










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.62 n.2 San José Apr./Jun. 2014
Histology of the regeneration of Paulownia tomentosa (Paulowniaceae) by organogenesis
Histología de la regeneración por organogésis en Paulownia tomentosa (Paulowniaceae)
Histología de la regeneración por organogésis en Paulownia tomentosa (Paulowniaceae)
*Dirección para correspondencia:
Abstract
Paulownia tomentosa is a fast-growing tree species with a considerable economic potential because of its value for wood as well as its high biomass production, and elevated stress tolerance. The objective of the present study was to evaluate the development of adventitious buds in leaves obtained from four-week-old shoots of P. tomentosa, in order to identify the cells involved in in vitro adventitious bud development. Leaves (proximal halves with the petiole) from the first node were excised from four-week-old micropropagated shoots, and cultured on Murashige and Skoog medium, supplemented with 3% (w/v) sucrose, 0.6% (w/v) Sigma agar, 22.7µM thidiazuron (TDZ) and 2.9µM indole-3-acetic acid for two weeks, explants were then transferred to the same medium with 0.44µM N6-benzyladenine for another four weeks. Five explants were collected daily during the two first weeks in TDZ treatment. A total of 140 samples were processed. Most of the buds developed indirectly from the callus formed in the petiole stub, and they became visible after eight-ten days of culture, although some buds were also observed in the area of the laminar cut at the level of the veins. The first histological changes could be observed after two-three days of culture, with the dedifferentiation of some subepidermal and inner parenchyma cells, which exhibited a large, prominent nucleus, densely-stained cytoplasm and a high nucleusto-cell area ratio. Proliferation of these cells gives rise to meristemoid formation after seven-ten days of culture. Organized cell division in meristemoids allows the formation of bud primordia that emerged from the explants surface. The progressive structural differentiation of the apical meristem, leaf primordia, and procambium strands, led to formation of complete buds that were observed in the exterior of the explants after 10-15 days of culture. Direct development of buds from cells in the subepidermic and/or epidermic layers were observed on the adaxial surface of the petiole. This protocol may be a useful tool for the application of genetic transformation techniques, as it enables to determine specific regions in the foliar explants where the meristemoids formation will take place, and therefore to determine which cells should be the object of genetic transformation.
Key words: bud induction, histological analysis, organogenesis, Paulownia tomentosa, TDZ.
Resumen
Paulownia tomentosa es un árbol de rápido crecimiento y con un gran potencial económico por su madera, su utilización para la producción de biocombustible, así como su alto rendimiento en la producción de biomasa y su elevada tolerancia al estrés. El objetivo del presente trabajo ha sido evaluar el desarrollo a nivel histológico de yemas adventicias en hojas de Paulownia tomentosa. Hojas del primer entrenudo de brotes de cuatro semanas cultivados in vitro, fueron cultivadas en medio de Murashige y Skoog complementado con 22.7µM tidiazuron y 2.9µM ácido indol acético durante dos semanas. Los explantos fueron posteriormente transferidos a igual medio con 0.44µM N6 –benciladenina durante otras cuatro semanas. Se recogieron cinco muestras diarias durante las dos primeras semanas de tratamiento en medio con TDZ, procesando un total de 140 muestras. La mayoría de las yemas se desarrollan indirectamente a partir del callo formado en la superficie de corte del pecíolo. Después de dos-tres días de cultivo se observan los primeros cambios histológicos, con la desdiferenciación de algunas células de las capas subepidérmicas y del parénquima interno. La posterior proliferación de estas células da lugar a la formación de los meristemoides después de siete-diez días de cultivo. La progresiva diferenciación de estos meristemoides da lugar a la formación de las yemas que son visibles al exterior a partir de los 10-15 días. En la superficie adaxial del pecíolo se observó la formación de yemas adventicias de forma directa. Este protocolo puede ser de gran utilidad para la determinación de las células más adecuadas para los procesos de transformación genética.
Palabras clave: histología, inducción de yemas, organogénesis, Paulownia tomentosa, TDZ.
The search for new sources of energy is currently of great interest. One potentially promising option is the use of plant biomass or production of biofuels like bioethanol and biodiesel, which are renewable and do not contribute to climate change (Mandpe, Kadlaskar, Degen, & Keppeler, 2005). Oil-based feedstock or biodiesel can be produced from vegetable oils obtained from agricultural plants such as rapeseed, sunflower, soybean, oil palm and groundnut (Johnson, Eswaran, & Sujatha, 2011). Bioethanol, with features similar to gasoline, is obtained from the fermentation of sugars from sugar cane, beet or cereals such as wheat, corn, or barley (Balat, Balat, & Öz, 2008). However, biofuels should be obtained from primary materials that are not also used as foodstuff, to prevent the price of such materials increasing and to make use of soils that would otherwise not be used for agricultural purposes. In this context, lignocellulosic biomass materials constitute a substantial renewable substrate for bioethanol production that do not compete with food production and animal feed (Limayem & Ricke, 2012). Paulownia tomentosa Steud., a lignocellulosic energy crop, is used to produce electricity, although species of this genus are being considered for the production of biofuels (López, Pérez, Zamudio, De Alva, & García, 2012). This species is becoming economically important because of its marketable value for wood and high biomass production as a result of its rapid growth in a wide variety of soil types, as well as its resistance to pathogens, and elevated stress tolerance (Bergmann, 1998; Corredoira, Ballester, & Vieitez, 2008; Doumett et al., 2008). Paulownia is also useful because of its high transpiration rates, widespread root system, and elevated tolerance to high concentrations of metals in both hydroponic and field studies, and is a good candidate for the phytoremediation of polluted soils (Doumett et al., 2008). The species is also used for the landreclamation of nutrient-poor soils (Marcotrigiano & Jagannathan, 1988), as a fast growing ornamental tree (Castellanos-Hernández et al., 2009), and its leaves and flowers can be used for medicinal purposes, they are rich in nitrogen, serving as good fertilizer and fodder (Zhu, Chao, Lu, & Xiong, 1986).
A prerequisite for genetic improvement of this biofuel feedstock plant (Kausch et al., 2010) is the establishment of an efficient transformation system. The use of transgenic tools to improve plant feedstock are required in order to diversify the energy sources, and to obtain plants that produce cellulases or ligninases, plants with lower lignin content or with increased biomass, suitable for producing biofuel (Beltrán, 2008). Although in vitro regenerated plants of P. tomentosa have been obtained by propagation of axillary shoots (Burger, 1989; Song, Sato, Saito, & Kihachiro, 1989) and organogenesis (Rao, Goh, & Kumar, 1996; Yang, Ho, Chen, & Chang, 1996; Bergmann & Moom, 1997; Corredoira et al., 2008), these regeneration systems have not been developed into efficient genetic transformation protocols. Attempts in our laboratory to transform Paulownia tissues by kanamycin selection have produced transgenic calluses, but have failed to produce regenerated transgenic plants. The major prerequisite for genetic transformation through Agrobacterium tumefaciens is the availability of a reliable plant regeneration system and a suitable method of transformation (Chateau, Sangwan, & Sanhgwan-Norreel, 2000; Chovelon, Restier, Giovinazzo, Dogimont, & Aarrouf, 2011). An understanding of the regeneration process and the identification of the optimal target tissue is essential for the success of genetic engineering techniques. It is therefore very important to locate and identify the cells involved in and responsible for in vitro plant organogenesis. Despite the large number of reports on in vitro plant regeneration of Paulownia species via organogenesis, the scant histological data available does not provide a good understanding of the process. The purpose of the present study was therefore to identify the cells involved in in vitro adventitious bud development in leaf explants derived from a mature P. tomentosa tree. This may lead to a better understanding of in vitro development and may be particularly useful for micropropagation and genetic transformation of this species.
Material and Methods
Plant material and culture conditions: Crown branches of a 17-year-old P. tomentosa tree were collected in May and cut into 25-30cm segments. The branch segments were forced to flush by placing them upright in flats of moistened perlite in a growth chamber at 25ºC and 90% relative humidity under a 16-h photoperiod. After three weeks the flushed shoots were surface sterilized and nodal segments and shoot tips were inoculated in Murashige & Skoog medium (MS) (1962) supplemented with 30g/L sucrose, 6g/L Sigma agar (basal medium) and 8.9µM N6-benzyladenine (BA). The development shoot were multiplied by axillary shoot development on basal medium supplemented with 0.88µM BA (proliferation medium) (Corredoira et al., 2008). In vitro shoots were maintained by subcultures every four weeks on a shoot proliferation medium. Leaves from the first node were excised from four-week-old micropropagated shoots and were cut transversally across the midvein. The proximal halves, with two-three mm of the petiole attached, were then placed in 90x150mm Petri dishes (10 explants per dish) containing 25mL of basal medium supplemented with 22.7μM thidiazuron (TDZ) and 2.9μM indol-3-acetic acid (IAA) for two weeks, and then transferred to medium with 0.44μM BA for another four weeks. All culture media were brought to pH 5.6 before autoclaving at 121ºC for 20min.
The cultures were maintained in a climatized growth chamber with photoperiodic lighting. White light of radiant flux density of 30μmol/m2.s was provided by fluorescent tubes (Mazdafluor 7D TF 36w/LJ), for a period of 16h. The temperature was maintained at 25ºC
during the 16 hours of light, and at 20ºC during the eight hours of darkness.
Histological procedure: The proximal halves of the foliar explants were collected daily during the two week period in culturemedium containing TDZ. Five samples were collected at each sampling time, and a total of 70 samples were processed. The experiment was repeated twice. Explants were fixed in FAA solution (formalin:acetic acid:ethanol 50, 1:1:18 (v/v/v)) for 48h at room temperature. The fixed tissue was dehydrated in agraded series of n-butanol (Jensen, 1962). The dehydrated tissue was infiltrated by transfer to paraffin wax: n-butanol (1:1, v/v) at 58ºC for 48h, and then to 100% paraffin wax at 58ºC for 48h. The paraffin wax infiltrated tissue was transferred to metal embedding moulds with melted paraffin wax, and the tissue sample was solidified at room temperature. Sections (10μm) were cut on a Reichert-Jung rotary microtome, and were later stained with safranin-fast green (Jensen, 1962) or with PAS-naphthol blue-black (O´Brien & McCully, 1981). The stained sections were mounted with Euckit® and the photomicrographs were taken with a Nikon-FXA microscope equipped with an Olympus DP71 digital camera. Macroscopic features were observed in a stereo microscope (Olympus SZX9) and photographed with an Olympus DP10 digital camera.
Results
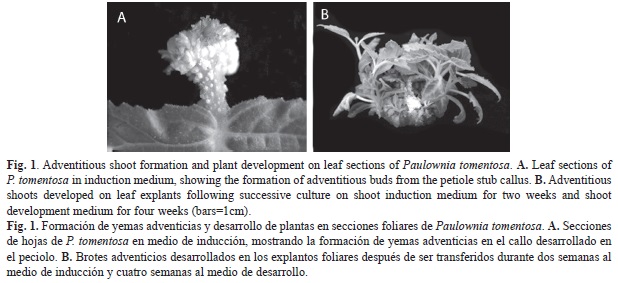
Morphological appearance: First morphological changes in the leaves were observed after six days, when the petioles began to swell and form a small callus on the cut surface. Most adventitious buds developed from the wound callus formed in the petiole stub, although some appeared in the leaf lamina, generally in association with the veins. Bud primordial became visible after eight-ten days of culture on medium with TDZ and IAA (Fig. 1A). The reddish-coloured buds were elongated on transfer to medium containing 0.44μM BA (Fig. 1B). As most of the buds appeared in the region of the petiole, histological study of the caulogenic process was focused on this area.
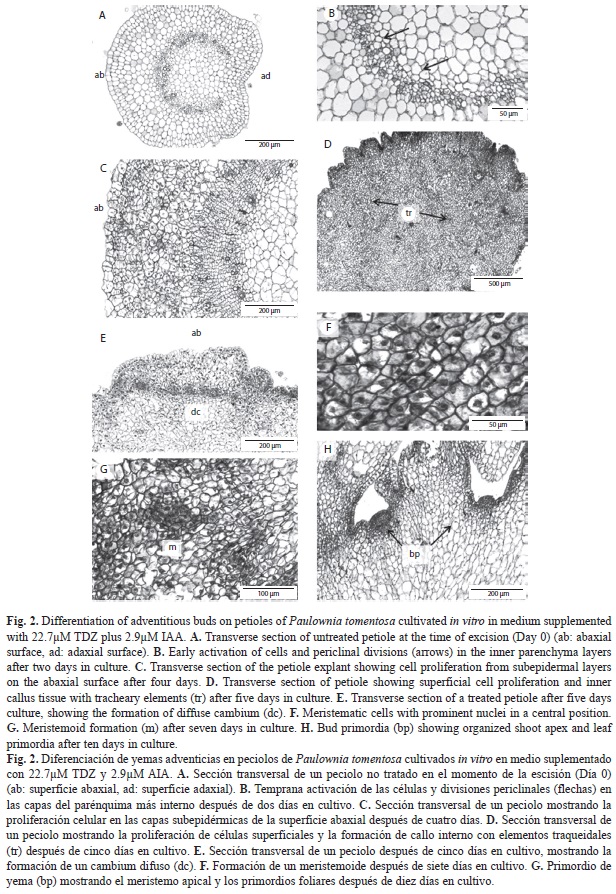
Histological study: Anatomical observations of transverse sections on zero day showed the typical dicotyledonous petiole-like structure, including the epidermis, formed by a layer of isodiametric cells with numerous trichomes, the subepidermis, and the inner parenchyma, composed by large highly vacuolated cells with the nuclei adjoining the cell walls, and with intercellular spaces between these cells. The outermost and innermost of the inner parenchyma cells were smaller than those in the intermediate portion, and were more densely packed. Internally, the vascular bundles were arranged in a continuous arc open towards the adaxial side of the petiole where additional bundles were also present. The petiolar bundles were collateral with the phloem external to the cambium, with discrete phloem fibers, and the xylem internal to the cambium, composed of cells with characteristic thickened walls (Fig. 2A).
Abaxial surface of the petiole: The morphogenetic response began shortly after initiation of the culture in medium containing 22.7µM TDZ and 2.9µM IAA. After two-three days in culture, histological sections revealed cell activation or dedifferentiation of some subepidermal cells and those of the inner parenchyma cells close to the vascular system. These cells showed prominent centrally-located nuclei with evident nucleoli. At this time, the first cell divisions were observed, mainly in the periclinal plane (Fig. 2B). Some anticlinal divisions also occurred in the epidermal cells. These changes mainly took place at the cut surface of the cut petiole, and were less pronounced in internal regions of the explant.
The continual, mainly periclinal, divisions that occurred on the following days (three-five days after culture initiation) in the subepidermis and the outermost zones of the inner parenchyma gave rise to the formation of callus, in which clusters of cells that shared a common cell wall can be observed (Fig. 2C). Proliferation of these cells gave the surface of the petiole a wavy appearance. Some anticlinal and oblique divisions, which increased the thickness of the explants, were also observed. Cell divisions occurred in different planes in the innermost zones of the inner parenchyma closest to the vascular bundles, giving rise to a cambial-type meristem, which led to formation of a type of callus in which numerous tracheary elements were differentiated (Fig. 2D).
Division of the new cells derived from the subepidermal layer led to the formation of rows of cells perpendicular to the surface of the explants. This region may be considered as a type of secondary meristem or diffuse cambium, and was evident on the abaxial surface of the petiole in contact with the culture medium (Fig. 2E). The cells in this region were rectangular and smaller than those from which they were derived; they were also vacuolated, with prominent laterally or centrally positioned nuclei. After five-seven days, some of the cells from this diffuse cambium began to dedifferentiate and acquired the appearance of meristem cells. In transverse sections, these cells were isodiametric, smaller than surrounding cells, with dense cytoplasm, small vacuoles, and a conspicuous nuclei, with prominent nucleoli, in a central position (Fig. 2F). These meristematic cells were characterized by a high ratio between the surface area of the nucleus and the cell. The dedifferentiation appeared to be associated with anticlinal divisions in cells that previously divided periclinally. The differentiation continued on the following days (seven to ten), giving rise to the formation of small groups of meristematic cells (meristemoids) with divisions in different planes (Fig. 2G). Meristemoids were placed in the outermost zones of the tissue derived from the subepidermal layer. Meristemoids gradually increased in size, forming more or less polarized structures (Fig. 2H). The meristemoids gradually developed into bud primordia by organised cell divisions and differentiation of a protodermis. Procambial bundles, which established connections with the adjacent vascular tissue formed within the callus, developed by around day 15. Structural differentiation of the apical meristem and the foliar primordia enabled formation of complete adventitious buds after 15-20 days in media containing TDZ. The formation of adventitious buds was an asynchronous process, and well formed buds were observed along with meristemoids and groups of fused buds and abnormal structures.
Adaxial surface of the petiole: Unlike on the abaxial surface of the petiole, differentiation of a diffuse cambium did not take place, and adventitious buds developed directly from the cells present in the explant. After two-three days culture in medium containing 22.7µM TDZ and 2.9µM IAA, cells in the subepidermic layer became activated. At this time, cells with prominent nuclei were observed, along with the first divisions mainly in the periclinal plane (Fig. 3A). Some divisions were also observed in epidermal cells. Gradual differentiation of these cells led to the formation of groups of cells perpendicular to the surface of the explant, and to the formation of meristemoids after five-seven days in culture (Fig. 3B). The gradual organization of these meristemoids gave rise to formation of the bud primordial (Fig. 3C). Their later development gave rise to the formation of adventitious buds, which were visible after ten days in culture.
Discussion
The development of a breeding programme associated with the biotechnological tools depends on the establishment of an efficient in vitro plant regeneration and transformation system (Chovelon et al., 2011). However, the application of such biotechnological approaches has not been possible for P. tomentosa, due to a lack of success in transformation. It is well known that meristemoids situated internally to the surface of the explants can reduce the efficiency of Agrobacterium mediated transformation, while shoots originating on the surface have a better chance of being transformed (Fernando, Vieira, Machado, & Appezzato-da Glória, 2007). The origin of the cells involved in the organogenic process must therefore be located and identified for successful transformation protocols.
Histological examination had provided details that help us to understand the process of organogenesis from P. tomentosa petioles cultured in vitro in medium containing TDZ. In the present study, meristemoids have been identified in both direct and indirect patterns of development. During the direct pathway, the formation of a meristem proceeds without intermediate proliferation of undifferentiated callus tissue. However, meristems can be formed indirectly from unspecialized cells in calluses or culture suspensions (Yancheva, Golubowicz, Fisher, Lev-Yadun, & Flaishman, 2003; Gahan & George, 2008). The direct and indirect origin of adventitious buds has been described for monocotyledons (e.g., Paphiopedilum spp., Chen, Chen, & Chang, 2004; Lycaste spp., Huang & Chung, 2011), dicotyledons (e.g., Citrus spp., Almeida et al., 2003; Malus x domestica, Yancheva et al., 2003; Passiflora edulis, Fernando et al., 2007; Centaurea ultreiae, Mallón, Rodríguez-Oubiña, & González, 2011) and conifers (e.g., Pinus massoniana, Zhang, Wei, Xi, & Shi, 2006; P. pinea, Cuesta, Rodríguez, Centeno, Ordas, & Fernández, 2009). In Paulownia, the cell divisions that lead to dedifferentiation of the primordial took place in subepidermal or epidermal cells. Direct meristemoids appeared to develop on the adaxial surface of the petiole from a single cell or small group of cells in the subepidermal or epidermal region of the explants. Whereas indirect meristemoids developed from rows of cells formed by superficial periclinally dividing secondary meristems (diffuse cambium-like zones) located on the abaxial surface of the petiole, which provided the bulk of cells for development of bud primordia. According to Gautheret (1959), a diffuse cambium is the result of the in vitro proliferation in a preferential direction, induced by contact with the culture medium. The presence of diffuse cambium during the formation of adventitious buds in foliar explants of beech have been described in detail by Cuenca & Vieitez (1999). Hu, Liu, Yan & Xie (2005) also observed differentiation of a long narrow meristematic zone in the subepidermis of petioles of Amorphophallus, from which the meristemoids were formed. The involvement of epidermic and subepidermic layers in bud formation have been reported in different plant species (Hervé et al., 2001; Budimir, 2003; Ghimire et al., 2010).
As occurs in the shoot, where actual cell division occurs, certain meristem cells undergo divisions in such a way that one product of division becomes a new body cell, denominated derivative and the other remains in the meristem, and is denominated initial (Esau, 1977). A similar pattern was observed in the present study, the formation of meristemoids may be considered as a two-stage process. In the first stage, the subepidermal cells dedifferentiate to form cambial-type cells that divide periclinally, giving rise to formation of secondary meristems. In the second stage, the outermost cells of these secondary meristems divide anticlinally and dedifferentiate to form primary meristems that gave rise to formation of meristemoids, described by Torrey (1966) as meristematic masses in the callus with capability of differentiating into roots, shoots or embryos. The models of division appear to be of great importance in defining the first meristematic cells that will lead to the organization of meristemoids. In a study carried out with foliar discs from Saintpaulia, Lo, Giles, & Sawhney (1997) observed that epidermal cells did not directly regenerate into adventitious shoots, periclinal divisions were required and then a daughter cell (target cell) from one of the periclinal divisions divided both periclinally and anticlinally to form a cell division centre (meristemoid), which was the precursor of an adventitious shoot. The “target” cell is considered the “competent” cell for shoot induction, and the formation of target cells may be interpreted in terms of competence acquisition. Similar observations of such changes in the cell division model during the differentiation of promeristemoids have been reported for other species, such as Fagus orientalis (Cuenca & Vieitez, 1999) and Jatropa curcas (Varshney, Sangapillai, Patil, & Johnson, 2011).
At the same time as the cellular dedifferentiation occurred in the most superficial zones of the leaf explants, proliferation of the inner parenchyma cells took place in the most internal zones. This proliferation contributed to form internal callus tissue with numerous tracheary elements, that established vascular connections with the differentiated buds. Differentiation of meristemoids has not been observed in the internal callus tissue. Hu et al. (2005) observed that when meristemoids were derived from inner tissues, abnormal shoot buds formed.
Independently of the origin of the organogenic process in the explant tissue, histological characterization of the process and identification of the morphogenic regions can provide information about alterations in the organogenic system that will favour the contact between the Agrobacterium and the morphogenetic region, enhancing the efficiency of genetic transformation. As also mentioned by Almeida, Mourao Filho, Mendes, & Rodríguez (2006), a direct regeneration pathway may be used for transformation purposes as a means to avoid the formation of chimeric plants and genetic variability, which can occur through regeneration from callus. This protocol may be a useful tool for micropropagation of the species as well as for the application of genetic transformation techniques, as it enables us to determine specific regions in the foliar explants where initiation of meristemoids will take place, and therefore to determine which cells should be the object of genetic transformation. The pre-culture of Paulownia tissues with phytohormones for three-five days before co-cultivation with A. tumefaciens may increase the transformation rates. This is consistent with the present results on the target explants, which showed dedifferentiated cells in subepidermal and/or epidermal layers. During pre-culture, cells were actively dividing and at the moment of bacterial inoculation, cells would be in a competent state for transformation.
In conclusion, in this study we have established the specific regions in the foliar explants where the differentiation takes place, and therefore which cells should be chosen as the target in the Agrobacterium studies. This article may contribute to improve the genetic transformation protocols in Paulownia.
Acknowledgments
We thank Daniel García Areán for technical support. Financial support from XUNTA GALICIA (Spain) through Project 09MRU-002400PR is also gratefully acknowledged.
References
Almeida, W. A. B. de, Mourao Filho, F. de A. A., Mendes, B. M. J., & Rodríguez, A. P. M. (2006). Histological characterization of in vitro adventitious organogenesis. Biologia Plantarum, 50, 321-325. [ Links ]
Almeida, W. A. B. de, Mourao Filho, F. de A. A., Pino, L. E., Boscariol, R. L., Rodríguez, A. P. M., & Mendes, B. M. J. (2003). Genetic transformation and plant recovery from mature tissues of Citrus sinensis L. Osbeck. Plant Science, 164, 203-211. [ Links ]
Balat, M., Balat, H., & Öz, C. (2008). Progress in bioethanol processing. Progress in Energy and Combustion Science, 34, 551-573. [ Links ]
Beltrán, J. P. (2008). Plantas para comer, plantas para quemar….¿Una cuestión de diseño genético? Mètode, 55, 203-209. [ Links ]
Bergmann, B. A. (1998). Propagation method influences first year field survival and growth of Paulownia. New Forests, 16, 251-264. [ Links ]
Bergmann, B. A. & Moon, H. K. (1997). In vitro adventitious shoot production in Paulownia. Plant Cell Reports, 16, 315-318. [ Links ]
Budimir, S. (2003). Developmental histology of organogenic and embryogenic tissue in Picea omorika culture. Biologia Plantarum, 47, 467-470. [ Links ]
Burger, D. W. (1989). Empress tree (Paulownia tomentosa Steud.). In Y. P. S. Bajaj (Ed.), Biotechnology in agriculture and forestry (pp. 359-369). Berlin: Springer. [ Links ]
Castellanos-Hernández, O. A., Rodríguez-Sahagún, A., Acevedo-Hernández, G. J., Rodríguez-Garay, B., Cabrera-Ponce, J. L., & Herrera-Estrella, L. H. (2009). Transgenic Paulownia elongata S. Y. Hu plants using biolistic-mediated transformation. Plant Cell Tissue and Organ Culture, 99, 175-181. [ Links ]
Chateau, S., Sangwan, R. S., & Sanhgwan-Norreel, S. (2000). Competence of Arabidopsis thaliana genotypes and mutants for Agrobacterium tumefaciensmediated transformation. Journal of Experimental Botany, 51, 1961-1968. [ Links ]
Chen, T. Y., Chen, J. T., & Chang, W. C. (2004). Plant regeneration through direct shoot bud formation from leaf cultures of Paphiopedilum orchids. Plant Cell Tissue and Organ Culture, 76, 11-15. [ Links ]
Chovelon, V., Restier, V., Giovinazzo, N., Dogimont, C., & Aarrouf, J. (2011). Histological study of organogenesis in Cucumis melo L. after genetic transformation: why is it difficult to obtain transgenic plants? Plant Cell Reports, 30, 2001-2011. [ Links ]
Corredoira, E., Ballester, A., & Vieitez, A. M. (2008). Thidiazuron-induced high frequency plant regeneration from leaf explants of Paulownia tomentosa mature trees. Plant Cell Tissue and Organ Culture, 95, 197-208. [ Links ]
Cuenca, B. & Vieitez, A. M. (1999). Histological study of in vitro development of adventitious buds on leaf explants of oriental beech (Fagus orientalis Lipsky). In Vitro Cellular & Developmental Biology – Plant, 35, 326-332. [ Links ]
Cuesta, C., Rodríguez, A., Centeno, M. L., Ordas, R. J., & Fernández, B. (2009). Caulogenic induction in cotyledons of Stone pine (Pinus pinea): relationship between organogenic response and benzyladenine trends in selected families. Journal of Plant Physiology, 166, 1162-1171. [ Links ]
Doumett, S., Lamperi, L., Checchini, L., Azzarello, E., Mugnai, S., Mancuso, S., Petruzelli, G., & Del Bubba, M. (2008). Heavy metal distribution between contamined soil and Paulownia tomentosa, in a pilot-scale assisted phytoremediation study: influence of different complexing agents. Chemosphere, 72, 1481-1490. [ Links ]
Esau, K. 1977. Anatomy of seed plants. New York: John Wiley & Sons. [ Links ]
Fernando, J. A., Vieira, M. L. C., Machado, S. R., & Appezzato-da-Glória, B. (2007). New insights into the in vitro organogenesis process: the case of Passiflora. Plant Cell Tissue and Organ Culture, 91, 37-44. [ Links ]
Gahan, P. B. & George, E. F. (2008). Adventitious regeneration. In E. F. George, M. A. Hall & G. J. de Klerk (Eds.), Plant propagation by tissue culture (pp. 355-401). Dordrecht: Springer. [ Links ]
Gautheret, R. J. (1959). La culture des tissus végétaux: techniques et réalisations. Paris: Massons. [ Links ]
Ghimire, B. K., Seong, E. S., Goh, E. J., Kim, N. Y., Kang, W. H., Kim, E. H., Yu, C. Y., & Chung, I. M. (2010). High-frequency direct shoot regeneration from Drymaria cordata Willd. leaves. Plant Cell Tissue and Organ Culture, 100, 209-217. [ Links ]
Hervé, P., Jauneau, A., Pâques, M., Marien, J. N., Boudet, A. M., & Teulières, C. (2001). A procedure for shoot organogenesis in vitro from leaves and nodes of an elite Eucalyptus gunnii clone: comparative histology. Plant Science, 16, 645-653. [ Links ]
Hu, J. B., Liu, J., Yan, H. B., & Xie, C. H. (2005). Histological observations of morphogenesis in petiole derived callus of Amorphophallis rivieri Durieu in vitro. Plant Cell Reports, 24, 642-648. [ Links ]
Huang, C. H. & Chung, J. P. (2011). Efficient indirect induction of protocorm-like bodies and shoot proliferation using field-grown axillary buds of a Lycaste hybrid. Plant Cell Tissue and Organ Culture, 106, 31-38. [ Links ]
Jensen, W. A. (1962). Botanical histochemistry. San Francisco: H. Freeman. [ Links ]
Johnson, T. S., Eswaran, N., & Sujatha, M. (2011). Molecular approaches to improvement of Jatropha curcas Linn. as a sustainable energy crop. Plant Cell Reports, 30, 1573-1591. [ Links ]
Kausch, A. P., Hague, J., Melvin Oliver, M., Li, Y., Daniell, H., Mascia, P., Watrud, L. S., & Steward, Jr. C. N. (2010). Transgenic perennial biofuel feedstocks and strategies for bioconfinement. Biofuels, 1, 163-176. [ Links ]
Limayem, A. & Ricke, S. C. (2012). Lignocellulosic biomass for bioethanol production: Current perspectives, potential issues and future prospects. Progress in Energy and Combustion Science, 38, 449-467. [ Links ]
Lo, K. H., Giles, K. L., & Sawhney, V. K. (1997). Histological changes associated with acquisition of competence for shoot regeneration in leaf discs of Saintpaulia ionantha x confuse hybrid (African violet) cultured in vitro. Plant Cell Reports, 16, 421-425. [ Links ]
López, F., Pérez, A., Zamudio M., De Alva, H. E., & García J. C. (2012). Paulownia as raw material for solid biofuel and cellulose pulp. Biomass and Bioenergy, 45, 77-86. [ Links ]
Mallón, R., Rodríguez-Oubiña, J., & González, M. L. (2011). Shoot regeneration from in vitro-derived leaf and root explants of Centaurea ultreiae. Plant Cell Tissue and Organ Culture, 106, 523-530. [ Links ]
Mandpe, S., Kadlaskar, S., Degen, W., & Keppeler, S. (2005). On road testing of advanced common rail diesed vehicles with biodiesel from the Jatrophacurcas plant. Society of Automotive Engineering International, 26, 356-364. [ Links ]
Marcotrigiano, M. & Jagannathan, L. (1988). Paulownia tomentosa somaclonal snowstorm. Hortscience, 23, 226-227. [ Links ]
Murashige, T. & Skoog, F. (1962). A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiologia Plantarum, 15, 473-497. [ Links ]
O’Brien, T. P. & McCully, M. E. (1981). The study of plant structure: principles and selected methods. Melbourne: Hermacarphi Pty Ltd. [ Links ]
Rao, C. D., Goh, C. J., & Kumar, P. P. (1996). High frequency adventitious shoot regeneration from excised leaves of Paulownia spp. cultured in vitro. Plant Cell Reports, 16, 204-209. [ Links ]
Song, S. L., Sato, T., Saito, A., & Kihachiro, O. (1989). Meristematic culture of seven Paulownia species. Journal of the Japanese Forestry Society, 71, 456-459. [ Links ]
Torrey, J. (1966). The initiation of organized development in plants. Advances in Morphogenesis, 5, 39-91. [ Links ]
Varshney, A., Sangapillai, R., Patil, M. S., & Johnson, T. S. (2011). Histological evidence of morphogenesis from various explants of Jatropa curcas L. Trees, 25, 689-694. [ Links ]
Yancheva, S. D., Golubowicz, S., Fisher, E., Lev-Yadun, S., & Flaishman, M. A. (2003). Auxin type and timing of application determine the activation of the developmental program during in vitro organogenesis in apple. Plant Science, 165, 299-309. [ Links ]
Yang, J. C., Ho, C. K., Chen, Z. Z., & Chang, S. H. (1996). Paulownia x taiwaniana (Taiwan Paulownia). In Y. P. S. Bajaj (Ed.), Biotechnology in agriculture and forestry (pp. 269-290). Berlin: Springer. [ Links ]
Zhang, Y., Wei, Z. M., Xi, M. L., & Shi, J. S. (2006). Direct organogenesis and plantlet regeneration from mature zygotic embryos of masson pine (Pinus massoniana L.). Plant Cell Tissue and Organ Culture, 84, 119-123. [ Links ]
Zhu, Z. H., Chao, C. J., Lu, X. Y., & Xiong, Y. G. (1986). Paulownia in China: cultivation and utilization. Asian Network of Biological Sciences & International Development Research Center (pp. 1-65) (Canada), Singapore. [ Links ]
Almeida, W. A. B. de, Mourao Filho, F. de A. A., Pino, L. E., Boscariol, R. L., Rodríguez, A. P. M., & Mendes, B. M. J. (2003). Genetic transformation and plant recovery from mature tissues of Citrus sinensis L. Osbeck. Plant Science, 164, 203-211. [ Links ]
Balat, M., Balat, H., & Öz, C. (2008). Progress in bioethanol processing. Progress in Energy and Combustion Science, 34, 551-573. [ Links ]
Beltrán, J. P. (2008). Plantas para comer, plantas para quemar….¿Una cuestión de diseño genético? Mètode, 55, 203-209. [ Links ]
Bergmann, B. A. (1998). Propagation method influences first year field survival and growth of Paulownia. New Forests, 16, 251-264. [ Links ]
Bergmann, B. A. & Moon, H. K. (1997). In vitro adventitious shoot production in Paulownia. Plant Cell Reports, 16, 315-318. [ Links ]
Budimir, S. (2003). Developmental histology of organogenic and embryogenic tissue in Picea omorika culture. Biologia Plantarum, 47, 467-470. [ Links ]
Burger, D. W. (1989). Empress tree (Paulownia tomentosa Steud.). In Y. P. S. Bajaj (Ed.), Biotechnology in agriculture and forestry (pp. 359-369). Berlin: Springer. [ Links ]
Castellanos-Hernández, O. A., Rodríguez-Sahagún, A., Acevedo-Hernández, G. J., Rodríguez-Garay, B., Cabrera-Ponce, J. L., & Herrera-Estrella, L. H. (2009). Transgenic Paulownia elongata S. Y. Hu plants using biolistic-mediated transformation. Plant Cell Tissue and Organ Culture, 99, 175-181. [ Links ]
Chateau, S., Sangwan, R. S., & Sanhgwan-Norreel, S. (2000). Competence of Arabidopsis thaliana genotypes and mutants for Agrobacterium tumefaciensmediated transformation. Journal of Experimental Botany, 51, 1961-1968. [ Links ]
Chen, T. Y., Chen, J. T., & Chang, W. C. (2004). Plant regeneration through direct shoot bud formation from leaf cultures of Paphiopedilum orchids. Plant Cell Tissue and Organ Culture, 76, 11-15. [ Links ]
Chovelon, V., Restier, V., Giovinazzo, N., Dogimont, C., & Aarrouf, J. (2011). Histological study of organogenesis in Cucumis melo L. after genetic transformation: why is it difficult to obtain transgenic plants? Plant Cell Reports, 30, 2001-2011. [ Links ]
Corredoira, E., Ballester, A., & Vieitez, A. M. (2008). Thidiazuron-induced high frequency plant regeneration from leaf explants of Paulownia tomentosa mature trees. Plant Cell Tissue and Organ Culture, 95, 197-208. [ Links ]
Cuenca, B. & Vieitez, A. M. (1999). Histological study of in vitro development of adventitious buds on leaf explants of oriental beech (Fagus orientalis Lipsky). In Vitro Cellular & Developmental Biology – Plant, 35, 326-332. [ Links ]
Cuesta, C., Rodríguez, A., Centeno, M. L., Ordas, R. J., & Fernández, B. (2009). Caulogenic induction in cotyledons of Stone pine (Pinus pinea): relationship between organogenic response and benzyladenine trends in selected families. Journal of Plant Physiology, 166, 1162-1171. [ Links ]
Doumett, S., Lamperi, L., Checchini, L., Azzarello, E., Mugnai, S., Mancuso, S., Petruzelli, G., & Del Bubba, M. (2008). Heavy metal distribution between contamined soil and Paulownia tomentosa, in a pilot-scale assisted phytoremediation study: influence of different complexing agents. Chemosphere, 72, 1481-1490. [ Links ]
Esau, K. 1977. Anatomy of seed plants. New York: John Wiley & Sons. [ Links ]
Fernando, J. A., Vieira, M. L. C., Machado, S. R., & Appezzato-da-Glória, B. (2007). New insights into the in vitro organogenesis process: the case of Passiflora. Plant Cell Tissue and Organ Culture, 91, 37-44. [ Links ]
Gahan, P. B. & George, E. F. (2008). Adventitious regeneration. In E. F. George, M. A. Hall & G. J. de Klerk (Eds.), Plant propagation by tissue culture (pp. 355-401). Dordrecht: Springer. [ Links ]
Gautheret, R. J. (1959). La culture des tissus végétaux: techniques et réalisations. Paris: Massons. [ Links ]
Ghimire, B. K., Seong, E. S., Goh, E. J., Kim, N. Y., Kang, W. H., Kim, E. H., Yu, C. Y., & Chung, I. M. (2010). High-frequency direct shoot regeneration from Drymaria cordata Willd. leaves. Plant Cell Tissue and Organ Culture, 100, 209-217. [ Links ]
Hervé, P., Jauneau, A., Pâques, M., Marien, J. N., Boudet, A. M., & Teulières, C. (2001). A procedure for shoot organogenesis in vitro from leaves and nodes of an elite Eucalyptus gunnii clone: comparative histology. Plant Science, 16, 645-653. [ Links ]
Hu, J. B., Liu, J., Yan, H. B., & Xie, C. H. (2005). Histological observations of morphogenesis in petiole derived callus of Amorphophallis rivieri Durieu in vitro. Plant Cell Reports, 24, 642-648. [ Links ]
Huang, C. H. & Chung, J. P. (2011). Efficient indirect induction of protocorm-like bodies and shoot proliferation using field-grown axillary buds of a Lycaste hybrid. Plant Cell Tissue and Organ Culture, 106, 31-38. [ Links ]
Jensen, W. A. (1962). Botanical histochemistry. San Francisco: H. Freeman. [ Links ]
Johnson, T. S., Eswaran, N., & Sujatha, M. (2011). Molecular approaches to improvement of Jatropha curcas Linn. as a sustainable energy crop. Plant Cell Reports, 30, 1573-1591. [ Links ]
Kausch, A. P., Hague, J., Melvin Oliver, M., Li, Y., Daniell, H., Mascia, P., Watrud, L. S., & Steward, Jr. C. N. (2010). Transgenic perennial biofuel feedstocks and strategies for bioconfinement. Biofuels, 1, 163-176. [ Links ]
Limayem, A. & Ricke, S. C. (2012). Lignocellulosic biomass for bioethanol production: Current perspectives, potential issues and future prospects. Progress in Energy and Combustion Science, 38, 449-467. [ Links ]
Lo, K. H., Giles, K. L., & Sawhney, V. K. (1997). Histological changes associated with acquisition of competence for shoot regeneration in leaf discs of Saintpaulia ionantha x confuse hybrid (African violet) cultured in vitro. Plant Cell Reports, 16, 421-425. [ Links ]
López, F., Pérez, A., Zamudio M., De Alva, H. E., & García J. C. (2012). Paulownia as raw material for solid biofuel and cellulose pulp. Biomass and Bioenergy, 45, 77-86. [ Links ]
Mallón, R., Rodríguez-Oubiña, J., & González, M. L. (2011). Shoot regeneration from in vitro-derived leaf and root explants of Centaurea ultreiae. Plant Cell Tissue and Organ Culture, 106, 523-530. [ Links ]
Mandpe, S., Kadlaskar, S., Degen, W., & Keppeler, S. (2005). On road testing of advanced common rail diesed vehicles with biodiesel from the Jatrophacurcas plant. Society of Automotive Engineering International, 26, 356-364. [ Links ]
Marcotrigiano, M. & Jagannathan, L. (1988). Paulownia tomentosa somaclonal snowstorm. Hortscience, 23, 226-227. [ Links ]
Murashige, T. & Skoog, F. (1962). A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiologia Plantarum, 15, 473-497. [ Links ]
O’Brien, T. P. & McCully, M. E. (1981). The study of plant structure: principles and selected methods. Melbourne: Hermacarphi Pty Ltd. [ Links ]
Rao, C. D., Goh, C. J., & Kumar, P. P. (1996). High frequency adventitious shoot regeneration from excised leaves of Paulownia spp. cultured in vitro. Plant Cell Reports, 16, 204-209. [ Links ]
Song, S. L., Sato, T., Saito, A., & Kihachiro, O. (1989). Meristematic culture of seven Paulownia species. Journal of the Japanese Forestry Society, 71, 456-459. [ Links ]
Torrey, J. (1966). The initiation of organized development in plants. Advances in Morphogenesis, 5, 39-91. [ Links ]
Varshney, A., Sangapillai, R., Patil, M. S., & Johnson, T. S. (2011). Histological evidence of morphogenesis from various explants of Jatropa curcas L. Trees, 25, 689-694. [ Links ]
Yancheva, S. D., Golubowicz, S., Fisher, E., Lev-Yadun, S., & Flaishman, M. A. (2003). Auxin type and timing of application determine the activation of the developmental program during in vitro organogenesis in apple. Plant Science, 165, 299-309. [ Links ]
Yang, J. C., Ho, C. K., Chen, Z. Z., & Chang, S. H. (1996). Paulownia x taiwaniana (Taiwan Paulownia). In Y. P. S. Bajaj (Ed.), Biotechnology in agriculture and forestry (pp. 269-290). Berlin: Springer. [ Links ]
Zhang, Y., Wei, Z. M., Xi, M. L., & Shi, J. S. (2006). Direct organogenesis and plantlet regeneration from mature zygotic embryos of masson pine (Pinus massoniana L.). Plant Cell Tissue and Organ Culture, 84, 119-123. [ Links ]
Zhu, Z. H., Chao, C. J., Lu, X. Y., & Xiong, Y. G. (1986). Paulownia in China: cultivation and utilization. Asian Network of Biological Sciences & International Development Research Center (pp. 1-65) (Canada), Singapore. [ Links ]
*Correspondencia: Mª del Carmen San José: Instituto de Investigaciones Agrobiológicas de Galicia, CSIC, Apartado 122, 15705 Santiago de Compostela, Spain; sanjose@iiag.csic.es
Mª José Cernadas: Instituto de Investigaciones Agrobiológicas de Galicia, CSIC, Apartado 122, 15705 Santiago de Compostela, Spain; cernadas@iiag.csic.es,
Elena Corredoira: Instituto de Investigaciones Agrobiológicas de Galicia, CSIC, Apartado 122, 15705 Santiago de Compostela, Spain; elenac@iiag.csic.es
1. Instituto de Investigaciones Agrobiológicas de Galicia, CSIC, Apartado 122, 15705 Santiago de Compostela, Spain; sanjose@iiag.csic.es, cernadas@iiag.csic.es, elenac@iiag.csic.es
Received 29-VII-2013. Corrected 07-XI-2013. Accepted 06-XII-2013.


{kind=link}
{kind=link}
{kind=link}