Hydrobiologia, 538, 153-165. [ Links ]
Martens, K., & Savatenalinton, S. (2011). A subjective checklist of the Recent, free-living, non-marine Ostracoda (Crustacea), Zootaxa, 2855, 1-79. [ Links ]
McKenzie, K., & Swain, F. M. (1967). Recent Ostracoda from Scammon Lagoon, Baja California. Journal of Paleontology, 41, 281-305. [ Links ]
Meisch, C. (2000). Freshwater Ostracoda of Western and Central Europe. Germany: Spektrum Akademischer Verlag, Heidelberg, Berlin. [ Links ]
Montiel-Martínez, A., Ciros-Pérez, J., Ortega-Mayagoitia, E., & Elías-Gutiérrez, M. (2008). Morphological, ecological, reproductive and molecular evidence for Leptodiaptomus garciai (Osorio-Tafall 1942) as a valid endemic species. Journal of Plankton Research, 30, 1079-1093. [ Links ]
Palacios-Fest, M., Carreño, A. L., Ortega-Ramírez, J., & Alvarado-Valdez, G. (2002). A paleoenvironmental reconstruction of Laguna Babícora, Chihuahua, Mexico based on ostracode paleoecology and trace element shell chemistry. Journal of Paleolimnology, 27, 185-206. [ Links ]
Pérez, L., Lorenschat, J., Brenner, M., Scharf, B., & Schwalb, A. (2010a). Extant freshwater ostracodes (Crustacea: Ostracoda) from Lago Petén Itzá, Guatemala. Revista de Biología Tropical, 58 (3), 871-895. [ Links ]
Pérez, L., Lorenschat, J., Bugja, R., Brenner, M., Scharf , B., & Schwalb, A. (2010b). Distribution, diversity and ecology of modern freshwater ostracodes (Crustacea), and hydrochemical characteristics of Lago Petén Itzá, Guatemala. Journal of Limnology, 69 (1), 146-159. [ Links ]
Pérez, L., Frenzel, P., Brenner, M., Escobar, E., Hoelzmann, P., Scharf, B., & Schwalb, A. (2011). Late Quaternary (24-10 ka BP) environmental history of the Neotropical lowlands inferred from ostracodes in sediments of Lago Petén Itzá, Guatemala. Journal of Paleolimnology, 46, 59-74. [ Links ]
Pérez, L., Lorenschat, J., Brenner, M., Scharf, B. & Schwalb, A. (2012). Non-marine ostracodes (Crustacea) of Guatemala. In E. Cano (Ed.). Biodiversidad de Guatemala, Vol 2. (pp. 121-131). Guatemala: Universidad del Valle de Guatemala. [ Links ]
Ramírez, P., & Vázquez, F. (1988). Contribuciones al estudio limnobotánico de la zona litoral de seis lagos cráter del estado de Puebla. Anales del Instituto de Ciencias del Mar y Limnología, Contribución No. 640. Universidad Nacional Autónoma de México. [ Links ]
Schön, I. (2001). PCR Primers and Conditions for Non-Marine Ostracods. Bio Techniques, 31, 1012-1020. [ Links ]
Stock, J., & Von Vaupel Klein, J. V. (1996). Mounting media revisited: The suitability of Reyne’s fluid for small crustaceans. Crustaceana, 69, 794-798. [ Links ]
Swain, F. (1967). Ostracoda from the Gulf of California. Geological Society of America, Memories 101. [ Links ]
Tressler, W. (1954). Fresh-water Ostracoda from Texas and Mexico. Journal of Washington Academy Science, 44, 138-149. [ Links ]
Alcocer, J., & Hammer, U. (1998). Saline lake ecosystems of Mexico. Aquatic Ecosystem Health and Management, 1, 291-315. [ Links ]
Brehm, V. (1939). La fauna microscópica del Lago Petén, Guatemala. Anales de la Escuela Nacional de Ciencias Biológicas, 1, 173-203. [ Links ]
Bridgwater, N., Heaton, T., & O’Hara, S. (1999a). A late Holocene paleolimnological record from central Mexico, based on faunal and stable-isotope analysis of ostracod shells. Journal of Paleolimnology, 22, 383-397. [ Links ]
Bridgwater, N., Holmes, J. A., & O’Hara, S. (1999b). Complex controls on the trace-element chemistry of nonmarine ostracods: an example from Lake Pátzcuaro, central Mexico. Palaeogeography, Palaeoclimatology, Palaeoecology, 148, 117-131. [ Links ]
Chávez-Lara, C., Priyadarsi, R., Caballero, M., Carreño, A. L., & Lakshumanan, C. (2012). Lacustrine ostracodes from the Chihuahuan Desert of Mexico and inferred Late Quaternary paleoecological conditions. Revista Mexicana de Ciencias Geológicas, 29, 422-431. [ Links ]
Cohuo-Durán, S., Elías-Gutiérrez, M., & Karanovic, I. (2013). On three new species of Cypretta Vávra, 1895 (Crustacea: Ostracoda) from the Yucatan Peninsula, Mexico. Zootaxa, 3636, 501-524. [ Links ]
Cole, G. (1949). A New Cytherid Ostracod from Minnesota. Transactions of the American Microscopical Society, 68, 350-354. [ Links ]
Danielopol, D. L., Martens, K., & Casale, L. M. (1989). Revision of the genus Leucocythere Kaufmann, 1892
(Crustacea, Ostracoda, Limnocytheridae), with the description of a new species and two new tribes. Bulletin de L’Institute Royal des Sciences Naturelles de Belgique, 59, 63-94. [ Links ]
Delachaux, T. (1928). Faune invertébrée d’eau douce des hauts plateaux du Pérou. Extrait du Bulletin de la Société Neucháteloise des Sciences Naturelles, Nouv Sér, 1, 45-77. [ Links ]
Delorme, L. (1971). Freshwater ostracodes of Canada. Part V: Families Limnocytheridae, Loxoconchidae. Canadian Journal of Zoology, 49, 43-64. [ Links ]
Escobar-Briones, E., & Alcocer, J. (2002). Caecidotea williamsi (Crustacea: Isopoda: Asellidae), a new species from a saline crater-lake in the eastern Mexican Plateau. Hydrobiologia, 477, 93-105. [ Links ]
Folmer, O., Black, M., Hoeh, W., Lutz, R., & Vrijenhoek, R. (1994). DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Molecular Marine Biology and Biotechnology, 3, 294-299. [ Links ]
Forester, R. (1985). Limnocythere bradburyi n. sp.: A Modern Ostracode from Central Mexico and a Possible Quaternary Paleoclimatic Indicator. Journal of Paleontology, 59, 8-20. [ Links ]
Furtos, N. (1936). On the Ostracoda from the cenotes of Yucatan and vicinity. Publ. Carnegie Institution of Washington, 457, 89-115. [ Links ]
Furtos, N. (1938). A new species of Cypridopsis from Yucatán. Publ. Carnegie Institution of Washington, 491, 155-157. [ Links ]
Garm, A. (2004). Revising the definition of the crustacean seta and setal classification systems based on examinations of the mouthpart setae of seven species of decapods. Zoological Journal of the Linnean Society, 142, 233-252. [ Links ]
Hartmann, G. (1959). Beitrag zur Kenntnis des Nicaragua- Sees unter besonderer Berücksichtigung seiner Ostracoden (mit Beschreibung von 5 neuen Arten). Zoologischer Anzeiger, 162, 269-294. [ Links ]
Juárez, D. (2005). Registro de cambios paleoambientales en sedimentos del lago “La Preciosa”, Puebla, con base en el estudio de ostrácodos. (M.Sc. unpublished thesis). Universidad Nacional Autónoma de México, México. [ Links ]
Karanovic, I. (2012). Recent freshwater Ostracods of the World. Crustacea, Ostracoda, Podocopida. Germany: Springer-Verlag Berlin and Heidelberg. [ Links ]
Kaźmierczak, J., Kempe, S., Kremer, B., López-García, P., Moreira, D., & Taver, R. (2011). Hydrochemistry and microbialites of the alkaline crater lake Alchichica, Mexico. Facies, 57 (4), 543-570. [ Links ]
Klie, W. (1934). Zur Kenntnis der Ostracoden-Gattung Limnocythere. Arch Naturgesch, Z Wiss Zool, Abt B, 3, 534-544. [ Links ]
Maddocks, R., Machain-Castillo, M., & Gío-Argáez, F. (2009). Podocopan Ostracoda (Crustacea) of the Gulf of Mexico. In D. Felder & D. K. Camp (Eds.). Gulf of Mexico–Origins, Waters, and Biota. Biodiversity (p. 877-894). Texas A & M Press, College. [ Links ]
Martens, K. (1990). Revision of African Limnocythere s.s. Brady, 1867 (Crustacea,Ostracoda), with special reference to the Rift Valley Lakes: morphology, taxonomy, evolution and (paleo-) ecology. Archiv für Hydrobiologie, Supplement 83, 453-524. [ Links ]
Martens, K. (1996). On Korannocythere gen. nov. (Crustacea, Ostracoda), a new genus of temporary pool limnocytherids from southern Africa, with the description of three new species and a generic reassessment of the Limnocytherinae. Bulletin de L’Institut Royal Des Sciences Naturelles de Belgique, 66, 51-72. [ Links ]
Martens, K. (1998). General morphology of non-marine Ostracoda. In K. Martens (Ed.). Sex and Parthenogenesis. Evolutionary Ecology of Reproductive Modes in Non-marine Ostracods (p. 57-75). Leiden,
Backhuys Publ. [ Links ]
Martens, K. (2000). Factors affecting the divergence of mate recognition systems in the Limnocytherinae (Crustacea, Ostracoda). Hydrobiologia, 419, 83-101. [ Links ]
Martens, K., Rossetti, G., Butlin, R., & Schön, I. (2005). Molecular and morphological phylogeny of the ancient asexual Darwinulidae (Crustacea, Ostracoda). Hydrobiologia, 538, 153-165. [ Links ]
Martens, K., & Savatenalinton, S. (2011). A subjective checklist of the Recent, free-living, non-marine Ostracoda (Crustacea), Zootaxa, 2855, 1-79. [ Links ]
McKenzie, K., & Swain, F. M. (1967). Recent Ostracoda from Scammon Lagoon, Baja California. Journal of Paleontology, 41, 281-305. [ Links ]
Meisch, C. (2000). Freshwater Ostracoda of Western and Central Europe. Germany: Spektrum Akademischer Verlag, Heidelberg, Berlin. [ Links ]
Montiel-Martínez, A., Ciros-Pérez, J., Ortega-Mayagoitia, E., & Elías-Gutiérrez, M. (2008). Morphological, ecological, reproductive and molecular evidence for Leptodiaptomus garciai (Osorio-Tafall 1942) as a valid endemic species. Journal of Plankton Research, 30, 1079-1093. [ Links ]
Palacios-Fest, M., Carreño, A. L., Ortega-Ramírez, J., & Alvarado-Valdez, G. (2002). A paleoenvironmental reconstruction of Laguna Babícora, Chihuahua, Mexico based on ostracode paleoecology and trace element shell chemistry. Journal of Paleolimnology, 27, 185-206. [ Links ]
Pérez, L., Lorenschat, J., Brenner, M., Scharf, B., & Schwalb, A. (2010a). Extant freshwater ostracodes (Crustacea: Ostracoda) from Lago Petén Itzá, Guatemala. Revista de Biología Tropical, 58 (3), 871-895. [ Links ]
Pérez, L., Lorenschat, J., Bugja, R., Brenner, M., Scharf , B., & Schwalb, A. (2010b). Distribution, diversity and ecology of modern freshwater ostracodes (Crustacea), and hydrochemical characteristics of Lago Petén Itzá, Guatemala. Journal of Limnology, 69 (1), 146-159. [ Links ]
Pérez, L., Frenzel, P., Brenner, M., Escobar, E., Hoelzmann, P., Scharf, B., & Schwalb, A. (2011). Late Quaternary (24-10 ka BP) environmental history of the Neotropical lowlands inferred from ostracodes in sediments of Lago Petén Itzá, Guatemala. Journal of Paleolimnology, 46, 59-74. [ Links ]
Pérez, L., Lorenschat, J., Brenner, M., Scharf, B. & Schwalb, A. (2012). Non-marine ostracodes (Crustacea) of Guatemala. In E. Cano (Ed.). Biodiversidad de Guatemala, Vol 2. (pp. 121-131). Guatemala: Universidad del Valle de Guatemala. [ Links ]
Ramírez, P., & Vázquez, F. (1988). Contribuciones al estudio limnobotánico de la zona litoral de seis lagos cráter del estado de Puebla. Anales del Instituto de Ciencias del Mar y Limnología, Contribución No. 640. Universidad Nacional Autónoma de México. [ Links ]
Schön, I. (2001). PCR Primers and Conditions for Non-Marine Ostracods. Bio Techniques, 31, 1012-1020. [ Links ]
Stock, J., & Von Vaupel Klein, J. V. (1996). Mounting media revisited: The suitability of Reyne’s fluid for small crustaceans. Crustaceana, 69, 794-798. [ Links ]
Swain, F. (1967). Ostracoda from the Gulf of California. Geological Society of America, Memories 101. [ Links ]
Tressler, W. (1954). Fresh-water Ostracoda from Texas and Mexico. Journal of Washington Academy Science, 44, 138-149. [ Links ]
*Correspondencia a:
Sergio Cohuo-Durán. El Colegio de la frontera sur, Av. Centenario Km 5.5, 77014, Chetumal, Quintana Roo, Mexico. Correo electrónico Sergiocd@comunidad.unam.mx
Liseth Pérez. Instituto de Geología, Universidad Nacional Autónoma de México, Ciudad Universitaria, 04510, Distrito Federal, México. Correo electrónico lcpereza@geologia.unam.mx
Ivana Karanovic. Hanyang University, Department of Life Science, Colleague of Natural Sciences, 17 Haengdang-dong, Seongdonggu, Seoul 133-791, Korea; Institute of Marine and Antarctic Studies, University of Tasmania, Private Bag 49, 7001, Hobart, Tasmania, Australia. Correo electrónico ivana.Karanovic@utas.edu.au
1. El Colegio de la frontera sur, Av. Centenario Km 5.5, 77014, Chetumal, Quintana Roo, Mexico. Correo electrónico Sergiocd@comunidad.unam.mx
2. Instituto de Geología, Universidad Nacional Autónoma de México, Ciudad Universitaria, 04510, Distrito Federal, México. Correo electrónico lcpereza@geologia.unam.mx
3. Hanyang University, Department of Life Science, Colleague of Natural Sciences, 17 Haengdang-dong, Seongdonggu, Seoul 133-791, Korea; Institute of Marine and Antarctic Studies, University of Tasmania, Private Bag 49, 7001, Hobart, Tasmania, Australia. Correo electrónico ivana.Karanovic@utas.edu.au
Received 12-III-2013. Corrected 10-VIII-2013. Accepted 11-IX-2013.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}