










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.62 n.2 San José Apr./Jun. 2014
Mesoamerican tree squirrels evolution (Rodentia: Sciuridae): a molecular phylogenetic analysis
Evolución de las ardillas arborícas mesoamericanas (Rodentia: Sciuridae): un análisis filogenético molecular
Evolución de las ardillas arborícas mesoamericanas (Rodentia: Sciuridae): un análisis filogenético molecular
*Dirección para correspondencia:
Abstract
The tribe Sciurini comprehends the genera Sciurus, Syntheosiurus, Microsciurus, Tamiasciurus and Rheinthrosciurus. The phylogenetic relationships within Sciurus have been only partially done, and the relationship between Mesoamerican species remains unsolved. The phylogenetic relationships of the Mesoamerican tree squirrels were examined using molecular data. Sequence data publicly available (12S, 16S, CYTB mitochondrial genes and IRBP nuclear gene) and cytochrome B gene sequences of four previously not sampled Mesoamerican Sciurus species were analyzed under a Bayesian multispecies coalescence model. Phylogenetic analysis of the multilocus data set showed the neotropical tree squirrels as a monophyletic clade. The genus Sciurus was paraphyletic due to the inclusion of Microsciurus species (M. alfari and M. flaviventer). The South American species S. aestuans and S. stramineus showed a sister taxa relationship. Single locus analysis based on the most compact and complete data set (i.e. CYTB gene sequences), supported the monophyly of the South American species and recovered a Mesoamerican clade including S. aureogaster, S. granatensis and S. variegatoides. These results corroborated previous findings based on cladistic analysis of cranial and post-cranial characters. Our data support a close relationship between Mesoamerican Sciurus species and a sister relationship with South American species, and corroborates previous findings in relation to the polyphyly of Microsciurus and Syntheosciurus’ paraphyly. Rev. Biol. Trop. 62 (2): 649-657. Epub 2014 June 01.
Key words: phylogenetics, cytochrome B, Sciurus, Microsciurus, Syntheosciurus, Mesoamerica.
Resumen
La tribu Sciurini comprende los géneros Sciurus, Syntheosciurus, Microsciurus, Tamiasciurus y Rheinthrosciurus. Las relaciones filogenéticas de Sciurus han sido resueltas parcialmente mientras que las relaciones de las especies Mesoamericanas permanecen sin resolverse. Las relaciones filogenéticas de las ardillas arborícolas mesoamericanas fueron estudiadas empleando datos moleculares. Datos de secuencias disponibles de forma pública (genes mitocondriales CYTB, 12S, 16S y gen nuclear IRBP) en conjunto con secuencias nuevas para el gen del Citocromo B de 4 especies mesoamericanas del genero Sciurus, fueron analizadas empleando un modelo bayesiano de coalescencia multi-especie. Los análisis filogenéticos del conjunto de datos multilocus mostraron que las especies neotropicales forman un clado monofilético. El género Sciurus resulto ser parafilético debido a la inclusión de las especies de Microsciurus (M. alfari y M. flaviventer). Las especies suramericanas S. aestuans y S. stramineus presentaron una relación de especies hermanas. El análisis de un solo locus basado en el conjunto de datos más compacto y completo (secuencias del gen del citocromo B), apoyó la naturaleza monofilética de las especies suramericanas y recuperó un clado mesoamericano que incluye a S. aureogaster, S. granatensis y S. variegatoides. Estos resultados corroboran los descubrimientos previos que emplearon datos morfológicos craneales y pos-craneales. Nuestros datos apoyan la relación cercana entre las especies de Sciurus Mesoamericanas y la relación hermana de estas con las especies de Suramérica, así como también corroboran la relación polifilética de Microsciurus y parafilética de Syntheosciurus previamente reportadas.
Palabras clave: filogenética, citocromo B, Sciurus, Microsciurus, Syntheosciurus, Mesoamerica.
Tree squirrels are distributed throughout Eurasia and America, and grouped in the tribe Sciurini (Black, 1963; Wilson & Reeder, 2005). Sciurini comprehends the genera Sciurus, Syntheosiurus, Microsciurus, Tamiasciurus and Rheinthrosciurus. Among these, the genus Sciurus, with seven subgenera and 28 species (Wilson & Reader, 2005), is the most diverse
and widespread, with representatives in Eurasia (S. vulgaris), the Japan Islands (S.lis), the Middle East (S. anomalus) and the Americas. In the Americas, 25 Sciurus species occur, with several species restricted to the Neartic region (S. carolinensis, S. niger, S. aberti, S griseus and S. arizonensis). The genus reaches its highest diversity in the Neotropical region with 20 species.
Among the Neotropical Sciurus species, S. variegatoides, S. yucatanensis and S. richmondi are restricted to the Mesoamerican region -the area between the Isthmus of Tehuantepec and the Isthmus of Panama-. Several other species (e.g. S. deppei, S. granatensis, and S. colliaei) occur in the Mesoamerican region albeit not exclusively. In addition to Sciurus, there are other Sciurini genera with Mesoamerican endemic taxa; the genus Microsciurus currently is composed by four species: M. alfari and M. mimuls distributed in lower Central America, and M. flaviventer and M. santandarensis found in Northern South America. The genus Syntheosciurus presents only one species with two subspecies, S. brochus poaensis and S. brocus brocus, endemic of Talamanca-Chiriqui mountain range in Costa Rica and Panama.
The most complete taxonomic revision of the Sciurini was made by Moore (1959) based on the analysis of qualitative characters from the cranium. In this study, the genus Sciurus was placed in the subtribe Sciurina together with Reinthrosciurus Gray and Guerlinguetus Gray, which was later placed as a subgenus in Sciurus (Hall, 1981). An additional subtribe (Microsciurina) was also proposed for Microsciurus and Syntheosciurus. In relation to Microsciurus, the genus was described initially as a Sciurus´ subgenus (Allen, 1895) and later elevated to full generic rank (Goldman, 1912). A similar situation occurred with Syntheosciurus, formerly considered a subgenus of Sciurus (Goodwin, 1943, 1946). These affinities between Mesosciurus (currently S. granatensis) and the species in Microsciurus were also suggested by Moore (1959).
The phylogenetic relationships within Sciurus have been only partially resolved. Several studies have corroborated the monophyly of S. vulgaris and S. lis (Oshida, Masuda, & Yoshida, 1996; Oshida & Masuda, 2000; Oshida, Arslan, & Noda, 2009). Also, previous analyses based on an incomplete taxon sampling have suggested that South American species might form a monophyletic clade (Hafner, Barkley & Chupasko, 1994; Steppan, Storz, & Hoffmann, 2004; Herron, Castoe, & Parkinson, 2004; Oshida et al., 2009). Moreover, the most species comprehensive phylogenetic study on neotropical tree squirrels to date (Villalobos & Cervantes, 2007) demonstrated the monophyly of the Mesoamerican squirrel species. In that study, Mesoamerican species were split in two clades including S. aureogaster, S. colliaei, S. variegatoides and Syntheosciurus brochus, and S. richomondi, S. granatensis, S. deppei, S. yucatanensis and Microsciurus alfari, respectively. These findings questioned the validity Synteosciurus, suggesting that S. brochus should be considered a Sciurus species, and also pointed to the possible polyphyly of Microsciurus.
The phylogenetic relationships within the Sciurini tribe have been the focus of several studies in recent years. An early study by Hafner et al. (1994) based on protein variation showed Microsciurus flaviventer in a Sciurus clade comprising S. carolinensis, S. niger and S. stramineus. The pioneering work of Mercer & Roth (2003), based on the mitochondrial (12S rDNA and 16S rDNA) and nuclear genes (IRBP), brought some information on the intratribe relationships and showed the paraphyletic nature of the Sciurus genera in relation to Syntheosciurus and Microsciurus, and the relationship of these genera with Tamiasciurus. These same results were recovered by Steppan et al. (2004) using two nuclear genes (RAG1 and cmy) and by Herron et al. (2004) using a mitochondrial gene (cytb). The close relationship between M. flaviventer and the Southamerican Sciurus species is a common feature throughout these studies. Despite these relevant results, the relationships within Sciurus and its related genera remain largely unresolved. The current data information include several genes (nuclear and mitochondrial for a total of 8) but remains poor in the number of taxa included and represented, precluding the possibilities of a more comprehensive estimation of the phylogenetic relationships. A recent combined supermatrix approach using all gene sequence publicly available (Pečnerová & Martínková, 2012) also revealed the same relationships between taxa. However, the phylogenetic relationships between Mesoamerican species remained unresolved, only supporting with low probability the sister relationship between M. alfari and S. granatensis.
The methodological approach of concatenated supermatrix multilocus analysis has been recently point it as a possible source of error in phylogenetic estimation. First revealed by Maddison (1997), the phylogenetic trees depicted by particular gene histories not necessarily correspond with the species tree phylogeny due to processes like horizontal transfer, lineage sorting, gene duplication and gene extinction. In conditions of short branches, closely related species and large population numbers the discrepancy between gene trees and species is expected to be more problematic (Madisson, 1997; Maddison & Knowles, 2006). Therefore, the use of several independent loci to estimate the species tree was suggested (Pamilo & Nei, 1998) and adopted as thought concatenation of several genes, as a common practice in phylogenetic reconstruction. However, recent studies into multigene phylogenetics have shown that concatenating genes can lead to poor estimation of species tree (Degnan & Rosenberg, 2006; Kubatko, 2007). When multiple loci are concatenate they are assumed to evolve according a single evolutionary tree, obviating the effect of recombination in decoupling the evolutionary histories of different loci (Dengan & Rosenberg, 2006). Under simulated conditions the supermatrix approach has shown a high probability of error estimating species trees, only 9 of 100 replicates contained the true species tree topology (Heled & Drummond, 2010).
In this study, we aim at resolving the phylogenetic relationships of the Mesoamerican tree squirrels using a Bayesian multispecies coalescence model. We provide new sequence data (CYTB) of four Mesoamerican Sciurus species and used the newly generated information together with published DNA sequences.
Materials and methods
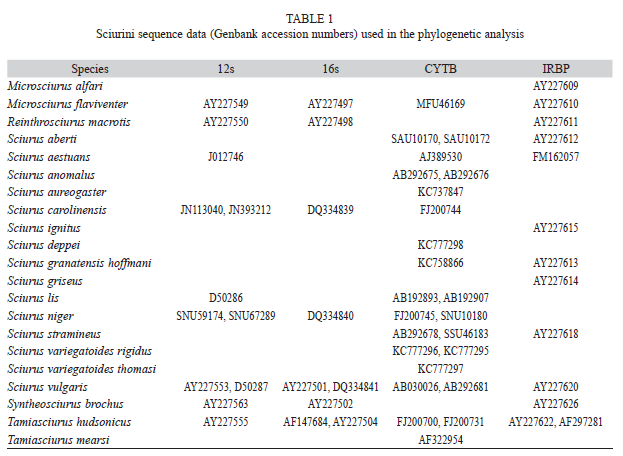
Specimens analyzed: Twenty one taxa, representing all recognized genera New World Tree squirrels, were included in the phylogenetic analysis, (Table 1). The outgroup taxa included Tamiasciurus hudsonicus, T. mearsi and Reinthrosciurus macrotis. More than one DNA sequences were included per species, depending on the availability in the GenBank database.
DNA extraction, amplification and sequencing: Genomic DNA was extracted from tissue (0.25g) or museum skin samples (50mm2) for six taxa (S. aureogaster, S. deppei, S. granatensis hoffmani, S.variegatoides atrirufus, S.variegatoides rigidus and S. variegatoides thomasi) using Wizzard Genomic Extraction Kit (Promega). A region of 1144bp from the mitochondrial cytochrome B gene was amplified using the primers H15910 and L14724 (Oshida, Liang-Kong, Masuda, & Yoshida, 2000). PCRs were set-up in a 25μL reaction volume containing the following reaction conditions: 1μL of template DNA, 2.5mm MgCl2, 200μM dNTPs, 0.5pmol/μL of each primer and 1.25U of Taq DNA polymerase (Fermentas). The amplification profile consisted of an initial denaturalization step of 94ºC for 4 min; followed by 40 cycles of amplification (94ºC for 1min., 55ºC for 1min, 72ºC for 1min.) and a final extension at 72ºC for 10min. PCR products were purified using Exo-sSap and sequenced in both directions on an ABI3130 automated Genetic Analyzer using BigDye 3.1 DNA sequencing chemistry (Applied Biosystems) and following manufacturer´s instructions. Sequences from this study are available under GenBank Accession Nos. KC737847, KC758866 KC777295, KC777296, KC777297, KC777298.
Phylogenetic analysis: Contig assemblies for cytochrome B sequences of both strands were made with GENEIOUS 5.6.4 (Biomatters Ltd., Auckland, New Zealand). All sequences obtained for this study plus Genbank sequences were aligned with Muscle using the default parameters (Edgar, 2004). The molecular substitution model for each gene was selected using JModelTest (Guindon & Gascuel, 2003; Posada, 2008) and the Akaike information criteria (Burnham & Anderson, 2003); mitochondrial 12s and 16s rDNA were analyzed under the GTR+G model, CYTB under the GTR G+I model; and the nuclear IRBP gene under the GTR+I model.
Multilocus phylogenetic reconstruction was performed using starBEAST (Heled & Drummond, 2010) for Bayesian inference, using the log-normal relaxed clock model with a Yule-prior for branch lengths. Two or more DNA sequences per taxa were considered when the information availability allows it, as recommend by Heled & Drummond (2010). Two independent runs for 100 million generations and sampled every 1 000 generations. The first 10 million generations were discarded as burnin. TRACER v1.5 (Drummond & Rambaut, 2007) was used to estimate the effective sample size for each parameter (all resulting effective sample sizes exceeded 100) and calculate the mean and upper and lower bounds of the 95% highest posterior density interval (95% HPD). The trees sampled were combined using LogCombiner (Drummond & Rambaut, 2007) and resampled every 2 000 generations. Single locus phylogenetic reconstruction using the most complete dataset (i.e. cytochrome B) was performed using BEAST v1.7.4 (Drummond & Rambaut, 2007) and the same conditions as the multilocus approach, however only one run of 100 million generations, sampled every 1 000 generations, was used.
Results
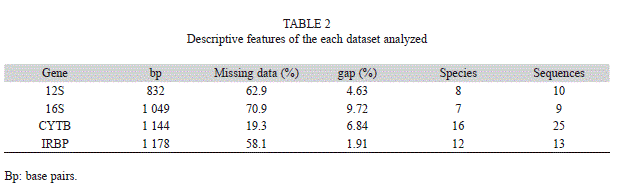
The four genes analyzed were variable in relation to the proportion of missing data, gap composition and number of species sampled (Table 2). The most complete data set was the cytochorme B gene followed by the nuclear gene IRBP. The proportion of sequences to taxa in the analysis was greater than 1.25 (12S rDNA) and up to 1.56 (CYTB). In total, the four loci included 4203bp. The implemented MCMC runs converged to a common stationary phase and the ESS for all estimated parameters was greater than 100.
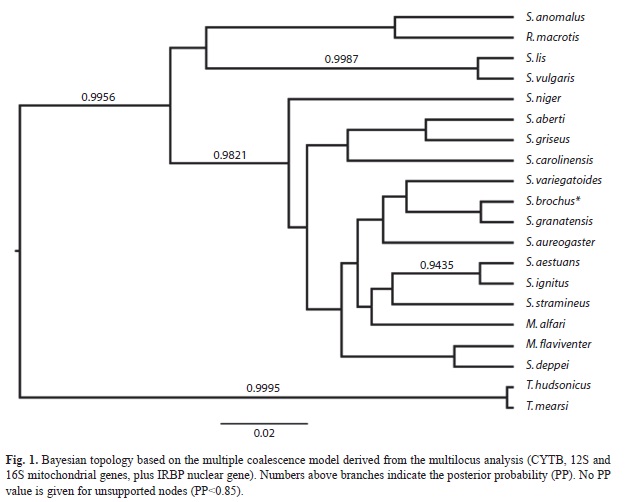
The sister taxa relationship between Tamiasciurus and Sciurus was recovered with high support in our multilocus Bayesian phylogeny (Fig. 1). The genus Sciurus was paraphyletic including Microsciurus species (M. alfari and M. flaviventer). Microsciurus was not monophyletic. Also, none of the phylogenetic relationships previously reported within Sciurus were recovered with high posterior probability. The Eurasian species S. vulgaris and S. lis were monophyletic. Also, an unsupported relationship between R. macrotis, S. anomalus (Middle East) and S. vulgaris-S. lis was recovered. The monophyletic nature of neotropical tree squirrels was corroborated but the internal relationships of this clade were unsupported. The only supported clade within this group includes S. aestuans and S. ignitus, both South American species, a relationship also recovered in previous studies.
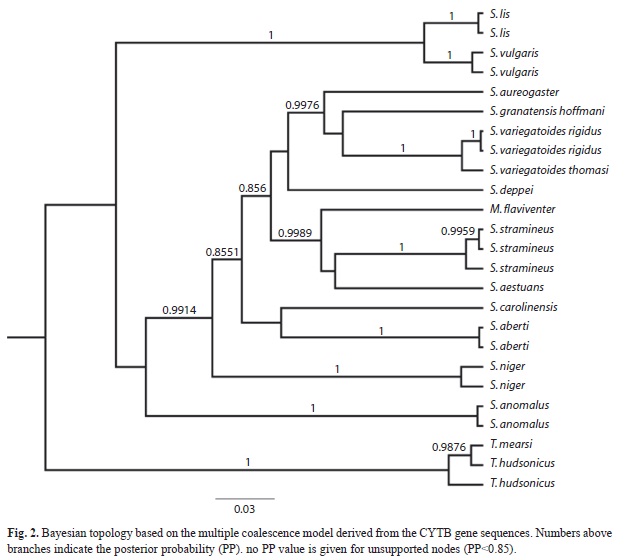
The single locus analysis (Fig. 2) based on CYTB sequences, recovered similar phylogenetic relationships for the Eurasian Sciurus species. However, we found with strong support (PP=0.9989) the monophyly of all South American species included in the analysis (M. flaviventer, S. stramineuos and S. aestuans). South American species were nested within a clade composed by all the Mesoamerican species. In this clade, a monophyletic group containing S. aureogaster, S. granatensis and S. variegatoides was recovered (PP: 0.9976). S. deppei is placed in a polytomy with South American and Mesoamerica clades. In relation to neartic species, S. aberti and S. carolinensis were placed them in a politomy with the Mesoamerican species, while S. niger was sister taxa to them.
Discussion
Our multilocus analysis using the most complete molecular data set recovered the general relationships previously reported in the literature (Oshida & Masuda, 2000; Mercer & Roth, 2003; Pečnerová & Martínková, 2012), but failed to unveil novel phylogenetic relationships among Sciurus species with high posterior probabilities. This could be due to the missing data in the genes analyzed. Consequently, our current best estimation on the phylogenetic relationships of Sciurus species rest on the cytochrome B gene tree.
The single locus analysis shows a close relationship between S. aureogaster, S. granatensis and S. variegatoides, however, the relationships between these species remains unsolved. A close relationship between S. aureograster, S. variegatoides and S colliaei has been previously suggested based on phenotypic and distributional data (Musser, 1969), and a cladistic analysis of cranial and postcranial characters (Villalobos & Cervantes, 2007) within Sciurini/Sciurus found these species belonging to the same clade. However, this same analysis suggested that S. granatensis was part of a sister clade, which included S. deppei. The relationship between S. deppei and the S. aureogaster + S. granatensis + S. variegatoides clade cannot be rejected. The cytochorome b gene tree indicates that S. deppei is part of a polytomy, future sampling of more molecular characters is need to resolve it, and test the phylogenetic relationships obtained using morphological characters.
The South American clade recovered in previous studies (Mercer & Roth, 2003; Pečnerová & Martínková, 2012) was also found in the single locus analysis. The sister taxa relationship of M. flaviventer, S. stramineus and S. aestuans was reported by Pečnerová & Martínková (2012) based on the cytochrome B, cmy exons and IRBP gene trees, all with an adequate posterior probability. Our current analysis also suggests this relationship.
Based on the molecular clock analysis (Mercer & Roth, 2003) the divergence observed between the Mesoamerican species clade and the South American clade was estimated to take place 3.4 to 3.1 MYA. However, this speciation event may not be associated to the Panamian Isthmus closure as previously suggested (Mercer & Roth, 2003), but to the formation of savanna-like environments caused by Northern ice sheets formation (Molnar, 2008; Montes et al., 2012). Novel geologic mapping evidence in Central Panama suggest that the Panamian Isthmus was a continuous uninterrupted chain above sea level from the late Eocene to early Miocene, with a closure at 15 MYA (Montes et al., 2012). The South America colonization and subsequent diversification may be expected to be a recent event. The central and Northern Andes deformation reach its current extension by late Oligocene to early Miocene (Oncken et al., 2006; Mora et al., 2010), providing a physiographic landscape similar to its present-day conformation. Therefore, the vicariant events associated to squirrel diversification in South America must be related to glacial cycles as has been suggested for the Mesoamerican Sciurus species (Villalobos, 2013).
The phenotypic variation observed among subspecies in some Sciurus may be associated to isolation events during glacial cycles (Moncrief et al., 2012). S. variegatoides comprises 15 subespecies with a high variation in color and coat pattern, distributed throughout Mesoamerica from the Tehuantepec Isthmus to the Panamian Isthmus (Harris, 1937; Hall, 1981). Seven subspecies are present and restricted to Costa Rica high and lowlands. Among these, S. v. rigidus and S. v. thomasi are related and allopatric: S. v. rigidus is distributed in Costa Rica`s central valley highlands and into the Pacific coast, while S. v. thomasi is found mainly in the Caribbean coast and into the Talamanca Cordillera (Hall, 1981). The CYTB gene tree shows both subspecies as sister taxa, suggesting a potential phylogeographic pattern. These results contrast with the findings reported in S. niger subspecies (Moncrief, Lack & Van den Bussche, 2010; Moncrief et al., 2012). S. niger is also a highly diverse species containing 10 subspecies with a marked diversity in coat and color pattern. However, cytochrome B gene analysis has shown a lack of phylogeographic structure attributed to a rapid post glaciation range expansion and rapid phenotypic divergence (Moncrief et al., 2012). In order to evaluate the mechanism responsible for S. variegatoides subspecies diversity and to corroborate these preliminary results, a more extensive sampling is required.
Our data support a close relationship between Mesoamerican Sciurus species and a sister relationship with South American species, both groups conform a larger clade distinct from the Neartic species. This study corroborates previous findings in relation to the polyphyly of Microsciurus and Syntheosciurus’ paraphyly. Future studies remain necessary to clarify the phylogenetic position of the Mexican species of Sciurus and their relation to Mesoamerican species. Also, the diversification of South American species remains unresolved but centered in the Andes Cordillera as the region of endemism and speciation. At a lesser evolutionary scale, our data suggest the existence of phylogeographic structure within S. variegatoides subspecies, which deserves to be further analyzed.
Acknowledgments
This work is dedicated to Marién. Special thanks to Bernal Morera, Sergio Vargas, and Ginny Abrahams provided fruitful discussion, comments and suggested improvements to the manuscript. This work was supported by CONICIT grant FI-179-05.
References
Allen, J. A. (1895). Description of new American mammals. Bulletin of the American Museum of Natural History, 7, 333. [ Links ]
Black, C. C. (1963). A review of the North American tertiary Sciuridae. Bulletin of the Museum of Comparative Zoology, 130, 109-248. [ Links ]
Burnham, K. P. & Anderson, D. R. (2003). Model selection and multimodel inference: a practical information theoretic approach. New York: Springer-Verlag. [ Links ]
Degnan, J. H. & Rosenberg, N. A. (2006). Discordance of species trees with their most likely gene trees. PLoS Genetics, 2, e68. [ Links ]
Drummond, A. J. & Rambaut, A. (2007). BEAST: Bayesian evolutionary analysis by sampling trees. BMC Evolutionary Biology, 7, 214. [ Links ]
Edgar, R. C. (2004). MUSCLE: a multiple sequence alignment method with reduced time and space complexity. BMC Bioinformatics, 5, 113. [ Links ]
Goldman, E. A. (1912). Descriptions of twelve new species and subspecies of mammals from Panama. Smithsonian Miscellaneous Collections, 56, 1-11. [ Links ]
Goodwin, G. G. (1943). Two new squirrels from Costa Rica. American Museum Noviates, 1218, 1-2. [ Links ]
Goodwin, G. G. (1946). Mammals of Costa Rica. Bulletin of the American Museum of Natural History, 87, 275-473. [ Links ]
Guindon, S. & Gascuel, O. (2003). A simple, fast and accurate method to estimate large phylogenies by maximum-likelihood. Systematic Biology, 52, 696-704. [ Links ]
Hall, E. R. (1981). The mammals of North America. New York: John Wiley and Sons. [ Links ]
Hafner, M. S., Barkley, L. J., & Chupasko, J. M. (1994). Evolutionary genetics of New World tree squirrels (tribe Sciurini). Journal of Mammalogy, 75, 102-109. [ Links ]
Herron, M. D., Castoe, T. A., & Parkinson, C. L. (2004). Sciurid phylogeny and the paraphyly of Holarctic ground squirrels (Spermophilus). Molecular Phylogenetics and Evolution, 31, 1015-1030. [ Links ]
Harris, W. P. (1937). Revision of Sciurus variegatoides, a species of Central American squirrel. Miscellaneous Publications Museum of Zoology, University of Michigan, 38, 1-38. [ Links ]
Heled, J. & Drummond, A. J. (2010). Bayesian Inference of Species Trees from Multilocus Data. Molecular Biology and Evolution, 27, 570-580. [ Links ]
Kubatko, L. S. (2007). Inconsistency of phylogenetic estimates from concatenated data under coalescence. Systematic Biology, 56, 17-24. [ Links ]
Maddison, W. P. (1997). Gene Trees in Species Trees. Systematic Biology, 46, 523-536. [ Links ]
Maddison, W. P. & Knowles, L. L. (2006). Inferring Phylogeny despite Incomplete Lineage Sorting. Systematic Biology, 55, 21-30. [ Links ]
Mercer, J. M. & Roth, L. (2003). The Effects of Cenozoic Global Change on Squirrel Phylogeny. Science, 299, 1568-1572. [ Links ]
Molnar, P. (2008). Closing of the Central American Seaway and the ice age: a critical review. Paleoceanography, 23, PA2201. [ Links ]
Moncrief, M. D., Lack, J. B., Maldonado, J. E., Bryant, K. L, Edwards, C. W., & Van Den Bussche, R. A. (2012). General lack of phylogeographic structure in two sympatric, forest obligate squirrels (Sciurus niger and S. carolinensis). Journal of Mammalogy, 93, 1247-1264. [ Links ]
Moncrief, N. D., Lack, J. B., & Van den Bussche, R. A. (2010). Eastern fox squirrel (Sciurus niger) lacks phylogeographic structure: recent range expansion and phenotypic differentiation. Journal of Mammalogy, 91, 1112-1123. [ Links ]
Montes, C., Cardona, A., McFadden, R., Morón, S. E., Silva, C. A., Restrepo-Moreno, S., Ramiréz, D. A., Hoyos, N., Farris, D., Bayona, G. A., Jaramillo, C. A., Valencia, V., Bryan, J., & Flores, J. A. (2012). Evidence for middle Eocene and younger land emergence in Central Panama: implications for Isthmus closure. Geological Society of America Bulletin, 124, 780-799. [ Links ]
Moore, J. C. (1959). Relationships among the living species of the Sciurinae. Bulletin of American Museum of Natural History, 118, 153-206. [ Links ]
Mora, A., Horton, B., Mesa, A., Rubiano, J., Ketchum, R., Parra, M., Blanco, M., García, D., & Stockli, D. (2010). Migration of Cenozoic deformation in the Eastern Cordillera of Colombia interpreted from fission track results and structural relationships: implications for petroleum systems. American Association of Petroleum Geologists Bulletin, 94, 1543-1580. [ Links ]
Musser, G. G. (1969). A systematic study of the Mexican and Guatemalan gray squirrel, Sciurus aureogaster F. Cuvier (Rodentia: Sciuridae). Miscellaneous Publications, Museum of Zoology, University of Michigan, 137, 1-112. [ Links ]
Oncken, O., Hindle, D., Kley, J., Elger, K., Victor, P., & Schemmann, K. (2006). Deformation of the Central Andean Upper Plate system–facts, fiction, and constraints for Plateau models. In O. Oncken, G. Chong, G. Franz, P. Giese, P., Götze, H-J., V. A. Ramos, M. R. Strecker, & P. Wigger (Eds.), The Andes–Active Subduction Orogeny (pp. 3-27). Berlin: Springer Publishing. [ Links ]
Oshida, T., Arslan, A., & Noda, M. (2009). Phylogenetic relationships among the old world Sciurus squirrels. Folia Zoologica, 58, 14-25. [ Links ]
Oshida, T., Liang-Kong, L., Masuda, R., & Yoshida, M. C. (2000). Phylogenetic Relationships among Asian species of Petaurista (Rodentia, Sciuridae), Inferred from Mitochondrial Cytochrome B Gene Sequences. Zoological Science, 17, 123-128. [ Links ]
Oshida, T., & Masuda, R. (2000). Phylogeny and zoogeography of six squirrel species of the genus Sciurus (Mammalia, Rodentia), inferred from cytochrome B gene sequences. Zoological Science, 17, 405-409. [ Links ]
Oshida, T., Masuda, R., & Yoshida, M. C. (1996). Phylogenetic relationships among Japanese species of the family Sciuridae (Mammalia, Rodentia), inferred from nucleotide sequences of mitochondrial 12S ribosomal RNA genes. Zoological Science, 13, 615-620. [ Links ]
Pamilo, P., & Nei, M. (1988). Relationships between gene trees and species trees. Molecular Biology and Evolution, 5, 568-583. [ Links ]
Pečnerová, P., & Martínková, N. (2012). Evolutionary history of tree squirrels (Rodentia, Sciurini) based on multilocus phylogeny reconstruction. Zoologica Scripta, 41, 211-219. [ Links ]
Posada, D. (2008). jModelTest: Phylogenetic Model Averaging. Molecular Biology and Evolution, 25, 1253-1256. [ Links ]
Steppan, S. J., Storz, B. L., & Hoffmann, R. S. (2004). Nuclear DNA phylogeny of the squirrels (Mammalia: Rodentia) and the Evolution of arboreality from c-myc and rag1. Molecular Phylogenetics and Evolution, 30, 703-719. [ Links ]
Villalobos, F. (2013). Tree squirrels: A key to understand the historic biogeography of Mesoamerica? Mammalian Biology, 78, 258-266. [ Links ]
Villalobos, F. & Cervantes-Reza, F. (2007). Phylogenetic relationships of Mesoamerican species of the genus Sciurus (Rodentia: Sciuridae). Zootaxa, 1525, 31-40. [ Links ]
Wilson, D. E. & Reeder, D. M. (2005). Mammal species of the world. A Taxonomic and Geographic Reference (3rd ed.). Baltimore: John Hopkins University Press. [ Links ]
Black, C. C. (1963). A review of the North American tertiary Sciuridae. Bulletin of the Museum of Comparative Zoology, 130, 109-248. [ Links ]
Burnham, K. P. & Anderson, D. R. (2003). Model selection and multimodel inference: a practical information theoretic approach. New York: Springer-Verlag. [ Links ]
Degnan, J. H. & Rosenberg, N. A. (2006). Discordance of species trees with their most likely gene trees. PLoS Genetics, 2, e68. [ Links ]
Drummond, A. J. & Rambaut, A. (2007). BEAST: Bayesian evolutionary analysis by sampling trees. BMC Evolutionary Biology, 7, 214. [ Links ]
Edgar, R. C. (2004). MUSCLE: a multiple sequence alignment method with reduced time and space complexity. BMC Bioinformatics, 5, 113. [ Links ]
Goldman, E. A. (1912). Descriptions of twelve new species and subspecies of mammals from Panama. Smithsonian Miscellaneous Collections, 56, 1-11. [ Links ]
Goodwin, G. G. (1943). Two new squirrels from Costa Rica. American Museum Noviates, 1218, 1-2. [ Links ]
Goodwin, G. G. (1946). Mammals of Costa Rica. Bulletin of the American Museum of Natural History, 87, 275-473. [ Links ]
Guindon, S. & Gascuel, O. (2003). A simple, fast and accurate method to estimate large phylogenies by maximum-likelihood. Systematic Biology, 52, 696-704. [ Links ]
Hall, E. R. (1981). The mammals of North America. New York: John Wiley and Sons. [ Links ]
Hafner, M. S., Barkley, L. J., & Chupasko, J. M. (1994). Evolutionary genetics of New World tree squirrels (tribe Sciurini). Journal of Mammalogy, 75, 102-109. [ Links ]
Herron, M. D., Castoe, T. A., & Parkinson, C. L. (2004). Sciurid phylogeny and the paraphyly of Holarctic ground squirrels (Spermophilus). Molecular Phylogenetics and Evolution, 31, 1015-1030. [ Links ]
Harris, W. P. (1937). Revision of Sciurus variegatoides, a species of Central American squirrel. Miscellaneous Publications Museum of Zoology, University of Michigan, 38, 1-38. [ Links ]
Heled, J. & Drummond, A. J. (2010). Bayesian Inference of Species Trees from Multilocus Data. Molecular Biology and Evolution, 27, 570-580. [ Links ]
Kubatko, L. S. (2007). Inconsistency of phylogenetic estimates from concatenated data under coalescence. Systematic Biology, 56, 17-24. [ Links ]
Maddison, W. P. (1997). Gene Trees in Species Trees. Systematic Biology, 46, 523-536. [ Links ]
Maddison, W. P. & Knowles, L. L. (2006). Inferring Phylogeny despite Incomplete Lineage Sorting. Systematic Biology, 55, 21-30. [ Links ]
Mercer, J. M. & Roth, L. (2003). The Effects of Cenozoic Global Change on Squirrel Phylogeny. Science, 299, 1568-1572. [ Links ]
Molnar, P. (2008). Closing of the Central American Seaway and the ice age: a critical review. Paleoceanography, 23, PA2201. [ Links ]
Moncrief, M. D., Lack, J. B., Maldonado, J. E., Bryant, K. L, Edwards, C. W., & Van Den Bussche, R. A. (2012). General lack of phylogeographic structure in two sympatric, forest obligate squirrels (Sciurus niger and S. carolinensis). Journal of Mammalogy, 93, 1247-1264. [ Links ]
Moncrief, N. D., Lack, J. B., & Van den Bussche, R. A. (2010). Eastern fox squirrel (Sciurus niger) lacks phylogeographic structure: recent range expansion and phenotypic differentiation. Journal of Mammalogy, 91, 1112-1123. [ Links ]
Montes, C., Cardona, A., McFadden, R., Morón, S. E., Silva, C. A., Restrepo-Moreno, S., Ramiréz, D. A., Hoyos, N., Farris, D., Bayona, G. A., Jaramillo, C. A., Valencia, V., Bryan, J., & Flores, J. A. (2012). Evidence for middle Eocene and younger land emergence in Central Panama: implications for Isthmus closure. Geological Society of America Bulletin, 124, 780-799. [ Links ]
Moore, J. C. (1959). Relationships among the living species of the Sciurinae. Bulletin of American Museum of Natural History, 118, 153-206. [ Links ]
Mora, A., Horton, B., Mesa, A., Rubiano, J., Ketchum, R., Parra, M., Blanco, M., García, D., & Stockli, D. (2010). Migration of Cenozoic deformation in the Eastern Cordillera of Colombia interpreted from fission track results and structural relationships: implications for petroleum systems. American Association of Petroleum Geologists Bulletin, 94, 1543-1580. [ Links ]
Musser, G. G. (1969). A systematic study of the Mexican and Guatemalan gray squirrel, Sciurus aureogaster F. Cuvier (Rodentia: Sciuridae). Miscellaneous Publications, Museum of Zoology, University of Michigan, 137, 1-112. [ Links ]
Oncken, O., Hindle, D., Kley, J., Elger, K., Victor, P., & Schemmann, K. (2006). Deformation of the Central Andean Upper Plate system–facts, fiction, and constraints for Plateau models. In O. Oncken, G. Chong, G. Franz, P. Giese, P., Götze, H-J., V. A. Ramos, M. R. Strecker, & P. Wigger (Eds.), The Andes–Active Subduction Orogeny (pp. 3-27). Berlin: Springer Publishing. [ Links ]
Oshida, T., Arslan, A., & Noda, M. (2009). Phylogenetic relationships among the old world Sciurus squirrels. Folia Zoologica, 58, 14-25. [ Links ]
Oshida, T., Liang-Kong, L., Masuda, R., & Yoshida, M. C. (2000). Phylogenetic Relationships among Asian species of Petaurista (Rodentia, Sciuridae), Inferred from Mitochondrial Cytochrome B Gene Sequences. Zoological Science, 17, 123-128. [ Links ]
Oshida, T., & Masuda, R. (2000). Phylogeny and zoogeography of six squirrel species of the genus Sciurus (Mammalia, Rodentia), inferred from cytochrome B gene sequences. Zoological Science, 17, 405-409. [ Links ]
Oshida, T., Masuda, R., & Yoshida, M. C. (1996). Phylogenetic relationships among Japanese species of the family Sciuridae (Mammalia, Rodentia), inferred from nucleotide sequences of mitochondrial 12S ribosomal RNA genes. Zoological Science, 13, 615-620. [ Links ]
Pamilo, P., & Nei, M. (1988). Relationships between gene trees and species trees. Molecular Biology and Evolution, 5, 568-583. [ Links ]
Pečnerová, P., & Martínková, N. (2012). Evolutionary history of tree squirrels (Rodentia, Sciurini) based on multilocus phylogeny reconstruction. Zoologica Scripta, 41, 211-219. [ Links ]
Posada, D. (2008). jModelTest: Phylogenetic Model Averaging. Molecular Biology and Evolution, 25, 1253-1256. [ Links ]
Steppan, S. J., Storz, B. L., & Hoffmann, R. S. (2004). Nuclear DNA phylogeny of the squirrels (Mammalia: Rodentia) and the Evolution of arboreality from c-myc and rag1. Molecular Phylogenetics and Evolution, 30, 703-719. [ Links ]
Villalobos, F. (2013). Tree squirrels: A key to understand the historic biogeography of Mesoamerica? Mammalian Biology, 78, 258-266. [ Links ]
Villalobos, F. & Cervantes-Reza, F. (2007). Phylogenetic relationships of Mesoamerican species of the genus Sciurus (Rodentia: Sciuridae). Zootaxa, 1525, 31-40. [ Links ]
Wilson, D. E. & Reeder, D. M. (2005). Mammal species of the world. A Taxonomic and Geographic Reference (3rd ed.). Baltimore: John Hopkins University Press. [ Links ]
*Correspondencia a:
Federico Villalobos. Laboratorio de Genética Evolutiva, Escuela de Ciencias Biológicas, Universidad Nacional de Costa Rica, Heredia, Costa Rica. fvillalo@gmail.com
Programa de Doctorado en Ciencias, Facultad de Ciencias, Universidad de Costa Rica, San José, Costa Rica; fvillalo@gmail.com
Gustavo Gutierrez-Espeleta. Escuela de Biología, Universidad de Costa Rica, San José, Costa Rica; gustavo.gutierrez@ucr.ac.cr
1. Laboratorio de Genética Evolutiva, Escuela de Ciencias Biológicas, Universidad Nacional de Costa Rica, Heredia, Costa Rica. fvillalo@gmail.com
2. Programa de Doctorado en Ciencias, Facultad de Ciencias, Universidad de Costa Rica, San José, Costa Rica; fvillalo@gmail.com
3. Escuela de Biología, Universidad de Costa Rica, San José, Costa Rica; gustavo.gutierrez@ucr.ac.cr
Received 22-vii-2013. Corrected 13-i-2014. Accepted 23-i-2014.


{kind=link}
{kind=link}
{kind=link}
{kind=link}