










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.62 n.2 San José Apr./Jun. 2014
Yasuní – a hotspot for jaguars Panthera onca (Carnivora: Felidae)? Camera-traps and jaguar activity at Tiputini Biodiversity Station, Ecuador
Yasmí- Un hotspot para jaguares Panthera onca (Carnivora: Felidae)? Cámaras trampa y actividad del jaguar en la Estación de Biodiversidad Tiputini, Ecuador
Yasmí- Un hotspot para jaguares Panthera onca (Carnivora: Felidae)? Cámaras trampa y actividad del jaguar en la Estación de Biodiversidad Tiputini, Ecuador
John G. Blake1*, Diego Mosquera2*, Jaime Guerra3*, Bette A. Loiselle4*,5, David Romo6* & Kelly Swing7*
*Dirección para correspondencia:
Abstract
Jaguars (Panthera onca) are the largest predator in lowland forests of Amazonia but there have been few studies on their occurrence and activity in such forests. Here, we used camera traps to document the occurrence and activity of jaguars within a local area (~650ha) of lowland forest of Eastern Ecuador, over two sample periods (2005-2008, 7 222 trap days; 2010-2012, 6 199 trap days). We accumulated 151 independent photos of jaguars (189 total photographs) that represented 21 different individuals, including 11 males (114 photographs), seven females (32 photographs), and three that could not be assigned to a sex. Individual jaguars varied in the number of months they were recorded in the area; ten were photographed in only one month; five were photographed over periods of 8 to 22 months; and five from 45 to 81 months. Capture rates across all camera stations averaged 10.6/1 000 trap days; capture rates did not differ between the two sample periods. Male jaguars were more active during the day (06:00am-18:00pm; 71% of photographs), whereas females were equally active during the day and night. Monthly activity was variable but showed no consistent pattern. Although the study area is much smaller than typical home ranges of jaguars, the area is clearly visited by a large number of different individuals, some of whom repeatedly visit the area, indicating that it forms part of their home range. Other individuals likely were simply passing through the area. Based on the number of jaguars recorded during this study, it is clear that the region is an important area for conservation. Continued protection will be needed to ensure that populations of jaguars and other species remain viable. Rev. Biol. Trop. 62 (2): 689-698. Epub 2014 June 01.
Key words: Amazonia, daily activity, lowland forest, spatial variation, temporal variation.
Resumen
El jaguar (Panthera onca) es el depredador más grande de los bosques bajos de la Amazonía, pero existen pocos estudios sobre su presencia y actividad en los mismos. En este estudio utilizamos cámaras trampa para documentar la presencia y actividad de jaguares dentro de una área de bosque bajo de la Amazonía este de Ecuador (~650 ha) en dos períodos (2005-2008, 7 222 trampas noche; 2010-2012, 6 199 trampas noche). Acumulamos 151 fotos independientes de jaguares (189 fotografías en total) que representaron 21 individuos, incluyendo 11 machos (114 fotos), siete hembras (32 fotos) y tres a los que no se les pudo asignar sexo. Los jaguares individuales variaron en el número de meses en que fueron registrados en el área; diez fueron fotografiados solo en un mes; cinco fueron fotografiados en un periodo entre 8 y 22 meses; y cinco jaguares entre 45 y 81. Las tasas de captura a través de las estaciones de trampeo promediaron 10.6/1 000 trampas noche; las tasas de captura no presentaron diferencias en los dos períodos de muestreo. Los jaguares machos fueron más activos durante el día (06:00am-18:00pm; 71% de las fotografías) mientras que las hembras fueron igualmente activas tanto en el día como en la noche. La actividad mensual fue variable pero no mostró un patrón consistente. Aunque el área de estudio es mucho más pequeña que el rango de vida típico para un jaguar, el área es claramente visitada por un alto número de individuos diferentes, algunos de los cuales visitan repetidamente el lugar, indicando que este forma parte de su rango de vida. Es probable que otros individuos solamente sean transeúntes del área.
Palabras clave: actividad diaria, Amazonia, bosque lluvioso, variación espacial, variación temporal.
The jaguar (Panthera onca) is the largest predator in Central and South America and, as a consequence, fulfills an important role in the functioning of many different tropical ecosystems (Terborgh, 1988). Tropical felids are, in general, difficult to study, particularly in closed-canopy forest habitats, given their low densities, large home ranges, elusive nature, and frequent nocturnal behavior. Yet, information on such felids and other predators are clearly needed-often to assess the conservation status of a particular area (Maffei, Cuéllar & Noss, 2004; Silver et al., 2004).
Camera traps have proven to be an useful technique for studying Neotropical felids, including ocelots (Leopardus pardalis), margays (Leopardus wiedii), pumas (Puma concolor), and jaguars (Kelly, 2003; Silver et al., 2004; Maffei, Noss, Silver, & Kelly, 2011; Blake et al., 2012). Although various studies have used camera traps to evaluate jaguar populations in a variety of different habitats, including rain forests, dry forests, and grasslands (Kelly, 2003; Scognamillo, Maxit, Sunquist, & Polisar, 2003; Silveira, Jácomo, & Diniz-Filho, 2003; Maffei et al., 2004; Silver et al., 2004; Soisalo & Cavalcanti, 2006), few studies have been conducted in lowland wet forests of the Amazon (Bolivia: Wallace, Gómez, Ayala, & Espinoza, 2003; Silver et al., 2004; Ecuador: Espinoza-Andrade, 2012; Peru: Tobler, Carrillo-Percastegui, Zúñiga Hartley, & Powell, 2013).
Studies on jaguars, and other large cats, typically are conducted over large areas (Silver et al., 2004), given their large home ranges (Rabinowitz & Nottingham, 1986; Crawshaw & Quigley, 1991; Maffei et al., 2004) and generally low densities (Kelly, 2003; Maffei et al., 2004, 2011). Home ranges of male and female jaguars are known to overlap (e.g., Soisalo & Cavalcanti, 2006) so it is possible that multiple individuals might co-occur in a given area of forest. Yet, we know relatively little about the extent of temporal and spatial overlap of individual jaguars at more local scales, particularly in lowland Amazonian forest (Emmons, 1987). Variation in local abundance might reflect differences in habitat, prey populations, hunting pressures, or other human activities.
Here we used camera traps to document occurrence of jaguars within Tiputini Biodiversity Station, a 650ha research station located in undisturbed forest of Eastern Ecuador. The station is part of Yasuní Biosphere Reserve, and adjacent to Yasuní National Park, one of the most biologically rich regions in the world (Bass et al., 2010). We use photographic records, collected over ~7 years, longer than most studies, to examine numbers of individual jaguars, activity patterns, and temporal and spatial overlap within the boundaries of the station. It is important to note that we are not attempting to estimate density; the area is far too small to allow such estimates. Rather, we are evaluating the degree to which multiple individuals overlap in their use of a relatively small area of forest.
Materials and methods
Study site: We conducted our research at Tiputini Biodiversity Station (TBS), Orellana Province, Ecuador (~0o38’ S, 76o09’ W, 190-270m asl). TBS was founded in 1994 by the Universidad San Francisco de Quito (USFQ) on a tract of undisturbed lowland rainforest within the Yasuní Biosphere Reserve. Hunting is rare in the vicinity of TBS (personal observations), with little impact on the fauna (Blake, Mosquera, & Salvador, 2013). The closest Kichwa community is ~17km north of the station; jaguars moving in that direction might, on occasion, encounter hunters but are much less likely to do so when moving in any other direction. The station and nearby areas contain a variety of habitats including terra firme and várzea forest, palm swamps and other wetlands, as well as areas of natural succession that follow treefalls, windthrows, or other natural disturbances. The mean annual precipitation at Yasuní Research Station, approximately 30km WSW of TBS, is about 3 100mm (Blake et al., 2012). Rainiest months are from April through June; January and August can be relatively dry.
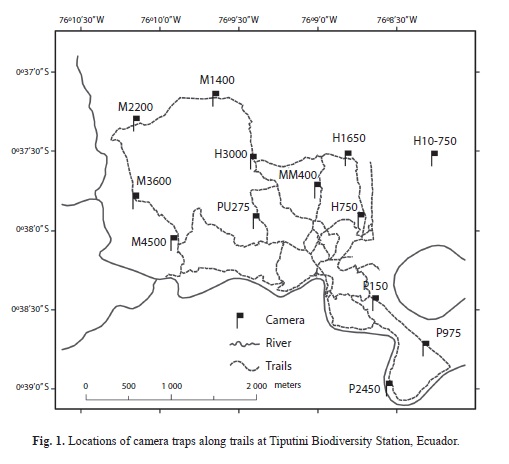
Camera trapping: Cameras triggered by an infrared heat-and-motion detector were deployed during two sample periods: mid- January 2005 - August 2008; February 2010 - January 2012. We used film‑based camera traps (Highlander Photoscout, PTC Technologies) during the first sampling period and digital camera traps (Cuddeback Capture) during the second. Pairs of cameras were located approximately 1-1.2km apart along narrow (generally <1m wide) preexisting trails within terra firme forest (Fig. 1). Cameras were placed at locations that showed evidence of jaguar activity (e.g., tracks) or where occurrence was deemed likely (based on topography and local knowledge). Two cameras were placed in each spot (eight locations during the first period; ten during the second), on opposite sides of the trail, approximately 0.50 to 0.75m off the ground, and about 2 to 3m back from the trail edge. We set cameras with a minimum time between photographs of 3 or 5min. Cameras remained continuously activated (except when malfunctions occurred); date and time were automatically stamped on each photograph. Cameras were checked at approximately monthly intervals to replace film, change SD cards, and check batteries. All images were labeled with location, camera, date, and time.
Analyses: We summarized images by individual, hour, and date. Individuals were identified on the basis of distinctive coat patterns. We classified photographs as belonging to independent records if more than 30min had elapsed between consecutive photographs of the same individual at a given location (see Blake et al., 2011). Activity was evaluated in terms of number of photographs, percentage of photographs or photographs/1 000 trap days, depending on the analysis. We calculated number of trap‑days from the time the camera was placed in operation to the time the film (or SD card) was replaced (i.e., starting a new sequence) or last photograph was taken if a malfunction had occurred (based on the date and time stamp on the photographs). We classified records by hour, starting at midnight, to examine hourly patterns of activity, and by month to examine seasonal activity irrespective of hour. Photographs were categorized by location to allow comparison of spatial distribution patterns of individuals and capture rates.
Several locations were not well sampled because of problems with cameras; these problems precluded calculations of numbers of days the cameras were in operation. Consequently, data from these sites were not used in analyses that required calculation of capture rates: MM400 and M1400 for both periods, and P975 and P2450 for the second period (see Fig. 1 for locations). Finally, an increasing number of cameras began to fail in 2008 so capture rates do not include data from 2008. In all cases, data from these sites were included in analyses of hourly variation in activity and to document presence in the study area (e.g., number of months over which an individual occurred within the study area).
We used a paired t-test to compare capture rates between the first and second sample periods; seven sites sampled well in both periods were included in the analysis. Similarly, we used a paired t-test to compare numbers of records for males and females by month. We used a two sample t-test to compare capture rates between two different areas of the station. Finally, we used correlation analyses (Pearson’s r) to determine if capture rates at a given location were correlated between the two sample periods (i.e., to determine if spatial differences in capture rates were consistent over time).
Results
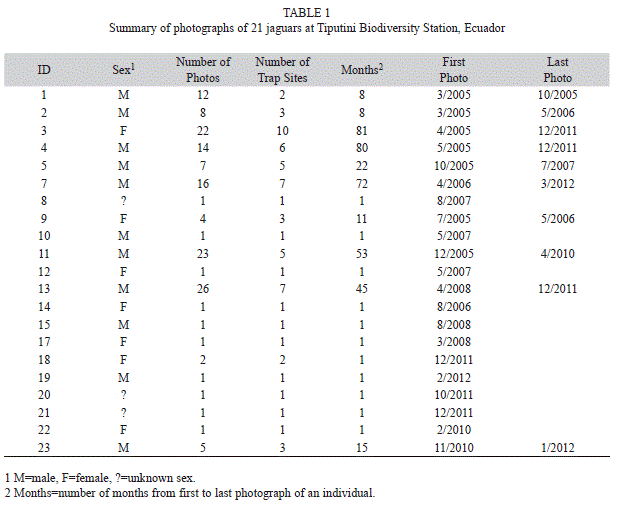
General summary: We accumulated 150 independent records of jaguars (189 total photographs) that represented 21 individuals (plus one photograph that could not be assigned to a specific individual), including 11 males (114 photographs with eight individuals photographed at least five times), seven females (32 photographs, 22 of the same individual), and three that could not be assigned to a sex (Table 1). One individual (unknown sex) was melanistic. There were 98 records during the first sample period (15 individuals) and 52 during the second (11 individuals); five individuals were photographed during both periods. Number of photographs per individual ranged from one (ten individuals) to more than 20 (three individuals) (Table 1). Capture periods (number of months from first to last photograph) ranged from one month (11 individuals) to more than 40 months (five individuals); one male and one female were photographed over an 80 month period each (Table 1).
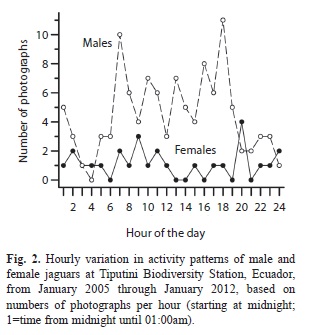
Temporal activity patterns: Jaguars were active both during daylight hours (06:00-18:00; 67% of photographs) and during the night. Males were more active during the day (71% of photographs), with pronounced peaks of activity from 06:00-07:00am and from 17:00- 18:00pm (Fig. 2). Females, in contrast, were equally active during the day (52% of photographs) and during the night. Highest peaks in activity were from 08:00-09:00am and from 19:00-20:00pm (Fig. 2), hours when male activity declined. Monthly activity showed substantial variation both for males and females (coefficient of variation in number of photographs per month: 86.5 for females and 49.3 for males) but was higher for males in all months except August, when there were five photos of each sex (paired t-test=4.31, d.f.=11, p=0.0012). Female capture rate showed a bimodal pattern, with peaks in June and December; males showed no clear seasonal pattern (Fig. 3). Monthly capture rates were not correlated between sexes (r=-0.17, p>0.20).
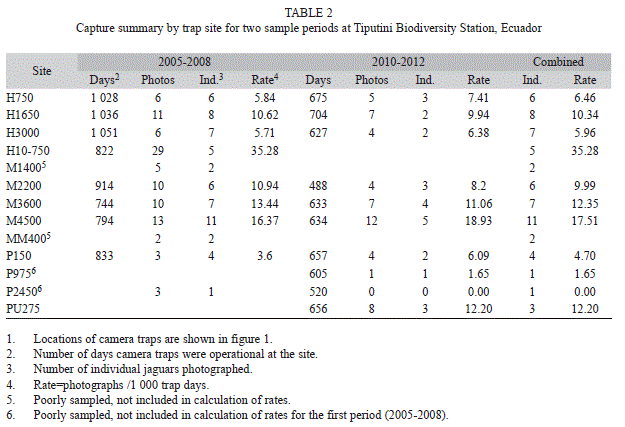
Spatial activity patterns: Jaguars were photographed at all locations during at least one sample period but number of photographs varied substantially among sites (Table 2, Fig. 1). Capture rates varied from a low of 0 at P2450 in the second sample period to a high of 35.3 at HP10-750 per 1 000 trap days during the first sample period (Table 2). There was, however, no difference in capture rates between sample periods (seven locations well sampled in both periods, paired t-test=0.26, d.f.=6, p=0.89); rates at a given location were highly correlated between periods (r=0.88, p<0.01). Three sites (P150, P975, P2450) located within a peninsular area of the station (Fig. 1) had lower capture rates than those elsewhere in the station during the second sample period (means ± SE for seven sites away from and three sites within the peninsula, 10.59±1.59 and 2.58±1.82, respectively; t-test=2.92, d.f.=8, p=0.019). Number of individuals recorded per site ranged from one to 11; in both years, one site (M4500) had the greatest number of individuals recorded (Table 2). Activity of individuals varied substantially among sites (Table 1). Although more males were recorded overall, one female was recorded at more sites (ten) than any individual male (two males recorded at seven sites each).
Discussion
During the period from January 2005 through January 2012, at least 21 individual jaguars are known to have been present, at one time or another, within the 650ha of Tiputini Biodiversity Station. The total number of jaguars that visited the station area within this time frame likely was higher, because 2008 was not well sampled and no cameras were deployed in 2009. On the other hand, given the relatively small area and close spacing of the cameras and considering that jaguars typically range over areas much larger than the station (Maffei et al., 2011; Tobler et al., 2013), it is likely that most individuals had a reasonably high probability of being detected when present (and when cameras were functioning properly).
Camera traps have been used in many previous studies of jaguars; in a recent review, Maffei et al. (2011) mentioned 83 different efforts to sample jaguars with cameras. These attempts to estimate jaguar abundance or density have relied on large study areas (range of 24 to 555km2 for camera polygons; 54 to 938km2 for effective sample areas). The wide range in study areas likely is a function both of logistics (it is hard to adequately sample very large areas) and geographic and habitat variation in jaguar densities and home range sizes (e.g., 10 to 40km2 home range in tropical moist forest in Belize, Rabinowitz & Nottingham, 1986; 52 to 176km2 in Pantanal grasslands of Brazil, Crawshaw & Quigley, 1991; Soisalo & Cavalcanti, 2006). In contrast, we examined the question of how many jaguars might spatially and temporally overlap within a relatively small (by jaguar home range standards) area. Home ranges for jaguars in lowland forests of Southwestern Peru (Tobler et al., 2013), ranged from 130 to 283km2 for females and males, respectively, much larger than the area encompassed by our traps.
Previous studies have demonstrated that multiple individuals may overlap in their use of space (Emmons, 1987; Soisalo & Cavalcanti, 2006); results of the present study support this conclusion, given that 11 males and seven females were recorded within the boundaries of TBS (i.e., an area much smaller than the smallest home range reported for jaguars). Multiple individuals also were photographed at the same trap locations and capture rates at locations were consistent over time, suggesting that different areas of the station were preferred as travel routes. In addition to the spatial overlap, individuals also overlapped on a daily basis. On two occasions, two jaguars were photographed the same day (two males; one male and one female); over two to five day periods, there were at least ten occasions when two to four individuals were present. Emmons (1987), reported three to five jaguars using the 7.5km2 area of Cocha Cashu Biological Station, Peru; two big cats (not specified whether jaguar or puma) were present on 20 out of 199 days and three were present in one day.
Some of the individuals photographed at TBS likely were simply transients (e.g., possibly dispersing individuals) or individuals whose home ranges only peripherally include parts of TBS (i.e., so that they may rarely encounter cameras at TBS). In contrast, other individuals clearly included TBS as a regular part of their range, with repeated occurrences recorded over months and years. One female, for example, was photographed 22 times at ten different locations within TBS, over an 81 month period and clearly is resident in the area. Similarly, four males were recorded multiple times over 45 to 80 month periods, suggesting that they were residents too. Overall, more males than females were repeatedly photographed at TBS. Males typically have larger home ranges than females (Rabinowitz & Nottingham, 1986; Soisalo & Cavalcanti, 2006; Tobler et al., 2013) so it is possible that home ranges of more males overlap a given area of forest, leading to more photographs and more repeated photographs of individuals. Most previous studies have recorded more males than females, with sex ratios (males:females) ranging from 1.5:1 to 4:1 and even 9:0 (Maffei et al., 2011) although some studies reported more females (five out of 39 studies reported in Maffei et al., 2011). Espinosa-Andrade (2012) reported a ratio of 2.6:1 from studies in Yasuní, somewhat higher than the one found in our study for the same region (1.6:1).
Males and females also differed in both hourly and monthly patterns of activity. Jaguars, in general, tend to be both diurnal and nocturnal in their activity, although their patterns of activity can vary geographically [e.g., primarily nocturnal in tropical moist forest, Belize (Weckel, Giuliano & Silver, 2006), Venezuelan llanos (Scognamillo et al., 2003), and dry forest, Bolivia (Maffei et al., 2011); nocturnal and crepuscular at four sites in Brazil (Foster et al., 2013); primarily diurnal in Ecuador, this study; see also Blake et al., 2012]. Few previous descriptions of hourly activity patterns have, however, separated males and females, as in this study where males were more diurnal in behavior than were females. Male and female activity patterns did not differ in dry forests of Bolivia (Romero-Muñoz, Maffei, Cuéllar & Noss, 2010). Differences in activity patterns, when they occur, might be one method to reduce possibilities for competition between sexes and/or to reduce the danger to cubs (i.e., by reducing the chances of encountering males that might kill cubs; Soares et al., 2006). No cubs were recorded in photographs during this study, which is consistent with most published studies [but see Maffei et al. (2011) for a summary of unpublished studies that do report cubs]. On the other hand, a cub has been seen with a female near TBS, so lack of photographs does not mean that cubs are not present. It is also possible that if cubs were following a female, they might not be photographed, given the time delay between photographs.
Variation in activity of preferred prey may account for some of the geographic variation in activity. Peccaries (e.g., Pecari tajacu, collared peccary), preferred prey of jaguars in some studies (e.g., Emmons, 1987; Weckel et al., 2006), are more nocturnal in Belize (Weckel et al., 2006) than in our study site (Blake et al., 2011, 2012). If sexes differ in preferred prey, this might explain some of the differences in hourly activity of males and females seen in this study. Previous studies on jaguar diets have not examined differences between sexes. Variation in prey abundance or activity may also influence monthly activity. Seasonal variation in distribution patterns of peccaries (Tayassu pecari, white-lipped peccary; collared peccary) apparently can influence the spatial distribution patterns of jaguars and pumas, respectively (Mendes-Pontes & Chivers, 2007). Herds of white-lipped peccaries at TBS show marked variation in abundance (personal observations) so it is possible that their movement patterns might affect the occurrence of jaguars within the station as well. Variation in abundance of other prey of jaguars, such as red brocket deer (Mazama americana) and armadillos (Dasypus novemcinctus), that are common at TBS (Blake et al. 2011, 2012), also might influence the distribution patterns of jaguars.
Capture rates of jaguars also vary substantially among studies but do not show a clear relationship with estimates of density. Overall capture rate in our study was intermediate compared to those of many other studies (see Maffei et al., 2011 for a review) that were conducted over much larger areas. Our study was conducted over a longer period of time than previous ones, which allowed us to compare capture rates over time. Although there was some variation among years, overall capture rates did not differ between the two periods included here. This lack of difference suggests that jaguar abundance, or at least activity, may be relatively stable in the area around TBS, at least for the period of this study (2005 to 2012). If so, this suggests that human activities (e.g., hunting, oil exploration) have not (yet) had an increasing impact on the fauna of the area.
At a smaller scale, capture rates did show some significant variation among the specific camera-trap locations within TBS. In part, this variation may have been related to individual variation in behavior. One location (HP10-750) experienced a much higher capture rate than other sites but also recorded fewer individuals than some sites that had lower overall captures. A large proportion of the photographs at HP10- 750 were of two individuals (21 of 29 photos) who repeatedly used this transect (part of a 100-ha study plot) as a travel route. In contrast, cameras located along the Parahuaco trail (P150, P975, P2450), which is located within a peninsular area of the station formed by a bend in the Tiputini River, experienced generally lower capture rates than other areas. Of the three sites along this trail, the two farthest ones had the lowest capture rates, suggesting that fewer jaguars were traveling in the area bounded by the river.
Considering conservation implications, Yasuní Biosphere Reserve (Ministerio del Ambiente del Ecuador, 2010) has been recognized as the most biologically diverse region in all Amazonia (Bass et al., 2010). The reserve also is home to indigenous groups, such as the Waorani and Kichwa, who depend on these forests for much of their subsistence. Much of the reserve, including parts of Yasuní National Park (YNP) also is subject to human disturbance, such as oil exploration and extraction, illegal logging, and hunting (Bass et al., 2010). Our study area is located in the border of YNP, approximately 45km from the ITT (Ishpingo-Tam bococha-Tiputini) oil concession within YNP, which has been the center of a unique conservation proposal (Larrea & Warnars, 2009; Finer, Moncel & Jenkins, 2010); that proposal has, however, recently been withdrawn. Given that top predators often serve as umbrella species in developing conservation plans, data such as those generated in the current study can help demonstrate the quality and importance of protected areas.
Although our study does not provide an estimate of actual density for jaguars in the area, it does demonstrate that jaguars are reasonably abundant and regularly use the area. Repeated occurrences of the same individuals within the boundaries of TBS also indicate that home ranges of jaguar overlap both spatially and temporally. Frequent overlap may further suggest that prey populations are high. Many of the prey upon which jaguars depend (e.g., deer, peccaries) also are frequent targets of human hunters (Mena, Stallings, Regalado & Cueva, 2000; Franzen, 2006; Bass et al., 2010). Less than 17km by foot to the North of TBS is the Kichwa community of El Eden and an oil field that is part of Ecuador’s Block 15; Kichwa hunters may, at times, come close to the station in search of wild protein. Similarly, the nearest Waorani community is about 30km West of the station and its hunters are occasionally encountered on the Tiputini River near TBS. Yet, an abundance of top predators suggests that prey populations also are abundant, as our previous studies indicated (Blake et al., 2011, 2012), and would suggest that hunting pressure is relatively slight in the area (Blake et al., 2013). If prey populations were reduced as a consequence of increased hunting activities, however, we could expect to see declines in predator populations as well (Espinosa-Andrade, 2012). Further information on population levels of potential prey of jaguars, in areas with and without human pressures, would be especially valuable (Suárez, Zapata-Rios, Utreras, Strindberg & Vargas, 2013). Yasuní represents an unique place because of its biodiversity, size, and lack of human intervention in most of the area. Continued protection will be needed to ensure that populations of jaguars and other species remain viable.
Acknowledgments
We thank the many staff and volunteers who helped check cameras during this study, particularly Rene Torres, Eduardo Gutierrez, Juanpablo Muñoz, Franklin Narvaez, Ramiro SanMiguel, and Jose Macanilla. We also appreciate the help of Consuelo de Romo in facilitating our work at Tiputini and the many staff that made working there such a pleasure. We thank Andrew Noss for helpful comments to improve this manuscript. Support for this study was provided by National Geographic Society, Universidad San Francisco de Quito, Tiputini Biodiversity Station, University of Missouri - St. Louis, University of Florida, and Walton Expeditions. Work at Tiputini Biodiversity Station was conducted in accordance with research permit No. 0005-FAU-MAE-DPOPNY, Ministerio del Ambiente, Ecuador.
References
Bass, M. S., Finer, M., Jenkins, C. N., Kreft, H., Cisneros-Heredia, D. F., McCracken, S. F., Pitman, N. C. A., English, P. H., Swing, K., Villa, G., Di Fiore, A., Voigt, C. C., & Kunz, T. H. (2010). Global conservation significance of Ecuador’s Yasuní National Park. PloS ONE 5, e8767. [ Links ]
Blake, J. G., Mosquera, D., Guerra, J., Loiselle, B. A., Romo, D., & Swing, K. (2011). Mineral licks as diversity hotspots in lowland forest of eastern Ecuador. Diversity, 3, 217-234. [ Links ]
Blake, J. G., Mosquera, D., Loiselle, B. A., Swing, K., Guerra, J., & Romo, D. (2012). Temporal activity patterns of terrestrial mammals in lowland rainforest of eastern Ecuador. Ecotropica, 18, 137-146. [ Links ]
Blake, J. G., Mosquera, D., & Salvador, J. (2013). Use of mineral licks by mammals and birds in hunted and non-hunted areas of Yasuní National Park, Ecuador. Animal Conservation, 16, 420-437. [ Links ]
Crawshaw Jr., P. G. & Quigley, H. B. (1991). Jaguar spacing, activity and habitat use in a seasonally flooded environment in Brazil. Journal of Zoology, 223, 357-370. [ Links ]
Emmons, L. H. (1987). Comparative feeding ecology of felids in a Neotropical rainforest. Behavioral Ecology and Sociobiology, 20, 271-283. [ Links ]
Espinosa-Andrade, S. R. (2012). Road development, bushmeat extraction and jaguar conservation in Yasuni Biosphere Reserve -Ecuador. (Doctoral dissertation). University of Florida, Gainesville, Florida, USA. [ Links ]
Finer, M., Moncel, J., & Jenkins, C. N. (2010). Leaving the oil under the Amazon: Ecuador’s Yasuní-ITT initiative. Biotropica, 42, 63-66. [ Links ]
Foster, V. C., Sarmento, P., Sollmann, R., Tôrres, N., Jácomo, A. T. A., Negrões, N., Fonseca, C., & Silveira, L. (2013). Jaguar and puma activity patterns and predator-prey interactions in four Brazilian biomes. Biotropica, 45, 373-379. [ Links ]
Franzen, M. (2006). Evaluating the sustainability of hunting: a comparison of harvest profiles across three Huaorani communities. Environmental Conservation, 33, 1-10. [ Links ]
Kelly, M. J. (2003). Jaguar monitoring in the Chiquibul Forest, Belize. Caribbean Geography, 13, 19-32. [ Links ]
Larrea, C. & Warnars, L. (2009). Ecuador’s Yasuni-ITT initative: Avoiding emissions by keeping petroleum underground. Energy and Sustainable Development, 13, 219-223. [ Links ]
Maffei, L., Cuéllar, E., & Noss, A. (2004). One thousand jaguars (Panthera onca) in Bolivia’s Chaco? Camera trapping in the Kaa-Iya National Park. Journal of Zoology, 262, 295-304. [ Links ]
Maffei, L., Noss, A. J., Silver, S. C., & Kelly, M. J. (2011). Abundance/density case study: Jaguars in the Americas. In A. F. O’Connell, J. D. Nichols & K. U. Karanth (Eds.), Camera traps in animal ecology - methods and analyses, (pp. 119-144). New York, USA: Springer. [ Links ]
Mena, V. P., Stallings, J. R., Regalado, J. B., & Cueva, R. L. (2000). The sustainability of current hunting practices by the Huaorani. In J. G. Robinson & E. I. Bennett (Eds.), Hunting for sustainability in tropical forests (pp. 57-78). New York, USA: Columbia University Press. [ Links ]
Mendes-Pontes, A. R. & Chivers, D. J. (2007). Peccary movements as determinants of the movements of large cats in Brazilian Amazonia. Journal of Zoology, 273, 257-265. [ Links ]
Ministerio del Ambiente del Ecuador. (2010). Reservas de Biosfera del Ecuador: lugares excepcionales. Quito, Ecuador: GTZ/GESORENDED-WCS- NCI-UNESCO. [ Links ]
Rabinowitz, A. R. & Nottingham Jr., B. G. (1986). Ecology and behaviour of the jaguar (Panthera onca) in Belize, Central America. Journal of Zoology, 210, 149-159. [ Links ]
Romero-Muñoz, A., Maffei, L., Cuéllar, E., & Noss, A. J. (2010). Temporal separation between jaguar and puma in the dry forests of southern Bolivia. Journal of Tropical Ecology, 26, 303-311. [ Links ]
Scognamillo, D. I., Maxit, E., Sunquist, M., & Polisar, J. (2003). Coexistence of jaguar (Panthera onca) and puma (Puma concolor) in a mosaic landscape in the Venezuelan llanos. Journal of Zoology, 259, 269-279. [ Links ]
Silveira, L., Jácomo, A. T. A., & Diniz-Filho, J. A. F. (2003). Camera trap, line transect census and track surveys: a comparative evaluation. Biological Conservation, 114, 351-355. [ Links ]
Silver, S. C., Ostro, L. E. T., Marsh, L. K., Maffei, L., Noss, A. J., Kelly, M. J., Wallace, R. B., Gómez, H., & Ayala, G. (2004). The use of camera traps for estimating jaguar Panthera onca abundance and density using capture/recapture analysis. Oryx, 38, 148-154. [ Links ]
Soisalo, M. K. & Cavalcanti, S. M. C. (2006). Estimating the density of a jaguar population in the Brazilian Pantanal using camera-traps and capture-recapture sampling in combination with GPS radio-telemetry. Biological Conservation, 129, 487-496. [ Links ]
Soares, T. N., Telles, M. P. C., Resende, L. V., Silveira, L., Jácomo, A. T. A., Morato, R. G., Diniz-Filho, J. A. F., Eizirik, E., Brondani, R. P. V., & Brondani, C. (2006). Paternity testing and behavioral ecology: A case study of jaguars (Panthera onca) in Emas National Park, Central Brazil. Genetics and Molecular Biology, 29, 735-740. [ Links ]
Suárez, E., Zapata-Ríos, G., Utreras, V., Strindberg, S., & Vargas, J. (2013). Controlling access to oil roads protects forest cover, but not wildlife communities: a case study from the rainforest of Yasuní Biosphere Reserve (Ecuador). Animal Conservation, 16, 265-274. [ Links ]
Terborgh, J. 1988. The big things that run the world – a sequel to E. O. Wilson. Conservation Biology, 2, 402-403. [ Links ]
Tobler, M. W., Carrillo-Percastegui, S. E., Zúñiga Hartley, A., & Powell, G. V. N. (2013). High jaguar densities and large population sizes in the core habitat of the southwestern Amazon. Biological Conservation, 159, 375-381. [ Links ]
Wallace, R. B., Gómez, H., Ayala, G., & Espinoza, F. (2003). Camera trapping for jaguar (Panthera onca) in the Tuichi valley, Bolivia. Mastozología Neotropical, 10, 133-139. [ Links ]
Weckel, M., Giuliano, W., & Silver, S. (2006). Jaguar (Panthera onca) feeding ecology: distribution of predator and prey through time and space. Journal of Zoology, 270, 25-30. [ Links ]
Blake, J. G., Mosquera, D., Guerra, J., Loiselle, B. A., Romo, D., & Swing, K. (2011). Mineral licks as diversity hotspots in lowland forest of eastern Ecuador. Diversity, 3, 217-234. [ Links ]
Blake, J. G., Mosquera, D., Loiselle, B. A., Swing, K., Guerra, J., & Romo, D. (2012). Temporal activity patterns of terrestrial mammals in lowland rainforest of eastern Ecuador. Ecotropica, 18, 137-146. [ Links ]
Blake, J. G., Mosquera, D., & Salvador, J. (2013). Use of mineral licks by mammals and birds in hunted and non-hunted areas of Yasuní National Park, Ecuador. Animal Conservation, 16, 420-437. [ Links ]
Crawshaw Jr., P. G. & Quigley, H. B. (1991). Jaguar spacing, activity and habitat use in a seasonally flooded environment in Brazil. Journal of Zoology, 223, 357-370. [ Links ]
Emmons, L. H. (1987). Comparative feeding ecology of felids in a Neotropical rainforest. Behavioral Ecology and Sociobiology, 20, 271-283. [ Links ]
Espinosa-Andrade, S. R. (2012). Road development, bushmeat extraction and jaguar conservation in Yasuni Biosphere Reserve -Ecuador. (Doctoral dissertation). University of Florida, Gainesville, Florida, USA. [ Links ]
Finer, M., Moncel, J., & Jenkins, C. N. (2010). Leaving the oil under the Amazon: Ecuador’s Yasuní-ITT initiative. Biotropica, 42, 63-66. [ Links ]
Foster, V. C., Sarmento, P., Sollmann, R., Tôrres, N., Jácomo, A. T. A., Negrões, N., Fonseca, C., & Silveira, L. (2013). Jaguar and puma activity patterns and predator-prey interactions in four Brazilian biomes. Biotropica, 45, 373-379. [ Links ]
Franzen, M. (2006). Evaluating the sustainability of hunting: a comparison of harvest profiles across three Huaorani communities. Environmental Conservation, 33, 1-10. [ Links ]
Kelly, M. J. (2003). Jaguar monitoring in the Chiquibul Forest, Belize. Caribbean Geography, 13, 19-32. [ Links ]
Larrea, C. & Warnars, L. (2009). Ecuador’s Yasuni-ITT initative: Avoiding emissions by keeping petroleum underground. Energy and Sustainable Development, 13, 219-223. [ Links ]
Maffei, L., Cuéllar, E., & Noss, A. (2004). One thousand jaguars (Panthera onca) in Bolivia’s Chaco? Camera trapping in the Kaa-Iya National Park. Journal of Zoology, 262, 295-304. [ Links ]
Maffei, L., Noss, A. J., Silver, S. C., & Kelly, M. J. (2011). Abundance/density case study: Jaguars in the Americas. In A. F. O’Connell, J. D. Nichols & K. U. Karanth (Eds.), Camera traps in animal ecology - methods and analyses, (pp. 119-144). New York, USA: Springer. [ Links ]
Mena, V. P., Stallings, J. R., Regalado, J. B., & Cueva, R. L. (2000). The sustainability of current hunting practices by the Huaorani. In J. G. Robinson & E. I. Bennett (Eds.), Hunting for sustainability in tropical forests (pp. 57-78). New York, USA: Columbia University Press. [ Links ]
Mendes-Pontes, A. R. & Chivers, D. J. (2007). Peccary movements as determinants of the movements of large cats in Brazilian Amazonia. Journal of Zoology, 273, 257-265. [ Links ]
Ministerio del Ambiente del Ecuador. (2010). Reservas de Biosfera del Ecuador: lugares excepcionales. Quito, Ecuador: GTZ/GESORENDED-WCS- NCI-UNESCO. [ Links ]
Rabinowitz, A. R. & Nottingham Jr., B. G. (1986). Ecology and behaviour of the jaguar (Panthera onca) in Belize, Central America. Journal of Zoology, 210, 149-159. [ Links ]
Romero-Muñoz, A., Maffei, L., Cuéllar, E., & Noss, A. J. (2010). Temporal separation between jaguar and puma in the dry forests of southern Bolivia. Journal of Tropical Ecology, 26, 303-311. [ Links ]
Scognamillo, D. I., Maxit, E., Sunquist, M., & Polisar, J. (2003). Coexistence of jaguar (Panthera onca) and puma (Puma concolor) in a mosaic landscape in the Venezuelan llanos. Journal of Zoology, 259, 269-279. [ Links ]
Silveira, L., Jácomo, A. T. A., & Diniz-Filho, J. A. F. (2003). Camera trap, line transect census and track surveys: a comparative evaluation. Biological Conservation, 114, 351-355. [ Links ]
Silver, S. C., Ostro, L. E. T., Marsh, L. K., Maffei, L., Noss, A. J., Kelly, M. J., Wallace, R. B., Gómez, H., & Ayala, G. (2004). The use of camera traps for estimating jaguar Panthera onca abundance and density using capture/recapture analysis. Oryx, 38, 148-154. [ Links ]
Soisalo, M. K. & Cavalcanti, S. M. C. (2006). Estimating the density of a jaguar population in the Brazilian Pantanal using camera-traps and capture-recapture sampling in combination with GPS radio-telemetry. Biological Conservation, 129, 487-496. [ Links ]
Soares, T. N., Telles, M. P. C., Resende, L. V., Silveira, L., Jácomo, A. T. A., Morato, R. G., Diniz-Filho, J. A. F., Eizirik, E., Brondani, R. P. V., & Brondani, C. (2006). Paternity testing and behavioral ecology: A case study of jaguars (Panthera onca) in Emas National Park, Central Brazil. Genetics and Molecular Biology, 29, 735-740. [ Links ]
Suárez, E., Zapata-Ríos, G., Utreras, V., Strindberg, S., & Vargas, J. (2013). Controlling access to oil roads protects forest cover, but not wildlife communities: a case study from the rainforest of Yasuní Biosphere Reserve (Ecuador). Animal Conservation, 16, 265-274. [ Links ]
Terborgh, J. 1988. The big things that run the world – a sequel to E. O. Wilson. Conservation Biology, 2, 402-403. [ Links ]
Tobler, M. W., Carrillo-Percastegui, S. E., Zúñiga Hartley, A., & Powell, G. V. N. (2013). High jaguar densities and large population sizes in the core habitat of the southwestern Amazon. Biological Conservation, 159, 375-381. [ Links ]
Wallace, R. B., Gómez, H., Ayala, G., & Espinoza, F. (2003). Camera trapping for jaguar (Panthera onca) in the Tuichi valley, Bolivia. Mastozología Neotropical, 10, 133-139. [ Links ]
Weckel, M., Giuliano, W., & Silver, S. (2006). Jaguar (Panthera onca) feeding ecology: distribution of predator and prey through time and space. Journal of Zoology, 270, 25-30. [ Links ]
*Correspondencia a:
John G. Blake. Department of Wildlife Ecology and Conservation, University of Florida, Gainesville, Florida, 32611, USA; john.blake@ufl.edu
Diego Mosquera. Estación de Biodiversidad Tiputini, Universidad San Francisco de Quito, P.O. Box 17-1200-841. Quito, Ecuador; dimosb@rocketmail.com
Jaime Guerra. Estación de Biodiversidad Tiputini, Universidad San Francisco de Quito, P.O. Box 17-1200-841. Quito, Ecuador; jaime_fguerra@yahoo.com
Bette A. Loiselle. Department of Wildlife Ecology and Conservation, University of Florida, Gainesville, Florida, 32611, USA.
Center for Latin American Studies, University of Florida, Gainesville, Florida, 32611, USA; loiselleb@ufl.edu
David Romo. Estación de Biodiversidad Tiputini, Universidad San Francisco de Quito, P.O. Box 17-1200-841. Quito, Ecuador; dromo@usfq.edu.ec
Kelly Swing. Estación de Biodiversidad Tiputini, Universidad San Francisco de Quito, P.O. Box 17-1200-841. Quito, Ecuador; kswing@usfq.edu.ec
1. Department of Wildlife Ecology and Conservation, University of Florida, Gainesville, Florida, 32611, USA; john.blake@ufl.edu
2. Estación de Biodiversidad Tiputini, Universidad San Francisco de Quito, P.O. Box 17-1200-841. Quito, Ecuador; dimosb@rocketmail.com
3. Estación de Biodiversidad Tiputini, Universidad San Francisco de Quito, P.O. Box 17-1200-841. Quito, Ecuador; jaime_fguerra@yahoo.com
4. Department of Wildlife Ecology and Conservation, University of Florida, Gainesville, Florida, 32611, USA.
5. Center for Latin American Studies, University of Florida, Gainesville, Florida, 32611, USA; loiselleb@ufl.edu
6. Estación de Biodiversidad Tiputini, Universidad San Francisco de Quito, P.O. Box 17-1200-841. Quito, Ecuador; dromo@usfq.edu.ec
7. Estación de Biodiversidad Tiputini, Universidad San Francisco de Quito, P.O. Box 17-1200-841. Quito, Ecuador; kswing@usfq.edu.ec
Received 29-VII-2013. Corrected 15-X-2013. Accepted 15-XI-2013.


{kind=link}
{kind=link}
{kind=link}
{kind=link}