










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.62 n.2 San José Apr./Jun. 2014
Phytoplankton variability in Lake Fraijanes, Costa Rica, in response to local weather variation
Variabilidad del fitoplancton en el lago Fraijanes, Costa Rica, en respuesta a la variación local del clima
Variabilidad del fitoplancton en el lago Fraijanes, Costa Rica, en respuesta a la variación local del clima
*Dirección para correspondencia:
Abstract
Phytoplankton species show a variety in morphology which is the result of adaptations to pelagic life including responses to fluctuations in water column dynamics driven by weather conditions. This has been reported in the oceans and in Northern temperate lakes. In order to observe whether tropical freshwater phytoplankton responds to seasonal variation in weather, the weekly variation in temperature of the water column and phytoplankton composition was studied in Lake Fraijanes, Costa Rica, a shallow (6.2m) lake at 1 640m above sea level. A chain of data loggers for temperature was placed in the deepest point in the lake to register temperature every hour at four different depths, and phytoplankton samples were retrieved every week for a year. Additional monthly samples for nutrients were taken at two depths. Notwithstanding its shallowness, the lake developed a thermal gradient which kept the water column stratified for several months during dry season. Whole lake overturns occurred during cold spells with intense precipitation. Phytoplankton changed throughout the year mainly through a shift in dominant taxa. From September to February the lake was frequently mixed by rain storms and windy weather. At this time, phytoplankton was dominated by Chlorococcal green algae. From March to June, the lake was stratified and warmer. Phytoplankton became dominated by Cyanobateria, mainly colonial Chroococcales. The rainy season started again in May 2009. During June and July the lake started to mix intermittently during rain events and phytoplankton showed a brief increase in the contribution of Chlorococcales. These changes fitted well to a general model of phytoplankton succession based on functional groups identified according to their morphology and adaptations.
Key words: tropical limnology, lakes, phytoplankton, weather variation, stratification, ecology.
Resumen
Las especies del fitoplancton muestran una variada morfología que es el resultado de su adaptación a la vida pelágica la cual incluye respuestas a las fluctuaciones en la dinámica de la columna de agua inducida por las condiciones del tiempo atmosférico. Esto ha sido reportado para el océano y lagos de zonas templadas. Con el fin de observar si el fitoplancton de lagos tropicales responde de la misma manera a los cambios estacionales se estudió la variación semanal de la temperatura del agua y la composición del fitoplancton en el lago Fraijanes, Costa Rica, un lago somero (6.2m de profundidad) localizado a 1 640m sobre el nivel del mar. Se colocó una cadena de registradores automáticos de temperatura en el punto más profundo del lago programados para tomar una lectura cada hora a cuatro profundidades y se tomaron muestras superficiales del fitoplancton cada semana por un periodo de un año. Adicionalmente se tomaron muestras mensuales para la determinación de nutrientes a dos profundidades. A pesar de su poca profundidad, el lago desarrolló un gradiente térmico que mantuvo la columna de agua estratificada por varios meses durante la estación seca. Los eventos de mezcla total ocurrieron durante episodios fríos y de lluvia intensa. El fitoplancton cambió a través del año principalmente por cambios en los táxones dominantes. De septiembre a febrero el lago se mezcló frecuentemente por la lluvia y el viento. En este periodo el fitoplancton estuvo dominado por algas verdes Chloroccocales. De marzo a junio el lago estuvo estratificado y cálido. El fitoplancton pasó a ser dominado por Cyanobacteria principalmente Chroococcales. La estación lluviosa inició de nuevo en mayo 2009. Durante junio y julio el lago se mezcló intermitentemente y el fitoplancton mostró breves episodios en los cuales las algas verdes aumentaron de nuevo. Estos cambios se ajustan bien al modelo general de sucesión del fitoplancton basado en grupos funcionales definidos de acuerdo a su morfología y adaptaciones.
Palabras clave: limnología tropical, lagos, fitoplancton, variaciones en el tiempo local, estratificación, ecología.
Lakes are open systems that are affected by the influence of external forces such as wind, solar radiation and rains, the input of substances from the catchment in the surface runoff or subterranean seepage and the move-ment of organisms in and out of the lake. In response to these factors, the lake is modified from the circulation and turnover of its waters to changes in the productivity and identity of the organisms that inhabit its waters. In accordance, several models have been devised to explain the behavior of lakes and their response to changes in environmental factors, such as the excess input of nutrients (e.g. Vollenweider, 1968; Schindler et al., 1973), acid rain (e.g. Henrikson, Hindlar & Abrahamsson, 2005; Schindler, 1988), exotic species (eg. Mills, Leach, Carlton & Secor, 1994), among others. They are intended to explain patterns in the dynamics of phytoplankton in terms of physical (light, mixing) (e.g. Harris, 1978; Reynolds, 1984), chemistry (nutrients) (e.g. Tilman, Kilham & Kilham, 1982) and biological (herbivory) (e.g. Sterner, 1989) factors, as well as the functional morphology of individual species (Margalef, 1978; Reynolds 1984). However these models are primarily based on studies made in temperate zone lakes (Reynolds, 1988; Sommer 1989; Sommer, Gliwicz, Lampert & Duncan, 1986) and few of them have been tested in tropical lakes. Lewis (1978, 1986) performed long-term studies in some tropical lakes such as Lake Lanao (Phillipines) and Lake Valencia (Venezuela); proposed that it is possible to observe the same sequence of temporal changes in phytoplankton composition in tropical lakes as in temperate lakes following a deep mixing event, but that these changes occur in a shorter time period in the former, which can be reverted or stopped if weather conditions change.
The study of the phytoplankton is basic to understand the dynamics of the lacustrine ecosystem since it is the fundament of the food chain, which is utilized not only by herbivorous zooplankton but also by some fish species (Schiemer, 1996). It also furnishes the organic matter that is used by bacteria, which also serve as food for the same zooplankton (Kalff, 2002; Lampert & Sommer, 1997; Weisse, 2004). Furthermore, phytoplankton has an important influence on the water quality of the lake; it absorbs nutrients and releases organic substances that interact with dissolved substances in the water (such as chelating agents for heavy metals). It can even constitute a hazard for public health due to its high reproduction rates that may produce algal blooms which affect negatively the scenic value and water quality. Some species are capable of producing toxins deleterious to surrounding fauna (Hudnell, 2008; Kalff, 2002; Lampert & Sommer, 1997; Manahan, 2000). Understanding how the different species react to environmental changes will help to predict community changes and help implement better management and recovery programs for tropical lakes.
Notwithstanding that temperature changes vary little with season in tropical regions, the small changes during hemispherical seasons are enough to modify the stratification condition of the water column in tropical lakes (Lewis, 1973, 1984; Osborne, 2000). In addition it has long been recognized the importance of other climatic elements such as direct solar radiation and winds (Lewis, 1996; Osborne, 2000; Serruya & Pollingher, 1983; Talling, 1969), which are subject to strong variations along the year in tropical regions.
In a former study an almost continuous weekly sampling program was carried out for a year in Lake Barva. The results revealed the occurrence of important changes in phyto-plankton composition over the course of a few days, but it was not possible to ensure that changes were a response to weather variations affecting the lake in the short term (Umaña, 2010). Other former studies have demonstrated the direct influence of the weather on circulation patterns of the Costa Rican lakes (Gocke, 1997; Umaña, 1993). Nevertheless all these studies made it clear that more detailed information on temperature changes with time is needed. In this study weekly variation in phyto-plankton composition is examined in Lake Fraijanes over the course of one year and it is related to the observed changes in temperature in the water column and in weather conditions.
Material and Methods
Description of the lake: The study was performed in Lake Fraijanes (Alajuela, Costa Rica). It is located at 1 650m above sea level, on the Pacific slope of Poas volcano. It is within the Lower Montane wet forest life zone according to Holdridge’s system, which means there is a marked seasonality in precipitation and wind speed (Coen, 1991; Herrera, 1984). It has 2.2Hm2 of surface area and a maximum depth of 6.2m. The lake forms part of a recreational park, and it is surrounded by cypress trees. Around the park there are plantations of ornamental plants grown under shaded greenhouses. The lake is eutrophic. The study was carried out with weekly sampling visits between September 2008 and August 2009, with an additional sampling in November 2009. On some visits a vertical profile of oxygen and temperature was performed at the deepest point in the lake.
Data of daily precipitation, temperature, solar radiation and hours of direct sunshine was obtained from a weather station near the lake (Station Number 84030, 10°08’14.36” lat. N, 84°11’36.58” long. W). Additionally, a portable weather tracker (Krestel® 4500 NV) provided with a weathervane was used on every sampling date, located on the shore, to measure wind speed and direction. Observations of wind strength were also kept according to Beaufort scale on each visit. Four data loggers for temperature (Hobbo®) were placed at different depths (0m, 1m, 3m, and 5m) from a buoy at the deepest place of the lake. The buoy was moored with two anchors to avoid its displacement with currents and entanglement of the rope with the loggers. The loggers were programmed to take a reading every hour. Additionally data with an electronic sensor (YSI®) were made of temperature and dissolved oxygen on several occasions. Loggers were retrieved on December 2008 for data download, but due to delays in obtaining a required permit to continue the study the following year, as well as the occurrence of an earthquake in January 2009, the placement of the loggers back in the lake was delayed until March 2009. They remained in place until the end of the study in August 2009. They were retrieved on September 1st 2009. During the additional field trips samples were taken at two depths (epilimnion and hypolimnion) for the determination of nutrients (nitrate, nitrite, ammonium and orthophosphate). Analyses were performed at the Laboratory of Chemical Oceanography at CIMAR (UCR). A multiple regression analysis was performed to determine the degree of association of water temperature in the lake with weather data.
Surface samples were retrieved every week to determine phytoplankton composition at a spot near the center of the lake. Samples were preserved with acid Lugol’s solution and latter counted according to the Utermöhl (1958) inverted microscope technique. An un-weighted average (UPGMA) cluster analysis using the Morisita similarity index was used with the most abundant and frequent species to determine if different species assemblages occurred with time, followed by a SIM-PER analysis to determine which species were more responsible for the resulting groups. A Correspondece Analysis was performed with the selected species to confirm the results of the cluster analysis. Finally the different assemblages were compared with the classification developed by Reynolds (1984, 2006) of seasonal composition of phytoplankton in lakes. The individual species were also assigned to the morphological based functional groups proposed by Kruk et al. (2010) according to their morphology, to see if changes in the relative contribution of each functional group followed a pattern corresponding to weather variations. All statistical analysis was performed with PAST (Hammer, Harper & Ryan, 2001).
Results
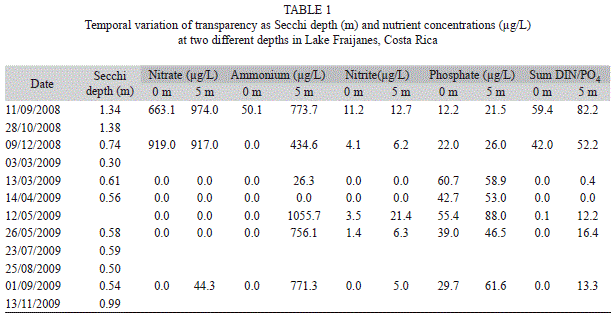
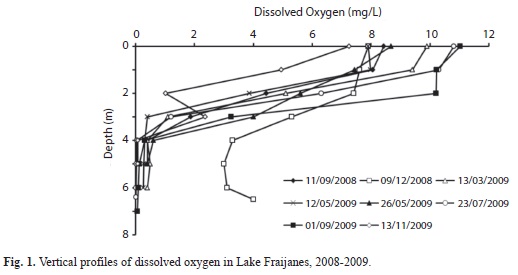
During the study water level fluctuated according to seasonality of rains. The surface level started to drop with the beginning of the dry season, and outflow from the lake stopped between April and August 2009, although the level started to rise by the end of May 2009. Secchi depth varied between 1.38m in October 2008 and 0.3m in March 2009. It remained at 0.5m until the end of the study (Table 1). In November 2009 it was 0.9m, not reaching the level of the previous year. The maximum transparency was observed after the heaviest rains of the season. The hypolimnion was anoxic on most of the occasions in which it was measured. Only in December 2008 there was a small increase in the oxygenation of the hypolimnion (Fig. 1). Nutrient data from epilimnion and hypolimnion also showed a shift (Table 1). At first nitrogen compounds (nitrate, nitrite and ammonium) were high; particularly nitrate concentration in September and December 2008. Later these concentrations were much lower. In contrast, orthophosphate showed slightly higher concentrations between March and September 2009. This change produced a shift in the ratio of N to P that implies a relative scarcity of nitrogen after March 2009.
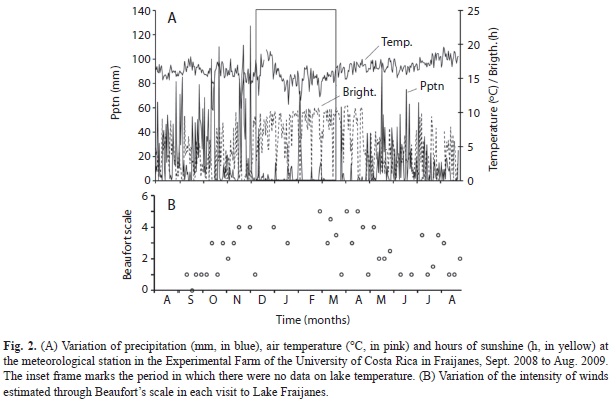
Weather data from the station near the lake showed that the greater precipitation events occurred in October, November and December 2008. There were some cold spells between November 2008 and February 2009 (Fig. 2A). Wind intensity according to Beaufort scale shows that wind is highly variable at the location of the lake. Stronger winds were registered in November and December, but in March and April 2009 there was also some windy weather (Fig. 2B). Data from the smaller weather tracker used beginning December 2008 on each sampling occasion, between 9:00 and 10:00am, showed maximum wind gusts of 11.3m/s in April 2009.
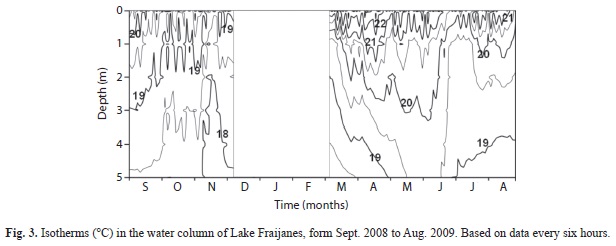
Water temperature of the lake varied between 17.0 and 19.8°C at the bottom and between 17.0 and 27.3°C at the surface. In September 2008 there was a slight thermal gradient down the water column, but the lake remained almost homeothermal for the rest of that year (Fig. 3). During 2008 bottom temperatures cooled down for brief periods followed by a stabilization phase at 18.5°C, a little warmer than before. In March 2009 a warming phase began that builds up gradually in surface waters up to May 2009, when the dry season was at its peak and lake level at its lowest one. A thermal gradient persisted throughout most of this period and the lake remained stratified as indicated by the fact that bottom waters were anoxic on most occasions. Short episodes of partial circulation down to 3m depth occurred during this time. Between May and June 2009 stratification was lost occasionally with the first rains of the season, but stratification was re-established in July and August 2009, when bottom waters again cooled below 19°C.
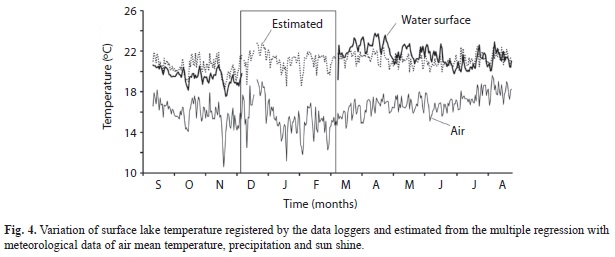
In order to relate lake temperature with weather data, a multiple regression was performed with the mean daily temperature (Tair), precipitation (Pptn) and solar brightness (Brgt) from the weather station close to the lake and the daily average of surface lake temperature (T0m). The regression was significant (F3,247=81.4, p<0.001) (T0m=14.418+0.386٠Tair+0.318٠Brgt+0.0095٠Pptn). It is of interest that deviations between estimated and observed surface temperature don’t distribute randomly over time. From September to December 2008 actual lake temperature was lower than estimated. In contrast from March to July 2009 the observed value was higher than estimate, coinciding with the period of low lake level and inactive surface outlet (Fig. 4). From July to September 2009 estimates match more closely observed values when lake level was rising again with accumulated water from the new rainy season.
Phytoplankton of Lake Fraijanes included 103 taxa, which belonged to the all major algae groups. The most diverse group was green algae (Chlorophyta) with 65 species. They are followed by blue-greens (Cyanobacteria) with eight species. Bacillariophyceae, Crytophyta, Dinophyceae and Euglenophyta were represented by six species each. The least diverse group was Chrysophyta with three species. The more abundant and frequent species were Aphanocapsa sp.1, Coelastrum reticulatum, C. proboscideum, Cosmarium contractum and Didymocystis fina.
Phytoplankton total abundance increased during the study period, mainly as a result of the increase of Cyanobateria (Fig. 5A). The relative contribution of the taxonomic groups shifted with time (Fig. 5B). At the beginning, in September 2008, during the rainy season there was a dominance of Chlorophyta, mainly Chlorococcales, but Cryptophyta was also important. In April 2009 the abundance of Cyanobacteria started to increase and in three weeks’ time they shifted from 10% to 90% of total abundance, mainly a species of the colonial Aphanocapsa. In the months of June and July 2009, there was a short period of deeper mixing, and the dominance of Cyanobacteria was less strong at that time. After that, stratification was reestablished, but with lower surface temperatures, during which Cyanobacteria maintained their dominance but at lower levels. Each of the dominant species was present throughout the study period, but relative abundances shifted throughout the year, following the main seasonal shifts in weather.
The cluster analysis yielded two groups of samples. The first group includes samples from September 11th 2008 to April 14th 2009 (From week 1 to 32). The second group includes samples from April 21st 2009 to September the 1st 2009 (From week 32 to 52) and the additional sampling on November 13th 2009 (week 61). A SIMPER analysis was used to help to determine which species contributed the most to the observed separation of groups. Each of the following species accounted for more than 2% of the variance: Aphanocapsa sp.01, Coelastrum proboscideum, Coelastrum reticulatum, Didymocystis fina, Oocystis lacustris, Chlorella sp.01, Tetraedron incus, Cyclotella meneghiniana and Cosmarium contractum. Together they explain more than 67% of the formation of the groups. In figure 6 are depicted the results of a Correspondence Analysis based on these nine species, that reinforces the identification of the two groups just described.
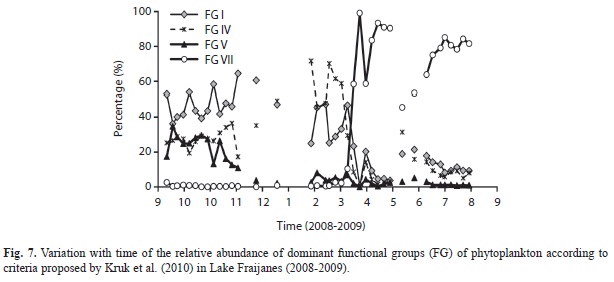
Only functional groups (FG) I, IV, V, VI and VII (as described by Kruk et al., 2010) were present in the lake. Most of the species belonged to FG I. This group was dominant from the beginning of the study in September 2008 until February 2009 (Fig. 7). The FG V was not dominant at any time, but its abundance was also higher at the beginning of the sampling period, from September 2008 until November 2008. FG IV increased after October 2008 and plummeted back in March 2009. FG VI was always present at low densities. Finally FG VII had a sharp increase in March 2009, and remained the most abundant until the end of the study. In May-June 2009 there was a brief period when FG VII decreased slightly and FG I increased at the same time.
Discusion
The presence and persistence of a thermal gradient in the water column of Lake Fraijanes, is noteworthy given that the maximum depth of the lake is less than the 10m that theoretically is required for stratification to occur (Lewis, 1996). The gradual thermal gradient in the lake does not allow for a precise determination of mixing depth, but nevertheless hinders mixing. The stratification of the water column is evidenced by the anoxic conditions below 3m of depth. The lake has been stratified on most other occasions that the author has visited it since the decade of 1970 (unpublished data). There are several possible explanations for this behavior of the lake. One is related to the input of cold ground water as the main source of water which maintains a layer of cool water at the bottom, where heat from the Sun does not reach due to the high turbidity. This turbidity itself could be a second reason. Mazumber and Taylor (1994) also reported on the influence of turbidity on the depth of the thermocline. A similar case was studied by Serruya and Pollingher (1983) in Malaysia (Lake Subang, with 8m of depth) . A third reason is the lowering of the lake level after March and its gradual rise after the onset of the following rainy season. This has been attributed by the managers of the park to the effect of water withdrawal from the aquifer by farmers during the dry season. As a result of the lack of any flow, heat accumulates at the surface. Thus the lake responds not only to direct solar radiation but also to shifts in hydrology.
Phytoplankton variation or seasonality has been studied in detail in lakes of the temperate zone, where it has been possible to predict fluctuations in phytoplankton composition due to drastic changes in temperature and light availability, which have an influence on stratification and mixing (Reynolds 1984, 1996, 2006; Sommer et al 1986). For tropical lakes, Lewis (1978, 1996) predicted that phytoplankton must show the same compositional changes not on a seasonal basis, but more sporadically in response to the occurrence of cold spells that spark the mixing of the lake. Becker, Huszar, Naselli-Flores and Padisák (2008) studied a reservoir in Southern Brazil and discovered a sequence of three periods which they named equilibrium phases according to the Sommer, Padisák, Reynolds & Juhász-Nagy (1993) criterion. One phase was dominated by Cyanobacteria (Anabaena crassa), the second dominated by a green alga (Nephrocytium sp.), and a third phase dominated by a diatom (Asterionella formosa). They attributed the changes to the conditions of stratification and mixing in the reservoir. Also L. G. Barboza, Barboza and Bicudo (2013) found that species of desmids with different morphology based on cell size and surface to volume ratios occur in those habitats for which they are best adapted in response to stratification and low light availability. Barinova, Keshri, Gosh and Sikdar (2012) also found evidence for changes in phytoplankton community composition in different phases of the monsoon cycle in a shallow pond in West Bengal, India, based on changes in relative abundances of the same species through the year.
In the case of Lake Fraijanes data shows that it can stratify for periods relatively long, which coincide with seasonality of winds and rain in the zone. The observed changes in phyto-plankton seem to be responding to changes in weather conditions, which do not preclude the effects from other factors such as nutrient availability and herbivory. For example, there was a shift in N:P ratio, with a relative scarcity of N during the period when Cyanobacteria were dominant which could also contribute to their dominance. The observed shifts in composition of the phytoplankton show close agreement with expected changes according to the model of plankton succession by Reynolds (1984). Cyanobacteria, mainly Aphanocapsa sp., a buoyant species that may form surface blooms (Komárek, 2003) was dominant when stratification was more pronounced and with higher temperature, whereas during deeper mixing and lower temperature periods green algae, mainly Chlorococcales as Coelastrum spp. Oocystis lacustris, Pediasttrum spp., and desmids: Cosmarium contractum and Staurastrum chatoceros were dominant. Another important species was Rhodomonas sp. (Cryptophyta). The most abundant diatom in the lake (Cyclotella stelligera) showed small increments at times of deeper mixing following a stratification period, but did not become dominant or highly abundant at any time. This behavior of Cyanobacteria has been documented in other systems where they are favored during times of relative calm weather, with stagnant waters and higher temperatures (Oliver & Ganf, 2000; Posch, Köster, Salcer & Pernthaler, 2012; Winder & Sommer, 2012). It can be explained by their capacity to develop air vacuoles which allows them to remain in suspension in conditions of low turbulence.
The variation in the relative dominance of phytoplankton groups in Lake Fraijanes coincides broadly with predicted changes according to the model by Reynolds (1996) based on data from temperate zone lakes. This model, which is based on Grime’s (1974) model of reproductive strategies for plants, explains succession of phytoplankton species along a cycle of stratification and mixing and of resource availability according with the following sequence: during stratification period nutrient limitation exacerbates with time and this induces a shift from C (competitors) species to S (stress tolerant) species as nutrients become less available. When the lake is mixed again it returns to a situation of limited light availability due to deep mixing and phytoplankton becomes dominated by R (ruderals) strategists with rapid reproduction which take advantage of the newly available nutrients brought up by the mixing. As stratification is reestablished, the favored strategy shifts again to C species closing the cycle. Reynolds (1984, 1996) created a classification scheme of functional groups that has been modified and improved by Padisák, Crossetti and Naselli-Flores (2009). Reynolds (2006) also recognizes the existence of intermediate strategies in the phytoplankton. These functional groups are distributed into the three strategies described above to explain the seasonal succession of phytoplankton. The model has been elaborated to include the effect of the trophic level of lakes. According to this model, phytoplankton in Lake Fraijanes fluctuated between the dominance of species of J groups (Pediastrum, Coelastrum, and Oocystis) that occur in mixing conditions in eutrophic lakes, and species of group K (Cyanobacteria, Aphanocapsa, and Aphanothece) that occur under more stressful conditions of low nutrient concentrations and stratification. Species typical of deep mixing and light limitation in the Reynolds’ system were not highly abundant, probably because the lake is shallow. Thus, in general terms, phytoplankton of Lake Fraijanes fits well with Reynold’s model. Nevertheless it is advisable to expand these studies to other tropical lakes in order to determine if the scheme of the model is suited to describe tropical conditions in general or to adjust it. The changes in phytoplankton composition in Lake Fraijanes also fitted well with the functional groups proposed by Kruk et al. (2010). During the first period of deep mixing, the lake was dominated by FG I which is characterized by small sized cells, either flagellated or not, having mucilage or not, without a siliceous structure. During the second period of stable stratification FG VII, formed by species with large mucilaginous colonies was dominant. FG IV which corresponds to species with large cell size without mucilage or siliceous structures, flagella or aerotopes, increased during the transition from wet season to dry season and the development of stratification. Finally it is interesting to note that FG VII decreased briefly after the first rains in 2009 and at the same time FG I showed a brief increase, which highlights the importance of weather events on the short term variation of phytoplankton in this small lake. It is also worth mentioning that the use of the functional groups in this case was based on dominant species at each time, but there were other species that belong to other functional groups that did not reach high densities in Lake Fraijanes. If the criterion to detect equilibrium phases of Sommer et al. (1993) is applied to the phytoplankton species composition in Lake Fraijanes, only the period when Aphanocapsa sp. was dominant fits that model. When Chlorophyta was dominant, species diversity was higher and none species reached a dominance level higher than 80%. Finally, it is important to mention that according to Reynold’s model, Lake Fraijanes classifies as hypetrophic, which is matched with the high turbidity and anoxic hypolimnion of the lake. This condition is mainly the result of the influence of human development around the lake with farms and a small town just outside of the recreational park where it is located.
Acknowledgments
This work was possible through the financial support of the Vicerectory of Research of the Universidad de Costa Rica to the Center for Research on Marine Sciences and Limnology (CIMAR), Proyect number 808-A4-513. I am in debt with Juan Carlos Zuñiga, Sheila Castillo, Delia Zavala, Aldo Farah, Anne Marie Gavlas who helped with field trips, to the administration of the INDER, and to Mr. Rafael Rojas Paniagua director of the Recreational Park of Fraijanes and the park personnel for their support and guarding of the buoy and sensors in the lake during the study.
References
Barinova, S., Keshri, J. P., Gosh, S., & Sikdar, J. (2012). The influence of the monsoon climate on phyto-plankton in the Shibpukur pool of Shiva Temple in Burdwann, Weat Bengal, India. Limnological Review, 12, 47-63. doi:10.2478/v10194-011-0044-y [ Links ]
Barboza, L. G., Barboza, F. A. R., & Bicudo, C. E. M. (2013). Adaptive strategies of desmids in two tropical monomictic lakes in southeast Brazil: do morphometric differences promote life strategies selection? Hydrobiologia, 710, 157-171. doi:10.1007/ s10750-013-1458-0. [ Links ]
Becker, V., Huszar, V. L. M., Naselli-Flores, L., & Padisák, J. (2008). Phytoplankton equilibrium phases during thermal stratification in a deep subtropical reservoir. Freshwater Biology, 53, 952-963. [ Links ]
Coen, E. (1991). Clima. In D. H. Janzen (Ed.), Historia natural de Costa Rica. (pp. 35-46). San José: Universidad de Costa Rica. [ Links ]
Grime, J. P. (1974). Vegetation classification by reference to strategies. Nature, 250, 26-31. [ Links ]
Hammer, Ø., Harper, D. A. T., & Ryan, P. D. (2001). PAST: Paleotological Statistics software package for education and data analysis. Paleontologia Electronica, 4, 9. [ Links ]
Harris, G. P. (1978). Photosynthesis, productivity and growth. Ergebnisse der Limnologie, 10, 1-163. [ Links ]
Henrikson, L., Hindlar, A., & Abrahamsson, I. (2005). Restoring acidified lakes: An overview. In P. E., O´Sullivan & C. S. Reynolds (Eds.), The lakes handbook: lake restoration and rahabilitation. Vol 2. (pp. 483-510). Oxford, UK: Blackwell. [ Links ]
Herrera, W. (1984). Clima de Costa Rica. In L. D. Gómez (Ed.), Vegetación y clima de Costa Rica. Vol. 2. San José, C.R.: EUNED. [ Links ]
Hudnell, H. K. (Ed.). (2008). Cyanobacterial harmful algal blooms: state of the science and research needs. Advances in Experimental Medicine and Biology, 619, 1-950. [ Links ]
Kalff, J. (2002). Limnology. New Jersey: Prentice. [ Links ]
Komárek, J. (2003). Coccoid and colonial Cyanobacteria. In J. D. Wehr & R. G. Sheath (Eds.) Freshwater algae of North America (pp. 59-116). San Diego, CA: Academic. [ Links ]
Kruk, C., Huszar, V. L. M., Peeters, E. T. H. M., Bonilla, S., Costa, L., Lurling, M., Reynolds, C. S., & Scheffer, M. (2010). A morphological classification capturing functional variation in phytoplankton. Freshwater Biology, 55, 614-27. [ Links ]
Lampert, W., & Sommer, U. (1997). Limnoecology: the ecology of lakes and streams. New York, NY: Oxford. [ Links ]
Lewis, W. M., Jr. (1978). Dynamics and succession of the phytoplankton in a tropical lake: Lake Lanao, Philippines. Journal of Ecology, 66, 849-880. [ Links ]
Lewis, W. M., Jr. (1986). Phytoplankton succession in Lake Valencia, Venezuela. Hydrobiologia, 138, 189-203. [ Links ]
Lewis, W. M., Jr. (1984). The thermal regime of Lake Lanao (Philippines) and its theoretical implications for tropical lakes. Limnology and Oceanography, 18, 200-217. [ Links ]
Lewis, W. M., Jr. (1996). Tropical lakes: how latitude makes a difference. In F. Schiemer & K. T. Boland (Eds.), Perspectives in tropical limnology (pp. 43-64). Amsterdam: SPB Academic Publishing. [ Links ]
Manahan, S. E. (2000). Environmental Chemistry. 7th Ed. Boca Raton, CA: CRC. [ Links ]
Margalef, R. (1978). Life-forms of phytoplankton as survival alternatives in an unstable environment. Oceanologica Acta, 1, 493-509. [ Links ]
Mazumder, A., & Taylor, W. D. (1994). Thermal structure of lakes varying in size and water clarity. Limnology and Oceanography, 39, 968-976. [ Links ]
Mills, E. L., Leach, J. H., Carlton, J. T., & Secor, C. L. (1994). Exotic species and the integrity of the Great Lakes. BioScience, 44, 666-676. [ Links ]
Oliver, R. L. & Ganf, G. G. (2000). Freshwater blooms. In B. A. Whitton & M. Potts (Eds.), The ecology of Cyanobacteria I: their diversity in time and space (pp. 149-194). España: Kluwer. [ Links ]
Osborne, P. L. (2000). Tropical ecosystems and ecological concepts. UK, Cambridge: Cambridge. [ Links ]
Padisak, J., Crossetti, L. O., & Naselli-Flores, L. (2009). Use and misuse in the application of the phyto-plankton functional classification: a critical review with updates. Hydrobiologia, 621, 1-19. [ Links ]
Posch, T., Köster, O., Salcher, M. M., & Pernthaler, J. (2012). Harmful filamentous cyanobacteria favored by reduced water turnover with lake warming. Nature Climate Change DOI: 10.1038/NCLIMATE1581. [ Links ]
Reynolds, C. S. (1984). Phytoplankton periodicity: the interactions of form, function and environmental variability. Freshwater Biology, 14, 111-142. [ Links ]
Reynolds, C. S. (1988). Funcional morphology and the adaptive strategies of freshwater phytoplankton. In C. D. Sandgren (Ed.), Growth and reproductive strategies of freshwater phytoplankton (pp. 388-433). Cambridge, UK: Cambridge. [ Links ]
Reynolds, C. S. (1996). The plant life of the pelagic. Ver-handlungen Internationale Vereinigung für Theoretis-che und Angewandte Limnologie, 26, 97-113. [ Links ]
Reynolds, C. S. (2006). Ecology of Phytoplankton. Cambridge, UK: Cambridge. [ Links ]
Reynolds, C. S. (1988). Funcional morphology and the adaptive strategies of freshwater phytoplankton. In C. D. Sandgren (Ed.), Growth and reproductive strategies of freshwater phytoplankton (pp. 388-433). Cambridge, UK: Cambridge. [ Links ]
Schiemer, F. (1996). Significance of fifter-feeding fish in tropical freshwaters. In F. Schiemer & K. T. Boland (Eds.), Perspectives in tropical limnology (pp. 65-76). Amsterdam, Holand: SPB Academic. [ Links ]
Schindler, D. W., Kling, H., Schmidt, R. V., Prokopowich, J., Frost, V. E., Reid, R. A., & Capel, M. (1973). Eutrophication of lake 227 by addition of phosphate and nitrate: the second, third and fourth years of enrichment, 1970, 1971 and 1972. Canadian Journal of Fisheries Research Board, 40, 1425-1440. [ Links ]
Schindler, D. W. (1988). Effects of acid rain on freshwater ecosystems. Science, 239, 149-157. [ Links ]
Serruya, C. & Pollingher, U. (1983). Lakes of the warm belt. Cambridge, UK: Cambridge. [ Links ]
Sommer, U. (Ed.). (1989). Plankton ecology: succession in plankton communities. Berlin, Germany: Springer. [ Links ]
Sommer, U. Z., Gliwicz, M., Lampert, W., & Duncan, A. (1986). The PEG-model of seasonal succession of phytoplanktonic events in fresh waters. Archiv für Hydrobiologie, 106, 433-471. [ Links ]
Sommer, U., Padisák, J., Reynolds, C. S., & Juhász-Nagy, P. (1993) Hutchinson‘s heritage: the diversity-disturbance relationship in phytoplankton. Hydrobiologia, 249, 1-7. [ Links ]
Sterner, R. W. (1989). The role of grazers in phytoplankton succession. In Sommer, U. (Ed.), Plankton ecology: succession in plankton communities (pp. 107-171). Berlin, Germany: Springer. [ Links ]
Talling, J. F. (1969). The incidence of vertical mixing, and some biological and chemical consequences, in tropical African lakes. Verhandlungen Internationale Vereinigung für Theoretische und Angewandte Limnologie, 17, 998-1012. [ Links ]
Tilman, D., Kilham, S. S., & Kilham, P. (1982). Phyto-plankton community ecology: the role of limiting nutrients. Annual Review of Ecology and Systematics, 13, 349-372. [ Links ]
Umaña, V. G. (1993). The planktonic association of Lake Hule. Revista de Biología Tropical, 41, 163-171. [ Links ]
Umaña, V. G. (2010). Comparison of basic limnological aspects of some crater lakes in the Cordillera Volcánica Central, Costa Rica. Revista Geológica de América Central, 43, 137-145. [ Links ]
Utermöhl, H. (1958). Zur Vervolkommung der quantitativen Phytoplankton-Methodik. Mitteilungen Internationale Vereinigung für Theoretische und Angewandte Limnologie, 9, 1-38. [ Links ]
Vollenweider, R. A. (1968). Scientific fundamentals of the eutrophication of lakes and flowing waters, with particular reference to nitrogen and phosphorus as factors of europhication. Technical Report DAS/ CSI/68.27. OECD, Paris. [ Links ]
Weisse, T., (2004). Pelagic microbes – protozoa and the microbial food web. In P. E. O’Sullivan & C. S. Reynolds (Eds.), The lakes handbook: limnology and limnetic ecology. Vol. 1. (pp. 417- 460). U.K., Oxford: Blackwell. [ Links ]
Winder, M., & Sommer, U. (2012). Phytoplankton response to a climate change. Hydrobiologia, 698, 5-16. doi:10.1007/s10750-012-1149-2. [ Links ]
Barboza, L. G., Barboza, F. A. R., & Bicudo, C. E. M. (2013). Adaptive strategies of desmids in two tropical monomictic lakes in southeast Brazil: do morphometric differences promote life strategies selection? Hydrobiologia, 710, 157-171. doi:10.1007/ s10750-013-1458-0. [ Links ]
Becker, V., Huszar, V. L. M., Naselli-Flores, L., & Padisák, J. (2008). Phytoplankton equilibrium phases during thermal stratification in a deep subtropical reservoir. Freshwater Biology, 53, 952-963. [ Links ]
Coen, E. (1991). Clima. In D. H. Janzen (Ed.), Historia natural de Costa Rica. (pp. 35-46). San José: Universidad de Costa Rica. [ Links ]
Grime, J. P. (1974). Vegetation classification by reference to strategies. Nature, 250, 26-31. [ Links ]
Hammer, Ø., Harper, D. A. T., & Ryan, P. D. (2001). PAST: Paleotological Statistics software package for education and data analysis. Paleontologia Electronica, 4, 9. [ Links ]
Harris, G. P. (1978). Photosynthesis, productivity and growth. Ergebnisse der Limnologie, 10, 1-163. [ Links ]
Henrikson, L., Hindlar, A., & Abrahamsson, I. (2005). Restoring acidified lakes: An overview. In P. E., O´Sullivan & C. S. Reynolds (Eds.), The lakes handbook: lake restoration and rahabilitation. Vol 2. (pp. 483-510). Oxford, UK: Blackwell. [ Links ]
Herrera, W. (1984). Clima de Costa Rica. In L. D. Gómez (Ed.), Vegetación y clima de Costa Rica. Vol. 2. San José, C.R.: EUNED. [ Links ]
Hudnell, H. K. (Ed.). (2008). Cyanobacterial harmful algal blooms: state of the science and research needs. Advances in Experimental Medicine and Biology, 619, 1-950. [ Links ]
Kalff, J. (2002). Limnology. New Jersey: Prentice. [ Links ]
Komárek, J. (2003). Coccoid and colonial Cyanobacteria. In J. D. Wehr & R. G. Sheath (Eds.) Freshwater algae of North America (pp. 59-116). San Diego, CA: Academic. [ Links ]
Kruk, C., Huszar, V. L. M., Peeters, E. T. H. M., Bonilla, S., Costa, L., Lurling, M., Reynolds, C. S., & Scheffer, M. (2010). A morphological classification capturing functional variation in phytoplankton. Freshwater Biology, 55, 614-27. [ Links ]
Lampert, W., & Sommer, U. (1997). Limnoecology: the ecology of lakes and streams. New York, NY: Oxford. [ Links ]
Lewis, W. M., Jr. (1978). Dynamics and succession of the phytoplankton in a tropical lake: Lake Lanao, Philippines. Journal of Ecology, 66, 849-880. [ Links ]
Lewis, W. M., Jr. (1986). Phytoplankton succession in Lake Valencia, Venezuela. Hydrobiologia, 138, 189-203. [ Links ]
Lewis, W. M., Jr. (1984). The thermal regime of Lake Lanao (Philippines) and its theoretical implications for tropical lakes. Limnology and Oceanography, 18, 200-217. [ Links ]
Lewis, W. M., Jr. (1996). Tropical lakes: how latitude makes a difference. In F. Schiemer & K. T. Boland (Eds.), Perspectives in tropical limnology (pp. 43-64). Amsterdam: SPB Academic Publishing. [ Links ]
Manahan, S. E. (2000). Environmental Chemistry. 7th Ed. Boca Raton, CA: CRC. [ Links ]
Margalef, R. (1978). Life-forms of phytoplankton as survival alternatives in an unstable environment. Oceanologica Acta, 1, 493-509. [ Links ]
Mazumder, A., & Taylor, W. D. (1994). Thermal structure of lakes varying in size and water clarity. Limnology and Oceanography, 39, 968-976. [ Links ]
Mills, E. L., Leach, J. H., Carlton, J. T., & Secor, C. L. (1994). Exotic species and the integrity of the Great Lakes. BioScience, 44, 666-676. [ Links ]
Oliver, R. L. & Ganf, G. G. (2000). Freshwater blooms. In B. A. Whitton & M. Potts (Eds.), The ecology of Cyanobacteria I: their diversity in time and space (pp. 149-194). España: Kluwer. [ Links ]
Osborne, P. L. (2000). Tropical ecosystems and ecological concepts. UK, Cambridge: Cambridge. [ Links ]
Padisak, J., Crossetti, L. O., & Naselli-Flores, L. (2009). Use and misuse in the application of the phyto-plankton functional classification: a critical review with updates. Hydrobiologia, 621, 1-19. [ Links ]
Posch, T., Köster, O., Salcher, M. M., & Pernthaler, J. (2012). Harmful filamentous cyanobacteria favored by reduced water turnover with lake warming. Nature Climate Change DOI: 10.1038/NCLIMATE1581. [ Links ]
Reynolds, C. S. (1984). Phytoplankton periodicity: the interactions of form, function and environmental variability. Freshwater Biology, 14, 111-142. [ Links ]
Reynolds, C. S. (1988). Funcional morphology and the adaptive strategies of freshwater phytoplankton. In C. D. Sandgren (Ed.), Growth and reproductive strategies of freshwater phytoplankton (pp. 388-433). Cambridge, UK: Cambridge. [ Links ]
Reynolds, C. S. (1996). The plant life of the pelagic. Ver-handlungen Internationale Vereinigung für Theoretis-che und Angewandte Limnologie, 26, 97-113. [ Links ]
Reynolds, C. S. (2006). Ecology of Phytoplankton. Cambridge, UK: Cambridge. [ Links ]
Reynolds, C. S. (1988). Funcional morphology and the adaptive strategies of freshwater phytoplankton. In C. D. Sandgren (Ed.), Growth and reproductive strategies of freshwater phytoplankton (pp. 388-433). Cambridge, UK: Cambridge. [ Links ]
Schiemer, F. (1996). Significance of fifter-feeding fish in tropical freshwaters. In F. Schiemer & K. T. Boland (Eds.), Perspectives in tropical limnology (pp. 65-76). Amsterdam, Holand: SPB Academic. [ Links ]
Schindler, D. W., Kling, H., Schmidt, R. V., Prokopowich, J., Frost, V. E., Reid, R. A., & Capel, M. (1973). Eutrophication of lake 227 by addition of phosphate and nitrate: the second, third and fourth years of enrichment, 1970, 1971 and 1972. Canadian Journal of Fisheries Research Board, 40, 1425-1440. [ Links ]
Schindler, D. W. (1988). Effects of acid rain on freshwater ecosystems. Science, 239, 149-157. [ Links ]
Serruya, C. & Pollingher, U. (1983). Lakes of the warm belt. Cambridge, UK: Cambridge. [ Links ]
Sommer, U. (Ed.). (1989). Plankton ecology: succession in plankton communities. Berlin, Germany: Springer. [ Links ]
Sommer, U. Z., Gliwicz, M., Lampert, W., & Duncan, A. (1986). The PEG-model of seasonal succession of phytoplanktonic events in fresh waters. Archiv für Hydrobiologie, 106, 433-471. [ Links ]
Sommer, U., Padisák, J., Reynolds, C. S., & Juhász-Nagy, P. (1993) Hutchinson‘s heritage: the diversity-disturbance relationship in phytoplankton. Hydrobiologia, 249, 1-7. [ Links ]
Sterner, R. W. (1989). The role of grazers in phytoplankton succession. In Sommer, U. (Ed.), Plankton ecology: succession in plankton communities (pp. 107-171). Berlin, Germany: Springer. [ Links ]
Talling, J. F. (1969). The incidence of vertical mixing, and some biological and chemical consequences, in tropical African lakes. Verhandlungen Internationale Vereinigung für Theoretische und Angewandte Limnologie, 17, 998-1012. [ Links ]
Tilman, D., Kilham, S. S., & Kilham, P. (1982). Phyto-plankton community ecology: the role of limiting nutrients. Annual Review of Ecology and Systematics, 13, 349-372. [ Links ]
Umaña, V. G. (1993). The planktonic association of Lake Hule. Revista de Biología Tropical, 41, 163-171. [ Links ]
Umaña, V. G. (2010). Comparison of basic limnological aspects of some crater lakes in the Cordillera Volcánica Central, Costa Rica. Revista Geológica de América Central, 43, 137-145. [ Links ]
Utermöhl, H. (1958). Zur Vervolkommung der quantitativen Phytoplankton-Methodik. Mitteilungen Internationale Vereinigung für Theoretische und Angewandte Limnologie, 9, 1-38. [ Links ]
Vollenweider, R. A. (1968). Scientific fundamentals of the eutrophication of lakes and flowing waters, with particular reference to nitrogen and phosphorus as factors of europhication. Technical Report DAS/ CSI/68.27. OECD, Paris. [ Links ]
Weisse, T., (2004). Pelagic microbes – protozoa and the microbial food web. In P. E. O’Sullivan & C. S. Reynolds (Eds.), The lakes handbook: limnology and limnetic ecology. Vol. 1. (pp. 417- 460). U.K., Oxford: Blackwell. [ Links ]
Winder, M., & Sommer, U. (2012). Phytoplankton response to a climate change. Hydrobiologia, 698, 5-16. doi:10.1007/s10750-012-1149-2. [ Links ]
*Correspondencia a:
Gerardo Umaña-Villalobos. Centro de Investigación en Ciencias del Mar y Limnología (CIMAR) / Escuela de Biología, Universidad de Costa Rica. Correo electrónico gerardo.umana@ucr.ac.cr
1. Centro de Investigación en Ciencias del Mar y Limnología (CIMAR) / Escuela de Biología, Universidad de Costa Rica. Correo electrónico gerardo.umana@ucr.ac.cr
Received 28-viii-2013. Corrected 13-i-2014. Accepted 29-i-2014.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}