










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.62 n.2 San José Apr./Jun. 2014
Larval development of Evermannia zosterura (Perciformes: Gobiidae)
*Dirección para correspondencia:
Abstract
Gobiidae is the most specious fish family in the world with almost 2 000 species, however only 11% of them have been described for their larval stages. The entire life cycle information is essential to understand the biology and ecology of this important fish group. Previous studies on zooplankton samples from Ensenada de La Paz, México, have shown the presence of several Gobiidae larvae and juveniles which were identified as Evermania zosterura. The main objective of this work was to describe the larval stages of this species, widely distributed in the Eastern tropical Pacific. The development of E. zosterura larvae was described based on 66 specimens. A total of 53 specimens were used to describe morphometrics and pigmentation patterns, while 13 specimens were cleared and stained, to obtain meristic characteristics. Cleared specimens had 30 to 31 total vertebrae; dorsal-fin elements: IV; I, 13-14, anal-fin elements: I, 13-14, and most had pterygiophore formula 4-111100. The combination of these characteristics confirmed these specimens as E. zosterura. The pigment pattern is similar throughout ontogeny. Larvae are characterized by having three to five dendritic melanophores along the post-anal ventral margin, four to nine smaller melanophores along the ventral margin between the isthmus and anus, and one on the midpoint of the dorsal margin of the tail. There is one small pigment spot on the angle of the jaw, and other on the tip of lower lip. There is an elongated internal pigment under the notochord, between the head and gas bladder. Notochord flexion starts near 3.5mm BL and ends at 4.6mm BL; transformation to the juvenile stage is at about 13.6mm BL. Our conclusion is that the most useful characters to distinguish this species early-larval stages from those of similar species in the area, are the number of myomeres, the large melanophores (approximately uniformly in size) on the post anal ventral margin, and the elongate internal pigment under the notochord, anterior to the gas bladder. Rev. Biol. Trop. 62 (2): 603-610. Epub 2014 June 01.
Key words: Gobiidae, early development, pigmentation pattern, ontogeny, larvae, Gulf of California.
Resumen
La familia Gobiidae es la más diversa de los peces en el mundo con casi 2 000 especies, sin embargo solo el 11% de ellos han sido descritos en sus estadios larvarios. El conocimiento del ciclo de vida completo es esencial para entender la biología y ecología de este importante grupo de peces. Muestras de zooplancton obtenidas de la Ensenada de La Paz, México, mostraron la presencia de varias larvas y juveniles de Gobiidae, las cuales fueron identificadas como Evermania zosterura. El principal objetivo de este trabajo fue describir los estadios larvarios de esta especie ampliamente distribuida en el Pacífico tropical Oriental. Se describió el desarrollo larvario de E. zosterura con base en 66 especímenes recolectados en la Ensenada de La Paz, México. Sólo 53 especímenes se usaron para describir la morfometría y el patrón de pigmentación, mientras que 13 ejemplares transparentados y teñidos se utilizaron para obtener las características merísticas. Los especímenes transparentados tuvieron de 30 a 31 vertebras totales; los elementos de las aletas dorsales fueron IV; I, 13-14, los de la aleta anal I, 13-14 y la mayoría tuvo una formula pterigiofórica de 4-111100. La combinación de estas características, confirmó que pertenecen a E. zosterura. El patrón de pigmentación es muy similar a lo largo del desarrollo. Las larvas se caracterizan por tener de tres a cinco melanóforos de tipo dendrítico sobre el borde ventral post-anal, de cuatro a nueve melanóforos más pequeños sobre el borde ventral pre-anal, entre el istmo y el ano, otro melanóforo se presenta a la mitad del margen dorsal de la cola. Hay una pequeña mancha de pigmento en el ángulo de la maxila y otra en la punta del labio inferior. Hay un pigmento interno alargado por debajo de la notocorda, entre la cabeza y la vejiga natatoria. La flexión de la notocorda se inicia a los 3.5mm BL y termina a los 4.6mm BL; la transformación al estadio juvenil es cercana a los 13.6mm BL. Nuestra conclusión es que los caracteres más importantes para distinguir las larvas de esta especie de aquellas similares en el área son el número de miómeros, los melanóforos grandes (aproximadamente iguales en tamaño) en el margen ventral post-anal y el pigmento interno alargado debajo de la notocorda, anterior a la vejiga gaseosa.
Palabras clave: Gobiidae, desarrollo larvario, patrón de pigmentación, ontogenia, larva, Golfo de California.
Studies of the early life history of fishes have been increasing in recent years. Most have the goal of obtaining basic information to understand changes in successful recruitment of the commercial fish populations (Bradford, 1992) or to add useful information for fishes phylogenetic studies (Moser et al. 1984). However, early developmental stages of many fish species have not been described yet. Kendall & Matarese (1994) reported that early development stages have been described for only 44% of fish species in the Eastern Pacific region. Although Moser (1996) increased this percent- age, most of the species considered in that work were from temperate waters, and larvae of the tropical species are still poorly known.
Gobiidae are relatively small fish that inhabit marine, brackish and fresh waters, with a preponderant tropical and subtropical distribution; this is the most species-rich family of marine fishes. Together with the blennioid fishes, gobiids are the dominant element of the benthic fish fauna of tropical seas in coastal areas. There are about 210 gobiid genera with nearly 1 950 species (Nelson, 2006), but other estimates suggest more than 2 000 species (Froese & Pauly, 2013). In ichthyoplankton studies, Gobiidae often is the most abundant family in shallow coastal waters (Grijalva-Chón, Castro-Longoria & Bustamante-Monge, 1992; Arreola-Lizarraga & González-Navarro, 1996; vazquez & Richards, 1999; Parmanne & Lindström, 2003; Costa & Souza-Conseição, 2009). However, information on the early life history of the Gobiidae is known for only 11% of the species (Borges, Faria, Gil & Gonçalves, 2011).
52 gobiid species have been reported from the Northwest coast of México (Castro-Aguirre, 1978; Birdsong, 1981; Allen & Robertson, 1994; González & Saldierna, 1997; Thompson, Findley & Kerstitch, 2000; Ahnelt & Göschl, 2003; Swift, Findley, Ellingson, Flessa & Jacobs, 2011; Froese & Pauly, 2013), but larval stages of only 14 have been described (Watson, 1996). Until now, none of the Evermannia species have been described. The genus Evermannia Jordan, 1895, from temperate waters of the American coast, is characterized by having four spines in the first dorsal fin, one dermal flap in the inner upper part of the branchial chamber, embedded scales that are not imbricate and which are visible only by the use of a microscope.
According to Hoese (1995), this genus includes four species: E. erici Bussing, 1983; E. longipinnis (Steindachner, 1879); E. panamensis Gilbert & Starks, 1904 and E. zosterura (Jordan & Gilbert, 1882). Bussing (1983) mentions that E. longipinnis is of uncertain identity, and a systematic revision of the genus is necessary. All species in this genus are dis- tributed mainly in the Eastern tropical Pacific in a habitat with few millimeters in depth at the intertidal zone of estuaries and sandy beaches. These species are characterized by having four spines in first dorsal fin and one spine and 13-17 rays in the second dorsal fin, one spine and 12-17 rays in the anal fin, 15 segmented rays in the caudal fin, and 30-32 total verte- brae, with 13-14 pre-caudal vertebrae (Bussing, 1983). These species, together with those in the genera Quietula and Ilypnus among others, have the first pterigyophore of the spiny dorsal fin inserted in interneural space 4-5, an unusual condition that distinguishes them from the remaining gobiids in the Pacific Ocean. Furthermore, Ilypnus and Quietula have more spines in the first dorsal fin (V) (Birdsong, Murdy & Pezold, 1988).
E. longipinnis and E. zosterura are found in the Gulf of California, but the later ranges South to Colombian waters. The remaining two species (E. panamensis and E. erici) are found from Southern México to Central America. Adults of E. zosterura are distinguishable from the other species by having a spot in the upper region of the opercle, and in males the first dorsal spine extending into a black filament and the rest of the body with violet tones. This is a colorful fish whose males stand out from other gobiids (Jordan & Evermann, 1898). E. zosterura can be distinguished by a combination of some meristic characters: E. erici, E. panamensis and E. longipinnis have more second dorsal fin rays (E. zosterura 13-15 vs 14-16 in E. erici and E. panamensis, and 16-17 in E. longipinnis) and more pectoral fin rays (E. zosterura 17-19 vs 20-21 in E. erici and 19-21 in E. panamensis), (Bussing, 1983); also E. zosterura has fewer anal fin elements than E. longipinnis (13-14 vs 16-17), (Jordan & Evermann 1898). In this work the larval development from preflexion to juvenile stage of E. zosterura is presented.
Materials and methods
Larvae was caught in Ensenada de La Paz, Gulf of California, México (24º08’00” N-110º25’00” W) with surface zooplankton tows using a conical plankton net of 2m length, 0.60m diameter and a net mesh of 250μm (February 27th, 1988). Additional specimens (larvae, juveniles and adults), were obtained with an epi-benthic net with mouth dimensions of 70x40cm, and a net mesh of 1mm (October 31th, 1991) and with a beach seine of 6x1.20m, and a net mesh of 1mm (September 28th, 1994). All specimens were fixed with a 5% formalin solution buffered with sodium borate, and after this, preserved in 5% formalin solution buffered with sodium phosphate (Markle, 1984).
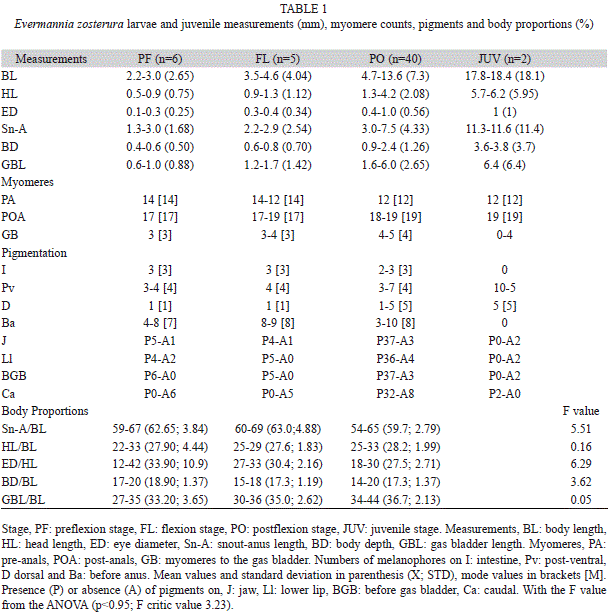
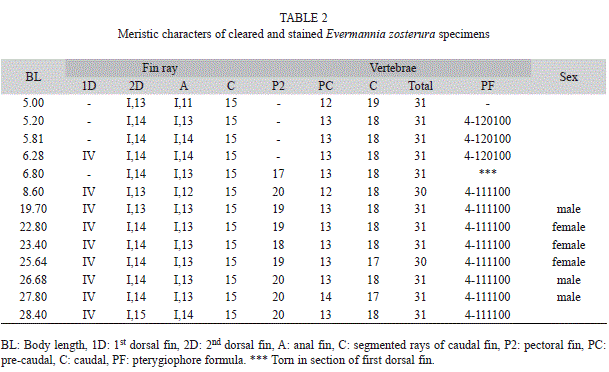
The description of the larvae and juvenile of E. zosterura was based on 53 specimens (51 larvae and two juveniles) between 2.2mm and 18.4mm BL (Table 1). Larvae from preflexion to postflexion stage with similar characteristics, were grouped using the pigmentation pattern (mainly number and shape of melanophores on the post anal ventral region as well as the internal pigment before the gas bladder), general morphology and meristic characteristics such as number of myomeres and fin elements when they were present. Flexion and postflexion stage larvae, juveniles and adults (5.0-28.4mm BL N=13), were cleared and stained according to Potthoff (1984). Larvae were identified as E. zosterura by having 31 total vertebrae and 15 elements in both the second dorsal and the anal fin.
The larval development description follows the three main larval stages proposed by Ahlstrom & Ball (1954): preflexion, flexion, and postflexion stages, plus the juvenile stage. For each of these stages, morphometric measurements were done on the left side of the specimens using a stereomicroscope with a micrometric ocular. The morphometric measurements were: body length (BL), head length (HL), eye diameter (ED) and snout anus length (Sn-A), as described in Moser 1996; body depth (BD), (Neira, Miskiewicz & Trnski, 1998). In addition, gas bladder length (GBL): distance between the tip of the mouth and the posterior margin of the gas bladder.
In order to know if the larval growth was isoor allometric, we calculated the body proportions with respect to BL and HL, and a one-way ANOvA, Sokal & Rohlf (2012), was used to detect statistical significant differences between the body proportions analyzed through the developmental stages.
Results
The cleared and stained specimens had between 12 to 14 pre-caudal vertebrae, 17 to 19 caudal vertebrae and 30 to 31 total vertebrae. From 6mm BL onwards, the fin-ray formula was: IV; I, 13-15 in the dorsal; I, 11-14 in the anal and 18 to 20 rays in the pectoral. The mode of the pterygiophore formula was 4-111100. Using these characteristics, it was determined that the specimens were E. zosterura (Table 2).
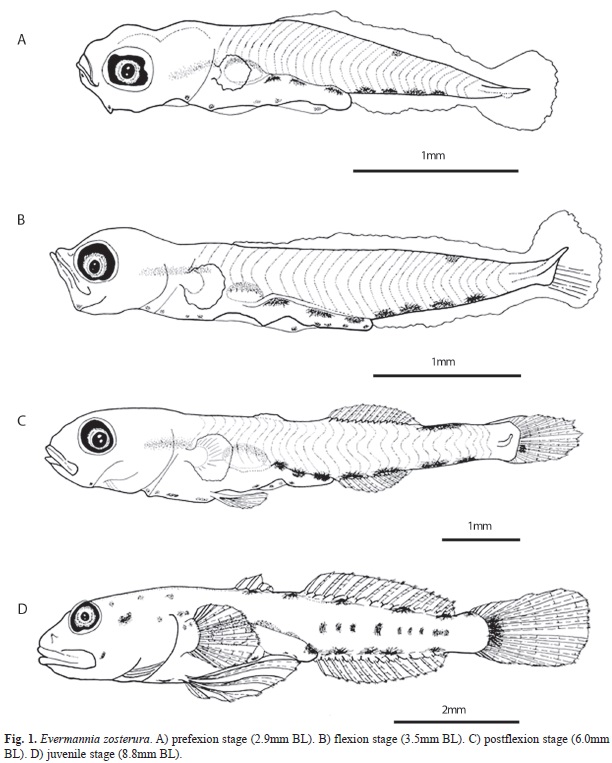
Distinctive characteristics: Larvae of E. zosterura can be recognized from early development by the pigmentation pattern of a series of dendritic melanophores along the mid-ven- tral margin, three to five in the post-anal region and four to nine smaller melanophores in the pre-anal region, and one melanophore located internally under the notochord before the gas bladder. This internal pigment is elongated and seems to be characteristic of the species, but it can be observed only in preflexion to early postflexion stages, since the increased muscular mass of the body obscures it in late postflexion specimens (Fig. 1). Beginning with the preflexion stage, there is a melanophore on the dorsal midline located more or less at the midpoint between the end of the intestine and the caudal fin and it is present until the end of the larval development when up to four additional dorsal melanophores form. The flexion stage begins near 3.5mm BL and is completed near 4.6mm BL. E. zosterura reach the juvenile stage at 13.6mm BL when scales form (Fig. 1A to D).
Morphology: The smallest specimens were 2.2mm BL, lack yolk traces and have ocular pigments which suggest that hatching could occur before this size.
Larval E. zosterura are thin and the snout- anus length is about 60% of the body length. Larval growth is isometric for HL/BL, BD/ BL and GBL/BL (p<0.05) and allometric for Sn-A/BL, and ED/HL, which decrease during development (p<0.05) (Table 1).
Pigmentation: As in almost all gobiid larvae, E. zosterura has a well developed gas bladder pigmented over its dorsal region.
During the preflexion stage (until 3.0mm BL), larvae have three to four (typically four) dendritic melanophores in the post-anal region of the mid-ventral line, which are bigger than the four to nine similar melanophores in the pre-anal region.
There is one melanophore on the mid-dorsal line located about mid-way between the end of the intestine and the end of the notochord. Three melanophores are present on the dorsal surface of the intestine, one on the lower angle of the jaw, and another at the tip of lower jaw. There is an internal, elongated pigment band under the notochord, just before the gas bladder (Fig. 1A).
In the flexion stage (3.5mm≤BL<=4.6mm) the pigmentation pattern is unchanged except that the number of melanophores on the pre-anal mid ventral line increases from eight to ten (Fig. 1B).
During the postflexion stage (4.7mm≤BL <=13.6mm), a pigment appears on the base of the caudal fin and the melanophores on the mid dorsal line increase gradually to five. Both characteristics start to be evident at about 5.6mm BL (Table 1). Late in postflexion stage, a series of internal pigments become apparent over the caudal vertebrae and several pigments are dispersed in areas of the head (Figs. 1C, 1D). Throughout postflexion stage, the pigments on the pre-anal mid-ventral margin start to decrease in number and disappear in the juvenile stage.
The main pigmentation changes in the juvenile stage (BL≥13.6mm) are the increase in the size of the dorsal pigments, and the internal pigments over the vertebrae can increase in size and numbers.
Fin development: The development of the anal and second dorsal fins starts at 3.8mm BL and is completed almost simultaneously with the caudal fin at 4.9mm. The formation of the first dorsal fin starts at 6.0mm BL and is completed at 7.0mm BL.
The pectoral fin buds are present from the preflexion stage, but the rays start to develop at 5.3mm BL and finish at 7.5mm BL. The ventral fins start to develop in synchrony with the pectoral fin rays and they are completely formed at 7.0mm BL.
Discussion
All the specimens of E. zosterura reported in the Panama region by Bussing (1983) had 30 vertebrae, urostyle included. However, in our samples from the Gulf of California, only two of the 13 cleared and stained specimens had 30 vertebrae. Thus, our results extend the range of the total vertebrae to 31 with 12 to 14 pre-caudal and 17 to 19 caudal vertebrae. It is known that the number of vertebrae in fishes can be variable depending on the phylogenetic position, the geographic area as well as the environmental variables such as temperature and salinity among others (Fowler, 1970; Lindsey, 1975, Billerbeck, Orti & Conover, 1997). More information is needed in order to under- stand the reasons of this variability.
This is the first Evermannia species for which the larval stages are described. Among the Gobiidae inhabiting the Eastern tropical Pacific, there is no other species with 30 to 31 myomeres (vertebrae). Some Eastern Pacific Bollmannia, Elacatinus, Gobiosoma and Microgobius have 26 to 28 vertebrae, while species of Ilypnus and Quietula have 32 to 36. Beltrán-León & Ríos-Herrera (2000), present seven forms of gobiid larvae from Colombia, but none of them has the pigmentation pattern or the number of vertebrae or myomeres (all with 27 or less) of E. zosterura.
Larval E. zosterura resemble larval Quietula y-cauda and Ilypnus gilberti which have similar pigmentation patterns and can have 32 myomeres in the study area (personal observations). These hree species are sympatric in the coastal areas of the Gulf of California, causing possible difficulty in identification. The similarity of the morphology and pigmentation pattern is more evident during the earlier larval stages, when it is not easy to count myomeres. However, E. zosterura and Q. y-cauda have three pigments in the dorsal region of the intestine while I. gilberti has only one to two. Also Quietula lacks the internal pigment anteriorly under the notochord until late in the postflexion stage, and even then is less extensive than in E. zosterura.
The most important characteristics that allow differentiating E. zosterura from the other two species at early stages are the following: the internal pigment under the notochord located before the gas bladder; the four to five melanophores located on the post-anal mid-ventral margin, which are uniformly large in E. zosterura but of different sizes in the other two species, as well as the fact that hatching in E. zosterura could occur at smaller sizes (2.2mm BL) than in Q. y-cauda (2.8-3.0mm BL) and I. gilberti (3.1mm BL).
When compared to those of Watson (1996), our results showed that the flexion stage starts and completes at smaller sizes in E. zosterura (near 3.5 to 4.6mm BL), than in Q. y-cauda (5.0 to 5.8mm BL) and I. gilberti (5.0 to 6.0mm BL).
Acknowledments
The authors wish to thank to Deneb Saldierna Cálapiz whom performed the statistical technics. Instituto Politécnico Nacional and Secretaría de Investigación y Posgrado provided financial support through the Project SIP 20120878. RSM and GAM thank to Estímulo al Desempeño de la Investigación (EDI), Comisión de Operación y Fomento de Actividades Académicas (COFAA), Concejo Nacional de Ciencia y Tecnología (CONA-CYT) and Sistema Nacional de Investigadores (SNI) for additional support.
References
Ahlstrom, E. H. & Ball, O. P. (1954). Description of eggs and larvae of jack mackerel (Trachurus symmetricus) and distribution and abundance of larvae in 1950 and 1951. Fishery Bulletin U.S., 56, 209-245. [ Links ]
Ahnelt, H. & Göschl, J. (2003). Morphological differences between the eastern Pacific gobiid fishes Quietula guaymasiae and Quietula y-cauda (Teleostei: Gobiidae) with emphasis on the topography of the lateral line system. Cybium, 27(3), 185-197. [ Links ]
Allen, G. R. & Robertson, D. R. (1994). Fishes of the tropical Eastern Pacific. Honolulu: University of Hawaii. [ Links ]
Arreola-Lizárraga, J. A. & González-Navarro, E. (1996). Composición, abundancia y distribución de las larvas de peces en la Ensenada de la Paz, B.C.S., México. Revista de Investigación Científica. Universidad Autónoma de Baja California Sur (Serie Ciencias del Mar), 7(1-2), 23-39. [ Links ]
Beltrán-León, B. S. & Ríos-Herrera, R. (2000). Estadios tempranos de peces del Pacífico Colombiano, Tomo 2. República de Colombia: Ministerio de Agricultura y Desarrollo Rural. INPA. [ Links ]
Billerbeck, J. M., Orti, G., & Conover, D. O. (1997). Latitudinal variation in vertebral number has a genetic basis in the Atlantic silverside, Menidia menidia. Canadian Journal of Fisheries and Aquatic Sciences,54(8), 1796-1801. [ Links ]
Birdsong, R. S. (1981). A review of the gobiid fish Genus Microgobius Poey. Bulletin of Marine Science, 31(2), 267-306. [ Links ]
Birdsong, R. S., Murdy, E. O., & Pezold, F. L. (1988). A study of the vertebral column and median fin osteology in gobioid fishes, with comments on gobioid relationships. Bulletin of Marine Science, 42(2), 174-214. [ Links ]
Borges, R., Faria, C., Gil, F., & Gonçalves, E. J. (2011). Early development of Gobies. In Patzner, R. A., Van Tassell, J. L., Kovačić, M. & Kapoor, B. G. (Eds.), The Biology of Gobies (pp. 403-462). Enfield: Science Publishers. [ Links ]
Bradford, M. J. (1992). Precision of recruitment predictions from early stages of marine fishes. Fishery Bulletin, 90(3), 439-453. [ Links ]
Bussing, W. A. (1983). Evermannia erici, a new burrowing gobiid fish from the Pacific coast of Costa Rica. Revista de Biología Tropical, 31(1), 125-131. [ Links ]
Castro-Aguirre, J. L. (1978). Catálogo sistemático de los peces marinos que penetran en las aguas continentales de México, con aspectos zoogeográficos y ecológicos. Instituto Nacional de Pesca, México. Serie Científica, 19, 1-298. [ Links ]
Costa, M. D. P. & Souza-Conceição, J. M. (2009). Composição e abundância de ovos e larvas de peixes na baía da Babitonga, Santa Catarina, Brazil. Pan-American Journal of Aquatic Sciences, 4(3), 372-382. [ Links ]
Fowler, J. A. (1970). Control of vertebral number in teleosts. an embriological problem. Quarterly Review of Biology, 45(2), 148-167. [ Links ]
Froese, R. & Pauly, D. (2013). Fishbase. World Wide Web electronic publication. Retrieved from: www.fishbase.org [ Links ]
González, E. A. & Saldierna, R. J. (1997). Adiciones y comentarios a los Gobiidae (Pisces) de la Bahía de La Paz, B.C.S. México. Oceánides, 12(2), 117-120. [ Links ]
Grijalva-Chon, J. M., Castro-Longoria, R., & Bustamante- Monge, A. (1992). Distribución, abundancia y diversidad de larvas de peces en la Laguna Costera Santa Rosa, Sonora, México. Ciencias Marinas, 18(2), 153-169. [ Links ]
Hoese, D. F. (1995). Gobiidae. Gobios, chanquetes y guasetas. In W. Fischer, F. Krupp, W. Schneider, C. K. Sommer, E. Carpenter, & V. H. Niem (Eds.), Guía FAO para la Identificación de Especies para los Fines de la Pesca. Pacífico Centro-Oriental (pp.1129-1135.). Rome: FAO. [ Links ]
Jordan, D. S. & Evermann, B. W. (1898). The fishes of North and Middle America: a descriptive catalogue of the species of fish-like vertebrates found in the waters of North America, north of the Isthmus of Panama. Part III. Bulletin United States National Museum, 47, 2183-3136. [ Links ]
Kendall, A. W. Jr. & Matarese, A. C. (1994). Status of early life history descriptions of marine teleosts. Fishery Bulletin, 92(4), 725-736. [ Links ]
Lindsey, C. C. (1975). Pleomerism, the widespread tendency among related fish species for vertebral number correlated with maximum body length. Journal of the Fisheries Research Board of Canada, 32, 2453-2469. [ Links ]
Markle, D. F. (1984). Phosphate buffered formalin for long term preservation of formalin fixed ichthyoplankton. Copeia, 1984(2), 525-528. [ Links ]
Moser, H. G. (Ed.). (1996). The Early Stages of Fishes in the California Current Region. CalCOFI Atlas 33. Lawrence: Allen [ Links ]
Moser, H. G., Richards, W. J., Cohen, D. M., Fahay, M. P., Kendall, A. W. Jr., & Richardson, S. L. (Eds.). (1984). Ontogeny and Systematics of Fishes. Lawrence: Allen. [ Links ]
Neira, F. J., Miskiewicz A. G., & Trnski, T. (1998). Larvae of Temperate Australian Fishes. Laboratory Guide for Larval Fish Identification. Nedlands: University Western Australia. [ Links ]
Nelson, J. S. (2006). Fishes of the World. New Jersey: Wiley. [ Links ]
Parmanne, R. & Lindström, V. (2003). Annual variation in gobiid larval density in the northern Baltic Sea. Journal of Fish Biology, 62, 413-426. [ Links ]
Potthoff, T. (1984). Clearing and staining techniques. In H. G. Moser, W. J. Richards, D. M. Cohen, M. P. Fahay, A. W. Kendall Jr., & S. L. Richardson (Eds.), Ontogeny and Systematics of Fishes (pp. 35-37). Lawrence: Allen. [ Links ]
Sokal, R. R. & Rohlf, F. J. (2012). Biometry: the Principles and Practice of Statistics in Biological Research. New York: W. H. Freeman and Company. [ Links ]
Swift, C. C., Findley, L. T., Ellingson, R. A., Flessa, K. W., & Jacobs, D. K. (2011). The delta mudsucker, Gillichthys detrusus, a valid species (Teleostei: Gobiidae) endemic to the Colorado River Delta, northernmost Gulf of California, Mexico. Copeia, 2011(1), 93-102. [ Links ]
Thompson, D. A., Findley, L. T., & Kerstitch, A. N. (2000). Reef Fishes of the Sea of Cortez. Austin: University of Texas. [ Links ]
Vazquez, L. & Richards, W. (1999). Variación estacional del ictioplancton de la Bahía de la Ascensión, Reserva de la Biosfera de Sian Ka’an (1990-1994). Revista de Biología Tropical, 47(Supl. 1), 197-207. [ Links ]
Watson, W. (1996). Gobiidae. In H. G. Moser (Ed.), The Early Stages of Fishes in the California Current Region. CalCOFI Atlas 33 (pp. 1214-1245). Lawrence: Allen. [ Links ]
*Correspondencia a:
Enrique González-Navarro: Universidad Autónoma de Baja California Sur. Departamento de Biología Marina. Apartado postal 19-B, CP 23080. La Paz, B.C.S., México; enriquegleznavarro@gmail.com
Ricardo Javier Saldierna-Martínez: Instituto Politécnico Nacional. Centro Interdisciplinario de Ciencias Marinas. Departamento de Plancton y Ecología Marina. Apartado postal 592, CP 23000. La Paz, B.C.S., México; rsaldier@ipn.mx
Gerardo Aceves-Medina: Instituto Politécnico Nacional. Centro Interdisciplinario de Ciencias Marinas. Departamento de Plancton y Ecología Marina. Apartado postal 592, CP 23000. La Paz, B.C.S., México; gaceves@ipn.mx
1. Universidad Autónoma de Baja California Sur. Departamento de Biología Marina. Apartado postal 19-B, CP 23080. La Paz, B.C.S., México; enriquegleznavarro@gmail.com
2. Instituto Politécnico Nacional. Centro Interdisciplinario de Ciencias Marinas. Departamento de Plancton y Ecología Marina. Apartado postal 592, CP 23000. La Paz, B.C.S., México; rsaldier@ipn.mx, gaceves@ipn.mx
Received 22-VII-2013. Corrected 10-XI-2013. Accepted 12-XII-2013.


{kind=link}
{kind=link}
{kind=link}