










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.62 n.2 San José Apr./Jun. 2014
A distinctive avian assemblage (Aves: Passeriformes) in Western Darién, Panama is uncovered through a disease surveillance program
Un conglomerado distinto de aves Aves: Passeriformes) descubierto en el Darién occidental, Panamá por un programa de vigilancia de enfermedades
Un conglomerado distinto de aves Aves: Passeriformes) descubierto en el Darién occidental, Panamá por un programa de vigilancia de enfermedades
*Dirección para correspondencia:
Abstract
Basic knowledge about the distribution of flora and fauna is lacking for most tropical areas. Improving our knowledge of the tropical biota will help address contemporary global problems, including emerging tropical diseases. Less appreciated is the role that applied studies can have in improving our understanding of basic biological patterns and processes in the tropics. Here, I describe a novel avifauna assemblage uncovered in Western Darién province in the Republic of Panama that was uncovered during a vector-borne disease surveillance program. I compared the passerine bird species composition at 16 sites using records from recent ornithological expeditions sponsored by the Smithsonian Tropical Research Institute in Central and Eastern Panama. Based on the results of a Mantel test, geographic distance did not correlate with pairwise distinctiveness of sites. Instead, based on an index of distinctiveness modified from the Chao-Jaccard index, most sites were more or less similarly distinctive, with one site, Aruza Abajo, significantly more distinctive than the rest. I found that the distinctiveness of this site was due not only to the presence of several rare and range-restricted taxa, but also to the absence of taxa that are common elsewhere. This finding provides more evidence of high species composition turnover (beta-diversity) in the Panamanian biota, which appears to be driven by a combination of soil and climate differences over narrow distances. Rev. Biol. Trop. 62 (2): 711-717. Epub 2014 June 01.
Key words: community composition, Darién, rare birds, beta-diversity.
Resumen
El conocimiento básico sobre la distribución de la flora y fauna es escaso en la mayoría de las zonas tropicales. Mejorar el conocimiento de la biota tropical ayudará a enfrentar los problemas globales contemporáneos, incluyendo las enfermedades tropicales emergentes. Menos apreciado es el papel que los estudios aplicados pueden tener en mejorar nuestra comprensión de los patrones biológicos y procesos básicos en estas regiones. A continuación, describo un conglomerado distinto de aves encontrado en la provincia occidental de Darién en la República de Panamá, el cual fue descubierto durante un programa de vigilancia de enfermedades transmitidas por insectos vectores. En este estudio comparé la composición de especies de aves paseriformes de 16 sitios con registros de expediciones ornitológicas recientes en Panamá Central y Oriental patrocinadas por el Instituto Smithsonian de Investigaciones Tropicales. Basado en los resultados de la prueba de Mantel, el carácter distintivo de los sitios en comparaciones pareadas no se correlacionó con la distancia geográfica entre sitios. En cambio, con base a un índice de disimilitud modificado a partir del índice de Chao-Jaccard, la mayoría de los sitios presentaron grados de disimilitud más o menos equivalentes, sin embargo Aruza Abajo fue mucho más distintivo que el resto. Yo encontré que el carácter distintivo de Aruza Abajo fue la presencia de varias especies raras y de rango restringido, pero también existe ausencia de especies comunes en otros sitios. Este hallazgo proporciona evidencia del elevado reemplazo de especies de aves (diversidad beta) en la avifauna panameña, la cual parece estar determinada por una combinación de diferencias del suelo y clima en cortas distancias geográficas.
Palabras clave: composición de la comunidad, Darién, aves poco-comunes, beta-diversidad.
For most tropical areas, we continue to lack basic knowledge regarding distributional patterns of biodiversity as well the processes that create and maintain this biodiversity. At the same time, this knowledge gap impedes our ability to address problems of conservation, agriculture, and mitigation of emerging tropical diseases using the best available science. Recently, many natural history museums have begun to close this knowledge gap in both basic and applied biology through expeditions in collaboration with applied scientists (Winker, 2004; Pyke & Ehrlich, 2010). While it is routinely acknowledged that basic scientific discovery informs applied scientific inquiry (Yasué et al. 2006), less attention is given to the role that applied scientific endeavors can play in improving our basic knowledge about biodiversity patterns, especially in the tropics. Here, I describe an instance where a collection trip to survey birds and mosquitos for equine encephalitic virus uncovered a unique and overlooked bird assemblage in Panama, a country with one of the best-studied avifauna in the Neotropics.
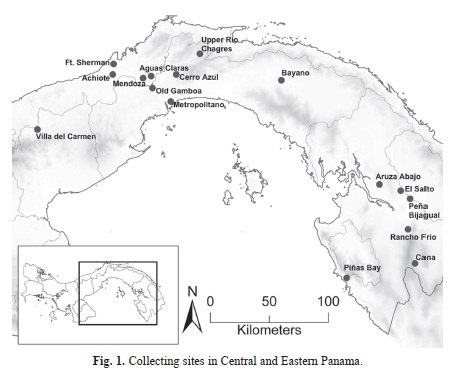
The Smithsonian Tropical Research Institute Bird Collection (STRIBC) was founded in 2007 in order to better describe patterns of avian biodiversity across the Isthmus of Panama as well as to understand interactions between birds and other environmental factors and biological agents, especially with regard to emerging tropical diseases. In 2010, at the onset of an outbreak of equine encephalitis in Eastern Panama that had claimed at least two human lives and caused considerable loss of livestock, the Panamanian Ministry of Agriculture requested that the STRIBC sample wild birds and mosquitoes in Aruza Abajo (geographical coordinates: 8.36, −77.95, Fig. 1), an affected site in Western Darién Province. Equine encephalitis outbreaks occur periodically in Eastern Panama, but almost nothing is know about the ecology of viral transmission in the area, including which mosquito species are local vectors (Navia-Gine et al. 2013) or whether rodents or birds are natural hosts (Arrigo, Adams, & Weaver, 2010).
At Aruza Abajo, I was impressed by many of the birds we encountered. For example, we collected the second Middle American specimen of the flycatcher Tolmomyias flaviventris, the third, fourth and fifth Middle American specimens of the woodpecker Colaptes punctigula, and several specimens of Phaethornis anthophilus, being the most abundant hummingbird at the site. This species had only been recorded in Panama from five specimens collected more than 45 years ago. The ornithological literature for Panama (Ridgely & Gwynne, 1989; Angehr & Dean, 2010) refers to the Eastern portion of Darién province as being the most distinctive in Panama, but it is clear from these sources and others (González, Eberhard, Lovette, Olson, & Bermingham, 2003; Siegel & Olson, 2008) that Western Darién province has been systematically overlooked by ornithologists. In order to test my observation that the avifaunal assemblage in Western Darién province, and especially Aruza Abajo, was distinctive, I compared our records from Aruza Abajo to those of other locations where my colleagues and I have collected birds in Panama since 2002.
Materials and methods
I generated species incidence lists for 16 locations (Fig. 1) where the STRIBC or STRIsponsored foreign natural history museums have collected birds in central and Eastern Panama since 2002. I participated in the collecting efforts at 15 of the 16 locations. Field work at each site varied from two and 13 days. Most expeditions were disease ecology surveys, while a few were solely biodiversity surveys, but all shared the same goal of sampling broadly across the local bird community, rather than focused efforts to collect particular species of interest. In all cases, the majority of collecting employed standard ornithological mistnets, but occasionally collecting was supplemented by shotgun, where permitted. Data come from the digital collections databases managed by STRI. Although our fieldwork included collecting both passerines and non-passerines, in order to improve consistency among sites, I restricted my analyses to resident passerines only. All scientific collecting in Panama sponsored by STRI was approved by ANAM, Panama’s Environmental Authority (permit numbers: DNPN-01-2002, DNPN-01-2003, DNPN-01-2004, DNPN-01-2004, SE/A-60-10, SE/A-137-10, SE/A-96-09, SE/A-44-10, SE/A-66-11, SE/A-2-12); likewise, scientific collecting undertaken by the STRI Bird Collection is done under IACUC approval (permits: 2007-03-03-15-07, 2011-0927-2014-03, 2013-0801-2016).
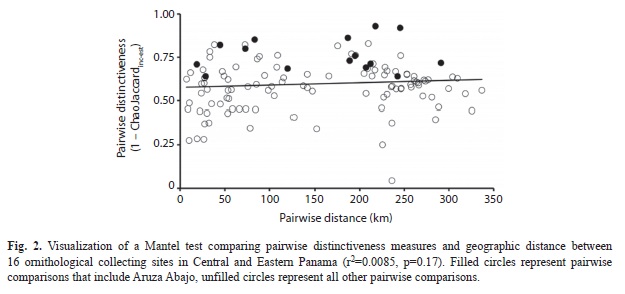
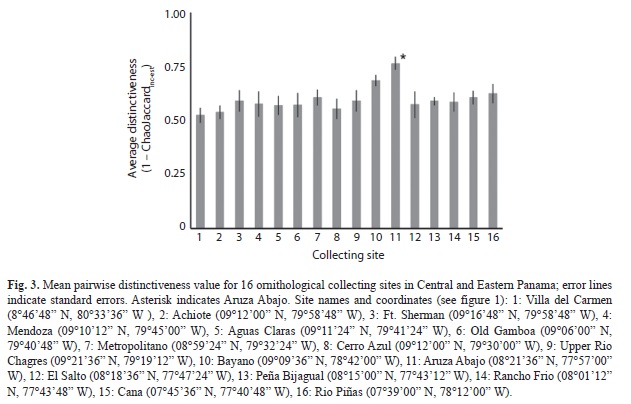
I used the Chao-Jaccard similarity index modified for replicated incidence data (Chao Chao, Chazdon, Colwell, & Shen, 2005) to measure passerine species assembly distinctiveness among the 16 localities. For each site, I tallied daily incidence (i.e. presence vs. absence). Therefore, for a given site, the total incidence for any given species could vary from 0 (never collected at that site) to n, where n equals the number of days of fieldwork at that site. I chose this replicated incidence approach because effort at each site varied in several ways: the number of days at a site, the number of nets employed and the hours of operation in a day, whether shotgun collecting was allowed, and because the total maximum number of collected specimens per species from a given site varied among species based either on our Panamanian collecting permit or our overall research objectives. Details about number of field personnel, mistnets deployed, and mistnet-hours was not available. Therefore, replicated incidence rather than a raw abundance approach should best deal with these differences in sampling effort, as well as the problem of incomplete sampling at a given site (Chao et al., 2005, 2006). It is certain that at none of the 16 sites our collection efforts approached a full sampling of the local passerine community. The modified Chao-Jaccard index (ChaoJaccardinc-est) was computed pairwise among all 16 sites in the program EstimateS 9 (Colwell, 2013). However, for the rest of this paper, I will be discussing assemblage distinctiveness, which I define as simply 1- ChaoJaccardinc-est. Thus, a distinctiveness value of 0 represents two sites where avifaunal assemblages overlap completely, whereas a value of 1 indicates two sites with completely different bird faunas. I tested for a relationship between pairwise site distinctiveness and the geographic distance using a Mantel test implemented in R (R Core Team 2012). In order to compare the distinctiveness among sites, I generated 16 t-tests, comparing each sites average pairwise distinctiveness measure to that of all other comparisons (that did not include that site). To correct for multiple comparisons, I employed a sequential Bonferroni correction which constrains the familywise error rate (e.g. overall P-value) to 0.05.
Results
Across the 16 sites, 183 species of resident passerines were recorded. Among the 16 sites, raw passerine species richness varied between 16 and 71 (mean S: 42.4±18.0), while species incidence (i.e. total species days) varied from 20 and 219 (mean: 88.3±51.6). No species occurred on all 16 collecting tally lists, and only one species (Mionectes oleagineus) was recorded from 15 locations, with five additional species collected in at least 13 sites. Instead, the distribution of species shows a long tail of relatively rare taxa: 116 species (63.3% of all species recorded in the study) were collected in three or fewer sites and nearly a third of all species in the study (N=60) were collected in only one location. Data for all 16 collecting sites are available at the Figshare depository (DOI: 10.6084/m9.figshare.941075).
Among the 16 sites, distinctiveness averaged 0.60, and ranged from 0.52 to 0.76, with the latter representing Aruza Abajo. Among all pairwise comparisons, the two sites estimated to be most similar were Old Gamboa and El Salto, whereas the most distinctive were found to be Mendoza and Aruza Abajo. A Mantel test failed to rejected the null hypothesis that pairwise distinctive was independent of geographic distance (r=0.092, p=0.17, Fig. 2). The results of the 16 pairwise t-tests found that three locations had significantly higher distinctiveness scores than the remaining pair-wise comparisons before correcting for multiple tests (Villa del Carmen: p=0.03; Bayano: p=0.009; Aruza Abajo: p=2.0×10-7, Fig. 3), with only Aruza Abajo remaining significant after sequential Bonferroni correction (corrected α=0.003).
Discussion
The passerine avifauna at Aruza Abajo represents a distinctive fauna in Central and Eastern Panama unlike any other of the 15 ornithological sites surveyed by STRI-affiliated ornithologists. Whether this distinctive avifauna is restricted only to the area immediately around Aruza Abajo, or extends across the Western Darién Province is unknown, as bird distribution data from most of this region is lacking (Siegel & Olson, 2008). Likewise, it remains to be determined if other faunal groups in this region (such as rodents and mosquitoes) are also distinctive, which may influence vector-borne disease patterns. What is clear from the data is that across the region, local assemblages of birds are not homogenous, but instead tend to vary considerably in composition. Importantly, similarity among sites is not a function of geographical proximity, with bird assemblages varying over relatively narrow geographic scales. This finding has implications for our understanding of both avian biodiversity and the transmission ecology of bird-borne diseases in Panama.
Several of the passerines collected at Aruza Abajo represented near-endemic species found only in Eastern Panama and adjacent Northern Colombia, including the flycatcher Tolmomyias flaviventris, the wren Campylorhynchus albobrunneus, and the conebill tanager Conirostrum leucogenys. These taxa were collected only at Aruza Abajo, or in the case of the wren, at one additional site. However, other taxa that were collected exclusively or almost exclusively at Aruza Abajo include more widespread taxa such as the antbird Myrmeciza longipes and the wren Cantorchilus leucotis. Although these species can be found across much of Panama, they were more routinely collected in Aruza Abajo relative to the remaining 15 sites. Finally, it is important to note that while these findings refer to only passerines it is likely that the assemblage of non-passerines at Aruza Abajo is similarly distinctive; the hummingbird and woodpecker examples mentioned earlier are two species of non-passerines that were almost never collected outside Aruza Abajo.
The avifauna of Aruza Abajo is distinctive not just for what is there, but also for what is apparently missing. Only 11 of the 27 species that were collected at half of the study sites were collected at Aruza Abajo, whereas the median representation of these taxa at the other 15 sites was 20 of 27. One possible explanation for missing species is that they are replaced by novel taxa. The most likely example of taxon replacement is the antshrike Thamnophilus atrinucha, which tied for second place among most frequently collected species across the 16 sites (N=14), but appears to be replaced at Aruza Abajo by Thamnophilus nigriceps, an antshrike endemic to Eastern Panama and adjacent Northern Colombia. T. nigriceps was collected at just four sites in this study.
In general, our results agree with earlier studies that have highlighted the high betadiversity found in Panamanian biotic communities. Unlike Chust’s finding for Panamanian trees (Chust et al., 2006), there is no evidence that distance drives the turnover in bird species in central and Eastern Panama, agreeing with a more recent study of trees of central Panama (Jones et al., 2013). Often, beta diversity in Panamanian systems has been attributed to the strong rainfall gradient between Caribbean and Pacific Panama, especially in the canal area (e.g. phytophagous beetles: Ødegaard, 2006; and birds: Rompré, Robinson, Desrochers, & Angehr, 2007). Alternatively, Jones et al. (2013) suggest that soil type along with rainfall best predict species turnover of trees and ferns in central Panama. Interestingly, evidence that soils contribute to patterns of tropical bird species distributions comes from recent studies that document a unique avifauna in Amazonian white-sand forests (Alonso & Whitney 2003; Alonso, Metz, & Fine, 2013). Determining the cause of the distinctive avifauna assemblage in Western Darién province, and whether such differences can be found in other flora and faunal assemblages, remains to be determined. It is also important to note that variation in the bird assemblage among Panamanian field sites may be related to the extremely high degree of phylogeographic variation (i.e. within species genetic variation) that has been routinely observed in lowland Panamanian plants (Jones, Cerón Souza, Hardesty, & Dick, 2013) and animals (e.g. birds: González et al., 2003; Miller et al., 2008; Miller, Bermingham, Klicka, Escalante, & Winker, 2010; frogs: Wang, Crawford, & Bermingham, 2008; and bats: Clare, Lim, Fenton, & Hebert, 2011; Hauswaldt, Ludewig, Vences, & Pröhl, 2011).
A poor understanding of faunal dynamics in tropical areas not only limits our ability to understand patterns of tropical biodiversity but also impedes our response to applied scientific problems such as vector-borne tropical diseases. In the case of the fatal 2010 outbreak of equine encephalitis in Western Darién, recent phylogenetic evidence indicates that the virus is likely endemic in the area (S. Weaver, pers. comm.), yet as demonstrated by this study, even the most basic understanding of potential vertebrate host communities in the area is lacking. These findings highlight the continuing need to re-examine the depth of basic biodiversity knowledge even in supposedly well-known tropical systems, and demonstrates the value of applied research programs to improving our knowledge of tropical biodiversity patterns.
Acknowledgments
I would like to thank Panama’s Environmental Ministry (ANAM) for over 10 years of consistent support of scientific collecting in Panama. More than a dozen Latin American students and interns have dedicated their time to helping the STRI Bird Collection grow, and I am indebted to them. The STRI Bird Collection received supported from an NIH/NSF “Ecology and Evolution of Infectious Diseases” award from the Fogarty International Center 3R01-TW005869-05S1 and an inter-agency award from the US Centers for Disease Control (“Effect of Anthropogenic Climate Change on the Ecology of Zoonotic and Vector-borne Diseases”). Prior to 2007, the LSU Museum of Vertebrate Zoology, UNLV Marjorie Barrick Museum, University of Alaska Museum, and the US National Museum sponsored collecting trips in concert with STRI that form the basis of some of the data analyzed here. S. Weaver, University of Texas Medical Branch, provided information regarding his team’s ongoing research on equine encephalitic virus in Eastern Panama. J. Touchon, J. Loaiza, A. Crawford, and two anonymous reviewers, provided helpful comments on the manuscript.
References
Alonso, J. A., & Whitney, B. M. (2003). New distributional records of birds from white sand forests of the northern Peruvian Amazon, with implications for biogeography of northern South America. Condor, 105, 552-566. [ Links ]
Alonso, J. A., Metz, M. R., & Fine, P. V. (2013). Habitat specialization by birds in western Amazonian whitesand forests. Biotropica, 43, 365-372. doi:10.1111/ btp.12020 [ Links ]
Angehr, G. & Dean, R. (2010). The birds of Panama: a field guide. Ithaca, USA: Zona Tropical. [ Links ]
Arrigo, N., Adams, A., & Weaver, S. (2010). Evolutionary patterns of Eastern equine encephalitis virus in North versus South America suggest ecological differences and taxonomic revision. Journal of Virology, 84, 1014-1025. [ Links ]
Chao, A., Chazdon, R. L., Colwell, R. K., & Shen, T. J. (2005). A new statistical approach for assessing similarity of species composition with incidence and abundance data. Ecology Letters, 8, 148-159. [ Links ]
Chao, A. A., Chazdon R. L., Colwell, R. K., & Shen, T. J. (2006). Abundance-based similarity indices and their estimation when there are unseen species in samples. Biometrics, 62, 361-371. [ Links ]
Chust, G., Chave, J., Condit, R., Aguilar, S., Lao, S., & Pérez, R. (2006). Determinants and spatial modeling of tree β-diversity in a tropical forest landscape in Panama. Journal of Vegetation Science, 17, 83-92. [ Links ]
Clare, E. L., Lim, B. K., Fenton, M. B., & Hebert, P. D. N. (2011). Neotropical bats: estimating species diversity with DNA barcodes. PLoS ONE, 6, e22648. [ Links ]
Colwell, R. K. (2013). EstimateS: Statistical estimation of species richness and shared species from samples. Version 9. User’s Guide and application published at: http://purl.oclc.org/estimates. [ Links ]
González, M., Eberhard, J., Lovette, I., Olson, S., & Bermingham, E. (2003). Mitochondrial DNA phylogeography of the Bay Wren (Troglodytidae: Thryothorus nigricapillus) complex. Condor, 105, 228-238. [ Links ]
Hauswaldt, J. S., Ludewig, A. K., Vences, M., & Pröhl, H. (2011). Widespread co-occurrence of divergent mitochondrial haplotype lineages in a Central American species of poison frog (Oophaga pumilio). Journal of Biogeography, 38, 711-26. [ Links ]
Jones, M. M., Ferrier, S., Condit, R., Manion, G., Aguilar, S., & R. Pérez. (2013). Strong congruence in tree and fern community turnover in response to soils and climate in central Panama. Journal of Ecology, 101, 506-516. doi:10.1111/1365-2745.12053. [ Links ]
Jones, F. A., Cerón Souza, I., Hardesty, B. D., & Dick, C. W. (2013). Genetic evidence of Quaternary demographic changes in four rain forest tree species sampled across the Isthmus of Panama. Journal of Biogeography, 40, 720-731. doi:10.1111/jbi.12037. [ Links ]
Miller, M. J., Bermingham, E., Klicka, J., Escalante, P., & Winker, K. (2010). Neotropical birds show a humped distribution of within-population genetic diversity along a latitudinal transect. Ecology Letters, 13, 576-586. [ Links ]
Miller, M. J., Bermingham, E., Klicka, J., Escalante, P., do Amaral, F. S. R., Weir, J. T., & Winker, K. (2008). Out of Amazonia again and again: episodic crossing of the Andes promotes diversification in a lowland forest flycatcher. Proceedings of the Royal Society B: Biological Sciences, 275, 1133-1142. [ Links ]
Navia-Gine, W.G., Loaiza, J.R., & Miller, M.J. Mosquitohost interactions during and after an outbreak of equine viral encephalitis in eastern Panama. PLOS ONE DOI: 10.1371/journal.pone.0081788. [ Links ]
Ødegaard, F. (2006). Host specificity, alpha-and beta-diversity of phytophagous beetles in two tropical forests in Panama. Biodiversity and Conservation, 15, 83-105. [ Links ]
Pyke, G. H. & Ehrlich, P. R. (2010). Biological collections and ecological/environmental research: a review, some observations and a look to the future. Biological Reviews, 85, 247-266. [ Links ]
Ridgely, R. & Gwynne, J. A. (1989). A guide to the birds of Panama with Costa Rica, Nicaragua, and Honduras. Princeton, USA: Princeton University Press. [ Links ]
Rompré, G., Robinson, W. D., Desrochers, A., & Angehr, G. (2007). Environmental correlates of avian diversity in lowland Panama rain forests. Journal of Biogeography, 34, 802-815. [ Links ]
Siegel, D. C. & Olson, S. L. (2008). The birds of the Republic of Panama. Part 5. Gazetteer and bibliography. Shipman, USA: Buteo Books. [ Links ]
Wang, I. J., Crawford, A. J., & Bermingham, E. (2008). Phylogeography of the Pygmy Rain Frog (Pristimantis ridens) across the lowland wet forests of isthmian Central America. Molecular Phylogenetics and Evolution, 47, 992-1004. [ Links ]
Winker, K. 2004. Natural history museums in a postbiodiversity era. BioScience, 54, 455 459. [ Links ]
Yasué, M., Feare, C.J., Bennun, L. & Fiedler, W. (2006). The epidemiology of H5N1 avian influenza in wild birds: why we need better ecological data. BioScience, 56, 923-929. [ Links ]
Alonso, J. A., Metz, M. R., & Fine, P. V. (2013). Habitat specialization by birds in western Amazonian whitesand forests. Biotropica, 43, 365-372. doi:10.1111/ btp.12020 [ Links ]
Angehr, G. & Dean, R. (2010). The birds of Panama: a field guide. Ithaca, USA: Zona Tropical. [ Links ]
Arrigo, N., Adams, A., & Weaver, S. (2010). Evolutionary patterns of Eastern equine encephalitis virus in North versus South America suggest ecological differences and taxonomic revision. Journal of Virology, 84, 1014-1025. [ Links ]
Chao, A., Chazdon, R. L., Colwell, R. K., & Shen, T. J. (2005). A new statistical approach for assessing similarity of species composition with incidence and abundance data. Ecology Letters, 8, 148-159. [ Links ]
Chao, A. A., Chazdon R. L., Colwell, R. K., & Shen, T. J. (2006). Abundance-based similarity indices and their estimation when there are unseen species in samples. Biometrics, 62, 361-371. [ Links ]
Chust, G., Chave, J., Condit, R., Aguilar, S., Lao, S., & Pérez, R. (2006). Determinants and spatial modeling of tree β-diversity in a tropical forest landscape in Panama. Journal of Vegetation Science, 17, 83-92. [ Links ]
Clare, E. L., Lim, B. K., Fenton, M. B., & Hebert, P. D. N. (2011). Neotropical bats: estimating species diversity with DNA barcodes. PLoS ONE, 6, e22648. [ Links ]
Colwell, R. K. (2013). EstimateS: Statistical estimation of species richness and shared species from samples. Version 9. User’s Guide and application published at: http://purl.oclc.org/estimates. [ Links ]
González, M., Eberhard, J., Lovette, I., Olson, S., & Bermingham, E. (2003). Mitochondrial DNA phylogeography of the Bay Wren (Troglodytidae: Thryothorus nigricapillus) complex. Condor, 105, 228-238. [ Links ]
Hauswaldt, J. S., Ludewig, A. K., Vences, M., & Pröhl, H. (2011). Widespread co-occurrence of divergent mitochondrial haplotype lineages in a Central American species of poison frog (Oophaga pumilio). Journal of Biogeography, 38, 711-26. [ Links ]
Jones, M. M., Ferrier, S., Condit, R., Manion, G., Aguilar, S., & R. Pérez. (2013). Strong congruence in tree and fern community turnover in response to soils and climate in central Panama. Journal of Ecology, 101, 506-516. doi:10.1111/1365-2745.12053. [ Links ]
Jones, F. A., Cerón Souza, I., Hardesty, B. D., & Dick, C. W. (2013). Genetic evidence of Quaternary demographic changes in four rain forest tree species sampled across the Isthmus of Panama. Journal of Biogeography, 40, 720-731. doi:10.1111/jbi.12037. [ Links ]
Miller, M. J., Bermingham, E., Klicka, J., Escalante, P., & Winker, K. (2010). Neotropical birds show a humped distribution of within-population genetic diversity along a latitudinal transect. Ecology Letters, 13, 576-586. [ Links ]
Miller, M. J., Bermingham, E., Klicka, J., Escalante, P., do Amaral, F. S. R., Weir, J. T., & Winker, K. (2008). Out of Amazonia again and again: episodic crossing of the Andes promotes diversification in a lowland forest flycatcher. Proceedings of the Royal Society B: Biological Sciences, 275, 1133-1142. [ Links ]
Navia-Gine, W.G., Loaiza, J.R., & Miller, M.J. Mosquitohost interactions during and after an outbreak of equine viral encephalitis in eastern Panama. PLOS ONE DOI: 10.1371/journal.pone.0081788. [ Links ]
Ødegaard, F. (2006). Host specificity, alpha-and beta-diversity of phytophagous beetles in two tropical forests in Panama. Biodiversity and Conservation, 15, 83-105. [ Links ]
Pyke, G. H. & Ehrlich, P. R. (2010). Biological collections and ecological/environmental research: a review, some observations and a look to the future. Biological Reviews, 85, 247-266. [ Links ]
Ridgely, R. & Gwynne, J. A. (1989). A guide to the birds of Panama with Costa Rica, Nicaragua, and Honduras. Princeton, USA: Princeton University Press. [ Links ]
Rompré, G., Robinson, W. D., Desrochers, A., & Angehr, G. (2007). Environmental correlates of avian diversity in lowland Panama rain forests. Journal of Biogeography, 34, 802-815. [ Links ]
Siegel, D. C. & Olson, S. L. (2008). The birds of the Republic of Panama. Part 5. Gazetteer and bibliography. Shipman, USA: Buteo Books. [ Links ]
Wang, I. J., Crawford, A. J., & Bermingham, E. (2008). Phylogeography of the Pygmy Rain Frog (Pristimantis ridens) across the lowland wet forests of isthmian Central America. Molecular Phylogenetics and Evolution, 47, 992-1004. [ Links ]
Winker, K. 2004. Natural history museums in a postbiodiversity era. BioScience, 54, 455 459. [ Links ]
Yasué, M., Feare, C.J., Bennun, L. & Fiedler, W. (2006). The epidemiology of H5N1 avian influenza in wild birds: why we need better ecological data. BioScience, 56, 923-929. [ Links ]
*Correspondencia a:
Matthew J. Miller. Smithsonian Tropical Research Institute, Apartado Postal 0843-03092, Panamá, República de Panamá; millerma@si.edu
1. Smithsonian Tropical Research Institute, Apartado Postal 0843-03092, Panamá, República de Panamá; millerma@si.edu
Received 22-vii-2013. Corrected 25-X-2013. Accepted 28-Xi-2013.


{kind=link}
{kind=link}
{kind=link}