










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.62 n.2 San José Apr./Jun. 2014
Histology and ultrastructure of male reproductive system of the Indian Spiny lobster Panulirus homarus (Decapoda: Palinuridae)
Histología y ultraestructura del sistema reproductor masculino de la langosta espinosa de la India Panulirus homarus (Decapoda: Palinuridae)
Histología y ultraestructura del sistema reproductor masculino de la langosta espinosa de la India Panulirus homarus (Decapoda: Palinuridae)
*Dirección para correspondencia:
Abstract
The spiny lobster Panulirus homarus, distributed along the Southeast and Southwest coasts of India, is an important commercial species having mariculture potential. Despite its importance, the structural and ultrastructure features of male gonads from this species have received scarce attention. Hence this study was aimed to describe the male reproductive tract of the species, using standard histological and electron microscopy techniques. Gonads from 94 specimens of P. homarus ranging in carapace length 37mm-92mm from vizhinjam (Southwest coast of India.) were obtained and processed for the study (Histology-70 numbers & ultrastructure-24 numbers). The male reproductive system consists of paired testis and vas deferens located in the cephalothoracic region. Macroscopically, the reproductive tract was observed in lobsters >35mm carapace length. In immature testis, spermatogonia were seen which measured 6.9-13.8µm in diameter and in the mature testis primary (5.4-5.9µm) and secondary spermatocytes (2.8-3µm) and spermatids (2.2-2.4µm) were present. Each vas deferens consists of proximal and distal portions. The spermatophoric mass begins formation in the proximal vas deferens. In the distal vas deferens the spermatophoric mass containing the spermatozoa are arranged in packets towards the periphery by the gelatinous matrix produced by the typhlosole. Ultrastructurally, the spermatogonia have lamina, nucleus and mitochondria like bodies, the primary spermatocytes have nucleus, dense chromatin and vacuolated cytoplasm and the spermatids have mitochondria, endoplasmic reticulum and centrioles. The endoplasmic reticulum and the nuclear envelope in the spermatids form the acrosome. The radial arms with microtubules are formed in association with the dense endoplasmic reticulum, near the nucleus. The sperm has a spherical structure with the nucleus, lamellar region, spikes and acrosome. This is the first comprehensive report of the structure of the male gametes and spermatogenesis in P. homarus from Indian waters.Rev. Biol. Trop. 62 (2): 533-541. Epub 2014 June 01.
Key words: reproduction, spermatogenesis, spermiogenesis, spermatophore, lobster, Panulirus homarus.
Resumen
La langosta espinosa Panulirus homarus, distribuida a lo largo de las costas sudeste y sudoeste de la India, es una especie de importancia comercial con gran potencial para la mari-cultura. A pesar de su importancia, las características estructurales y ultraestructurales de las gónadas masculinas de esta especie han sido poco estudiadas. Debido a esto, el objetivo de este estudio fue describir el aparato reproductor masculino de dicha especie, utilizando técnicas convencionales de microscopía histológica y electrónica. Se procesaron 94 ejemplares de P. homarus de vizhinjam (costa suroeste de la India) (70 individuos para histología y 24 para ultraestructura), cuyos caparazones variaron de 37 mm a 92 mm de longitud. El sistema reproductor masculino de esta especie consistió en un par de testículos y un conducto deferente situados en la región céfalo-torácica. Macroscópicamente, el aparato reproductor se observó en langostas con una longitud de caparazón >35mm. En testículos inmaduros, la espermatogonia midió 6.9-13.8μm de diámetro y se encontró presente en los testículos maduros primarios (5.4-5.9μm), espermatocitos secundarios (2.8 a 3 μm) y espermátidas (2.2-2.4μm). Cada conducto deferente consistió de porciones proximales y distales. La formación de la masa espermatofórica comienza en los conductos deferentes proximales. En el conducto deferente distal espermatofórico, la masa que contiene los espermatozoides está dispuesta en paquetes hacia la periferia, en una matriz gelatinosa producida por el tiflosol. Ultraestructuralmente, las espermatogonias presentan una lámina, núcleo y mitocondrias, los espermatocitos primarios tienen núcleo, cromatina densa y citoplasma vacuolado, mientras que las espermátidas tienen mitocondrias, retículo endoplasmático y centríolos. En las espermátidas, el retículo endoplásmico y la envoltura nuclear forman el acrosoma. Los brazos radiales con microtúbulos se forman en asociación con el retículo endoplásmico denso, cerca del núcleo. El esperma presenta una estructura esférica con el núcleo, la región laminar, las espinas y el acrosoma. Este documento constituye el primer informe exhaustivo de la estructura de los gametos masculinos y espermatogénesis en P. homarus de la India.
Palabras clave: reproducción, espermatogénesis, espermiogénesis, espermatóforo, langosta, Panulirus homarus.
Spiny lobsters are one of the world’s most valued and highly prized seafood and they support a large commercial fishery in many parts of the world. They inhabit the temperate and tropical seas, but majority of the species and the highest abundance are found in the tropics (Holthuis, 1991). In India, lobsters form an important export commodity although they form just 0.12% of the total marine landings. Panulirus homarus Linnaeus, 1758 is an important commercial spiny lobster species in India, found at a depth between 20-90m. Increase in global demand, high production value and concern on the sustainability of wild stock have prompted research into commercial seed production for replenishing of wild population as well as lobster mariculture (Cox & Johnston, 2003). Wide tolerance of environmental condition and higher growth rate make them a suitable candidate species for commercial aquaculture. The progress of lobster mariculture depends on better understanding of reproductive biology to ensure sufficient supply of high quality larvae. Despite the importance of P. homarus as a candidate species for aquaculture as well as a valuable crustacean fishery resource, knowledge on the basic biology of reproduction of this species is far from well understood (Cobb & Phillips, 1994). Most of the earlier researchers addressed the physiology of female reproduction rather than that of the male reproductive biology, even though the reproductive performance of males also play an equally important role in the productivity of the captive broodstock.
Knowledge of male reproductive biology of P. homarus has mainly come from the morphological investigation of Berry and Heydorn (1970). Later, Radha and Subramoniam (1985) studied the origin and nature of spermatophoric mass of this species. Kooda-Cisco and Talbot (1986) described the fine structure of the cells of the proximal vas deferens, in relation to their secretory role in producing the spermatophore layers. In Panulirus interruptus, the fine structure of the freshly extruded and hardened spermatophores was investigated (Martin et al.,1987). Talbot & Summers (1978) reported the structure of the sperm from Panulirus argus and Panulirus guttatus with special reference to acrosome. Besides these, the ultrastructure of sperm of the genus Homarus, Nephrops and Jasus have been described (Pochon-Masson, 1965; Tudge, Scheltinga & Jamieson, 1986). The internal anatomy and ultrastructure of the male reproductive system of Nephrops norvegicus was reported recently (Rotllant, Ribes, Company & Durfort, 2012). Haley (1986) described the ultrastructure of spermatogenesis in the reef lobster Enoplometopus occidentalis.
However, there has been little information on the male reproductive tract and spermatogenesis of P. homarus from India. The present work hence describes in detail the histology and ultrastructure of the male reproductive tract which may lead to a better understanding of development of male gonads of the species, and consequently permit creation of baseline information for further studies on reproductive biology.
Materials and methods
Sampling area: Live specimens of male P. homarus were obtained from lobster fishers of vizhinjam (Kerala, Southwest India). Lobsters were fished by gill nets and trammel nets from a depth of 80-90m. This study examined 70 and 24 numbers of P. homarus (carapace length 37mm-92mm) for histological and ultrastructure studies, respectively.
Laboratory procedures: The animals were transported to the laboratory in aerated jerry cans (100L capacity) containing seawater of salinity 35ppt. They were maintained in one ton rectangular tanks and fed with fresh frozen diet (mussel: Perna viridis; Clam: Villorita sp.). Parts of testicular lobes and vas deferens were fixed in Davidson’s fixative (Bell & Lightner, 1988) for 24h and later transferred to 50% iso propyl alcohol until processing for histological studies. These tissues were then dehydrated in ascending series of ethanol, embedded in paraffin wax (melting point: 58-62oC) and serially sectioned at 7µm and stained with Mayer’s haematoxylin and eosin. Images were taken under Olympus CX41 light microscope with an Olympus digital camera CAMEDIA C-5050 zoom, at various magnifications.
For transmission electron microscopic studies, the testis, proximal and distal vas deferens were cut into 1mm size and fixed in 3% glutaraldehyde for 4-5h in the refrigerator, then washed in ice cold cacodylate buffer. The tissues were post fixed in freshly prepared 1% osmium tetroxide in 0.1M cacodylate buffer for 2h at 4oC, washed five times in cacodylate buffer for 15 minutes each and refrigerated. The tissues were dehydrated using ascending grades of acetone and infiltrated with Spurr’s low viscosity resin (Spurr, 1969). The tissues were embedded in fresh resin and polymerized at 70 ºC. Ultrathin sections were prepared using a glass knife, and double-stained with uranyl acetate and lead citrate. The stained sections were viewed and photographed with a JEOL JEM 100SX Transmission Electron Microscope.
Results
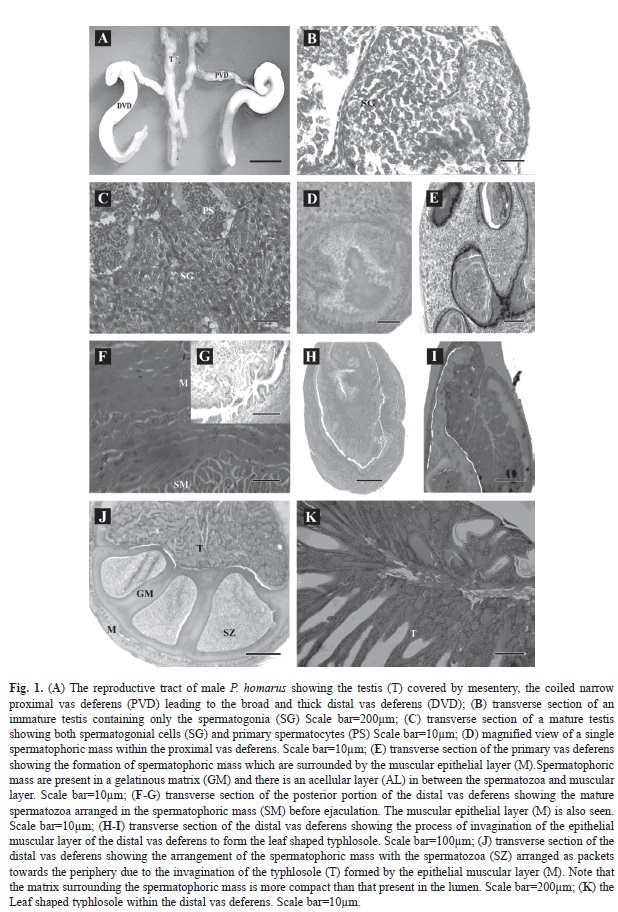
Morphological features of reproductive tract: The male reproductive system located in the cephalo-thoracic region consists of paired testis and vas deferens (Fig. 1A). The semi- transparent ‘H’ shaped testis is surrounded by the mesenteries, and it extends from the level of eyes to the insertion of abdomen. In animals with 35-55mm CL, the testicular lobe if left unwound measure 20-22mm in length and animals 55mm in CL, measure 35-55mm. Each vas deferens extends from the mid region of the testis to the gonopore, located at the base of the fifth walking leg. The two distinct regions of the vas deferens - the proximal region on each side of the body is highly coiled and dull white and the distal vas deferens is thick, opaque and opens out to the gonopore.
Histological observations of germ cells: Under the light microscope the testis has numerous acini with developing follicles, arranged around the central seminiferous duct (Fig. 1B), leading into the proximal vas deferens. The immature testis contains only spermatogonia measuring 6.9-13.8mm in diameter. The primary spermatocytes (5.4-5.9µm), secondary spermatocytes (2.8-3µm) and spermatids (2.2-2.4µm) are present in the mature testis (Fig. 1C). A number of nutritive granules are also present in the acini. They may provide the necessary energy for the developing germ cells. The acini within the testis are surrounded by a thin layer of connective tissue. Spermatophoric mass are present as discrete masses in the proximal vas deferens (Fig. 1E) and are arranged as packets towards the periphery in the distal vas deferens. The glandular epithelial layer of the distal vas deferens form the leaf shaped typhlosole (Fig. 1K) which has columnar epithelium with multiple lobes at the peripheral region. The terminal portion of the distal vas deferens is surrounded by thick muscular wall enclosing packets of sperms ready for ejaculation during mating (Fig. 1F). These spermatophores emerge through the gonopore located at the base of the fifth walking leg and attach to the sternum of the female.
Spermatogenesis: The development of male gametes (spermatogonia through primary and secondary spermatocytes to spermatids) is synchronous in P. homarus, occurs in the acini. In the testis, each acinus has germ cells in varying stages of spermatogenesis. The germ cells are conveyed to the proximal vas deferens from the testis by the seminiferous duct. In the proximal vas deferens, acellular material is secreted from its innermost columnar epithelial layers, around the germ cells to initiate the formation of the spermatophoric mass (Fig. 1D). These secretions coalasce the sperm cells into a coherent mass (Fig. 1E). In the distal vas deferens, the glandular epithelial layer invaginates (Fig. 1H, I) into a leaf like structure and produces matrix that pushes the spermatophoric mass conveyed from the proximal vas deferens, towards the periphery (Fig. 1J). The matrix around the spermatophoric mass is more compact than the matrix in the lumen. Mature sperms within the spermatophore are stored in the terminal end of the distal vas deferens (Fig.1G) until its ejaculation as a sticky mass, during mating. Thus spermatogenesis continues as the spermatocytes travel down the vas deferens from the testis and final form is attained in the distal vas deferens. During the course of its formation, the spermatohore develops several protective layers around it from both the proximal and distal vas deferens.
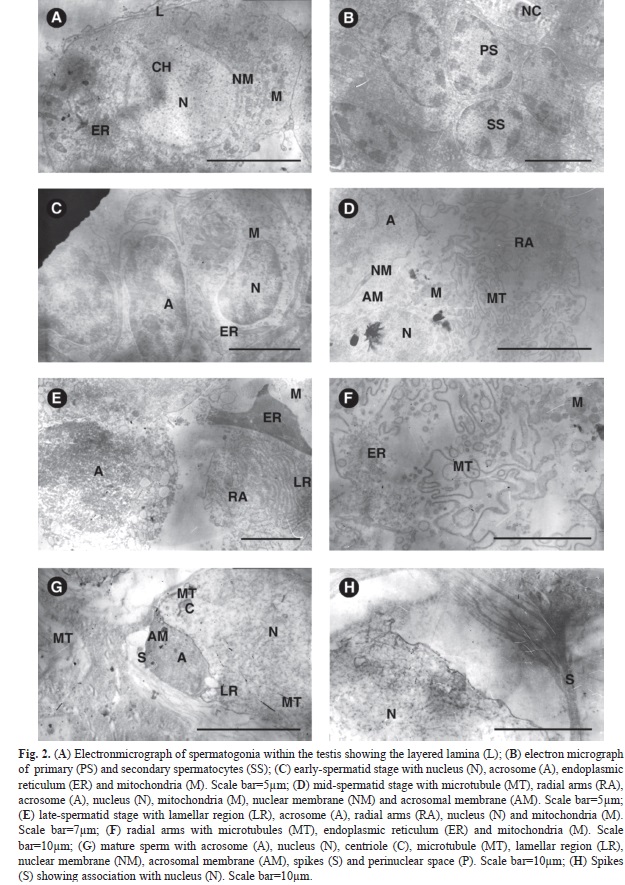
Ultrastructure of germ cells: Under the transmission electron microscope the spermatogonia has four discontinuous layered lamina, nucleus and mitochondria like bodies (Fig. 2A). Lamina is not observed around other germ cells. The spermatogonia are large cells and their nucleus and cytoplasm seem to occupy the cell space almost equally. The nucleus is round in shape with almost evenly distributed chromatin. The primary spermatocytes have nucleus, dense chromatin and vacuolated cytoplasm (Fig. 2B). The secondary spermatocytes are smaller and round with dense chromatin. The mature sperm (Fig. 2G) consists of nucleus, lamellar region, spikes and acrosome with the nucleus occupying the major share of the cell space. The sperm lacks flagella and is surrounded by a membrane. The nuclear boundary is thick and the nucleus and cytoplasm do not seem to be separated from each other. The lamellar region has folds of membranes and lies to one side of the acrosome but outside the nuclear envelope. The acrosome is lens like and surrounded by the acrosomal membrane. The perinuclear material is present near the acrosome along with folds of lamellar region and at the base of the acrosome are the centrioles. Nuclear envelope is not evident at the region where the nucleus and lamellar region meet. Numerous spikes are present in the area anterior to the nucleus and they extend posteriorly (Fig. 2H).
Spermiogenesis: The acrosome and lamellar region are formed in the spermatids and the spermatozoa are produced by gradual changes in the cytoplasm without further division of the spermatids. In early-stage spermatids (Fig. 2C) the acrosome is formed from the coalescence of endoplasmic reticulum and nuclear envelope. The inner and outer membranes of the nuclear envelope fuse and become identical. In middle-stage spermatids, the membrane of the nucleus and the layers of lamellar region become continuous with each other (Fig. 2D). The lamellar region seems to be formed from the association of the nuclear membranes and endoplasmic reticulum. The acrosomal material condenses and becomes electron dense and is present in close contact with the nucleus, with the lamellar region to one side (Fig. 2E). In late-stage spermatids, the nuclear profile differentiates numerous slender arms (Fig. 2F). Radial arms are formed with microtubules containing rounded mitochondria. They are arranged around the nucleus and the acrosome and form the spikes. The spikes contain microtubules which pass through the lamellar region as they extend from one arm to another. The chromatin condenses giving rise to a homogenous fine filament like nucleoplasm in the spermatids. Mitochondria seem to degenerate as they are absent in mature sperms. In the mature sperm, the electron dense acrosome is placed to one side and the fibrillar nucleoplasm occupies most of the space within the sperm.
Discussion
The present study provides a valuable description of the morphology of male gametes and their formation in the spiny lobster P. homarus for the first time in India by transmission electron microscopy. The male reproductive system of P. homarus with minor variations in structure shares similarities with other species of spiny lobsters, Jasus lalandei (Fielder, 1964), P. argus (Talbot & Summers, 1978) and Nephropid lobster N. norvegicus (Rotllant et al., 2012). For example, in P. homarus, the testis is multi-lobed whereas Mathews (1951) described a racimose testis in Panulirus pencillatus with freely branching ducts terminating in acini. The reef lobster E. occidentalis has multiple lobes in the testis (Haley, 1986). P. homarus has synchronous spermatogenesis. In J. lalandei Fielder (1964), crayfish Procambarus clarkii (Lu et al., 1973) and the Carribean spiny lobster P. argus (Melville-Smith, 1987), transverse sections of the mature testis have follicles of varying degrees of maturity. Rotllant et al. (2012) described asynchronous nature of testis in N. norvegicus with only one type of germinal cells. The general anatomical design of the testis and vas deferens of P. homarus differs from that described in the freshwater crayfish P. clarkii-tri-lobate testis and single vas deferens (Scovacricchi, Luise & Magni, 1998; Mirheydari et al., 2012) and bi-lobate testis with longer, thicker, coiled right vas deferens, short, transparent, straight left vas deferens (Huner & Barr, 1991; Saad & Hassan, 2010).
The highly coiled proximal vas deferens and their secretions in P. homarus signify their secretory nature. In decapod crustaceans, the highly coiled proximal vas deferens may serve to increase the surface area for storage, secretory and absorptive function (Adiyodi & Anilkumar, 1988). In P. homarus, the typhlosole shares similarity with P. homarus from African waters (Berry, 1970), whereas differ in the nature of the extruded spermatophoric mass, which is sticky and thick in P. homarus and granular in the African species. Berry and Heydorn (1970) described the typhlosole in Panulirus gilchristi to have extensive proliferation of the epithelium consisting of a central core of muscle and connective tissue merely lined with glandular epithelium. The homogenous secretion of the typhlosole in P. homarus, pushes the spermatophoric mass towards the periphery as observed in the Carribean spiny lobster P. argus (Haley, 1986). Unlike in P. homarus, the spermatophore in J. lalandei (Berry & Heydorn, 1970) is convoluted randomly throughout the matrix but shares similarity in the storage and ejaculatory nature of the terminal region of the distal vas deferens as evident from the circular thick muscle wall. The extruded spermatophore hardens and remains attached to the ventral side of the female sternum until fertilisation which is external. Hardening of the spermatophore protects the sperms until spermatophore rupture/dehiscence. Sperm attain their final form in the distal vas deferens as in the brachyuran crab Charybdis smithii (Balasubramanian & Suseelan, 2000), pandalid shrimp Pandalopsis japonicus (Medina, 1994) but unlike in the crab Uca uruguayensis (Cuartas & Petriella, 1995), the spermatophore are formed at the junction of the testis and vas deferens and in Homarus americanus (Kooda-Cisco & Talbot, 1982) and N. norvegicus (Rotllant et al., 2012), they are completely formed in the testis.
The spermatozoa of P. homarus, has a nucleus, lens shaped acrosome, spikes and microtubules. Studies show the decapod sperm to consist of an acrosome, a cup shaped nucleus with several radiating processes and varying quantities of such cellular organelles as centrioles, mitochondria and microtubules (Moses, 1960; Brown, 1966; Hinsch, 1969; Rotllant et al., 2012). Numerous studies of crustacean sperm (Brown, 1966; Reger, 1970) also indicate that species differences occur regarding the presence of these organelles.
The fine structure of the spermatogonia has never been described in the genus Panulirus. The present study reports four discontinuous layers of lamina around the spermatogonia, in contrast to the 2-14 layers described in E. occidentalis (Haley, 1986) which he opined might serve to limit the steroid producing meiotic events in spermatogenesis from spermatogonia.
The presence of electron opaque and granular nucleoplasm of the spermatozoa has been reported in other lobsters such as Homarus vulgaris (Pochon-Masson, 1965) and N. norvegicus (Chevaillier & Maillet, 1965). The lens shaped structurally complex acrosome that lacks acrosomal tubule is similar to Homarus and brachyuran crabs. In the present study the number of layers limiting the boundary of the mature sperm could not be distinguished although layers were discerned around the spermatogonia. Chevaillier (1966) observed tight limiting boundary in the hermit crab Eupagurus bernhardus, which he presumed to be tripartite. Sperm data provides valuable information for determining relationships between crustacean taxa. In certain groups, ultrastructural variations in sperm morphology are highly significant so that some classifications have been questioned on this ground, like the appearance of microtubule bundles in the spermatozoan of P. japonicus is an exception among dendrobranchiate, which is of phylogenetic interest (Medina, 1994).
This study prove that the histology and ultrastructure of the reproductive tract in P. homarus has features common to the genus and congeneric species and provides information for further studies on reproductive biology. Future extensive studies on other species in the region may lead to valuable information and contribute to our understanding of the genus.
Acknowledgments
The first author thanks E. V. Radhakrishnan, former Head of Crustacean Fisheries Division, CMFRI for his suggestions and Pushpa vishwanathan, Cancer Research Institute, Chennai for assistance with the Electron microscope.
References
Adiyodi, K. G. & Anilkumar, G. (1988). Arthropoda-Crustacea. In K. G. Adiyodi & R. G. Adiyodi (Eds.), Reproductive Biology of Invertebrates: Accessory sex glands (vol. 3, pp. 261-318). New Delhi, India: Oxford and IBH Publishing Co. [ Links ]
Balasubramanian, C. P. & Suseelan, C. 2000. Male reproductive system and spermatogenesis in the deepwater crab Charybdis smithii McLeay (Brachyura: Portunidae). Indian Journal of Fisheries, 47(4), 275-282. [ Links ]
Bell, T. A. & Lightener, D. V. (1988). A handbook of normal penaeid shrimp histology. Baton Rouge, Lousiana, USA: World Aquaculture Society. [ Links ]
Berry, P. F. (1970). Mating behaviour, oviposition and fertilization in the spiny lobster Panulirus homarus (Linnaeus). Oceanographic Research Institute (Durban) Investigation Report, 24, 1-16. [ Links ]
Berry, P. F. & Heydorn, A. E. F. (1970). A comparison of the spermatophoric masses and mechanisms of fertilization in Southern African spiny lobsters (Palinuridae). South Africa Oceanographic Research Institute Investigation Report, 25, 1-18. [ Links ]
Brown, G. G. (1966). Ultrastructural studies of sperm morphology and spermegg interaction in the decapod, Callinectes sapidus. Journal of Ultrastructure Research, 14, 425-440. [ Links ]
Chevaillier, P. & Maillet, P. L. (1965). Structure fine et constitution cytochimique du spermatozoide de la Langoustine Nephrops norvegicus L. (Crustace, Decapode). Journal of Microscopy, 4, 679-700. [ Links ]
Chevaillier, P. (1966). Structure et constitution cytochimique de la capsule du spermatozoide des Crustaces Decapodes. Paris, France: C.R. Academic Science. [ Links ]
Cobb, J. S. & Phillips, B. F. (1994). In B. F Phillips, J. S Cobb, & J. Kittaka (Eds.), Spiny Lobster Management (pp. 529-535). Oxford, England: Fishing News Book. [ Links ]
Cox, S. L. & Johnston, D. J. (2003). Developmental changes in the structure and function of mouth parts of phyllosoma larvae of packhorse lobster, Jasus verreauxi (Decapoda; Palinuridae). Journal of Experimental Marine Biology & Ecology, 296, 35-47. [ Links ]
Cuartas, E. I. & Petriella, A. M. (2007). Initial formation of spermatophores in the testicles of the crab Uca uruguayensis (Brachyura: Ocypodidae). Revista de Biología Tropical, 55(Suppl. 1), 9-14. [ Links ]
Fielder, D. R. (1964). The spiny lobster, Jasus lalandii (H. Milne Edwards) in South Australia. II. Reproduction. Australian Journal of Marine & Freshwater Research, 15, 133-144. [ Links ]
Haley, S. R. (1986). Spermatogenesis and spermatophore production in the Hawaiian red lobster Enoplometopus occidentalis (Randall) (Crustacea, Nephropidae). Journal of Morphology, 180, 181-193. [ Links ]
Hinsch, G. W. (1969). Microtubules in the sperm of the spider crab Libinia emarginata. Journal of Ultrastructure Research, 29, 525-534. [ Links ]
Holthuis, L. B. (1991). FAO species catalogue. Marine lobsters of the world. FAO Fisheries synopsis. Rome, Italy: Food and Agriculture Organization of the United Nations. [ Links ]
Huner, J. V. & Barr, Z. E. (1991). Red swamp crawfish biology and exploitation (3rd ed.). Baton Rouge, Louisiana: Louisiana Sea Grant College Program Centre for Wetland Resources Louisiana State University. [ Links ]
Kooda-Cisco, M. J. & Talbot, P. (1982). A structural analysis of the freshly extruded spermatophore from the lobster, Homarus americanus. Journal of Morphology, 172, 193-207. [ Links ]
Kooda-Cisco, M. J. & Talbot, P. (1986). Ultrastructure and role of the lobster vas deferens in spermatophore formation. The proximal segment. Journal of Morphology, 188(1), 91-103. [ Links ]
Lu, C. C., Clark, W. H., & Franklin, L. E. (1973). Spermatogenesis of the decapod Penaeus setiferus. Journal of Cell Biology, 59, 202. [ Links ]
Martin, G. G, Herzeg, C., & Narimatsu, G. (1987). Fine structure and histochemistry of the freshly extruded and hardened spermatophore of the spiny lobster Panulirus interruptus. Journal of Morphology, 192, 237-246. [ Links ]
Mathews, D. C. (1951). The origin, development and nature of the spermatophoric mass of the spiny lobster, Panulirus pencillatus (Olivier). Pacific Science, 5, 359-371. [ Links ]
Medina, A. (1994). Spermiogenesis and sperm structure in the shrimp Parapenaeus longirostris (Crustacea: Dendrobranchiata): Comparative aspects among decapods. Marine Biology, 119(3), 449-460. [ Links ]
Melville-Smith, R. (1987). The reproductive biology of Greyon maritae (Decapoda, Brachyura) of Southwest Africa, Namibia. Crustaceana, 53, 259-275. [ Links ]
Mirheydari, S. M., Matinfar, A., Soltani, M., Kamali, A., Ousalou, Y. A. & Roomiani, L. (2012). Survey of seasonal histology of male reproductive organ in narrow clawed crayfish A. leptodactylus in Aras Dam Lake, Iran. World Journal of Fish and Marine Sciences, 4(6), 692-701. [ Links ]
Moses, M. J. (1960). A light and electron microscope study of spermiogenesis in decapod crustacea. Anatomical Records, 130, 343. [ Links ]
Pochon-Masson, J. (1965). Schema general du spermatozoide vesiculaire des decapods. Paris, France: C.R. Academic Science. [ Links ]
Radha, T. & Subramoniam, T. (1985). Origin and nature of spermatophoric mass of the spiny lobsters, Panulirus homarus. Marine Biology, 86, 13-19. [ Links ]
Reger, J. F. (1970). Studies on the fine structure of sper- matids and spermatozoa of the crab, Pinmixia sp. Journal of Morphology, 132, 89-100. [ Links ]
Rotllant, G., Ribes, E., Company, J. B., & Durfort, M. (2012). Internal anatomy and ultrastructure of the male reproductive system of the Norway lobster Nephrops norvegicus (Decapoda: Astacidea). Journal of Morphology, 273, 572-585. [ Links ]
Saad, M. M. & Hassan, M. M. (2010). Anatomical and histological studies on the male reproductive system of the red swamp crayfish Procambarus clarkii. Egyptian Journal of Aquatic Biology & Fisheries, 14(1), 87-100. [ Links ]
Scovacricchi, T., de Luise, G., & Magni, A. (1998). Notes on the presence of a single sperm duct in Procambarus clarkii (Girard, 1852) (Decapoda: Cambaridae) from Italian waters. Journal of Natural History, 32(10-11), 1747-1751. [ Links ]
Spurr, A. R. (1969). A low viscosity epoxy resin embedding medium for electron microscopy. Journal of Ultrastructure Research, 26, 31-43. [ Links ]
Talbot, P. & Summers, R. G. (1978). The structure of sperm from Panulirus, the spiny lobster with special regard to the acrosome. Journal of Ultrastructure Research, 64, 341-351. [ Links ]
Tudge, C. C., Scheltinga, D. M., & Jamieson, G. M. (1986). Spermatozoal ultrastructure in the spiny lobster Jasus novaehollandiae Holthuis, 1963 (Palinuridae, Palinura, Decapoda). Journal of Morphology, 236(2), 117-126. [ Links ]
Balasubramanian, C. P. & Suseelan, C. 2000. Male reproductive system and spermatogenesis in the deepwater crab Charybdis smithii McLeay (Brachyura: Portunidae). Indian Journal of Fisheries, 47(4), 275-282. [ Links ]
Bell, T. A. & Lightener, D. V. (1988). A handbook of normal penaeid shrimp histology. Baton Rouge, Lousiana, USA: World Aquaculture Society. [ Links ]
Berry, P. F. (1970). Mating behaviour, oviposition and fertilization in the spiny lobster Panulirus homarus (Linnaeus). Oceanographic Research Institute (Durban) Investigation Report, 24, 1-16. [ Links ]
Berry, P. F. & Heydorn, A. E. F. (1970). A comparison of the spermatophoric masses and mechanisms of fertilization in Southern African spiny lobsters (Palinuridae). South Africa Oceanographic Research Institute Investigation Report, 25, 1-18. [ Links ]
Brown, G. G. (1966). Ultrastructural studies of sperm morphology and spermegg interaction in the decapod, Callinectes sapidus. Journal of Ultrastructure Research, 14, 425-440. [ Links ]
Chevaillier, P. & Maillet, P. L. (1965). Structure fine et constitution cytochimique du spermatozoide de la Langoustine Nephrops norvegicus L. (Crustace, Decapode). Journal of Microscopy, 4, 679-700. [ Links ]
Chevaillier, P. (1966). Structure et constitution cytochimique de la capsule du spermatozoide des Crustaces Decapodes. Paris, France: C.R. Academic Science. [ Links ]
Cobb, J. S. & Phillips, B. F. (1994). In B. F Phillips, J. S Cobb, & J. Kittaka (Eds.), Spiny Lobster Management (pp. 529-535). Oxford, England: Fishing News Book. [ Links ]
Cox, S. L. & Johnston, D. J. (2003). Developmental changes in the structure and function of mouth parts of phyllosoma larvae of packhorse lobster, Jasus verreauxi (Decapoda; Palinuridae). Journal of Experimental Marine Biology & Ecology, 296, 35-47. [ Links ]
Cuartas, E. I. & Petriella, A. M. (2007). Initial formation of spermatophores in the testicles of the crab Uca uruguayensis (Brachyura: Ocypodidae). Revista de Biología Tropical, 55(Suppl. 1), 9-14. [ Links ]
Fielder, D. R. (1964). The spiny lobster, Jasus lalandii (H. Milne Edwards) in South Australia. II. Reproduction. Australian Journal of Marine & Freshwater Research, 15, 133-144. [ Links ]
Haley, S. R. (1986). Spermatogenesis and spermatophore production in the Hawaiian red lobster Enoplometopus occidentalis (Randall) (Crustacea, Nephropidae). Journal of Morphology, 180, 181-193. [ Links ]
Hinsch, G. W. (1969). Microtubules in the sperm of the spider crab Libinia emarginata. Journal of Ultrastructure Research, 29, 525-534. [ Links ]
Holthuis, L. B. (1991). FAO species catalogue. Marine lobsters of the world. FAO Fisheries synopsis. Rome, Italy: Food and Agriculture Organization of the United Nations. [ Links ]
Huner, J. V. & Barr, Z. E. (1991). Red swamp crawfish biology and exploitation (3rd ed.). Baton Rouge, Louisiana: Louisiana Sea Grant College Program Centre for Wetland Resources Louisiana State University. [ Links ]
Kooda-Cisco, M. J. & Talbot, P. (1982). A structural analysis of the freshly extruded spermatophore from the lobster, Homarus americanus. Journal of Morphology, 172, 193-207. [ Links ]
Kooda-Cisco, M. J. & Talbot, P. (1986). Ultrastructure and role of the lobster vas deferens in spermatophore formation. The proximal segment. Journal of Morphology, 188(1), 91-103. [ Links ]
Lu, C. C., Clark, W. H., & Franklin, L. E. (1973). Spermatogenesis of the decapod Penaeus setiferus. Journal of Cell Biology, 59, 202. [ Links ]
Martin, G. G, Herzeg, C., & Narimatsu, G. (1987). Fine structure and histochemistry of the freshly extruded and hardened spermatophore of the spiny lobster Panulirus interruptus. Journal of Morphology, 192, 237-246. [ Links ]
Mathews, D. C. (1951). The origin, development and nature of the spermatophoric mass of the spiny lobster, Panulirus pencillatus (Olivier). Pacific Science, 5, 359-371. [ Links ]
Medina, A. (1994). Spermiogenesis and sperm structure in the shrimp Parapenaeus longirostris (Crustacea: Dendrobranchiata): Comparative aspects among decapods. Marine Biology, 119(3), 449-460. [ Links ]
Melville-Smith, R. (1987). The reproductive biology of Greyon maritae (Decapoda, Brachyura) of Southwest Africa, Namibia. Crustaceana, 53, 259-275. [ Links ]
Mirheydari, S. M., Matinfar, A., Soltani, M., Kamali, A., Ousalou, Y. A. & Roomiani, L. (2012). Survey of seasonal histology of male reproductive organ in narrow clawed crayfish A. leptodactylus in Aras Dam Lake, Iran. World Journal of Fish and Marine Sciences, 4(6), 692-701. [ Links ]
Moses, M. J. (1960). A light and electron microscope study of spermiogenesis in decapod crustacea. Anatomical Records, 130, 343. [ Links ]
Pochon-Masson, J. (1965). Schema general du spermatozoide vesiculaire des decapods. Paris, France: C.R. Academic Science. [ Links ]
Radha, T. & Subramoniam, T. (1985). Origin and nature of spermatophoric mass of the spiny lobsters, Panulirus homarus. Marine Biology, 86, 13-19. [ Links ]
Reger, J. F. (1970). Studies on the fine structure of sper- matids and spermatozoa of the crab, Pinmixia sp. Journal of Morphology, 132, 89-100. [ Links ]
Rotllant, G., Ribes, E., Company, J. B., & Durfort, M. (2012). Internal anatomy and ultrastructure of the male reproductive system of the Norway lobster Nephrops norvegicus (Decapoda: Astacidea). Journal of Morphology, 273, 572-585. [ Links ]
Saad, M. M. & Hassan, M. M. (2010). Anatomical and histological studies on the male reproductive system of the red swamp crayfish Procambarus clarkii. Egyptian Journal of Aquatic Biology & Fisheries, 14(1), 87-100. [ Links ]
Scovacricchi, T., de Luise, G., & Magni, A. (1998). Notes on the presence of a single sperm duct in Procambarus clarkii (Girard, 1852) (Decapoda: Cambaridae) from Italian waters. Journal of Natural History, 32(10-11), 1747-1751. [ Links ]
Spurr, A. R. (1969). A low viscosity epoxy resin embedding medium for electron microscopy. Journal of Ultrastructure Research, 26, 31-43. [ Links ]
Talbot, P. & Summers, R. G. (1978). The structure of sperm from Panulirus, the spiny lobster with special regard to the acrosome. Journal of Ultrastructure Research, 64, 341-351. [ Links ]
Tudge, C. C., Scheltinga, D. M., & Jamieson, G. M. (1986). Spermatozoal ultrastructure in the spiny lobster Jasus novaehollandiae Holthuis, 1963 (Palinuridae, Palinura, Decapoda). Journal of Morphology, 236(2), 117-126. [ Links ]
*Correspondencia a:
S. Lakshmi Pillai: Central Marine Fisheries Research Institute, P.B.No.1603, Ernakulam North, P.O., Kochi-18, India;slakshmipillai@rediffmail.com
M. Nasser: Department of Zoology, University of Calicut, Thenipalam, Kerala, India; drnasher@gmail.com
N. K. Sanil: Central Marine Fisheries Research Institute, P.B.No.1603, Ernakulam North, P.O., Kochi-18, India; nksanil@gmail.com
1. Central Marine Fisheries Research Institute, P.B.No.1603, Ernakulam North, P.O., Kochi-18, India;slakshmipillai@rediffmail.com , nksanil@gmail.com
2.Department of Zoology, University of Calicut, Thenipalam, Kerala, India; drnasher@gmail.com
Received 24-IV-2013. Corrected 10-X-2013. Accepted 15-XI-2013.


{kind=link}
{kind=link}