










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.62 n.2 San José Apr./Jun. 2014
Variability of coastal suprabenthic assemblages from sandy beaches of the Caribbean coast of Venezuela
Variabilidad de agregaciones suprabetónicas costeras de playas arenosas de la costa Caribe de Venezuela
Variabilidad de agregaciones suprabetónicas costeras de playas arenosas de la costa Caribe de Venezuela
*Dirección para correspondencia:
Abstract
The suprabenthos or hyperbenthos is the macrofaunal assemblage of small-sized organisms that interact for some time in the benthic boundary layer. Information about the taxonomic composition and role of suprabenthic species, especially in littoral zones, is scarce and scattered. This work attempts to contribute alleviate this problem. We analyze the temporal and spatial variations of suprabenthic assemblages in the swash-zone from four beaches of the littoral coast of Venezuela. For each beach, two sites were chosen, and special attention was given to water and sediment characteristics. 12 environmental variables were measured: Dissolved oxygen, oxygen saturation percentage, pH, salinity, surface temperature, total, organic and inorganic suspended solids, total organic carbon, organic matter in sediment, grain size of sediment, and amount of dragged material of sample. All faunal samples were taken on a monthly basis during 2011; these were extracted using a manual suprabenthic sledge towed parallel to the shoreline. Samples were sorted and identified to their lowest possible taxonomic level. A total of 24 141 specimens (mean abundance: 26.16±55.35ind./m2) belonging to 21 taxonomic groups were identified. Analysis suggests that seasonality does not explain observed changes either in fauna or environmental variables. It was found that suprabenthic assemblages, total suprabenthos density, richness and environmental variables changed in a dissimilar fashion between months and beaches. The most frequent groups were amphipods and decapods; and at the species/categories level post-larval shrimp (Penaeidae), Grapsidae crab megalopae and Arenaeus cribarius megalopae were common. Dissimilarity between months in each beach was primarily explained by the abundance of amphipods, ctenophores, decapods and mysids. For particular months and selected beaches very high abundances of ctenophores were found. This group dominated the sample even though it is not usually a representative group in suprabenthos. Samples showed low correlations between suprabenthos and environmental variables. A somewhat stronger correlation could be established between water characteristics and dragged material abundance. The studied suprabenthos assemblage was found to have high taxa richness and very dynamic behaviour at spatial and temporal scale. Further analysis suggested that there is no evident pattern of distribution and that causality can not be directly attributed to temporal variation only. Possibly there is an influence of a synergy of environmentals or biological factors, rather than a single variable. The species Americamysis bahia and Americamysis taironana are reported for the first time in Venezuela. This study represents the first ecological research of the suprabenthos in the Caribbean region.
Key words: suprabenthos, assemblage structure, environmental variables, sandy beach, spatial variations, temporal variations, Venezuelan littoral coast.
Resumen
El suprabentos o hiperbentos es la agregación de organismos de pequeño tamaño que interactúan por cierto tiempo en la capa de límite bentónico. La información de la composición taxonómica y el papel de las especies suprabentónicas, especialmente en la zona litoral, es escasa. Este trabajo trata de contribuir a solventar este problema. Se analizó la variación espacial y temporal de la agregación suprabentónica en la zona de rompiente de cuatro playas en la costa litoral de Venezuela. Se dio especial atención al sedimento y características del agua. Todas las muestras fueron tomadas mensualmente durante el 2011. Las muestras fueron extraídas utilizando un trineo suprabentónico manual paralelo a la línea de costa. En cada playa se escogieron dos sitios. Las muestras fueron separadas e identificadas hasta el nivel taxonómico más bajo posible. Se midieron doce variables ambientales: oxígeno disuelto, porcentaje de saturación de oxígeno, pH, salinidad, temperatura superficial, sólidos suspendidos totales, inorgánicos y orgánicos, carbono orgánico total, materia orgánica en sedimento, tamaño del grano de sedimento y cantidad de material arrastrado en cada muestra. Se identificaron un total de 24 141 individuos (densidad promedio: 26.16±55.35ind./m2), pertenecientes a 21 grupos taxonómicos. Los análisis sugieren que la estacionalidad no explica los cambios observados en la fauna ni en las variables ambientales. La agregación del suprabentos, la densidad total, riqueza y variables ambientales cambiaron de manera diferente entre meses y playas. Los grupos más frecuentes fueron anfípodos y decápodos. A nivel de especies/categorías fueron las post-larvas de camarón (Penaeidae), las megalopas de cangrejos Grapsidae y de Arenaeus cribarius (Lamarck, 1818). La disimilitud entre meses en cada playa se debe principalmente a la abundancia de anfípodos, ctenóforos, decápodos y misidáceos. En meses particulares y algunas playas, se encontraron altas abundancias de ctenóforos. Este grupo dominó esas muestras, aún cuando no son frecuentemente un grupo representativo del suprabentos. Se encontraron bajas correlaciones entre el suprabentos y las variables ambientales. Una relación un poco más fuerte fue establecida con las características del agua y la abundancia de material arrastrado. La agregación de suprabentos estudiada tuvo una alta riqueza taxonómica y fue muy dinámica tanto espacial como temporalmente. Los análisis sugieren que no hay un patrón evidente de distribución y el azar no puede ser atribuido sólo a la variación temporal. Posiblemente hay influencia de una sinergia de factores ambientales o biológicos, más que de una variable en particular. Las especies Americamysis bahia (Molenock, 1969) y Americamysis taironana (Brattegard, 1973) se reportan por primera vez para Venezuela. Este es el primer estudio ecológico del suprabentos en el Mar Caribe.
Palabras clave: suprabentos, agregación, variables ambientales, playa arenosa, variaciones espaciales, variaciones temporales, costa litoral venezolana.
The suprabenthos or hyperbenthos communities are macrofaunal assemblages of small-sized organisms living in the benthic boundary layer for different periods of their lives or at different times of the day (Mees & Jones, 1997; Ligas et al., 2007). Suprabenthos may also be considered as the assemblage composed of small swimming animals, mainly crustaceans, that live directly above the sediment and can migrate on a daily or seasonal basis (Brunel et al., 1978; Munilla & San Vicente, 2005). This complex assemblage is mainly composed of permanent suprabenthos and other taxa which have a more direct relation with water-sediment interface (Fanelli, 2007).
This has been studied in many environments, from estuaries to abyssal zones (Mees & Jones, 1997). These communities are a major link in coastal food webs. They consume detritus, algae and zooplankton, and are prey for demersal fish and shrimps (Cartes & Sorbe, 1999; Cartes, Grémare, Maynou, Villora-Moreno, & Dinet, 2002; Fanelli et al., 2011).
Despite the importance of this assemblage in the coastal ecosystem dynamics, information about the taxonomic composition is scarce and scattered (Cartes, Papiol, Palanques, Guillen, & Demestre, 2007). Most studies concerning the suprabenthos are conducted in temperate areas as the Mediterranean Sea (Munilla & Corrales, 1995; Cartes, 1998; Munilla, Corrales, & San Vicente, 1998; Madurell & Cartes, 2003; Fanelli, Cartes, Badalamenti, Rumolo, & Sprovieri, 2009; Fanelli et al., 2011). While studies in tropical waters is quite limited, for example Melo et al. (2010) in Brazil and Domínguez et al. (2004) in Ecuador. Unfortunately, these kind of studies for the Caribbean Sea are non-existent, and this work represents the first ecological research of suprabenthos for this region.
Coastal and estuarine environments are typically subjected to major environmental fluctuations. These are associated with a variety of oceanographic processes: changes in water masses (temperature, salinity, river inputs, among others), pulses of organic matter derived from primary production, and vertical fluxes with direct influence on the suprabenthos dynamics (Cartes et al., 2007; Cartes, Ligas, De Biasi, Pacciardi, & Sartor, 2009). Composition and richness of communities that inhabit the swash zone are influenced by physical and temporal variations. Physical fluctuations include the degree of wave exposure, sediment grain size and water turbity (Clark, Bennett, & Lamberth, 1996). While temporal variations take into account seasonal changes, moon phases, tide levels and diurnal changes (Marín, 2007). Given these influences, it is paramount to have a detailed knowledge of the beaches’ main characteristics in order to understand the variability patterns of the marine communities.
We address questions on community composition changes (temporal and spatial), exploring the relationships between these changes with respect to seasonality and environmental variables. The beaches studied in this work present a high variability in hydrodynamics, beach characteristics and different waterbodies sources, there should be an evident variability between fauna. Strong relationship can also be expected between faunal temporal variation and precipitation. Given the high turbulence observed in the swash zone it is anticipated that water and sediment variables should influence the composition of the suprabenthic fauna.
In this work we investigated the spatial and temporal variability of suprabenthic assemblages in the swash zone of four beaches in the Venezuelan Caribbean coast, including their relationship with some environmental variables.
Material and Methods
Study area: The area encompasses a large stretch of sandy beaches that extends from Cabo Codera to Barcelona in the Caribbean coast of Venezuela. Surveys were conducted at four sandy beaches: Agua Sal (10°28’00” N - 66°05’04” W), Los Timones (10°28’44” N - 66°05’38” W), Las Cabañas (10°29’45” N - 66°06’27” W) and Valle Seco (10°31’19” N - 66°06’56” W). They are characterized by fine sands, a dissipative profile of intermediate energy (depending on the season), a relatively low subtidal slope and having a small decline (Bone et al., 1998). The beaches have a high turbidity due to the nutrients received from the Curiepe, Capaya and Tuy rivers (Bone et al., 1998). Additionally, this area is influenced by upwelling events originated from the nearby Cariaco Trench, located at Northeast of the study area. These events generate phytoplank-ton blooms and sediment resuspension (Herrera & Bone, 2011).
Suprabenthos sampling: Organisms were collected from the swash-zone (<1m depth) of each beach on a monthly basis during 2011. Samples were extracted using a manual suprabenthic sledge (mouth aperture: height 20cm×width 50cm) equipped with a 500μm mesh plankton net. Towing for fauna extraction was performed by a single operator wading the swash zone parallel to the shoreline. The distance covered was 5m and determined by a pilot sample. At each beach two sampling sites were chosen to be approximately separated by 120m. At each site four replicate tows were performed. All collected material was preserved in 10% buffered formaldehyde. Samples were sorted and identified to their lowest possible taxonomic level.
We used general zooplankton books for differentiation among taxonomic groups, like Boltovskoy (1981), Harris, Wiebe, Lenz, Skjoldal, & Huntley (2000). For some groups we also used specialized keys such as: Poly-chaetes: Fauchald (1977), Uebelacker & Johnson (1984), Delgado-Blas (2009); Copepods: Campos & Suarez (1994); Amphipods: Díaz (2001), Ortiz, Martín, Winfield, Díaz, & Atienza (2004); Isopods: Kensley & Schotte (1989); Cumaceans: Heard, Roccagliata, & Petrescu (2007); Misids: Brattegard (1969, 1970a,b, 1973, 1974a,b, 1975), Price (1982), Heard, Price, Knott, King, & Allen (2006); Decapods: Hart (1971), Baez (1997), Puls (2001), Pessani, Tirelli, & Flagella (2004), Koettker, Sumida, Lopes, & Freire (2012); Fishes: Fahay (1983).
Due to the fact that each stage has a particular habitat and lifestyle, for some groups we counted every stage of the life cycle as a separate category (Beyst, Buysse, Dewicke, & Mees, 2001), e.g. distinguishing protozoea, zoea, megalopae and post-larvae of the decapods. This is necessary because the supraben-thos is composed by many larval stages more than permanent suprabenthos, but just particular ones are abundant. In order to understand the composition of these communities it is necessary to identify life-cycle stages.
Therefore we had two sets of data, one for groups and one for species/categories. After extracting the organisms, dragged material (e.g. wood debris, bryozoans, macroalgae detritus) contained in the sample was dried for 24h at 75°C. Then weighted in a 0.1g balance, in order to judge whether the amount of material influences the variations of the community.
Environmental variables: Were taken or measured simultaneously with the fauna, using the experimental design described previously. Dissolved oxygen (mg/L), oxygen saturation (%), pH, salinity and surface temperature (°C) were measured with a portable multiparameter probe for each site. Precipitation data was obtained from the Giovanni online data system, NASA GES DISC (http://gdata1.sci.gsfc.nasa.gov/daac-bin/G3/gui.cgi?instance_ id=TRMM_Monthly). Rainy months are those which have an accumulated monthly precipitation greater than 60mm, as established by the Venezuelan Air Force. For this study the months considered to be dry were January, February and March.
The total, organic and inorganic suspended solids were measured in three water samples taken at each site. These were filtered using Whatman GF/A filters (American Public Health Asociation [A.P.H.A.], 1995). Four sediment samples in each site were used to measure total organic carbon percentage (TOC) and organic matter percentage (OM). A single sample was taken at each site for sediment mean grain size (GS). For TOC measurements, the Walkley-Black method described in Anderson & Ingram (1989) was used. Organic matter percentage in sediment was obtained by drying the samples at 75°C for 48h (dry weight) and subsequently heated at 450°C for 1h (ash weight). The sediment grain size was determined using the procedure described by Roa (1983).
Abundances were standardized to individuals/m2. Taxa richness was determined for the suprabenthic assemblage using the DIVERSE analysis routines of the PRIMER V6 package (Clarke & Gorley, 2006). Group or species/ categories frequencies were expressed as a percentage of total presence in a given month and beach. Group and species/categories matrices were analyzed independently.
A Bray-Curtis similarity matrix was constructed from the abundance data. In order to reduce the effect of absence of individuals among samples (Clarke, Somerfield, & Chapman, 2006), dummy values of one were added. Four factor permutational multivariate analyses of variance (PERMANOVA) tested the null hypotheses of no difference in assemblages among beaches (random, 4 levels), sites (fixed, 2 levels, nested in beaches), seasons (fixed, 2 levels, orthogonal to beaches) and month (random, 12 levels, nested in seasons) (Anderson, 2001). Significance was set to p=0.05 and p-values were obtained using 9999 permutations of residuals under unrestricted permutation of raw data (Anderson, Gorley & Clarke, 2008). Non-metric multidimensional scaling (nMDS) was conducted on the similarity matrix in order to visualize the patterns in the spatial and temporal distributions of the suprabenthic assemblages (Clarke & Gorley, 2006). When significant differences were found, greatest taxa contributions were detected using similarity percentage (SIMPER) analyses (Clarke, 1993; Clarke & Warwick, 2001).
Environmental data was averaged to month level. PERMANOVA was used to evaluate the group differentiation by factors of beach, site, season, month and their interactions. For environmental data, the previously described design was also used. When significant interactions were detected, a principal component analysis (PCA) was conducted over the normalized data. This was performed in order to understand the spatial and temporal relationships of environmental variables. On the other hand, the environmental variables that best explain the biological distributions patterns were chosen using the BIO-ENV procedure. All variables were analyzed using month levels. Environmental data was averaged while biological data was grouped using centroids.
Results
Suprabenthic assemblage and variations: A total of 24 141 specimens (mean abundance: 26.16±55.35ind./m2) belonging to21 taxonomic groups were collected: foraminifera, radiolarians, cnidarians, ctenophores, bivalves, gastropods, opisthobranchs, poly-chaetes, pycnogonids, ostracods, cladocerans, copepods, amphipods, cumaceans, isopods, mysids, tanaids, decapods, ophiurids, chaeto-gnaths and fishes. A detailed taxonomic list is reported in table 1.
The most frequent groups in all sampling beaches and months were amphipods (97.87%) and decapods (95.74%). At the level of species/ categories, post-larval shrimp (Penaeidae) were the most frequent (91.49%), followed by Grap-sidae crab megalopae and Arenaeus cribarius (Lamarck, 1818) megalopae (82.98% each) (Table 1). Of all the species/categories identified, 31% appeared on one beach in a single month, illustrating how diverse and variable this assemblage can be.
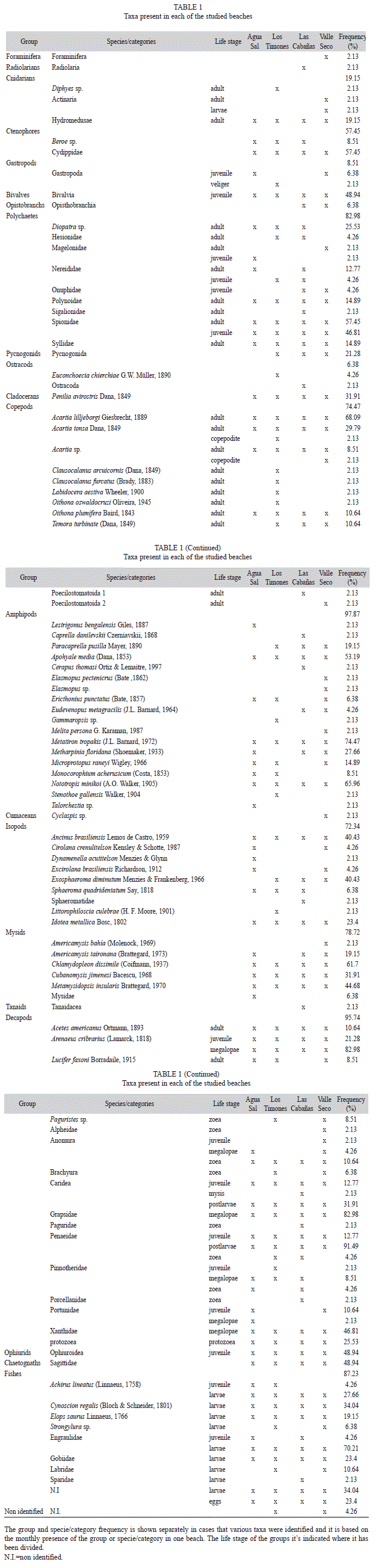
The maximum sample group richness was 14 at Los Timones during June (Fig. 1). Species richness ranged between 1 and 26 species/ categories by taxonomic group, being decapods the richest group (26 categories), followed by amphipods (18 species). The species/categories richness varied significantly between month-sites (PERMANOVA, p=0.0001) and month-beaches (PERMANOVA, p=0.0008) indicating temporal variation between beaches and sites.
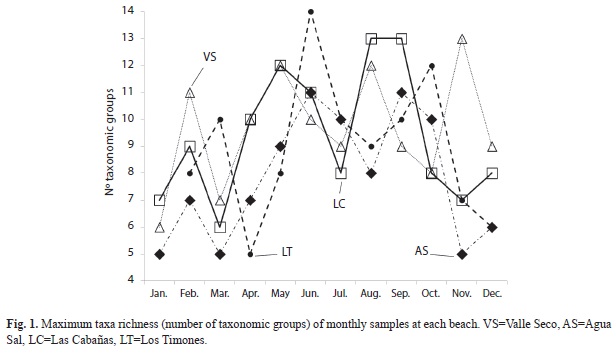
The total abundance of suprabenthos varied significantly between month-beaches (PERMANOVA, p=0.0001) and month-sites (PERMANOVA, p=0.0001). Large increments in abundances can be observed in January and November in Valle Seco, May for Los Timones and September for Las Cabañas. The highest abundances of all samples occurred in September in Las Cabañas. With mean value twice the maximum mean for the other months and beaches (Fig. 2). This high mean abundance was mainly attributed to an increase of Cydippidae ctenophores. As in the previous case, assemblages at the group and species/ categories levels, varied significantly between month-beaches (PERMANOVA, p=0.0001) and month-sites (PERMANOVA, p=0.0001). Thus assemblages can change monthly at sites and beaches.
During group data analysis, samples from each beach were not separated by nMDS. No clear patterns among beaches or months were observed at any level. SIMPER analysis was conducted for interactions between month-beaches at the group and species/categories level. The temporal assemblage’s differences for each beach exceeded 50% at both levels. The groups selected from the analysis only include those that contribute greater than 25% to dissimilarities between months for each beach. For Agua Sal and Valle Seco, the most contribution to beach dissimilarities was due to decapods, amphipods, ctenophores and mysids. On the other hand for Las Cabañas and Los Timones were amphipods, decapods, mysids, ctenophores and copepods.
The selected species/categories from the analysis include those that contribute 15% or more to dissimilarity between months for each beach. For Agua Sal beach, the species/categories that most contributed to the dissimilarity are: decapods (A. cribarius megalopae, Grap-sidae megalopae, Penaeidae post-larvae), cope-pods (Acartia lilljeborgi Giesbrecht, 1889), amphipods (Apohyale media (Dana, 1853)), polychaetes (Spionidae), and mysids (Chlamydopleon dissimile (Coifmann, 1937)). In the case of Valle Seco, the differences through the year were explained by the presence or abundance of amphipods (A. media, Nototropis minikoi (A. O. Walker, 1905), Metatiron tropakis (J. L. Barnard, 1972)), decapods (Penaeidae post-larvae, Grapsidae megalopae and proto-zoeas), isopods (Ancinus brasiliensis Lemos de Castro, 1959) and mysids (Cubanomysis jimenesi Bacescu, 1968). At Las Cabañas, decapods (A. cribarius megalopae, Penaeidae post-larvae), copepods (A. lilljeborgi, Acartia tonsa Dana, 1849), amphipods (N. minikoi, M. tropakis), chaetognath (Sagittidae) and the mysid (Metamysidopsis insularis Brattegard, 1970) contributed the most to temporal differences. Finally, in Los Timones the differences were due to the amphipods (N. minikoi, M. tropakis), a copepod (A. lilljeborgi), a decapod (A. cribarius megalopae), fish (Engraulidae larvae) and the mysid (C. dissimile).
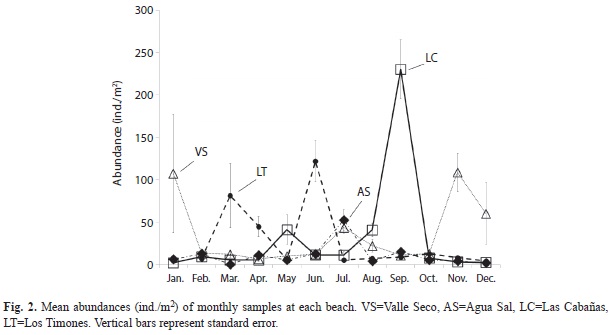
Environmental variables: Dissolved oxygen (DO) was similar on the four beaches, being very low in January with an increased in February and December. Values ranged from 1 to 7.67mg/L. The percentage of oxygen saturation ranged from 9% in January and 104% in July, remaining above 60% in most months for all beaches. The temperature ranged from 26.6 to 33.9°C with both extremes recorded from Valle Seco in April and August, respectively. The warmest months were between May and September. Salinity varied between 20 and 40ppm with February being the month with the highest and December the lowest. The pH tended to be alkaline throughout the year (7.2 to 9.95) with higher values in December. Average suspended solids remained close to zero during the year, but with higher values in July, October and November. Suspended organic matter varied between 4.16x10-7 and 6.28x10-4mg/L. Sediment medium grain size was 125µm for all beaches during the entire year. Sediments were greater than 60% of fine and very fine sands, with less than 1.4% total organic carbon and less than 10% of organic matter. An exception was observed in July at Los Timones, where organic matter content was as high as 24%. Likely due to the observed dredging activities in the creek that discharges onto this beach (Fig. 3).
Environmental variables for water and sediment have significant differences between month-beaches (PERMANOVA, p=0.0001). PCA analysis showed weak temporality in changes of the environmental variables, independently of the sampled beach. Sample groups did not have evident patterns. Samples from all beaches for October and those from Agua Sal, Valle Seco and Las Cabañas beaches for January were significantly different from the rest of the months, having low oxygen saturation percentage and DO concentration. Samples for Los Timones in July and September differed from the rest given the comparatively high percentage of OM and TOC.
Correlation with environmental variables: BIO-ENV routine revealed those environmental variables, either in isolation or in combination, that have the greatest influence on suprabenthic assemblage structure. At all levels, combinations of water characteristics best explained the changes in fauna assemblage.
When groups are used, the observed patterns in community assemblages were correlated with two or more environmental variables. On the contrary, the patterns in Agua Sal were explained only by salinity differences (Spearman, p=0.398). The fauna of Valle Seco was correlated with total suspended solids, suspended organic matter and suspended inorganic matter (Spearman, p=0.557). For Las Cabañas with organic matter percentage both in sediments and dragged material (Spearman, p=0.536). In respect to Los Timones, fauna was correlated to suspended inorganic matter, oxygen saturation percentage, total organic carbon percentage and dragged material (Spearman, p=0.457).
At species/categories level, low correlation was found between faunal changes and the combination of suspended organic matter and dragged material. The fauna of Valle Seco was correlated with suspended organic matter, suspended inorganic matter and dragged material (Spearman, p=0.430). That of Agua Sal with suspended organic matter and salinity (Spearman, p=0.270). For Las Cabañas beach fauna was correlated with organic matter percentage in sediment and dragged material (Spearman, p=0.403) and for Los Timones, with suspended organic matter, oxygen saturation percentage and dragged material (Spearman, p=0.224).
Discussion
The dominants groups of suprabenthos in the swash-zone can change in terms of abundance or frequency, depending on the studied area. Generally, peracarids are an important part of the community. In tropical waters of Brazil the suprabenthic fauna, in terms of frequency, was predominantly cumaceans, amphi-pods, ostracods, mysids and foraminifera; with copepods outranking all other groups on sandy substrates (Melo et al., 2010) while mysids were the most abundant group in Ecuador (Domínguez et al., 2004). In temperate waters, as there are more studies in diverse areas, abundance percentages of the main groups may be different, but it is common to find high abundances of amphipods, mysids, iso-pods and in a few cases cumaceans (Colman & Seagrove, 1955; Munilla & Corrales, 1995; Dauvin & Zouhiri, 1996; Munilla et al., 1998; Marín, 2007).
It is known that in terms of frequency for the tropical waters of Brazil, the suprabenthic fauna was predominantly cumaceans, amphi-pods, ostracods, mysids and foraminifera. Copepods outranked all other groups on sandy substrates (Melo et al., 2010). For beaches in Ecuador it is reported that mysids are the most abundant group (Domínguez et al., 2004). On the other hand, for temperate waters it is common to find high abundances of amphipods, mysids, isopods and in a few cases cumaceans (Colman & Seagrove, 1955; Munilla & Corrales, 1995; Dauvin & Zouhiri, 1996; Munilla et al., 1998; Marín, 2007).
For the littoral beaches of Venezuela, peracarids and decapods were the most frequent groups. For the decapods group, megalopae for crabs and post-larvae for shrimps were the most abundant stages in the benthic boundary layer. Results indicate that the benthic boundary layer probably plays a fundamental role for the development of these crustaceans. Extracted samples show presence of larval stages in the benthic boundary layer. This indicates that at least these larvaes form part of the faunal community of those beaches, but can migrate at latter stages.
Among peracarids, amphipods were the most frequent an abundant group. Particularly, A. media, is one of the most common in macroalgal assemblages (Tararam, Wakabara, & Mesquita, 1985). A. media is omnivorous, feeding by predation, scavenging, scraping and browsing (Tararam et al., 1985). Therefore, it is not surprising that individuals may live for some time in the benthic boundary layer, and even make use of the available substrate such as accumulated bryozoans (Valle Seco) and wood debris (Agua Sal). M. tropakis is a warm-temperate species reported from Virginia to Venezuela at depths between 3-157m on sandy bottoms (Dickinson, Wigley, Brodeur, & Brown-Leger, 1980). It is probably a common benthic species in this area which migrates to the benthic boundary layer to feed. N. minikoi is a cosmopolitan species reported in plankton samples and in sea weeds. Swarms of this species have been reported in shallow waters (Naomi, 1979). N. minikoi has also been reported as the preponderant species in a Brazilian estuary, and serves as an important food source for fishes (Wakabara, Nicoletti, & Tararam, 1996).
Within decapods, Penaeidae family includes almost all commercial shrimps, being mainly benthic species from littoral or deep zones (Rodriguez, 1980) . As they are frequently seen from shrimp artisanal fisheries in these beaches, it is probable that they have all their life cycle in those beaches, and suprab-enthos is required for the post larval development. Grapsidae crabs are common habitants of rocky substrates (Arteta-Bonivento, 2009); probably just their larvaes spend part of their lives in sampled beaches. A. cribarius is a decapod frequently found on sandy beaches, particularly abundant in estuarine and coastal beaches until 3m depth (Arteta- Bonivento, 2009). Probably this crab also has all its live cycle in these beaches, and megalopae stage inhabits in the suprabenthos.
For particular months in several beaches, this study found very high abundances of cteno-phores, which were often the dominant group for some samples (e.g. September in Agua Sal, September and October in Los Timones, and March, August and September in Las Cabañas). Those are not usually a representative group of the suprabenthos in other investigations. However, Wang, Thibeaut & Dauvin (1995) have studied the vertical migration of the ctenophore Pleurobrachia pileus (O. F. Müller, 1776) (Order Cydippida) and found that it normally inhabits in the suprabenthos zone.
Three species of mysids already reported for Venezuela (Sorbe, Martín, & Díaz, 2007) were also identified. These were C. dissimile, C. jimenesi and M. insularis which appeared in high abundance and frequency. An important contribution of this work is that we report for the first time in Venezuela the species Americamy-sis bahia (Molenock, 1969) and Americamy-sis taironana Brattegard, 1973. The species A. bahia has been previously reported in waters with salinities greater than 20ppm in the Gulf of Mexico (Price, 1982), while A. taironana has been found in Colombia and Brazil in the benthic boundary layer between 1 and 5m depth (Brattegard, 1973).
Suprabenthic assemblages in the studied area showed a high richness (21 taxonomic group) similar to other studies in tropical waters, such as in Brazil and Ecuador (Domínguez et al., 2004, Melo et al., 2010). However, it was higher than reported for temperate areas (Hamerlynck & Mees, 1991; Colman & Seagrove, 1955). The variations in richness of species/categories in most cases are unrelated to changes in the abundance of the organisms. But it is not comparable with other studies of suprabenthos since many groups are not identified to the lowest taxonomic levels.
Assemblages were found to be so variable and diverse that no evident pattern of distribution could be identified. Direct causality to temporal variation remains to be determined. Both abundance and community assemblage varied differently at temporal and spatial scales. No evident influence of the rainy and dry season could be detected. Mean suprabenthic abundances were low compared to those reported in tropical waters of Brazil (1 093±592ind./m2 sandy bottoms during day) (Melo et al., 2010) and Ecuador (60.73±35.90ind./m2) (Domínguez et al., 2004). Nevertheless, these were higher than those reported for European temperate waters, where abundances were less than 10ind./m2 (Munilla & Corrales, 1995; Cartes et al., 2007).
The swash zone in these beaches seems to be used extensively by the post-larvae of many species as reported by Beyst et al. (2001) in Belgium. In this sense, zoea, megalopae, decapod post-larvae and fish larvae were found throughout the year. All larva stages could be found around the year, indicating possible monthly or seasonal independence in their development. It has been shown that the swash zone provides a good environment and adequate food for larval and juvenile fish. Apparently, this environment is indispensable for the survival of animals that undergo metamorphosis in their life cycle (Beyst et al., 2001).
For this study, the rainy and dry seasons were not significant factors for the variability of environmental variables. This could be attributed to the fact that 2011 can be thought of as an atypical year in respect to rainfall. Only three months of drought were registered where the average is five months (Servicio de Hidrografia y Navegación de Venezuela).
Nevertheless, even the short drought season some temporary changes were observed in the environmental variables. Sediments of the four beaches consisted mainly of fine to very fine sand with an average grain size of 125μm. This is consistent with that reported by Herrera (2007) for this same area and matches the characteristics of a dissipative beach, such as very fine sand, a gentle slope and a well-developed surf zone (Short & Hesp, 1982; Wright & Short, 1984; Wright, Short, & Green, 1985).
The correlations between suprabenthic assemblages and the environmental variables (pH, salinity, temperature, solid, organic and inorganic suspended material, dissolved oxygen, and oxygen saturation percentage on water and organic matter, total organic carbon and grain size in sediment, dragged material) were weak. This is possibly due to the great environmental variability of the studied area. Overlap of factors may hamper detection of individually significant environmental factors. Coastal environments are not stable and can be affected by fluctuations associated with: discharges of organic matter, primary production of mac-rophytes in the surrounding areas, river discharges and vertical flows (Cartes et al., 2007; Fanelli, 2007). The bodies of water affecting the study area are primary the Tuy, Capaya and Curiepe rivers that have a significantly different influence per beach. The system is further complicated by the presence of a lagoon (La Reina), mangrove areas and Thalassia testudinum (Banks & Sol ex K. D. Koenig) beds.
In this sense, Agua Sal beach has an influence of the Tuy and Capaya rivers, while Las Cabañas is affected primarily by the Curiepe River. On the other hand, Valle Seco has a direct impact from the lagoon, the nearby T. testudinum beds and the mangrove areas. These ecosystems also influence Los Timones beach where samples showed accumulations of bryozoans that could only have proceeded from there.
It has been suggested that organisms considered good swimmers, including mysids, move away from the swash zone in order to avoid turbulence. This possibly can result in a decrease in density during severe weather conditions (Colman & Seagrove, 1955). On sampling days significant changes were observed in wave action caused by storms. This could explain temporal variations in the different groups throughout the study. No direct correlation between wave action and suprabenthos has been established. Although it is thought that it may have a significant impact on this community (Munilla et al., 1998; San Vicente & Sorbe, 1999).
Finally, Ligas et al. (2007) argued that seasonal patterns in the suprabenthos and suprabenthic crustacean fluctuations may be related to biological and ecological characteristics rather than environmental factors. Perhaps these biological factors may be the major determinants of the suprabenthic assemblages in Venezuela.
We concluded that the studied suprab-enthos is a very dynamic community. It can change significantly each month without a clear effect of any particular environmental variable. Suprabenthos is constituted mainly of peracarids and larval stages of other crustaceans. This is why we considered very important to continue the study of suprabenthos in the Caribbean Sea, for a better understanding of its composition, and the importance of this great community. More studies complementing suprabenthos samples with macrob-enthos and plankton are required for a better understanding of the biological interactions in coastal areas. Additionally, other environmental variables like wave action and tide levels should be measured.
Acknowledgments
The authors wish to thank all the participants of the field works, especially Williams Campos. To Juan José Cruz-Motta for his advices in data treatment. To Adriana Lopez, Joxmer Scott and Baumar Marin for their help in the identification of some taxa. To Andres Sajo for reviewing the manuscript and for editorial assistance. To reviewers by their comments which help to improve the manuscript.
References
American Public Health Association (A. P. H. A). (1995). Standard methods for the examination of water and wastewater. 19th edition. Washington, D.C., USA. [ Links ]
Anderson, M. J. (2001). Permutation tests for univariate or multivariate analysis of variance and regression. Canadian Journal of Fisheries and Aquatic Sciences, 58(3), 626-639. [ Links ]
Anderson, M. J., Gorley, R. N., & Clarke, K. R. (2008). PERMANOVA + for PRIMER: Guide to Software and Statistical Methods. PRIMER-E: Plymouth, UK. [ Links ]
Anderson, J. M. & Ingram J. S. I. (1989). Tropical soil biology and fertility: a handbook of methods. TSBF. CAB•International: Wallingford, UK. [ Links ]
Arteta-Bonivento, R. (2009). Cangrejos en el delta del río Ranchería, Riohacha (Colombia) (Crustacea: Deca-poda: Brachyura). Boletín Científico. Centro de Museos de Historia Natural, 13(1): 140-152. [ Links ]
Baez, P. (1997). Key to the families of decapod crustacean larvae collected off northern Chile during an El Niño event. Investigaciones Marinas, 25, 167-176. [ Links ]
Beyst, B., Buysse, D., Dewicke, A., & Mees, J. (2001). Surf zone hyperbenthos of Belgian Sandy Beaches: Seasonal Patterns. Estuarine, Coastal and Shelf Science, 53, 877-895. [ Links ]
Bone, D., Klein, E., Losada, F., Miloslavich, P., Penchasza-deh, P., Paredes, C., & Roa, V. (1998). Evaluación de impacto sobre la comunidad bentónica de la playa de Higuerote: Intensidad y valoración del daño. Informe preparado por INTECMAR para PDVSA. [ Links ]
Boltovskoy, D. (1981). Atlas del zooplancton del Atlántico Sudoccidental y métodos de trabajo con el zooplanc-ton marino. Mar del Plata, Argentina: Publicaciones Especiales INIDEP. [ Links ]
Brattegard, T. (1969). Marine biological Investigations in the Bahamas 10. Mysidacea from shallow water in the Bahamas and southern Florida. Part 1. Sarsia, 39, 17-106. [ Links ]
Brattegard, T. (1970a). Marine biological Investigations in the Bahamas 11. Mysidacea from shallow water in the Bahamas and southern Florida. Part 2. Sarsia, 41, 1-36. [ Links ]
Brattegard, T. (1970b). Mysidacea from shallow water in the Caribbean Sea. Sarsia, 43, 111-154. [ Links ]
Brattegard, T. (1973). Mysidacea from shallow water on the Caribbean Coast of Colombia. Sarsia, 54, 1-65. [ Links ]
Brattegard, T. (1974a). Additional Mysidacea from shallow water on the Caribbean Coast of Colombia. Sarsia, 57: 47-86. [ Links ]
Brattegard, T. (1974b). Mysidacea from shallow water on the Caribbean coast of Panama. Sarsia, 57, 87-108. [ Links ]
Brattegard, T. (1975). Shallow-water Mysidacea from the Lesser Antilles and other Caribbean regions. Studies on the Fauna of Curaçao and other Caribbean Islands, 157, 102-115. [ Links ]
Brunel, P., Besner, M., Messier, D., Poirier, L., Granger, D., & Weinstein, M. (1978). Le traîneau Macer-Giroq: appareil amélioré pour l’échantillonnage quantitatif étagé de la petite faune nageuse au voisinage du fond. Internationale Revue der Gesanten Hydrobiologie, 63, 815-829. [ Links ]
Campos, A. & Suárez, E. (1994). Copépodos pelágicos del Golfo de México y Mar Caribe I. Biología y Sistemática. México, D.F., México: Centro de Investigaciones de Quintana Roo. [ Links ]
Cartes, J. E. (1998). Dynamics of the bathyal Benthic Boundary Layer in the northwestern Mediterranean: depth and temporal variations in macrofaunal-mega-faunal communities and their possible connections within deep-sea trophic webs. Progress in Oceanography, 41, 111-139. [ Links ]
Cartes, J. E., Grémare, A., Maynou, F., Villora-Moreno, S., & Dinet, A. (2002). Bathymetric changes in the distributions of particulate organic matter and associated fauna along a deep-sea transect down the Catalan Sea slope (Northwestern Mediterranean). Progress in Oceanography, 53(1), 29-56. [ Links ]
Cartes, J. E., Ligas, A., De Biasi, A. M., Pacciardi, L., & Sartor, P. (2009). Small-spatial scale changes in productivity of suprabenthic and infaunal crustaceans at the continental shelf of Ebro Delta (western Mediterranean). Journal of Experimental Marine Biology and Ecology, 378(1-2), 40-49. [ Links ]
Cartes, J. E., Papiol, V., Palanques, A., Guillén, J., & Demestre, M. (2007). Dynamics of suprabentos off the Ebro Delta (Catalan Sea: western Mediterranean): Spatial and temporal patterns and relationships with environmental factors. Estuarine, Coastal and Shelf Sciences, 75, 501-515. [ Links ]
Cartes, J. E. & Sorbe, J. C. (1999). Estimating secondary production in bathyal suprabenthic peracarid crustaceans from the Catalan Sea slope (western Mediterranean; 391-1255m). Journal of Experimental Marine Biology and Ecology, 239(2), 195-210. [ Links ]
Clark, B. M., Bennett, B. A., & Lamberth, S. J. (1996). Factors affecting spatial variability in seine net catches of fish in the surf zone of False Bay, South Africa. Marine Ecology Progress Series, 131, 17-34. [ Links ]
Clarke, K. R. (1993). Non-parametric multivariate analyses of changes in community structure. Australian Journal of Ecology, 18(1), 117-143. [ Links ]
Clarke, K. R. & Gorley, R. N. (2006). PRIMER v6: User Manual/Tutorial. PRIMER-E: Plymouth, England. [ Links ]
Clarke, K. R. & Warrick, R. M. (2001). A further biodiversity index applicable to species list: variation in taxonomic distinctness. Marine Ecology Progress Series, 216, 265-278. [ Links ]
Clarke, K. R., Somerfield, P. J., & Chapman, M. G. (2006). On resemblance measures for ecological studies, including taxonomic dissimilarities and a zero-adjusted Bray-Curtis coefficient for denuded assemblages. Journal of Experimental Marine Biology and Ecology, 330, 55-80. [ Links ]
Colman, J. S. & Seagrove, F. (1955). The tidal plankton over Stoupe Beck Sands, Robin Hood’s Bay (Yorkshire, North Riding). Journal of Animal Ecology, 24, 445-462. [ Links ]
Dauvin, J. C. & Zouhiri, S. (1996). Suprabenthic crustacean fauna of a dense Ampelisca community from the English Channel. Journal of the Marine Biological Association of the United Kingdom, 76, 909-929. [ Links ]
Delgado-Blas, V. H. (2009). Spionidae Grube, 1850. In De J. A. León-González, R. Bastida-Zavala, L. F. Carrera-Parra, P. Hernández-Alcántara, S. I. Salazar-Vallejo, & V. Solís-Weiss (Comps). Poliquetos (Annelida: Polychaeta) de México: Generalidades y claves para identificación de species 3 (pp. 589-614). [ Links ]
Díaz, Y. J. (2001). Contribución al conocimiento del orden Amphipoda de la costa Centro-Occidental de Venezuela. (Master Thesis). Universidad Simón Bolívar, Caracas, Venezuela. [ Links ]
Dickinson, J. J., Wigley, L. R., Brodeur, R. D., & Brown-Leger, S. (1980). Distribution of Gammaridean Amphipoda (Crustacea) in the middle Atlantic Bight Region. NOAA Technical report NMFS SSRF-741: 1-46. [ Links ]
Domínguez, L., Fockedey, N., De Mey, M., Beyst, B., Cornejo, M., Calderon, J., & Vincx, M. (2004). Spatial patterns of the surf zone hyperbenthic fauna of Valdivia Bay (Ecuador). Hydrobiologia, 529, 205-224. [ Links ]
Fahay, M. (1983). Guide to the Early Stages of Marine Fishes Occurring in the Western North Atlantic Ocean, Cape Hatteras to the Southern Scotian Shelf. Journal of Northwest Atlantic Fishery Science, 4, 3-423. [ Links ]
Fanelli, E. (2007). Trophic relationships in demersal communities of western Mediterranean: case of studies from coastal and deep-sea ecosystems. (Ph.D. Thesis). Università degli Studi della Tuscia, Viterbo, Italia. [ Links ]
Fanelli, E., Cartes, J. E., Badalamenti, F., D’Anna, G., Pipitone, C., Azzurro, E., Rumolo, P., & Sprovieri, M. (2011). Mesoscale variability of coastal suprabenthic communities in the southern Tyrrhenian Sea (western Mediterranean). Estuarine, Coastal and Shelf Science, 91, 351-360. [ Links ]
Fanelli, E., Cartes, J. E., Badalamenti, F., Rumolo, P., & Sprovieri, M. (2009). Trophodynamics of suprabenthic fauna on coastal muddy bottoms of the southern Tyrrhenian Sea (western Mediterranean). Journal of Sea Research, 61, 174-187. [ Links ]
Fauchald, K. (1977) The polychaete worms. Definitions and keys to the orders, families, and genera. Natural History Museum of Los Angeles County in conjunction with The Allan Hancock Foundation University of Southern California, Science Series 28. [ Links ]
Hamerlynck, O. & Mees, J. (1991). Temporal and spatial structure in the hyperbenthic community of a shallow coastal area and its relation to environmental variables. Oceanologica Acta, 11, 205-212. [ Links ]
Harris, R., Wiebe, P., Lenz, J., Skjoldal, H. R., & Huntley, M. (2000). ICES Zooplankton Methodology Manual. Elsevier. [ Links ]
Hart, J. F. L. (1971). Key to planktonic larvae of families of decapod Crustacea of British Columbia. Syesis, 4, 227-234. [ Links ]
Heard, R. W., Price, W. W., Knott, D. M., King, R. A., & Allen, D. M. (2006). A taxonomic guide to the Mysids of the south Atlanctic Bight. NOAA Professional Paper NMFS, 4, 1-37. [ Links ]
Heard, R. W., Roccagliata, D., & Petrescu, I. (2007). An illustrated guide to Cumacea (Crustacea: Malacos-traca: Peracarida) from Florida coastal and shelf waters to depth of 100m. Tallahassee, Florida, U.S.A: Department of Environmental Protection, State of Florida. [ Links ]
Herrera, A. (2007). Influencia de los ríos sobre ecosistemas marino-costeros: ejemplos sobre comunidades de playas arenosas. (Ph.D. Thesis). Universidad Simón Bolívar, Caracas, Venezuela. [ Links ]
Herrera, A. & Bone, D. (2011). Influence of riverine outputs on sandy beaches of Higuerote, central coast of Venezuela. Latin American Journal of Aquatic Research, 39(1), 56-70. [ Links ]
Kensley, B. & Schotte, M. (1989). Guide to the marine Iso-pod Crustaceans of the Caribbean. Washington D.C. and London: Smithsonian Institution Press. [ Links ]
Koettker, A., Sumida, P. Y. G., Lopes, R. M., & Freire, A. S. (2012). Illustrated key for the identification of the known zoeal stages of brachyuran crabs (Crustacea: Decapoda) from tropical and subtropical Brazil, southwestern Atlantic. Zootaxa, 3204, 1-19. [ Links ]
Ligas, A., Sartor, P., Sartini, M., Vannucci, A., De Ranieri, S., & Cartes, J. E. (2007). Hyperbenthic Mysidae Haworth, 1825 (Peracarida, Mysida) from the continental shelf of the Northern Adriatic Sea. Crustaceana, 80(8), 969-977. [ Links ]
Madurell, T. & Cartes, J. E. (2003). The suprabenthic peracarid fauna collected at bathyal depths in the Ionian Sea (eastern Mediterranean). Crustaceana, 76(5), 611-624. [ Links ]
Marín, J. (2007). The ecology of surf zone fauna of dissipative sandy beaches in southern Oregon U. S. A. (Master Thesis). University of Oregon, Oregon, U.S.A. [ Links ]
Mees, J. & Jones, M. B. (1997). The hyperbenthos. Oceanography and Marine Biology: an Annual Review, 35, 221-255. [ Links ]
Melo, P., Silva, T. A., Neumann-Leitãão, S., Schwam-born, R., Gusmãão, L. M. O., & Neto, F. P. (2010). Demersal zooplankton communities from tropical habitats in the southwestern Atlantic. Marine Biology Research, 6, 530-541. [ Links ]
Munilla, T. & Corrales, M. (1995). Suprabentos de la playa de Rosas (Gerona, Mediterráneo occidental). Orsis, 10, 83-90. [ Links ]
Munilla, T., Corrales, M., & San Vicente, C. (1998). Supra-benthic assemblages from Catalan beaches: zoological groups. Orsis, 13, 67-7. [ Links ]
Munilla, T. & San Vicente, C. (2005). Suprabenthic biodiversity of Catalan beaches (NW Mediterranean). Acta Oecologica, 27, 81-91. [ Links ]
Naomi, T. S. (1979). On a swarm of amphipods Atylus minikoi (Walker) in the shallow waters of the Karwar bay. Indian Journal of Fisheries, 26(1&2): 227-228. [ Links ]
Ortiz, M., Martín, A., Winfield, I., Díaz, Y. J., & Atienza, D. (2004). Anfípodos (Crustacea: Gammaridea): Clave gráfica para la identificación de las familias, los géneros y las especies de los anfípodos marinos y estuarinos del Atlántico Occidental tropical. México: Editorial Universidad Nacional Autónoma de México. [ Links ]
Pessani, D., Tirelli, T., & Flagella, S. (2004). Key for the identification of Mediterranean brachyuran megalopae. Mediterranean Marine Science, 5(2), 53-64. [ Links ]
Price, W. W. (1982). Key to the shallow water Mysidacea of the Texas coast notes on their ecology. Hydrobiologia, 93, 9-21. [ Links ]
Puls, A. L. (2001). Arthropoda: Decapoda. In A. Shanks (Ed.), An identification guide to the larval marine invertebrates of the Pacific Northwest (pp. 179-250). Oregon, U.S.A.: Oregon State University Press. Corvallis. [ Links ]
Roa, V. (1983). Estudio comparativo de los sedimentos marinos del área de Punta Morón, Planta Centro, Estado Carabobo, Venezuela. In P. Penchaszadeh. (Ed.), Ecología del ambiente Marino Costero de Punta Morón (pp. 41-55). Caracas, Venezuela: INTECMAR-USB. [ Links ]
Rodriguez, G. (1980). Crustáceos decápodos de Venezuela. Caracas, Venezuela: Instituto Venezolano de Investigaciones Científicas. [ Links ]
San Vicente, C. & Sorbe, J. C. (1999). Spatio-temporal structure of the suprabenthic community from Creixell beach (Western Mediterranean). Acta Oecologica, 20(4), 377-389. [ Links ]
Short, A. & Hesp, P. (1982). Wave, beach and dune interactions in southeastern Australia. Marine Geology, 48(3-4), 259-284. [ Links ]
Sorbe, J. C., Martín, A., & Díaz, Y. J. (2007). New records of Mysida (Crustacea:Peracarida) from shallow waters of the Caribbean coast of Venezuela. Marine Biology Research, 3(3), 175-181. [ Links ]
Tararam, A. S., Wakabara, Y., & Mesquita, H. S. L. (1985). Feeding habits of Hyale media (Dana, 1853) (Crustacea-Amphipoda). Boletin Instituto Oceanografico da universidade de Sao Paulo, 33(2): 193-199. [ Links ]
Uebelacker, J. M. & Johnson, P. G. (Eds.). (1984). Taxonomic Guide to the Polychaetes of the Northern Gulf of México. Final Report to the Minerals Management Service, contract 14-12-001-29091. Mobile, Alabama: Barry A. Vittor & Associates, Inc. [ Links ]
Wakabara, Y., Nicoletti, M., & Tararam, A. S. (1996). Ingestion and selection of suprabenthic crustaceans by small-sized fishes in a lower saltmarsh system. Revista Brasileira de Oceanografia, 44(2), 89-103. [ Links ]
Wang, Z., Thibeaut, E., & Dauvin, J. C. (1995). Spring abundance and distribution of the ctenophore Pleurobrachia pileus in the Seine estuary: advective transport and diel vertical migration. Marine Biology, 124(2), 313-324. [ Links ]
Wright, L. D. & Short, A. D. (1984). Morphodynamic variability of surf zones and beaches: a synthesis. Marine Geology, 56(1-4), 93-118. [ Links ]
Wright, L. D., Short, A. D., & Green, M. O. (1985). Short-term changes in the morphodynamic states of beaches and surf zones: an empirical predictive model. Marine Geology, 62(3-4), 339-364. [ Links ]
Anderson, M. J. (2001). Permutation tests for univariate or multivariate analysis of variance and regression. Canadian Journal of Fisheries and Aquatic Sciences, 58(3), 626-639. [ Links ]
Anderson, M. J., Gorley, R. N., & Clarke, K. R. (2008). PERMANOVA + for PRIMER: Guide to Software and Statistical Methods. PRIMER-E: Plymouth, UK. [ Links ]
Anderson, J. M. & Ingram J. S. I. (1989). Tropical soil biology and fertility: a handbook of methods. TSBF. CAB•International: Wallingford, UK. [ Links ]
Arteta-Bonivento, R. (2009). Cangrejos en el delta del río Ranchería, Riohacha (Colombia) (Crustacea: Deca-poda: Brachyura). Boletín Científico. Centro de Museos de Historia Natural, 13(1): 140-152. [ Links ]
Baez, P. (1997). Key to the families of decapod crustacean larvae collected off northern Chile during an El Niño event. Investigaciones Marinas, 25, 167-176. [ Links ]
Beyst, B., Buysse, D., Dewicke, A., & Mees, J. (2001). Surf zone hyperbenthos of Belgian Sandy Beaches: Seasonal Patterns. Estuarine, Coastal and Shelf Science, 53, 877-895. [ Links ]
Bone, D., Klein, E., Losada, F., Miloslavich, P., Penchasza-deh, P., Paredes, C., & Roa, V. (1998). Evaluación de impacto sobre la comunidad bentónica de la playa de Higuerote: Intensidad y valoración del daño. Informe preparado por INTECMAR para PDVSA. [ Links ]
Boltovskoy, D. (1981). Atlas del zooplancton del Atlántico Sudoccidental y métodos de trabajo con el zooplanc-ton marino. Mar del Plata, Argentina: Publicaciones Especiales INIDEP. [ Links ]
Brattegard, T. (1969). Marine biological Investigations in the Bahamas 10. Mysidacea from shallow water in the Bahamas and southern Florida. Part 1. Sarsia, 39, 17-106. [ Links ]
Brattegard, T. (1970a). Marine biological Investigations in the Bahamas 11. Mysidacea from shallow water in the Bahamas and southern Florida. Part 2. Sarsia, 41, 1-36. [ Links ]
Brattegard, T. (1970b). Mysidacea from shallow water in the Caribbean Sea. Sarsia, 43, 111-154. [ Links ]
Brattegard, T. (1973). Mysidacea from shallow water on the Caribbean Coast of Colombia. Sarsia, 54, 1-65. [ Links ]
Brattegard, T. (1974a). Additional Mysidacea from shallow water on the Caribbean Coast of Colombia. Sarsia, 57: 47-86. [ Links ]
Brattegard, T. (1974b). Mysidacea from shallow water on the Caribbean coast of Panama. Sarsia, 57, 87-108. [ Links ]
Brattegard, T. (1975). Shallow-water Mysidacea from the Lesser Antilles and other Caribbean regions. Studies on the Fauna of Curaçao and other Caribbean Islands, 157, 102-115. [ Links ]
Brunel, P., Besner, M., Messier, D., Poirier, L., Granger, D., & Weinstein, M. (1978). Le traîneau Macer-Giroq: appareil amélioré pour l’échantillonnage quantitatif étagé de la petite faune nageuse au voisinage du fond. Internationale Revue der Gesanten Hydrobiologie, 63, 815-829. [ Links ]
Campos, A. & Suárez, E. (1994). Copépodos pelágicos del Golfo de México y Mar Caribe I. Biología y Sistemática. México, D.F., México: Centro de Investigaciones de Quintana Roo. [ Links ]
Cartes, J. E. (1998). Dynamics of the bathyal Benthic Boundary Layer in the northwestern Mediterranean: depth and temporal variations in macrofaunal-mega-faunal communities and their possible connections within deep-sea trophic webs. Progress in Oceanography, 41, 111-139. [ Links ]
Cartes, J. E., Grémare, A., Maynou, F., Villora-Moreno, S., & Dinet, A. (2002). Bathymetric changes in the distributions of particulate organic matter and associated fauna along a deep-sea transect down the Catalan Sea slope (Northwestern Mediterranean). Progress in Oceanography, 53(1), 29-56. [ Links ]
Cartes, J. E., Ligas, A., De Biasi, A. M., Pacciardi, L., & Sartor, P. (2009). Small-spatial scale changes in productivity of suprabenthic and infaunal crustaceans at the continental shelf of Ebro Delta (western Mediterranean). Journal of Experimental Marine Biology and Ecology, 378(1-2), 40-49. [ Links ]
Cartes, J. E., Papiol, V., Palanques, A., Guillén, J., & Demestre, M. (2007). Dynamics of suprabentos off the Ebro Delta (Catalan Sea: western Mediterranean): Spatial and temporal patterns and relationships with environmental factors. Estuarine, Coastal and Shelf Sciences, 75, 501-515. [ Links ]
Cartes, J. E. & Sorbe, J. C. (1999). Estimating secondary production in bathyal suprabenthic peracarid crustaceans from the Catalan Sea slope (western Mediterranean; 391-1255m). Journal of Experimental Marine Biology and Ecology, 239(2), 195-210. [ Links ]
Clark, B. M., Bennett, B. A., & Lamberth, S. J. (1996). Factors affecting spatial variability in seine net catches of fish in the surf zone of False Bay, South Africa. Marine Ecology Progress Series, 131, 17-34. [ Links ]
Clarke, K. R. (1993). Non-parametric multivariate analyses of changes in community structure. Australian Journal of Ecology, 18(1), 117-143. [ Links ]
Clarke, K. R. & Gorley, R. N. (2006). PRIMER v6: User Manual/Tutorial. PRIMER-E: Plymouth, England. [ Links ]
Clarke, K. R. & Warrick, R. M. (2001). A further biodiversity index applicable to species list: variation in taxonomic distinctness. Marine Ecology Progress Series, 216, 265-278. [ Links ]
Clarke, K. R., Somerfield, P. J., & Chapman, M. G. (2006). On resemblance measures for ecological studies, including taxonomic dissimilarities and a zero-adjusted Bray-Curtis coefficient for denuded assemblages. Journal of Experimental Marine Biology and Ecology, 330, 55-80. [ Links ]
Colman, J. S. & Seagrove, F. (1955). The tidal plankton over Stoupe Beck Sands, Robin Hood’s Bay (Yorkshire, North Riding). Journal of Animal Ecology, 24, 445-462. [ Links ]
Dauvin, J. C. & Zouhiri, S. (1996). Suprabenthic crustacean fauna of a dense Ampelisca community from the English Channel. Journal of the Marine Biological Association of the United Kingdom, 76, 909-929. [ Links ]
Delgado-Blas, V. H. (2009). Spionidae Grube, 1850. In De J. A. León-González, R. Bastida-Zavala, L. F. Carrera-Parra, P. Hernández-Alcántara, S. I. Salazar-Vallejo, & V. Solís-Weiss (Comps). Poliquetos (Annelida: Polychaeta) de México: Generalidades y claves para identificación de species 3 (pp. 589-614). [ Links ]
Díaz, Y. J. (2001). Contribución al conocimiento del orden Amphipoda de la costa Centro-Occidental de Venezuela. (Master Thesis). Universidad Simón Bolívar, Caracas, Venezuela. [ Links ]
Dickinson, J. J., Wigley, L. R., Brodeur, R. D., & Brown-Leger, S. (1980). Distribution of Gammaridean Amphipoda (Crustacea) in the middle Atlantic Bight Region. NOAA Technical report NMFS SSRF-741: 1-46. [ Links ]
Domínguez, L., Fockedey, N., De Mey, M., Beyst, B., Cornejo, M., Calderon, J., & Vincx, M. (2004). Spatial patterns of the surf zone hyperbenthic fauna of Valdivia Bay (Ecuador). Hydrobiologia, 529, 205-224. [ Links ]
Fahay, M. (1983). Guide to the Early Stages of Marine Fishes Occurring in the Western North Atlantic Ocean, Cape Hatteras to the Southern Scotian Shelf. Journal of Northwest Atlantic Fishery Science, 4, 3-423. [ Links ]
Fanelli, E. (2007). Trophic relationships in demersal communities of western Mediterranean: case of studies from coastal and deep-sea ecosystems. (Ph.D. Thesis). Università degli Studi della Tuscia, Viterbo, Italia. [ Links ]
Fanelli, E., Cartes, J. E., Badalamenti, F., D’Anna, G., Pipitone, C., Azzurro, E., Rumolo, P., & Sprovieri, M. (2011). Mesoscale variability of coastal suprabenthic communities in the southern Tyrrhenian Sea (western Mediterranean). Estuarine, Coastal and Shelf Science, 91, 351-360. [ Links ]
Fanelli, E., Cartes, J. E., Badalamenti, F., Rumolo, P., & Sprovieri, M. (2009). Trophodynamics of suprabenthic fauna on coastal muddy bottoms of the southern Tyrrhenian Sea (western Mediterranean). Journal of Sea Research, 61, 174-187. [ Links ]
Fauchald, K. (1977) The polychaete worms. Definitions and keys to the orders, families, and genera. Natural History Museum of Los Angeles County in conjunction with The Allan Hancock Foundation University of Southern California, Science Series 28. [ Links ]
Hamerlynck, O. & Mees, J. (1991). Temporal and spatial structure in the hyperbenthic community of a shallow coastal area and its relation to environmental variables. Oceanologica Acta, 11, 205-212. [ Links ]
Harris, R., Wiebe, P., Lenz, J., Skjoldal, H. R., & Huntley, M. (2000). ICES Zooplankton Methodology Manual. Elsevier. [ Links ]
Hart, J. F. L. (1971). Key to planktonic larvae of families of decapod Crustacea of British Columbia. Syesis, 4, 227-234. [ Links ]
Heard, R. W., Price, W. W., Knott, D. M., King, R. A., & Allen, D. M. (2006). A taxonomic guide to the Mysids of the south Atlanctic Bight. NOAA Professional Paper NMFS, 4, 1-37. [ Links ]
Heard, R. W., Roccagliata, D., & Petrescu, I. (2007). An illustrated guide to Cumacea (Crustacea: Malacos-traca: Peracarida) from Florida coastal and shelf waters to depth of 100m. Tallahassee, Florida, U.S.A: Department of Environmental Protection, State of Florida. [ Links ]
Herrera, A. (2007). Influencia de los ríos sobre ecosistemas marino-costeros: ejemplos sobre comunidades de playas arenosas. (Ph.D. Thesis). Universidad Simón Bolívar, Caracas, Venezuela. [ Links ]
Herrera, A. & Bone, D. (2011). Influence of riverine outputs on sandy beaches of Higuerote, central coast of Venezuela. Latin American Journal of Aquatic Research, 39(1), 56-70. [ Links ]
Kensley, B. & Schotte, M. (1989). Guide to the marine Iso-pod Crustaceans of the Caribbean. Washington D.C. and London: Smithsonian Institution Press. [ Links ]
Koettker, A., Sumida, P. Y. G., Lopes, R. M., & Freire, A. S. (2012). Illustrated key for the identification of the known zoeal stages of brachyuran crabs (Crustacea: Decapoda) from tropical and subtropical Brazil, southwestern Atlantic. Zootaxa, 3204, 1-19. [ Links ]
Ligas, A., Sartor, P., Sartini, M., Vannucci, A., De Ranieri, S., & Cartes, J. E. (2007). Hyperbenthic Mysidae Haworth, 1825 (Peracarida, Mysida) from the continental shelf of the Northern Adriatic Sea. Crustaceana, 80(8), 969-977. [ Links ]
Madurell, T. & Cartes, J. E. (2003). The suprabenthic peracarid fauna collected at bathyal depths in the Ionian Sea (eastern Mediterranean). Crustaceana, 76(5), 611-624. [ Links ]
Marín, J. (2007). The ecology of surf zone fauna of dissipative sandy beaches in southern Oregon U. S. A. (Master Thesis). University of Oregon, Oregon, U.S.A. [ Links ]
Mees, J. & Jones, M. B. (1997). The hyperbenthos. Oceanography and Marine Biology: an Annual Review, 35, 221-255. [ Links ]
Melo, P., Silva, T. A., Neumann-Leitãão, S., Schwam-born, R., Gusmãão, L. M. O., & Neto, F. P. (2010). Demersal zooplankton communities from tropical habitats in the southwestern Atlantic. Marine Biology Research, 6, 530-541. [ Links ]
Munilla, T. & Corrales, M. (1995). Suprabentos de la playa de Rosas (Gerona, Mediterráneo occidental). Orsis, 10, 83-90. [ Links ]
Munilla, T., Corrales, M., & San Vicente, C. (1998). Supra-benthic assemblages from Catalan beaches: zoological groups. Orsis, 13, 67-7. [ Links ]
Munilla, T. & San Vicente, C. (2005). Suprabenthic biodiversity of Catalan beaches (NW Mediterranean). Acta Oecologica, 27, 81-91. [ Links ]
Naomi, T. S. (1979). On a swarm of amphipods Atylus minikoi (Walker) in the shallow waters of the Karwar bay. Indian Journal of Fisheries, 26(1&2): 227-228. [ Links ]
Ortiz, M., Martín, A., Winfield, I., Díaz, Y. J., & Atienza, D. (2004). Anfípodos (Crustacea: Gammaridea): Clave gráfica para la identificación de las familias, los géneros y las especies de los anfípodos marinos y estuarinos del Atlántico Occidental tropical. México: Editorial Universidad Nacional Autónoma de México. [ Links ]
Pessani, D., Tirelli, T., & Flagella, S. (2004). Key for the identification of Mediterranean brachyuran megalopae. Mediterranean Marine Science, 5(2), 53-64. [ Links ]
Price, W. W. (1982). Key to the shallow water Mysidacea of the Texas coast notes on their ecology. Hydrobiologia, 93, 9-21. [ Links ]
Puls, A. L. (2001). Arthropoda: Decapoda. In A. Shanks (Ed.), An identification guide to the larval marine invertebrates of the Pacific Northwest (pp. 179-250). Oregon, U.S.A.: Oregon State University Press. Corvallis. [ Links ]
Roa, V. (1983). Estudio comparativo de los sedimentos marinos del área de Punta Morón, Planta Centro, Estado Carabobo, Venezuela. In P. Penchaszadeh. (Ed.), Ecología del ambiente Marino Costero de Punta Morón (pp. 41-55). Caracas, Venezuela: INTECMAR-USB. [ Links ]
Rodriguez, G. (1980). Crustáceos decápodos de Venezuela. Caracas, Venezuela: Instituto Venezolano de Investigaciones Científicas. [ Links ]
San Vicente, C. & Sorbe, J. C. (1999). Spatio-temporal structure of the suprabenthic community from Creixell beach (Western Mediterranean). Acta Oecologica, 20(4), 377-389. [ Links ]
Short, A. & Hesp, P. (1982). Wave, beach and dune interactions in southeastern Australia. Marine Geology, 48(3-4), 259-284. [ Links ]
Sorbe, J. C., Martín, A., & Díaz, Y. J. (2007). New records of Mysida (Crustacea:Peracarida) from shallow waters of the Caribbean coast of Venezuela. Marine Biology Research, 3(3), 175-181. [ Links ]
Tararam, A. S., Wakabara, Y., & Mesquita, H. S. L. (1985). Feeding habits of Hyale media (Dana, 1853) (Crustacea-Amphipoda). Boletin Instituto Oceanografico da universidade de Sao Paulo, 33(2): 193-199. [ Links ]
Uebelacker, J. M. & Johnson, P. G. (Eds.). (1984). Taxonomic Guide to the Polychaetes of the Northern Gulf of México. Final Report to the Minerals Management Service, contract 14-12-001-29091. Mobile, Alabama: Barry A. Vittor & Associates, Inc. [ Links ]
Wakabara, Y., Nicoletti, M., & Tararam, A. S. (1996). Ingestion and selection of suprabenthic crustaceans by small-sized fishes in a lower saltmarsh system. Revista Brasileira de Oceanografia, 44(2), 89-103. [ Links ]
Wang, Z., Thibeaut, E., & Dauvin, J. C. (1995). Spring abundance and distribution of the ctenophore Pleurobrachia pileus in the Seine estuary: advective transport and diel vertical migration. Marine Biology, 124(2), 313-324. [ Links ]
Wright, L. D. & Short, A. D. (1984). Morphodynamic variability of surf zones and beaches: a synthesis. Marine Geology, 56(1-4), 93-118. [ Links ]
Wright, L. D., Short, A. D., & Green, M. O. (1985). Short-term changes in the morphodynamic states of beaches and surf zones: an empirical predictive model. Marine Geology, 62(3-4), 339-364. [ Links ]
*Correspondencia a:
Ileana Ortega. Universidad Simón Bolívar. Laboratorio de Crustáceos Peracáridos. Apdo. 89000. Caracas 1086-A, Venezuela. Correo electrónico ileanaortega@gmail.com
Alberto Martín. Universidad Simón Bolívar. Centro de Biodiversidad Marina. Departamento de Estudios Ambientales. Laboratorio de Crustáceos Peracáridos. Apdo. 89000. Caracas 1086-A, Venezuela. Correo electrónico amartinz@usb.ve
Yusbelly J. Díaz. Universidad Simón Bolívar. Centro de Biodiversidad Marina. Departamento de Estudios Ambientales. Laboratorio de Crustáceos Peracáridos. Apdo. 89000. Caracas 1086-A, Venezuela. Correo electrónico yusdiaz@usb.ve
1. Universidad Simón Bolívar. Laboratorio de Crustáceos Peracáridos. Apdo. 89000. Caracas 1086-A, Venezuela. Correo electrónico ileanaortega@gmail.com
2. Universidad Simón Bolívar. Centro de Biodiversidad Marina. Departamento de Estudios Ambientales. Laboratorio de Crustáceos Peracáridos. Apdo. 89000. Caracas 1086-A, Venezuela. Correo electrónico amartinz@usb.ve, yusdiaz@usb.ve
Received 22-v-2013. Corrected 10-Xi-2013. Accepted 11-Xii-2013.


{kind=link}
{kind=link}
{kind=link}
{kind=link}