










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.61 n.4 San José Oct./Dec. 2013
Fast changes in seasonal forest communities due to soil moisture increase after damming
Cambios estacionales acelerados en comunidades boscosas debidos al aumento en la humedad del suelo despúes de la construcción de una represa
Cambios estacionales acelerados en comunidades boscosas debidos al aumento en la humedad del suelo despúes de la construcción de una represa
Vagner Santiago do Vale1*, Ivan Schiavini1, Glein Monteiro Araújo1, André Eduardo Gusson2*, Sérgio de Faria Lopes3*, Ana Paula de Oliveira1, Jamir Afonso do Prado-Júnior1, Carolina de Silvério Arantes1 & Olavo Custódio Dias-Neto4*
*Dirección para correspondencia
Abstract
Local changes caused by dams can have drastic consequences for ecosystems, not only because they change the water regime but also the modification on lakeshore areas. Thus, this work aimed to determine the changes in soil moisture after damming, to understand the consequences of this modification on the arboreal community of dry forests, some of the most endangered systems on the planet. We studied these changes in soil moisture and the arboreal community in three dry forests in the Araguari River Basin, after two dams construction in 2005 and 2006, and the potential effects on these forests. For this, plots of 20m x10m were distributed close to the impoundment margin and perpendicular to the dam margin in two deciduous dry forests and one semi-deciduous dry forest located in Southeastern Brazil, totaling 3.6ha sampled. Besides, soil analysis were undertaken before and after impoundment at three different depths (0-10, 20-30 and 40-50cm). A tree (minimum DBH of 4.77cm) community inventory was made before (T0) and at two (T2) and four (T4) years after damming. Annual dynamic rates of all communities were calculated, and statistical tests were used to determine changes in soil moisture and tree communities. The analyses confirmed soil moisture increases in all forests, especially during the dry season and at sites closer to the reservoir; besides, an increase in basal area due to the fast growth of many trees was observed. The highest turnover occurred in the first two years after impoundment, mainly due to the higher tree mortality especially of those closer to the dam margin. All forests showed reductions in dynamic rates for subsequent years (T2-T4), indicating that these forests tended to stabilize after a strong initial impact. The modifications were more extensive in the deciduous forests, probably because the dry period resulted more rigorous in these forests when compared to semideciduous forest. The new shorelines created by damming increased soil moisture in the dry season, making plant growth easier. We concluded that several changes occurred in the T0-T2 period and at 0-30m to the impoundment, mainly for the deciduous forests, where this community turned into a “riparian-deciduous forest” with large basal area in these patches. However, unlike other transitory disturbances, damming is a permanent alteration and transforms the landscape to a different scenario, probably with major long-term consequences for the environment.
Key words: mortality, recruitment, ingrowth, turnover, net changes, impoundment, tropical forest, riparian forest.
Resumen
Los cambios locales provocados por las represas pueden tener consecuencias para un ecosistema, ya que no solo cambian el régimen de las aguas, sino que también modifican las áreas a la orilla de los lagos. Por lo tanto, este trabajo tuvo como objetivo observar los cambios en la humedad del suelo después de la construcción de represas, para entender las consecuencias de esta modificación a la comunidad arbórea de los bosques secos, que son uno de los sistemas más amenazados del planeta. Después de la construcción de tres represas, se estudiaron los cambios en la humedad del suelo y la comunidad arbórea en 3 bosques secos de la cuenca del río Araguari, en donde se construyeron dos represas en 2005 y 2006. Se demarcaron parcelas de 20x10m que se distribuyeron cerca y perpendicular al margen de la represa en dos bosques secos caducifolios y un bosque seco semideciduo ubicado en el sureste de Brasil, en total se muestrearon 3.6 hectáreas. Se realizó un análisis de la humedad del suelo antes y después de la construcción del embalse en tres profundidades (0-10, 20-30, 40-50cm). El inventario de la comunidad arbórea (DAP de 4.77cm) se realizó antes (T0), dos (T2) y cuatro (T4) años a partir de la retención del agua. Además se calcularon las tasas anuales dinámicas de todas las comunidades, y se utilizaron pruebas estadísticas para confirmar los cambios en la humedad del suelo y las comunidades de árboles. Los análisis confirmaron aumento de humedad del suelo en todos los bosques, especialmente durante la estación seca y en los lugares más cercanos al embalse, con un aumento del área basal debido al rápido crecimiento de muchos árboles. El mayor volumen de modificaciones se produjo en los primeros dos años después de La construcción del embalse, principalmente debido a la mayor mortalidad de los árboles más cerca del margen de la represa. Sin embargo, todos los bosques mostraron reducción de las tasas dinámicas en los años siguientes (T2-T4), lo que indica que estos bosques tienden a estabilizarse después de un fuerte primer impacto. Las modificaciones fueron más severas en los bosques caducifolios, probablemente debido a una estación seca más severa en estos bosques en comparación con el semideciduo. Las nuevas márgenes creadas por el embalse aumentaron la humedad del suelo en la época seca facilitando el crecimiento de muchas plantas. Se concluye que los cambios que se produjeron en el período de T0-T2 y de 0-30m del embalse, principalmente en los bosques caducifolios estan transformando estas comunidades en “bosque de ribera-caducifolia” con gran área basal en estos parches. Sin embargo, este impacto es diferente a otras alteraciones transitorias, así la construcción del embalse es un cambio permanente y transforma el paisaje en otro escenario, probablemente con importantes consecuencias a largo plazo para el ambiente.
Palabras clave: mortalidad, reclutamiento, crecimiento interno, recambio, cambios netos, embalse, bosque tropical, bosque de ribera.
The consequences of artificial reservoirs can be seen in two different ways. Macro-scale consequences that are related to rapid changes in landscape, such as habitat fragmentation due to roads and highways, extensive flooding areas and consequent mortality among many biotic elements, downstream water flow reduction with loss of biomass due to death of trees, and lack of carbon assimilation by photosynthesis (Humborg, Ittekkot, Cociasu, & VonBodungen, 1997; Jansson, Nilsson, Dynesius, & Andersson, 2000; Nilsson & Berggren, 2000). The act of building dams causes large problems, with many changes to the landscape that are easily noticed; however, local modifications in the vegetation and soil near the lakeshore can be hard to be noticed. We consider these local changes as micro-scale consequences, related to problems that can be difficult to comprehend without detailed studies.
There are reports of several examples of micro-scale consequences after dam construction: generation of a large pulse of methane and carbon dioxide emissions (Duchemin, Lucotte, Canuel, & Chamberland, 1995; Fearnside, 2002; Soumis, Duchemin, Canuel, & Lucotte, 2004), changes in biochemistry of water (Humborg et al., 1997), explosion of disease vectors such as mosquitoes (Fearnside, 2005; Luz, 1994; Patz, Graczyk, Geller, & Vittor, 2000), increase in human illness due to stagnant water (Steinmann et al., 2006), and decreased diversity of fungi (Hu, Cai, Chen, Bahkali, & Hyde, 2010), herbs and shrubs (Dynesius, Jansson, Johansson, & Nilsson, 2004; Nilsson, Jansson, & Zinko, 1997). These local problems do not change the landscape over a short time period, but their long-term effects can have drastic consequences. After conversion of a running-water (lotic) system to a still-water (lentic) system, aquatic weeds cover the water, enhancing methane flux to the atmosphere (Fearnside, 2002) and the abundance of carnivorous fish increases, leading to a drastic reduction of fish diversity (Leite & Bittencourt, 1991). These changes in terrestrial environments are critical because plants represent primary producers and the basal component of most ecosystems (Loreau et al., 2001). However, most studies focus on dam impacts on grasses, herbs and shrubs (Mallik & Richardson, 2009; Nilsson, Ekblad, Gardfjell, & Carlberg, 1991; Nilsson & Svedmark, 2002) and are concentrated in temperate environments with low diversity (Dynesius et al., 2004; Jansson et al., 2000; Nilsson et al., 1997), although most dam construction occurs in high-diversity tropical systems dominated by trees (Guo, Li, Xiao, Zhang, & Gan, 2007; Johansson & Nilsson, 2002; Nilsson et al., 1997; Nilsson, Reidy, Dynesius, & Revenga, 2005). Comparisons between dammed and undammed rivers are also frequent (Nilsson et al., 1997; Nilsson & Svedmark, 2002); however, temporal studies that monitor dam consequences over time are lacking.
Monitoring studies that evaluate the dynamics of mature forests (Condit et al., 1999; Lewis et al., 2004; Phillips et al., 2004; Sheil et al., 2000) or tree community changes related to natural or anthropogenic disturbances (Chazdon, Brenes, & Alvarado, 2005; Chazdon et al, 2007; Condit et al., 2004; Machado & Oliveira-Filho, 2010; Oliveira, Curi, Vilela, & Carvalho, 1997) are widespread but do not evaluate dam consequences for forest communities, and this is a particular problem. The majority of the world´s large rivers have a regulated flow (Nilsson et al., 2005), and therefore, dynamic studies should reveal dam impacts on many environments. Moreover, most dams are built to generate electricity (Truffer et al., 2003) and are therefore established on mountainous terrain (Nilsson & Berggren, 2000) to increase the energy production (Truffer et al., 2003). Thus, we chose to evaluate the effects of an upstream dam on tropical dry forests in Southeastern Brazil.
Tropical dry forests are associated with mountainous or at least steep terrain and therefore are an excellent subject of study to infer changes in other forests with similar impact. Moreover, dry forests can also be subdivided into deciduous and semideciduous forests (Oliveira-Filho & Ratter, 2002). Both are physiognomically identical in terms of structural parameters (height of canopy, density and basal area), but deciduous forest occur in rocky soils, which are inefficient at retaining water (Oliveira-Filho & Ratter, 2002), and thus, the consequences of damming on these forests should be distinct. Thus, we chose to evaluate three dry forests under dam construction impacts: two deciduous forests and one semideciduous forest. These forests have a marked dry season with lack of rains, and the water approach after dam construction means a total change in water relations to flora with uncertain consequences. Besides, dry forests are a threatened environment (Espirito-Santo et al., 2009; Miles et al., 2006), so it is important to evaluate dam impacts on these communities.
Considering that even small changes in the water regime level induce changes in vegetation structure (Nilsson, 1996), this work aimed to determine empirically the changes in soil moisture close to the lakeshore after damming, to understand the consequences of this modification in the arboreal community of three dry forests, and to compare the damming impacts on deciduous and semideciduous forests. Since new shorelines created by dams could enhance soil moisture, we predicted that there would be many changes in arboreal structure, such as high dynamic rates (because dams create severe disturbances, and disturbed forests show high dynamic rates) and high ingrowth rates with an increase in basal area (because wet forests have a higher basal area than dry forests (Murphy & Lugo, 1986), and that the most impacted sites would be those closest to the impoundment, where the water approach would enhance soil moisture.
Materials and methods
Study area: This study was carried out in three dry forests (18°47´40´´S - 48°08’57´´W, 18°40´31´´S - 42°24´30´´W and 18°39´13´´S - 48°25´04´´ W) located in the Amador Aguiar Dam Complex (two dams located on the Araguari River with depths of 52m and 55m). All areas had sloped terrains; however, the inclinations of the deciduous forest (in some plots the inclination was more than 30°) were much more pronounced than those of the semideciduous forest. The predominant soil types were dystrophic and eutrophic podzolic soil and dystrophic cambissoil with basalt outcrops with micaxist and biotite gneiss (Baccaro, Medeiros, & Ferreira, 2004; Baruqui & Motta, 1983; Nishiyama, 1989). The first dam, at 624m above sea level (Amador Aguiar Hydroelectric Dam I-AD1), finished flooding in 2005, and has a flooded area of 18.66 km2 (CCBE, 2007). The second dam at 565m elevation (Amador Aguiar Hydroelectric Dam II-AD2), stopped flooding in 2006, and has a flooded area of 45.11km2 (CCBE, 2006).
After damming, three seasonal dry forests (two deciduous with steep slopes and one semideciduous with mild slopes), which had previously been at least 200m distant from any water source (Fig. 1A), now had the impoundment on their edge (since 2005 for AD1 and 2006 for AD2). Unlike other dams, the water level is controlled by the water flow of an upstream dam; therefore, there are no water fluctuations and no floods in any period of the year. The climate is Aw (Koppen, 1948) with a dry winter (April to September) and a rainy summer (October to March), with an average annual temperature of 22°C and average rainfall of around 1 595mm (Santos & Assunção, 2006).
Soil sampling and analysis: In each forest, we took ten soil samples at three different depths: 0-10cm, 20-30cm and 40-50cm, five samples near the riverbed (5m from the water line), and five samples 15m distant from the artificial lake. This separation was important because we aimed to measure how the water increase affected the soil moisture at different depths and the influence of distance from dam on soil moisture. To calculate soil moisture variation, we applied EMBRAPA methodology (EMBRAPA, 1997). We repeated this sampling protocol every three months to cover the middle and end of the rainy and dry seasons. Accordingly, for each year, we cataloged 40 soil samples for each soil depth range. We also repeated soil moisture collections in three distinct years: before spillway construction (T0: original condition without artificial lake influence created by dams), during the first year after impoundment (T1), and during the third year after damming (T3).
We performed some soil moisture analyses for the three periods (T0, T1, and T3). First, to check soil data normality we performed Lilliefors test (Lilliefors, 1967), but the soil data did not show normality. Therefore, to determine damming effects at the different distances, we carried out separate Friedman tests (Friedman, 1939) for 5m and 15m from the shore. These tests were made at each soil depth in each season (middle of rainy, end of rainy, middle of dry, and end of dry season), comparing the three years of measurement (T0, T1, and T3).
Finally, to compare damming effects on soil moisture between 5m (close to shore) and 15m (distant from shore), we performed Wilcoxon tests between each year (T0, T1 and T3) using all data samples from all forests. Thus, we compared soil moisture near and far from shore before damming (T0), one year after damming (T1), and three years after damming (T3) with a pairwise test. The intention was to show that near the shore, the moisture increases after damming more than at 15m distant from the shore. All the analyses were performed using Systat 10.2 (Wilkinson, 2002).
Plant sampling and analysis: The first inventory (T0) was carried out before damming in 2005 (at AD1) and 2006 (at AD2). In each forest, 60 permanent plots of 20m x10m were marked, totaling 1.2ha by area (total of 3.6ha sampled). A total of 10 plots (of 200m width) were established at the site where the river reached flood elevation after damming, and remaining plots were set up perpendicular to the river margin (Fig. 1B). Thus, samples were distributed every 10m perpendicular to the river (0-10m, 10-20m, 20-30m, 30-40m, 40-50m and 50-60m; Fig. 1B). All trees with diameter at breast height (DBH) of at least 4.77cm were tagged with aluminum labels. Stem diameter was measured at 1.30m from the ground and for multiple stems; all live tillers were measured at 1.30m. The first inventory (T0) results were published in 2009 (Kilca, Schiavini, Araújo, & Felfili, 2009; Siqueira, Araújo, & Schiavini, 2009). The second (T2) and third (T4) inventories were carried out two and four years after damming, respectively. These samplings followed the same procedure as the first inventory. New individuals that met inclusion criteria (recruits) were measured and identified, and mortality referred to standing dead trees, fallen trees, or individuals not found.
After testing the data regarding number of individuals and basal area in all forests for three measurement times for normality using the Lilliefors test (data were normal), we compared the number of individuals and basal area of three plant inventories (T0, T2, and T4) in each forest with an ANOVA test, followed by a post-hoc Tukey test. We then tested the number of dead trees and recruits in the T0-T2 and T2-T4 periods with a paired t-test. These tests are important to determine if the changes occurring in the forests truly affected the community structure. The same procedure was performed for the basal area of dead trees, recruits, increment and decrement. We also applied a paired t-test between plots on the basis of distance to river, comparing the T0-T2, T2-T4, and T0-T4 periods to determine in which period the forest changes were more pronounced. In this analysis, after some exploratory investigations, we combined both deciduous forests as a single forest (the damming effects were very similar for both forests) and separated plots into two distance groups: samples near riverbed (0-30m distance) and samples at a distance from riverbed (30-60m distance). All analyses were performed in the Systat 10.2 program (Wilkinson, 2002).
Dynamic rates: We based the community dynamics on mortality, recruitment, outgrowth, and ingrowth rates. Annual mortality (m) and recruitment (r) were calculated in terms of annual rates (see formulas in Sheil et al., 1995, 2000). Outgrowth annual rates refer to the basal area of dead trees plus dead branches and the basal area of living trees (decrement), and ingrowth annual rates refer to basal area of recruits plus growth in the basal area of surviving trees (increment). To evaluate the forest changes, we computed turnover rates for individuals and basal area through mortality-recruitment rates and outgrowth-ingrowth rates (Oliveira-Filho et al., 2007). We then evaluated the net change (Korning & Balslev, 1994) for individuals and basal area and computed an overall net change (ONC, average between individuals and basal area net change).
Results
Soil: Friedman tests near and far from the lakeshore showed increase in soil moisture due to the approach of the water line on the forest margin, for the several periods evaluated (Fig. 2). There was a significant increase in soil moisture after damming in deciduous forests (DFs); however, this was higher at the end of the dry season. In both DFs, soil moisture increased significantly at all depths (F>7.6, p<0.05,) both near (5m) and far from (15m) the lakeshore, but only at depths of 20-30cm and 40-50cm in the semideciduous forest (SF) (F>9.6, p<0.05). In the middle of the dry season, the results were very similar with a significant increase in soil moisture at all depths in DFs (F>7.6, p<0.05) and both near and far from the water line. The SF was an exception, with significant increase near the water line in the middle of the dry season only at 40-50cm (F=6.4, p<0.05). Damming effects on soil moisture were evident; before damming, soil moisture varied about 12-30% in rainy seasons, and 6-16% in dry seasons, otherwise showing higher soil moisture after damming, varying 17-48% in rainy seasons and 15-52% in dry seasons in deciduous forests. These values demonstrated a clear increase in soil moisture, and these changes are summarized in figure 2. We considered soil changes conclusive only when soil moisture analyses were significantly higher for both periods after damming (T1 and T3), when compared to previous dam construction conditions.
For the rainy season (middle and end), the results were more variable. In deciduous forest 2 (DF2) and the semideciduous forest (SF), there was an increase in soil moisture at the 0-10cm depth both near and 15m distant from the lakeshore (F>7.6, p<0.05), and for the middle of the rainy season after damming. In deciduous forest 1 (DF1), there was no significant increase. For 20-30cm depth near the shore, there was an increase in soil moisture in DF2 (F=7.6, p<0.05) after damming. At 15m distant from the shore, and 20-30cm deep, there was an increase in soil moisture only in DF2 (F=7.6, p<0.05). Finally, at 40-50cm deep, far from shore, soil moisture was higher after damming DF1 (F=6.4, p<0.05,) only three years after damming. At the end of the rainy season, all data were inconclusive. Sometimes soil moisture was higher in T0, and other times it was higher after damming (occasionally in T1, in other cases in T3, Fig. 2).
The Wilcoxon comparisons tests showed that before damming there were no differences in soil moistures (Z=0.13, p=0.90); however, after damming (T1 and T3) the soils showed higher moisture near the shore (in T1: Z=3.27, p=0.002; in T3: Z=2.2, p=0.027). Thus, on the basis of all soil results, we concluded that the soil was clearly moister both near and far from the water line after damming but more intensively when closer to the shore (Fig. 3). Damming consequences for soil moisture throughout the dry and rainy season are summarized in figure 3.
Structural changes and dynamics of the forests: There were no strong variations in the number of individuals for the three time periods in any forest (Table 1), with no significant differences (Table 2). Indeed, mortality and recruitment of trees in the T0-T2 and T2-T4 periods were similar (Table 1), masking possible effects on communities. However, basal area showed a clear increase (Table 1), statistically confirmed, in both deciduous forests (DFs) in T0-T4 but not in the semideciduous forest (Table 2). This enlargement occurred mainly due to strong tree increments during the four years after damming (Table 1). Even in SF, the basal area gain was high (11.59% in four years) not only due to increment and mortality in T2-T4 but also to low mortality in T2-T4 (Table 1).
There was a contrast between the first two years of damming and after the second year (T2-T4) for the deciduous and semideciduous forests. In the first two years, mortality and recruitment (number of individuals and basal area) were similar in all forests, while the increment in DFs was higher than in SF. However in T2-T4, mortality in SF was much lower than in deciduous, while recruitment and increment were similar between the three forests (Table 1).
These results led, in general, to high dynamic rates in all forests in T0-T4. However, the dynamic rates were high in T0-T2 and much smaller in T2-T4, especially in both DFs for all rates and for mortality and outgrowth in SF. The ingrowth rates, for example, decreased from 11%/year to less than 7.5%/year in deciduous forest, and recruitment decreased from ~7 to 4.7%/year in deciduous forest 1 and ~7 to ~2%/year in deciduous forest 2 (Table 1); in contrast, ingrowth and recruitment rates in the semideciduous forest were just slightly higher in the T0-T2 than in T2-T4 period (Table 1).
For mortality and outgrowth, T0-T2 rates were higher than those of T2-T4 in all forests; however, these rates declined sharply in the semideciduous forest (mortality from 5.2 to 1.6%/year and outgrowth from 4.5 to 1.6%/year; Table 1). The contrast between T0-T2 and T2-T4 led to high turnover rates (of individuals and basal area) in the first years of damming in all forests, but superior in both deciduous forests (Table 1).
Net change rates helped to differentiate damming effects in these forests, too. In the first two years in both DFs, net change in individuals was low, unlike the basal area, where net increase was large (Table 1). However, SF displayed a negative net change in individuals and a low net increase in basal area (Table 1); therefore, the first two years of damming modified the deciduous forests more than the semideciduous forest, and in a positive manner.
After two years of impoundment, increase in soil moisture showed distinct results. The subsequent period analyzed (T2-T4) showed low negative net chance of individuals (very close to 0) in the deciduous forests, but with a positive value for the semideciduous forest (Table 1) mainly due to very low mortality in the T2-T4 period in the latter (Table 1). The basal area net change was positive and similar for all forests; however, unlike the first two years, net change in the semideciduous forest was superior. Thus, overall net change (ONC) in the deciduous forests in the first two years was higher than in the semideciduous forest (Table 1); otherwise, in T2-T4, ONC was superior in the semideciduous forest.
The paired t-test confirmed that damming impacts were higher in T0-T2. Mortality was significantly higher in all forests (Table 3), but only in the deciduous forest was the recruitment greater in T0-T2. Regarding basal area, only increment was higher in the first two years in all forests, and recruitment in the basal area was superior in the deciduous forests in this period (Table 3). An interesting result was related to basal area mortality in SF, which was very much smaller than in the two DFs in T2-T4 (Table 1) and significantly less than the basal area of dead trees in the first two years of damming (Table 3).
The paired t-test between distances confirmed damming effects in all forests (Table 4). However, the largest variations occurred near the lakeshore (0-30m distance). The number of recruits was higher at 0-30m in T2-T4 in both deciduous and semideciduous forests but only in DFs in the T0-T4 period (Table 4). This occurred due to greater recruitment in the 30-60m samples in SF in the T0-T2 period; meanwhile, in this period, DFs recruited more trees in the 0-30m plots than 30--60m ones. The effect was clearer for the recruits’ basal, where recruitment was higher near the lakeshore than at 30-60m in the T0-T2 period (Table 4). In DFs, this also happened in T2-T4, reflecting more recruits near the dam after four years of damming. However, in SF, in the T2-T4 period, few trees were recruited near the stream (0-30m), and more recruits were found at a distance from the impoundment (30-60m), showing that damming effects were intense only in the first two years (Table 4).
Discussion
Soil moisture and vegetation: The soil moisture analysis successfully showed damming effects on soil moisture over the dry season (middle and end of this season) at all depths (0-10cm, 20-30cm, and 40-50cm) mainly for deciduous forests. Nevertheless, the data were inconclusive for rainy seasons, probably due to heavy rains making any soil moisture sample more influenced by rainfall than by water from the dam. Probably the lack of water is not too harsh in the semideciduous forest, so soil moisture analysis did not demonstrate many changes, but only tendencies of higher soil moisture after damming. Moreover, soil moisture increased both near the lakeshore and at 15m distance for all forests. These results inferred changes in soil hydrology in these forests leading to many forest alterations, mainly because 80% of the root biomass in dry forests is found in the first 50cm of soil depth (Raherison & Grouzis, 2005), and the same occurs generally in all kind of forests (Kiley & Schneider 2005; Tufekcioglu, Raich, Isenhart, & Schultz, 1998). In seasonal environments, soil water shortage is greatly due to lower overall rainfall in winter (Stefan et al., 2007), and thus, it is expected that changes in soil moisture up to 50cm of soil depth imply several changes in the structure of dry forests.
The approach of the water line may have two consequences for a single tree: in the worst case, the water coverage creates anaerobic conditions which can lead to anoxia and cell death in roots, inhibiting plant growth and consequently killing the tree (Blom & Voesenek, 1996; Nilsson & Berggren, 2000); otherwise, soil moisture increase can compensate for the lack of water, making the environment less stressful, reducing the chance of root desiccation, and enhancing plant growth. In deciduous forests, for example, soils are fertile (Kilca et al., 2009; Siqueira et al., 2009) but are deficient in water supply (Pennington, Lavin, & Oliveira-Filho, 2009) due to water shortage in the drought season. These droughts reduce photosynthetic efficiency (Goulden, Munger, Fan, Daube, & Wofsy, 1996; Reichstein et al., 2002) and have an adverse effect on growth (Suarez, Ghermandi, & Kitzberger, 2004). Therefore, if this lack of water is relieved, there are no restrictions to plant growth (Guilherme, Morellato, & Assis, 2004), and an increase in their metabolic efficiency (for example, in photosynthesis) should be expected, leading to fast tree growth.
Increase in basal area: In general, changes occurred mainly in the basal area with strong increment rates and gain (ingrowth) more than 9%/year in the deciduous forest compared to 6%/year in the semideciduous forest after four years of damming (corresponding to an increase in basal area of 4.5m2/ha in all forests). These values were higher than in major dynamic studies in other tropical rain forests (Korning & Balslev, 1994; Lewis et al., 2004; Phillips et al., 2004; Sheil et al., 2000), tropical seasonal forests (Carvalho & Felfili, 2011; Marin, Nygard, Rivas, & Oden, 2005; Swaine, Lieberman, & Hall, 1990), riparian forests (Braga & Rezende, 2007; Felfili, 1995), and even higher forests, which suffer intense impacts (Chazdon et al., 2007; Condit et al., 2004; Machado & Oliveira-Filho, 2010), mainly considering deciduous forests.
These high rates in deciduous forests are probably due to a combination of two favorable abiotic factors: the soil moisture supply (by water approach) and good soil nutrition support (already occurring in these forests). Drought in deciduous forests is more pronounced than in semideciduous forests due to rocky terrain with steep slopes; therefore, drought has a more negative impact on these vegetations. It is known that drought represents a major constraint on plant growth and productivity in most terrestrial plant communities due to a decline in plants’ photosynthetic capacity (Churkina & Running, 1998; Hinckley, Dougherty, Lassoie, Roberts, & Teskey, 1979); otherwise, soil fertility increases net productivity (Malhi et al., 2004; Phillips et al., 2004). Therefore, the combination of water supply and high fertility leads to greater above-ground forest productivity in deciduous more so than semideciduous forests. The ingrowth rates in semideciduous forests are lower because the soil is much less fertile than in deciduous forests (Kilca et al., 2009; Siqueira et al., 2009), which limits plant growth.
High turnovers in first years after damming: When a severe disturbance occurs, reduction in basal area and density is expected (Machado & Oliveira-Filho, 2010), but for the three forests studied, not only did the basal area increase but the density remained constant, due to offsetting of recruitment over mortality. While many small trees died (93% of dead trees were smaller than 20cm in diameter), a great number of trees were recruited (most recruits were between the minimum of 4.77cm and 10cm in diameter); therefore, this change of trees provided high turnover, not only in basal area but in density too. Turnover rates (individuals and basal area) were higher compared to those of the vast majority of other studies in forests (see studies cited in the preceding paragraphs), exemplifying the damming impact on dry forests and how these forests are disrupted.
This difference in turnover is probably due to the interaction of a number of factors. First, the long winter season limits net primary productivity (Pregitzer & Euskirchen, 2004; Stephenson & van Mantgem, 2005) by water scarcity in tropical zones, which means less change in basal area and individuals; however, this limitation was removed by water approach enhancing plant growth. Second, richer soils have higher turnover rates compared to less fertile soils (Phillips et al., 2004; Stephenson & van Mantgem, 2005). Therefore, a fertile soil favors the survival of more individuals if other environmental factors are not limiting. Third, the proximity of the water table reduces the seasonal effects on soil moisture, and a seasonal environments favor pathogens, leading to higher death rates (Givnish, 1999). Furthermore, the death of individuals opens spaces to other trees, leading to higher recruitment rates (Stephenson & van Mantgem, 2005) and, consequently, high turnover rates. In the absence of a significant disturbance, little net change in living mass is expected (Stephenson & van Mantgem, 2005). Other deciduous forests, for example, show less than 4.5%/year turnover rates (Carvalho & Felfili, 2011; Marin et al., 2005; Swaine et al., 1990; Werneck & Franceschinelli, 2004). However, in the dry forests analyzed here, not only small trees died but also some large trees (7% of all dead trees were bigger than 20cm in diameter). Even so, these big trees represented 43% of the total basal area lost, and when these individuals fall they can disturb small trees, enhancing gap creation (Pearson, Burslem, Goeriz, & Dalling, 2003); the turnover of trees in gaps tends to be faster than in non-gap environment. Turnover rates are important because they are positively correlated with net primary productivity (Runyon, Waring, Goward, & Welles, 1994), and they have important implications for carbon capture. More turnover means more carbon in organic matter in trees and more carbon exchange between trees and the entire community.
This result, however, is tendentious. The organic carbon allocated in these forests does not compensate for the carbon lost by flooding of the original riparian forest. Original riparian forests in this region covered about 40m2/ha (Rodrigues, Lopes, Araújo, & Schiavini, 2010), and many were entirely flooded. Furthermore, the increases in basal area and turnover rates were extremely high in the first two years after damming, but decreased in T2-T4 (the period of two to four years of measurement). This occurred for all parameters analyzed for individuals (mortality and recruitment) and basal area (mortality, recruitment, increment and decrement); however, deciduous forests follow one pattern and semideciduous follow another, as we will discuss below. T2-T4 dynamic rates were more comparable to other dry forest, where mortality varies between 2.3 and 5.0%/year and recruitment varies between 1.5 and 4.5%/year (Appolinário, Oliveira-Filho, & Guilherme, 2005; Machado & Oliveira-Filho 2010; Oliveira et al., 1997; Paiva, Araújo, & Pedroni, 2007; Silva & Araújo, 2009; Swaine et al., 1990), but many rates are still high, mainly ingrowth rates (highly influenced by increment in basal area, exceeding 5.45%/year).
Therefore, we expect a stabilization of the forest rates in the coming years, as occurs with a natural forest that suffers an anthropogenic disturbance (Toniato & Oliveira-Filho, 2004). In these forests, in the first years after the disturbance, there are high dynamic rates (mainly high turnover rates); however, over the years, rates tend to stabilize. The difference in dam impact was basal area gain over a few years after the impact. The disturbance caused by the dam was much more intense in the first two years, and therefore, we suggest that all forests with similar impact in the world should be monitored at least every two years and for a long time to get a better view of the effects for better management practices.
Forest heterogeneity and stabilization: Forest changes and stabilization do not follow the same pattern in deciduous and semideciduous forests or even within each forest. For the deciduous forest, recruitment (in individuals and basal area) and increment was intense near the shore (0-30m) but not in the semideciduous forest (with less recruitment near the shore in T0-T2 and more recruits at this distance in T2-T4, with no differences in four years of damming and no differences in increment between distances). Thus, we found a regular arboreal response pattern for the deciduous forest, where recruitment and increment of trees were positively affected after damming in patches near the water table, and were strongly affected in the first two years of impoundment.
The basal area could drive succession (van Breugel, Martinez-Ramos, & Bongers, 2006), and a high increase in basal area occurs in the early successional stage (Harper, Steininger, Tucker, Juhn, & Hawkins, 2007) and/or after disturbance (Aide, Zimmerman, Herrera, Rosario, & Serrano, 1995; van Breugel et al., 2006). The most impacted patch was near the lakeshore, which expected a succession period but an uncommon succession, because the forest before damming was not in an early succession stage. The water approach creates a new condition for water supplies that enhances environmental heterogeneity and divides the community into two main communities: a “riparian-deciduous forest” near the shore, and water stressed forest at a distance from the margin (more similar to the original deciduous forest). This means that the deciduous forest occupies just a narrow strip of the original area threatening these systems much more. Thus, in management and replanting programs, we need to recover, or at least consider recovering, not only the flooded forests but the forest sectors 0–30m from the artificial lakeshore that were lost due to their conversion to a “riparian-deciduous forest.” This means more extensive areas for the conservation of these forests, such as local parks and ecological reserves. For example, one of the remedial practices in the construction of the Amador Aguiar Hydroeletric Dams was the creation of the “Pau Furado State Park,” which includes native dry forests remaining in the region. However, the protected areas were situated only in the flooded forests, and did not consider changes to these forests near the impoundment. Moreover, these forests should be protected at least 30m up to the current protected area to maximize the conservation of the original conditions.
Riparian systems commonly show high heterogeneity perpendicular to the watercourse (Ribeiro & Walter, 2001; Rodrigues & Nave, 2000); however, this occurs naturally in the environment. Here, we had an increase in heterogeneity caused by dam construction, a human impact. The water supply improves tree growth, increasing basal area in many trees, which become thicker near the shore. Without rains in the dry season, low available subsoil moisture hinders plant performance (Borchert, 1994), limiting growth (Suarez et al., 2004). Of course those remote areas were impacted by the river dam, although to a lesser degree. This increase in heterogeneity and great increase in tree increment near the shore was less intense also in the T2-T4 period. Other works demonstrated that impacts on other taxa were more pronounced in the first years after dam disturbance, such as a rapid increase in aquatic vegetation (Nilsson, 1983) or an explosion of fish and invertebrate populations (Leite & Bittencourt, 1991; Luz, 1994; Patz et al., 2000), but many changes tend to stabilize over the years, and here the same occurred for trees, with a stabilization after the first two years following impact. Great impacts tend to change arboreal structure greatly, but this also tends to stabilize over the years (Aide et al., 1995, Chazdon et al., 2005). We could infer that the “riparian-deciduous forest” parameters tend to stabilize, even near the river. However, unlike other transitory disturbances (winds, intense dry periods, cold), damming is a permanent alteration and transforms the landscape to another scenario, probably with major long-term consequences for the environment, and thus, we consider this “riparian-deciduous forest” as a transition between the deciduous forest and the impoundment.
Acknowledgments
The authors thank the Foundation for Research Support of the State of Minas Gerais (FAPEMIG) for financial support. Dr. A. Leyva and Evandro Lisboa Freire helped with English editing of the manuscript.
References
Aide, T. M., Zimmerman, J. K., Herrera L., Rosario M., & Serrano, M. (1995). Forest recovery in abandoned tropical pastures in Puerto Rico. Forest Ecology and Management, 77(1), 77-86. [ Links ]
Appolinário, V., Oliveira-Filho, A. T., & Guilherme, F. A. G. (2005). Tree population and community dynamics in a Brazilian tropical semideciduous forest. Revista Brasileira de Botânica, 28(2), 347-360. [ Links ]
Baccaro, C. A. D., Medeiros, S. M., Ferreira, I. L., & Rodrigues, S. C. (2004). Mapeamento geomorfológico da bacia do rio Araguari (MG). In S.C. Lima, R. J. Santos (Eds.), Gestão Ambiental da Bacia do Rio Araguari–rumo ao desenvolvimento sustentável (p. 1-19). Uberlândia: EDUFU. [ Links ]
Baruqui, R. T., & Motta, P. E. F. (1983). Interpretação de um trecho do mapa de solos do Triângulo Mineiro. Informativo Agropecuário, 9, 45-63. [ Links ]
Blom, C., & Voesenek, L. (1996). Flooding: The survival strategies of plants. Trends in Ecology & Evolution, 11(7), 290-295. [ Links ]
Borchert, R. (1994). Soil and stem water storage determine phenology and distribution of tropical dry forest trees. Ecology, 75(5), 1437-1449. [ Links ]
Braga, F. M. S., & Rezende, A. V. (2007). Dinâmica da vegetação arbórea da Mata de Galeria do Catetinho, Brasilia-DF. Cerne, 13(2), 138-148. [ Links ]
Carvalho, F. A., & Felfili, J. M. (2011). Variações temporais na comunidade arbórea de uma floresta decidual sobre afloramentos calcários no Brasil Central: composição, estrutura e diversidade florística. Acta Botanica Brasilica, 25(1), 203-214. [ Links ]
CCBE. 2006. Informativo Capim Branco. Retrieved from http://www.ccbe.com.br/comunicacao/informativos/mar_abr_06.pdf. [ Links ]
CCBE. 2007. Informativo Capim Branco. Retrieved from http://www.ccbe.com.br/comunicacao/informativos/dez_07.pdf. [ Links ]
Chazdon, R. L., Brenes, A. R., & Alvarado, B. V. (2005). Effects of climate and stand age on annual tree dynamics in tropical second-growth rain forests. Ecology, 86(7), 1808-1815. [ Links ]
Chazdon, R. L., Letcher, S. G., van Breugel, M., Martinez-Ramos, M., Bongers, F., & Finegan, B. (2007). Rates of change in tree communities of secondary Neotropical forests following major disturbances. Biological Science, 362(1478), 273-289. [ Links ]
Churkina, G., & Running, S. W. (1998). Contrasting climatic controls on the estimated productivity of global terrestrial biomes. Ecosystems, 1(2), 206-215. [ Links ]
Condit, R., Aguilar, S., Hernandez, A., Perez, R., Lao, S., Angehr, G., Hubbell, S. P., & Foster, R. B. (2004). Tropical forest dynamics across a rainfall gradient and the impact of an El Niño dry season. Journal of Tropical Ecology, 20, 51-72. [ Links ]
Condit, R., Ashton, P. S., Manokaran, N., LaFrankie, J. V., Hubbell, S. P., & Foster, R. B. (1999). Dynamics of the forest communities at Pasoh and Barro Colorado: comparing two 50-ha plots. Philosophical Biological Sciences, 354(1391), 1739-1748. [ Links ]
Duchemin, E., Lucotte, M., Canuel, R., & Chamberland, A. (1995). Production of the greenhouse gases CH4 and CO2 by hydroelectric reservoirs of the boreal. Global Biogeochemical Cycles, 9(4), 529-540. [ Links ]
Dynesius, M., Jansson, R., Johansson, M. E., & Nilsson, C. (2004). Intercontinental similarities in riparian-plant diversity and sensitivity to river regulation. Ecological Applications, 14(1), 173-191. [ Links ]
EMBRAPA. (1997). Manual de métodos de análise do solo. Ministério de agricultura e do abastecimento. Rio de Janeiro: Centro Nacional de Pesquisa de Solos. [ Links ]
Espirito-Santo, M. M., Sevilha, A. C., Anaya, F. C., Barbosa, R., Fernandes, G. W., Sanchez-Azofeifa, G. A., Scariot, A., de Noronha, S. E., & Sampaio, C. A. (2009). Sustainability of tropical dry forests: Two case studies in southeastern and central Brazil. Forest Ecology and Management, 258(6), 922-930. [ Links ]
Fearnside, P. M. (2002). Greenhouse gas emissions from a hydroelectric reservoir (Brazil’s Tucurui Dam) and the energy policy implications. Water, Air, Soil & Pollution, 133(1), 69-96. [ Links ]
Fearnside, P. M. (2005). Brazil’s Samuel Dam: Lessons for hydroelectric development policy and the environment in Amazonia. Environmental Management, 35, 1-19. [ Links ]
Felfili, J. M. (1995). Diversity, structure and dynamics of a gallery in central Brazil. Vegetatio, 117(1), 1-15. [ Links ]
Friedman, M. (1939). The use of ranks to avoid the assumption of normality implicit in the analysis of variance. Journal of the American Statistical Association, 32(200), 675-701. [ Links ]
Givnish, T. J. (1999). On the causes of gradients in tropical tree diversity. Journal of Ecology, 87(2), 193-210. [ Links ]
Goulden, M. L., Munger, J. W., Fan, S. M., Daube, B. C., & Wofsy, S. C. (1996). Exchange of carbon dioxide by a deciduous forest: Response to interannual climate variability. Science, 271(5255), 1576-1578. [ Links ]
Guilherme, F. A. G., Morellato, L. P. C., & Assis, M. A. (2004). Horizontal and vertical tree community structure in a section of lowland Atlantic Rain Forest in the Intervales State Park, southeastern Brazil. Revista Brasileira de Botanica, 27, 725-737. [ Links ]
Guo, Z. W., Li, Y. M., Xiao, X. M., Zhang, L., & Gan, Y. L. (2007). Hydroelectricity production and forest conservation in watersheds. Ecological Applications, 17(6), 1557-1562. [ Links ]
Harper, G. J., Steininger, M. K., Tucker, C. J., Juhn, D., & Hawkins, F. (2007). Fifty years of deforestation and forest fragmentation in Madagascar. Environmental Conservation, 34(4), 325-333. [ Links ]
Hinckley, T. M., Dougherty, P. M., Lassoie, J. P., Roberts, J. E., & Teskey, R. O. (1979). Severe drought-Impact on tree growth, phenology, net photosynthetic rate and water relations. American Midland Naturalist, 102(2), 307-316. [ Links ]
Hu, D., Cai, L., Chen, H., Bahkali, A. H., & Hyde, K. D. (2010). Fungal diversity on submerged wood in a tropical stream and an artificial lake. Biodiversity and Conservation, 19(13), 3799-3808. [ Links ]
Humborg, C., Ittekkot, V., Cociasu, A., & VonBodungen, B. (1997). Effect of Danube River dam on Black Sea biogeochemistry and ecosystem structure. Nature, 386(6623), 385-388. [ Links ]
Jansson, R., Nilsson, C., Dynesius, M., & Andersson, E. (2000). Effects of river regulation on river-margin vegetation: A comparison of eight boreal rivers. Ecological Applications, 10(1), 203-224. [ Links ]
Johansson, M. E., & Nilsson, C. (2002). Responses of riparian plants to flooding in free-flowing and regulated boreal rivers: an experimental study. Journal of Applied Ecology, 39(6), 971-986. [ Links ]
Kilca, R. V., Schiavini, I., Araújo, G. M., & Felfili, J. M. (2009). Edaphic and structural differences between two seasonal forests in the Cerrado biome. Neotropical Biology and Conservation, 4(3), 150-163. [ Links ]
Kiley, D. K., & Schneider, R. L. (2005). Riparian roots through time, space and disturbance. Plant Soil, 269(1), 259-272. [ Links ]
Koppen, W. (1948). Climatologia: com um estúdio de los climas de la terra. México: Fondo de Cultura Econômica. [ Links ]
Korning, J., & Balslev, H. (1994). Growth-rates and mortality patterns of tropical lowland tree species and the relating to forest structure in amazonian Ecuador. Journal of Tropical Ecology, 10(2), 151-166. [ Links ]
Leite, R. A. N., & Bittencourt, M. M. (1991). Impacto de hidroelétricas sobre a ictiofauna amazônica: O exemplo de Tucuruí. In A. L. Val, R. Figiuolo, & E. Feldberg (Eds.), Bases Científicos para Estratégias de Preservação e Desenvolvimento da Amazônia: Fatos e Perspectivas (85-100). Manaus: Instituto Nacional de Pesquisas da Amazônia (INPA). [ Links ]
Lewis, S. L., Phillips, O. L., Sheil, D., Vinceti, B., Baker, T. R., Brown, S., Graham, A. W., Higuchi, N., Hilbert, D. W., Laurance, W. F., Lejoly, J., Malhi, Y., Monteagudo, A., Vargas, P. N., Sonke, B., Supardi, N., Terborgh, J. W., & Martinez, R. V. (2004). Tropical forest tree mortality, recruitment and turnover rates: calculation, interpretation and comparison when census intervals vary. Journal of Ecology, 92(6), 929-944. [ Links ]
Lilliefors, H. (1967). On the Kolmogorov-Smirnov test for normality with mean and variance unknown. Journal of the American Statistical Association, 62, 399-402. [ Links ]
Loreau, M., Naeem, S., Inchausti, P., Bengtsson, J., Grime, J. P., Hector, A., Hooper, D. U., Huston, M. A., Raffaelli, D., Schmid, B., Tilman, D., & Wardle, D. A. (2001). Ecology - Biodiversity and ecosystem functioning: Current knowledge and future challenges. Science, 294(5543), 804-808. [ Links ]
Luz, S. L. B. (1994). Estudo sobre os culicíneos (diptera: culicidar:culicinar) crepusculares e noturnos, vetores potenciais de patógenos de importância médica e veterinária na Estação Ecológica de Samuel-Rondônia. Rio de Janeiro: Universidade Federal Rural do Rio de Janeiro. [ Links ]
Machado, E. L. M., & Oliveira-Filho, A. T. (2010). Spatial patterns of tree community dynamics are detectable in a small (4 ha) and disturbed fragment of the Brazilian Atlantic forest. Acta Botanica Brasilica, 24(1), 250-261. [ Links ]
Malhi, Y., Baker, T. R., Phillips, O. L., Almeida, S., Alvarez, E., Arroyo, L., Chave, J., Czimczik, C. I., Di Fiore, A., Higuchi, N., Killeen, T. J., Laurance, S. G., Laurance, W. F., Lewis, S. L., Montoya, L. M. M., Monteagudo, A., Neill, D. A., Vargas, P. N., Patino, S., Pitman, N. C. A., Quesada, C. A., Salomao, R., Silva, J. N. M., Lezama, A. T., Martinez, R. V., Terborgh, J., Vinceti, B., & Lloyd, J. (2004). The above-ground coarse wood productivity of 104 Neotropical forest plots. Global Change Biology, 10(5), 563-591. [ Links ]
Mallik, A. U., & Richardson, J. S. (2009). Riparian vegetation change in upstream and downstream reaches of three temperate rivers dammed for hydroelectric generation in British Columbia, Canada. Ecological Engineering, 35(5), 810-819. [ Links ]
Marin, G. C., Nygard, R., Rivas, B. G., & Oden, P. C. (2005). Stand dynamics and basal area change in a tropical dry forest reserve in Nicaragua. Forest Ecology and Management, 208(1), 63-75. [ Links ]
Miles, L., Newton, A. C., DeFries, R. S., Ravilious, C., May, I., Blyth, S., Kapos, V., & Gordon, J. E. (2006). A global overview of the conservation status of tropical dry forests. Journal of Biogeography, 33(3), 491-505. [ Links ]
Lugo, A. E. (1986). Ecology of Tropical Dry Forest. Annual Review of Ecology and Systematics, 17, 67-88. [ Links ]
Nishyiama, L. (1989). Geologia do município de Uberlândia e áreas adjacentes. Sociedade e Natureza, 1(1), 9-16. [ Links ]
Nilsson, C. (1983). Frequency-distributions of vascular plants in the geolittoral vegetation along 2 rivers in northern Sweden. Journal of Biogeography, 10(4), 351-369. [ Links ]
Nilsson, C. (1996). Remediating river margin vegetation along fragmented and regulated rivers in the North: What is possible? Regulated Rivers-Research & Management, 12(4), 415-431. [ Links ]
Nilsson, C., & Berggren, K. (2000). Alterations of riparian ecosystems caused by river regulation. Bioscience, 50(9), 783-792. [ Links ]
Nilsson, C., & Svedmark, M. (2002). Basic principles and ecological consequences of changing water regimes: Riparian plant communities. Environmental Management, 30(4), 468-480. [ Links ]
Nilsson, C., Ekblad, A., Gardfjell, M., & Carlberg, B. (1991). Long-term effects of river regulation on river margin vegetation. Journal of Applied Ecology, 28(3), 963-987. [ Links ]
Nilsson, C., Jansson, R., & Zinko, U. (1997). Long-term responses of river-margin vegetation to water-level regulation. Science, 276(5313), 798-800. [ Links ]
Nilsson, C., Reidy, C. A., Dynesius, M., & Revenga, C. (2005). Fragmentation and flow regulation of the world’s large river systems. Science, 308(5720), 405-408. [ Links ]
Oliveira, A. T., Curi, N., Vilela, E. A., & Carvalho, D. A. (1997). Tree species distribution along soil catenas in a riverside semideciduous forest in southeastern Brazil. Flora, 192(1), 47-64. [ Links ]
Oliveira-Filho, A. T., Carvalho, W. A. C., Machado, E. L. M., Higuchi, P., Appolinário, V., Castro, G. C., Silva, A. C., Santos, R. M., Borges, L. F., Corrêa, B. S., & Alves, J. M. (2007). Dinâmica da comunidade e populações arbóreas da borda e interior de um remanescente florestal na Serra da Mantiqueira, Minas Gerais, em um intervalo de cinco anos (1999-2004). Revista Brasileira de Botânica, 30(1), 149-161. [ Links ]
Oliveira-Filho, A. T., & Ratter, J. A. (2002). Vegetation Physiognomies and Woody Flora of the Cerrado Biome. In P. S. liveira & R. J. Marquis (Eds.), The Cerrados of Brazil. New York: Columbia University Press. [ Links ]
Paiva, L. V., Araújo, G. M., & Pedroni, F. (2007). Structure and dynamics of a wood plant community of a tropical semi-deciduous seasonal forest in the “Estação Ecológica do Panga”, municipality of Uberlândia, Minas Gerais, Brazil. Revista Brasileira de Botânica, 30(3), 365-373. Retrieved from http://www.scielo.br/pdf/rbb/v30n3/03.pdf. [ Links ]
Patz, J. A., Graczyk, T. K., Geller, N., & Vittor, A. Y. (2000). Effects of environmental change on emerging parasitic diseases. International Journal for Parasitology, 30(12), 1395-1405. [ Links ]
Pearson, T. R. H., Burslem, D., Goeriz, R. E., & Dalling, J. W. (2003). Regeneration niche partitioning in neotropical pioneers: effects of gap size, seasonal drought and herbivory on growth and survival. Oecologia, 137(3), 456-465. [ Links ]
Pennington, R. T., Lavin, M., & Oliveira-Filho, A. (2009). Woody Plant Diversity, Evolution, and Ecology in the Tropics: Perspectives from Seasonally Dry Tropical Forests. Annual Review of Ecology Evolution and Systematics, 40, 437-457. [ Links ]
Phillips, O. L., Baker, T. R., Arroyo, L., Higuchi, N., Killeen, T. J., Laurance, W. F., Lewis, S. L., Lloyd, J., Malhi, Y., Monteagudo, A., Neill, D. A., Vargas, P. N., Silva, J. N. M., Terborgh, J., Martinez, R. V., Alexiades, M., Almeida, S., Brown, S., Chave, J., Comiskey, J. A., Czimczik, C. I., Di Fiore, A., Erwin, T., Kuebler, C., Laurance, S. G., Nascimento, H. E. M., Olivier, J., Palacios, W., Patino, S., Pitman, N. C. A., Quesada, C. A., Saldias, M., Lezama, A. T., & Vinceti, B. (2004). Pattern and process in Amazon tree turnover, 1976-2001. Philosophical Transactions of the Royal Society of London Series B-Biological Sciences, 359(1443), 381-407. [ Links ]
Pregitzer, K. S., & Euskirchen, E. S. (2004). Carbon cycling and storage in world forests: biome patterns related to forest age. Global Change Biology, 10(12), 2052-2077. [ Links ]
Raherison, S. M., & Grouzis, M. (2005). Plant biomass, nutrient concentration and nutrient storage in a tropical dry forest in the south-west of Madagascar. Plant Ecology, 180(1), 33-45. [ Links ]
Reichstein, M., Tenhunen, J. D., Roupsard, O., Ourcival, J. M., Rambal, S., Miglietta, F., Peressotti, A., Pecchiari, M., Tirone, G., & Valentini, R. (2002). Severe drought effects on ecosystem CO(2) and H(2)O fluxes at three Mediterranean evergreen sites: revision of current hypotheses? Global Change Biology, 8(10), 999-1017. [ Links ]
Ribeiro, J. F., & Walter, B. M. T. (2001). As matas de galeria no contexto do bioma Cerrado. In J. F. Ribeiro, C. E. L. Fonseca, & J. C. Sousa-Silva (Eds.), Cerrado: caracterização e recuperação de Matas de Galeria (29-45). Planaltina: Embrapa Cerrados. [ Links ]
Rodrigues, R. R., & Nave, A. G. (2000). Heterogeneidade florística das matas ciliares. In R. R. Rodrigues, & H. F. Leitão-Filho (Eds.), Matas ciliares: conservação e recuperação (45-71). São Paulo: EDUSP. [ Links ]
Rodrigues V. H. P., Lopes, S. F., Araújo G. M., & Schiavini, I. (2010). Composição, estrutura e aspéctos ecológicos da floresta ciliar do rio Araguari no Triângulo Mineiro. Hoehnea, 37, 87-105. [ Links ]
Runyon, J, Waring, R. H., Goward, S. N., & Welles, J. M. (1994). Environmental limits on net primary production and light-use efficiency across the Oregon Transect. Ecological Applications, 4(2), 226-237. [ Links ]
Santos, E. R., & Assunção, W. L. (2006). Distribuição espacial das chuvas na microbacia do Córrego do Amanhece, Araguari-MG. Caminhos da Geografia, 6, 41-55. [ Links ]
Sheil, D., Burslem, D., & Alder, D. (1995). The interpretation and misinterpretation of mortality-rate measures. Journal of Ecology, 83(2), 331-333. [ Links ]
Sheil, D., Jennings, S., & Savill, P. (2000). Long-term permanent plot observations of vegetation dynamics in Budongo, a Ugandan rain forest. Journal of Tropical Ecology, 16, 765-800. [ Links ]
Silva, M. R., & Araújo, G. M. (2009). Dinâmica da comunidade arbórea de uma floresta semidecidual em Uberlândia, MG, Brasil. Acta Botanica Brasilica, 23(1), 49-56. [ Links ]
Siqueira, A. S., Araújo, G. M., & Schiavini, I. (2009). Estrutura do componente arbóreo e características edáficas de dois fragmentos de floresta estacional decidual no vale do rio Araguari, MG, Brasil. Acta Botanica Brasilica, 23(1), 10-21. [ Links ]
Soumis, N., Duchemin, E., Canuel, R., & Lucotte, M. (2004). Greenhouse gas emissions from reservoirs of the western United States. Global Biogeochemical Cycles, 18(3), 1-18. [ Links ]
Stefan, J., McDonald, A. J., Johnson, M. S., Feldpaush, T. R., Couto, E. G., & Riha, S. J. (2007). Relationships between soil hydrology and forest structure and composition in the southern Brazilian Amazon. Journal of Vegetation Science, 18(2), 183-194. [ Links ]
Stephenson, N. L., & van Mantgem, P. J. (2005). Forest turnover rates follow global and regional patterns of productivity. Ecology Letters, 8(5), 524-531. [ Links ]
Suarez, M. L, Ghermandi, L., & Kitzberger, T. (2004). Factors predisposing episodic drought-induced tree mortality in Nothofagus - site, climatic sensitivity and growth trends. Journal of Ecology, 92(6), 954-966. [ Links ]
Swaine, M. D., Lieberman, D., & Hall, J. B. (1990). Structure and dynamics of a tropical dry forest in Ghana. Vegetatio, 88(1), 31-51. [ Links ]
Toniato, M. T. Z., & Oliveira-Filho, A. T. (2004). Variations in tree community composition and structure in a fragment of tropical semideciduous forest in southeastern Brazil related to different human disturbance histories. Forest Ecology and Management, 198(1), 319-339. [ Links ]
Truffer, B., Bratrich, C., Markard, J., Peter, A., Wuest, A., & Wehrli, B. (2003). Green Hydropower: The contribution of aquatic science research to the promotion of sustainable electricity. Aquatic Sciences, 65(2), 99-110. [ Links ]
Tufekcioglu, A., Raich, J. W., Isenhart, T. M., & Schultz, R. C. (1998). Fine root dynamics, coarse root biomass, root distribution, and soil respiration in a multispecies riparian buffer in Central Iowa, USA. Agroforestry Systems, 44(2), 163-174. [ Links ]
van Breugel, M., Martinez-Ramos, M., & Bongers, F. (2006). Community dynamics during early secondary succession in Mexican tropical rain forests. Journal of Tropical Ecology, 22, 663-674. [ Links ]
Wilkinson, L. (2002). Systat software 10.2. Chicago. [ Links ]
Appolinário, V., Oliveira-Filho, A. T., & Guilherme, F. A. G. (2005). Tree population and community dynamics in a Brazilian tropical semideciduous forest. Revista Brasileira de Botânica, 28(2), 347-360. [ Links ]
Baccaro, C. A. D., Medeiros, S. M., Ferreira, I. L., & Rodrigues, S. C. (2004). Mapeamento geomorfológico da bacia do rio Araguari (MG). In S.C. Lima, R. J. Santos (Eds.), Gestão Ambiental da Bacia do Rio Araguari–rumo ao desenvolvimento sustentável (p. 1-19). Uberlândia: EDUFU. [ Links ]
Baruqui, R. T., & Motta, P. E. F. (1983). Interpretação de um trecho do mapa de solos do Triângulo Mineiro. Informativo Agropecuário, 9, 45-63. [ Links ]
Blom, C., & Voesenek, L. (1996). Flooding: The survival strategies of plants. Trends in Ecology & Evolution, 11(7), 290-295. [ Links ]
Borchert, R. (1994). Soil and stem water storage determine phenology and distribution of tropical dry forest trees. Ecology, 75(5), 1437-1449. [ Links ]
Braga, F. M. S., & Rezende, A. V. (2007). Dinâmica da vegetação arbórea da Mata de Galeria do Catetinho, Brasilia-DF. Cerne, 13(2), 138-148. [ Links ]
Carvalho, F. A., & Felfili, J. M. (2011). Variações temporais na comunidade arbórea de uma floresta decidual sobre afloramentos calcários no Brasil Central: composição, estrutura e diversidade florística. Acta Botanica Brasilica, 25(1), 203-214. [ Links ]
CCBE. 2006. Informativo Capim Branco. Retrieved from http://www.ccbe.com.br/comunicacao/informativos/mar_abr_06.pdf. [ Links ]
CCBE. 2007. Informativo Capim Branco. Retrieved from http://www.ccbe.com.br/comunicacao/informativos/dez_07.pdf. [ Links ]
Chazdon, R. L., Brenes, A. R., & Alvarado, B. V. (2005). Effects of climate and stand age on annual tree dynamics in tropical second-growth rain forests. Ecology, 86(7), 1808-1815. [ Links ]
Chazdon, R. L., Letcher, S. G., van Breugel, M., Martinez-Ramos, M., Bongers, F., & Finegan, B. (2007). Rates of change in tree communities of secondary Neotropical forests following major disturbances. Biological Science, 362(1478), 273-289. [ Links ]
Churkina, G., & Running, S. W. (1998). Contrasting climatic controls on the estimated productivity of global terrestrial biomes. Ecosystems, 1(2), 206-215. [ Links ]
Condit, R., Aguilar, S., Hernandez, A., Perez, R., Lao, S., Angehr, G., Hubbell, S. P., & Foster, R. B. (2004). Tropical forest dynamics across a rainfall gradient and the impact of an El Niño dry season. Journal of Tropical Ecology, 20, 51-72. [ Links ]
Condit, R., Ashton, P. S., Manokaran, N., LaFrankie, J. V., Hubbell, S. P., & Foster, R. B. (1999). Dynamics of the forest communities at Pasoh and Barro Colorado: comparing two 50-ha plots. Philosophical Biological Sciences, 354(1391), 1739-1748. [ Links ]
Duchemin, E., Lucotte, M., Canuel, R., & Chamberland, A. (1995). Production of the greenhouse gases CH4 and CO2 by hydroelectric reservoirs of the boreal. Global Biogeochemical Cycles, 9(4), 529-540. [ Links ]
Dynesius, M., Jansson, R., Johansson, M. E., & Nilsson, C. (2004). Intercontinental similarities in riparian-plant diversity and sensitivity to river regulation. Ecological Applications, 14(1), 173-191. [ Links ]
EMBRAPA. (1997). Manual de métodos de análise do solo. Ministério de agricultura e do abastecimento. Rio de Janeiro: Centro Nacional de Pesquisa de Solos. [ Links ]
Espirito-Santo, M. M., Sevilha, A. C., Anaya, F. C., Barbosa, R., Fernandes, G. W., Sanchez-Azofeifa, G. A., Scariot, A., de Noronha, S. E., & Sampaio, C. A. (2009). Sustainability of tropical dry forests: Two case studies in southeastern and central Brazil. Forest Ecology and Management, 258(6), 922-930. [ Links ]
Fearnside, P. M. (2002). Greenhouse gas emissions from a hydroelectric reservoir (Brazil’s Tucurui Dam) and the energy policy implications. Water, Air, Soil & Pollution, 133(1), 69-96. [ Links ]
Fearnside, P. M. (2005). Brazil’s Samuel Dam: Lessons for hydroelectric development policy and the environment in Amazonia. Environmental Management, 35, 1-19. [ Links ]
Felfili, J. M. (1995). Diversity, structure and dynamics of a gallery in central Brazil. Vegetatio, 117(1), 1-15. [ Links ]
Friedman, M. (1939). The use of ranks to avoid the assumption of normality implicit in the analysis of variance. Journal of the American Statistical Association, 32(200), 675-701. [ Links ]
Givnish, T. J. (1999). On the causes of gradients in tropical tree diversity. Journal of Ecology, 87(2), 193-210. [ Links ]
Goulden, M. L., Munger, J. W., Fan, S. M., Daube, B. C., & Wofsy, S. C. (1996). Exchange of carbon dioxide by a deciduous forest: Response to interannual climate variability. Science, 271(5255), 1576-1578. [ Links ]
Guilherme, F. A. G., Morellato, L. P. C., & Assis, M. A. (2004). Horizontal and vertical tree community structure in a section of lowland Atlantic Rain Forest in the Intervales State Park, southeastern Brazil. Revista Brasileira de Botanica, 27, 725-737. [ Links ]
Guo, Z. W., Li, Y. M., Xiao, X. M., Zhang, L., & Gan, Y. L. (2007). Hydroelectricity production and forest conservation in watersheds. Ecological Applications, 17(6), 1557-1562. [ Links ]
Harper, G. J., Steininger, M. K., Tucker, C. J., Juhn, D., & Hawkins, F. (2007). Fifty years of deforestation and forest fragmentation in Madagascar. Environmental Conservation, 34(4), 325-333. [ Links ]
Hinckley, T. M., Dougherty, P. M., Lassoie, J. P., Roberts, J. E., & Teskey, R. O. (1979). Severe drought-Impact on tree growth, phenology, net photosynthetic rate and water relations. American Midland Naturalist, 102(2), 307-316. [ Links ]
Hu, D., Cai, L., Chen, H., Bahkali, A. H., & Hyde, K. D. (2010). Fungal diversity on submerged wood in a tropical stream and an artificial lake. Biodiversity and Conservation, 19(13), 3799-3808. [ Links ]
Humborg, C., Ittekkot, V., Cociasu, A., & VonBodungen, B. (1997). Effect of Danube River dam on Black Sea biogeochemistry and ecosystem structure. Nature, 386(6623), 385-388. [ Links ]
Jansson, R., Nilsson, C., Dynesius, M., & Andersson, E. (2000). Effects of river regulation on river-margin vegetation: A comparison of eight boreal rivers. Ecological Applications, 10(1), 203-224. [ Links ]
Johansson, M. E., & Nilsson, C. (2002). Responses of riparian plants to flooding in free-flowing and regulated boreal rivers: an experimental study. Journal of Applied Ecology, 39(6), 971-986. [ Links ]
Kilca, R. V., Schiavini, I., Araújo, G. M., & Felfili, J. M. (2009). Edaphic and structural differences between two seasonal forests in the Cerrado biome. Neotropical Biology and Conservation, 4(3), 150-163. [ Links ]
Kiley, D. K., & Schneider, R. L. (2005). Riparian roots through time, space and disturbance. Plant Soil, 269(1), 259-272. [ Links ]
Koppen, W. (1948). Climatologia: com um estúdio de los climas de la terra. México: Fondo de Cultura Econômica. [ Links ]
Korning, J., & Balslev, H. (1994). Growth-rates and mortality patterns of tropical lowland tree species and the relating to forest structure in amazonian Ecuador. Journal of Tropical Ecology, 10(2), 151-166. [ Links ]
Leite, R. A. N., & Bittencourt, M. M. (1991). Impacto de hidroelétricas sobre a ictiofauna amazônica: O exemplo de Tucuruí. In A. L. Val, R. Figiuolo, & E. Feldberg (Eds.), Bases Científicos para Estratégias de Preservação e Desenvolvimento da Amazônia: Fatos e Perspectivas (85-100). Manaus: Instituto Nacional de Pesquisas da Amazônia (INPA). [ Links ]
Lewis, S. L., Phillips, O. L., Sheil, D., Vinceti, B., Baker, T. R., Brown, S., Graham, A. W., Higuchi, N., Hilbert, D. W., Laurance, W. F., Lejoly, J., Malhi, Y., Monteagudo, A., Vargas, P. N., Sonke, B., Supardi, N., Terborgh, J. W., & Martinez, R. V. (2004). Tropical forest tree mortality, recruitment and turnover rates: calculation, interpretation and comparison when census intervals vary. Journal of Ecology, 92(6), 929-944. [ Links ]
Lilliefors, H. (1967). On the Kolmogorov-Smirnov test for normality with mean and variance unknown. Journal of the American Statistical Association, 62, 399-402. [ Links ]
Loreau, M., Naeem, S., Inchausti, P., Bengtsson, J., Grime, J. P., Hector, A., Hooper, D. U., Huston, M. A., Raffaelli, D., Schmid, B., Tilman, D., & Wardle, D. A. (2001). Ecology - Biodiversity and ecosystem functioning: Current knowledge and future challenges. Science, 294(5543), 804-808. [ Links ]
Luz, S. L. B. (1994). Estudo sobre os culicíneos (diptera: culicidar:culicinar) crepusculares e noturnos, vetores potenciais de patógenos de importância médica e veterinária na Estação Ecológica de Samuel-Rondônia. Rio de Janeiro: Universidade Federal Rural do Rio de Janeiro. [ Links ]
Machado, E. L. M., & Oliveira-Filho, A. T. (2010). Spatial patterns of tree community dynamics are detectable in a small (4 ha) and disturbed fragment of the Brazilian Atlantic forest. Acta Botanica Brasilica, 24(1), 250-261. [ Links ]
Malhi, Y., Baker, T. R., Phillips, O. L., Almeida, S., Alvarez, E., Arroyo, L., Chave, J., Czimczik, C. I., Di Fiore, A., Higuchi, N., Killeen, T. J., Laurance, S. G., Laurance, W. F., Lewis, S. L., Montoya, L. M. M., Monteagudo, A., Neill, D. A., Vargas, P. N., Patino, S., Pitman, N. C. A., Quesada, C. A., Salomao, R., Silva, J. N. M., Lezama, A. T., Martinez, R. V., Terborgh, J., Vinceti, B., & Lloyd, J. (2004). The above-ground coarse wood productivity of 104 Neotropical forest plots. Global Change Biology, 10(5), 563-591. [ Links ]
Mallik, A. U., & Richardson, J. S. (2009). Riparian vegetation change in upstream and downstream reaches of three temperate rivers dammed for hydroelectric generation in British Columbia, Canada. Ecological Engineering, 35(5), 810-819. [ Links ]
Marin, G. C., Nygard, R., Rivas, B. G., & Oden, P. C. (2005). Stand dynamics and basal area change in a tropical dry forest reserve in Nicaragua. Forest Ecology and Management, 208(1), 63-75. [ Links ]
Miles, L., Newton, A. C., DeFries, R. S., Ravilious, C., May, I., Blyth, S., Kapos, V., & Gordon, J. E. (2006). A global overview of the conservation status of tropical dry forests. Journal of Biogeography, 33(3), 491-505. [ Links ]
Lugo, A. E. (1986). Ecology of Tropical Dry Forest. Annual Review of Ecology and Systematics, 17, 67-88. [ Links ]
Nishyiama, L. (1989). Geologia do município de Uberlândia e áreas adjacentes. Sociedade e Natureza, 1(1), 9-16. [ Links ]
Nilsson, C. (1983). Frequency-distributions of vascular plants in the geolittoral vegetation along 2 rivers in northern Sweden. Journal of Biogeography, 10(4), 351-369. [ Links ]
Nilsson, C. (1996). Remediating river margin vegetation along fragmented and regulated rivers in the North: What is possible? Regulated Rivers-Research & Management, 12(4), 415-431. [ Links ]
Nilsson, C., & Berggren, K. (2000). Alterations of riparian ecosystems caused by river regulation. Bioscience, 50(9), 783-792. [ Links ]
Nilsson, C., & Svedmark, M. (2002). Basic principles and ecological consequences of changing water regimes: Riparian plant communities. Environmental Management, 30(4), 468-480. [ Links ]
Nilsson, C., Ekblad, A., Gardfjell, M., & Carlberg, B. (1991). Long-term effects of river regulation on river margin vegetation. Journal of Applied Ecology, 28(3), 963-987. [ Links ]
Nilsson, C., Jansson, R., & Zinko, U. (1997). Long-term responses of river-margin vegetation to water-level regulation. Science, 276(5313), 798-800. [ Links ]
Nilsson, C., Reidy, C. A., Dynesius, M., & Revenga, C. (2005). Fragmentation and flow regulation of the world’s large river systems. Science, 308(5720), 405-408. [ Links ]
Oliveira, A. T., Curi, N., Vilela, E. A., & Carvalho, D. A. (1997). Tree species distribution along soil catenas in a riverside semideciduous forest in southeastern Brazil. Flora, 192(1), 47-64. [ Links ]
Oliveira-Filho, A. T., Carvalho, W. A. C., Machado, E. L. M., Higuchi, P., Appolinário, V., Castro, G. C., Silva, A. C., Santos, R. M., Borges, L. F., Corrêa, B. S., & Alves, J. M. (2007). Dinâmica da comunidade e populações arbóreas da borda e interior de um remanescente florestal na Serra da Mantiqueira, Minas Gerais, em um intervalo de cinco anos (1999-2004). Revista Brasileira de Botânica, 30(1), 149-161. [ Links ]
Oliveira-Filho, A. T., & Ratter, J. A. (2002). Vegetation Physiognomies and Woody Flora of the Cerrado Biome. In P. S. liveira & R. J. Marquis (Eds.), The Cerrados of Brazil. New York: Columbia University Press. [ Links ]
Paiva, L. V., Araújo, G. M., & Pedroni, F. (2007). Structure and dynamics of a wood plant community of a tropical semi-deciduous seasonal forest in the “Estação Ecológica do Panga”, municipality of Uberlândia, Minas Gerais, Brazil. Revista Brasileira de Botânica, 30(3), 365-373. Retrieved from http://www.scielo.br/pdf/rbb/v30n3/03.pdf. [ Links ]
Patz, J. A., Graczyk, T. K., Geller, N., & Vittor, A. Y. (2000). Effects of environmental change on emerging parasitic diseases. International Journal for Parasitology, 30(12), 1395-1405. [ Links ]
Pearson, T. R. H., Burslem, D., Goeriz, R. E., & Dalling, J. W. (2003). Regeneration niche partitioning in neotropical pioneers: effects of gap size, seasonal drought and herbivory on growth and survival. Oecologia, 137(3), 456-465. [ Links ]
Pennington, R. T., Lavin, M., & Oliveira-Filho, A. (2009). Woody Plant Diversity, Evolution, and Ecology in the Tropics: Perspectives from Seasonally Dry Tropical Forests. Annual Review of Ecology Evolution and Systematics, 40, 437-457. [ Links ]
Phillips, O. L., Baker, T. R., Arroyo, L., Higuchi, N., Killeen, T. J., Laurance, W. F., Lewis, S. L., Lloyd, J., Malhi, Y., Monteagudo, A., Neill, D. A., Vargas, P. N., Silva, J. N. M., Terborgh, J., Martinez, R. V., Alexiades, M., Almeida, S., Brown, S., Chave, J., Comiskey, J. A., Czimczik, C. I., Di Fiore, A., Erwin, T., Kuebler, C., Laurance, S. G., Nascimento, H. E. M., Olivier, J., Palacios, W., Patino, S., Pitman, N. C. A., Quesada, C. A., Saldias, M., Lezama, A. T., & Vinceti, B. (2004). Pattern and process in Amazon tree turnover, 1976-2001. Philosophical Transactions of the Royal Society of London Series B-Biological Sciences, 359(1443), 381-407. [ Links ]
Pregitzer, K. S., & Euskirchen, E. S. (2004). Carbon cycling and storage in world forests: biome patterns related to forest age. Global Change Biology, 10(12), 2052-2077. [ Links ]
Raherison, S. M., & Grouzis, M. (2005). Plant biomass, nutrient concentration and nutrient storage in a tropical dry forest in the south-west of Madagascar. Plant Ecology, 180(1), 33-45. [ Links ]
Reichstein, M., Tenhunen, J. D., Roupsard, O., Ourcival, J. M., Rambal, S., Miglietta, F., Peressotti, A., Pecchiari, M., Tirone, G., & Valentini, R. (2002). Severe drought effects on ecosystem CO(2) and H(2)O fluxes at three Mediterranean evergreen sites: revision of current hypotheses? Global Change Biology, 8(10), 999-1017. [ Links ]
Ribeiro, J. F., & Walter, B. M. T. (2001). As matas de galeria no contexto do bioma Cerrado. In J. F. Ribeiro, C. E. L. Fonseca, & J. C. Sousa-Silva (Eds.), Cerrado: caracterização e recuperação de Matas de Galeria (29-45). Planaltina: Embrapa Cerrados. [ Links ]
Rodrigues, R. R., & Nave, A. G. (2000). Heterogeneidade florística das matas ciliares. In R. R. Rodrigues, & H. F. Leitão-Filho (Eds.), Matas ciliares: conservação e recuperação (45-71). São Paulo: EDUSP. [ Links ]
Rodrigues V. H. P., Lopes, S. F., Araújo G. M., & Schiavini, I. (2010). Composição, estrutura e aspéctos ecológicos da floresta ciliar do rio Araguari no Triângulo Mineiro. Hoehnea, 37, 87-105. [ Links ]
Runyon, J, Waring, R. H., Goward, S. N., & Welles, J. M. (1994). Environmental limits on net primary production and light-use efficiency across the Oregon Transect. Ecological Applications, 4(2), 226-237. [ Links ]
Santos, E. R., & Assunção, W. L. (2006). Distribuição espacial das chuvas na microbacia do Córrego do Amanhece, Araguari-MG. Caminhos da Geografia, 6, 41-55. [ Links ]
Sheil, D., Burslem, D., & Alder, D. (1995). The interpretation and misinterpretation of mortality-rate measures. Journal of Ecology, 83(2), 331-333. [ Links ]
Sheil, D., Jennings, S., & Savill, P. (2000). Long-term permanent plot observations of vegetation dynamics in Budongo, a Ugandan rain forest. Journal of Tropical Ecology, 16, 765-800. [ Links ]
Silva, M. R., & Araújo, G. M. (2009). Dinâmica da comunidade arbórea de uma floresta semidecidual em Uberlândia, MG, Brasil. Acta Botanica Brasilica, 23(1), 49-56. [ Links ]
Siqueira, A. S., Araújo, G. M., & Schiavini, I. (2009). Estrutura do componente arbóreo e características edáficas de dois fragmentos de floresta estacional decidual no vale do rio Araguari, MG, Brasil. Acta Botanica Brasilica, 23(1), 10-21. [ Links ]
Soumis, N., Duchemin, E., Canuel, R., & Lucotte, M. (2004). Greenhouse gas emissions from reservoirs of the western United States. Global Biogeochemical Cycles, 18(3), 1-18. [ Links ]
Stefan, J., McDonald, A. J., Johnson, M. S., Feldpaush, T. R., Couto, E. G., & Riha, S. J. (2007). Relationships between soil hydrology and forest structure and composition in the southern Brazilian Amazon. Journal of Vegetation Science, 18(2), 183-194. [ Links ]
Stephenson, N. L., & van Mantgem, P. J. (2005). Forest turnover rates follow global and regional patterns of productivity. Ecology Letters, 8(5), 524-531. [ Links ]
Suarez, M. L, Ghermandi, L., & Kitzberger, T. (2004). Factors predisposing episodic drought-induced tree mortality in Nothofagus - site, climatic sensitivity and growth trends. Journal of Ecology, 92(6), 954-966. [ Links ]
Swaine, M. D., Lieberman, D., & Hall, J. B. (1990). Structure and dynamics of a tropical dry forest in Ghana. Vegetatio, 88(1), 31-51. [ Links ]
Toniato, M. T. Z., & Oliveira-Filho, A. T. (2004). Variations in tree community composition and structure in a fragment of tropical semideciduous forest in southeastern Brazil related to different human disturbance histories. Forest Ecology and Management, 198(1), 319-339. [ Links ]
Truffer, B., Bratrich, C., Markard, J., Peter, A., Wuest, A., & Wehrli, B. (2003). Green Hydropower: The contribution of aquatic science research to the promotion of sustainable electricity. Aquatic Sciences, 65(2), 99-110. [ Links ]
Tufekcioglu, A., Raich, J. W., Isenhart, T. M., & Schultz, R. C. (1998). Fine root dynamics, coarse root biomass, root distribution, and soil respiration in a multispecies riparian buffer in Central Iowa, USA. Agroforestry Systems, 44(2), 163-174. [ Links ]
van Breugel, M., Martinez-Ramos, M., & Bongers, F. (2006). Community dynamics during early secondary succession in Mexican tropical rain forests. Journal of Tropical Ecology, 22, 663-674. [ Links ]
Wilkinson, L. (2002). Systat software 10.2. Chicago. [ Links ]
*Correspondencia a:
Vagner Santiago do Vale: Uberlândia Federal University, Laboratory of Plant Ecology. Uberlândia, Minas Gerais, Brazil. CEP: 38.400-902, Uberlândia-MG, Brazil; vsvale@hotmail.com
Ivan Schiavini: Uberlândia Federal University, Laboratory of Plant Ecology. Uberlândia, Minas Gerais, Brazil. CEP: 38.400-902, Uberlândia-MG, Brazil; ivanschiavini@gmail.com
Glein Monteiro Araújo: Uberlândia Federal University, Laboratory of Plant Ecology. Uberlândia, Minas Gerais, Brazil. CEP: 38.400-902, Uberlândia-MG, Brazil; glein@ufu.br
Ana Paula de Oliveira: Uberlândia Federal University, Laboratory of Plant Ecology. Uberlândia, Minas Gerais, Brazil. CEP: 38.400-902, Uberlândia-MG, Brazil; anadeoli@gmail.com
Jamir Afonso do Prado-Júnior: Uberlândia Federal University, Laboratory of Plant Ecology. Uberlândia, Minas Gerais, Brazil. CEP: 38.400-902, Uberlândia-MG, Brazil; jamirjunior@yahoo.com.br
Carolina de Silvério Arantes: Uberlândia Federal University, Laboratory of Plant Ecology. Uberlândia, Minas Gerais, Brazil. CEP: 38.400-902, Uberlândia-MG, Brazil; carolina.s.arantes@gmail.com
André Eduardo Gusson: Lutheran University of Brazil, Laboratory of Botany. Itumbiara, Goiás, Brazil. CEP: 75.522-100, Uberlândia - MG, Brazil; desrp4@hotmail.com
Sérgio de Faria Lopes: Paraiba State University, Laboratory of Plant Ecology. Campina Grande, Paraiba, Brazil. CEP: 58.429-500, Campina Grande - PB, Brazil; defarialopes@gmail.com
Olavo Custódio Dias-Neto: Mario Palmério Carmelitan Foundation, Institute of Biology. Monte Carmelo, Minas Gerais, Brazil. CEP: 38 500-000, Monte Carmelo - MG, Brazil; olavonneto@hotmail.com
1.Uberlândia Federal University, Laboratory of Plant Ecology. Uberlândia, Minas Gerais, Brazil. CEP: 38.400-902, Uberlândia-MG, Brazil; vsvale@hotmail.com Corresponding author, ivanschiavini@gmail.com, glein@ufu.br, anadeoli@gmail.com, jamirjunior@yahoo.com.br, carolina.s.arantes@gmail.com
2. Lutheran University of Brazil, Laboratory of Botany. Itumbiara, Goiás, Brazil. CEP: 75.522-100, Uberlândia - MG, Brazil; desrp4@hotmail.com
3. Paraiba State University, Laboratory of Plant Ecology. Campina Grande, Paraiba, Brazil. CEP: 58.429-500, Campina Grande - PB, Brazil; defarialopes@gmail.com
4. Mario Palmério Carmelitan Foundation, Institute of Biology. Monte Carmelo, Minas Gerais, Brazil. CEP: 38 500-000, Monte Carmelo - MG, Brazil; olavonneto@hotmail.com
Received 29-V-2012. Corrected 20-VI-2013. Accepted 22-VII-2013.


{kind=link}
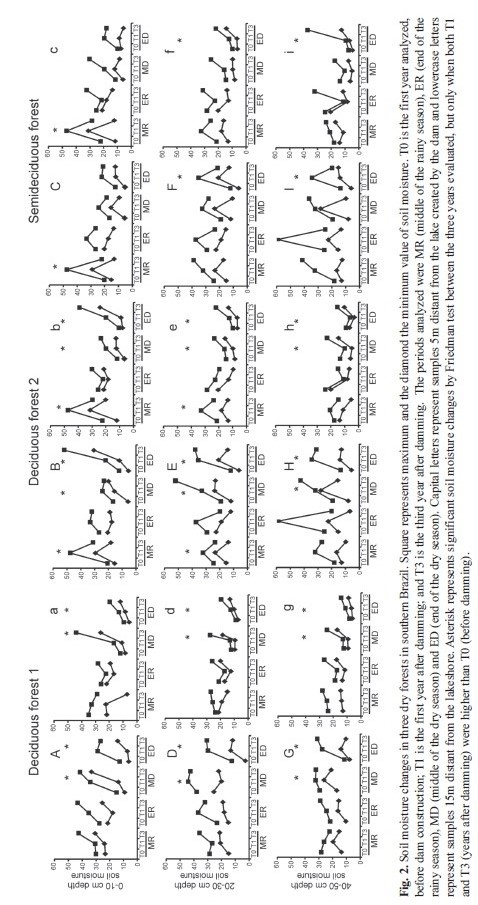{kind=link}
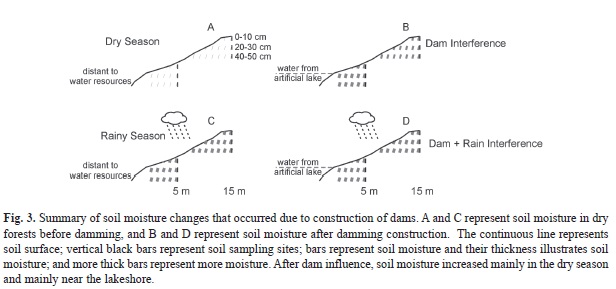{kind=link}
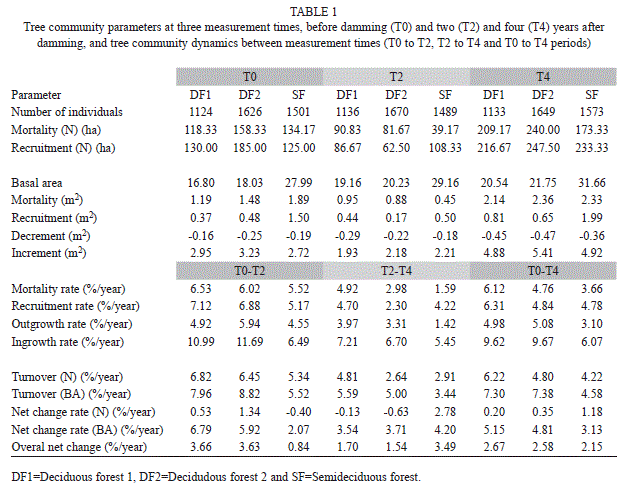{kind=link}
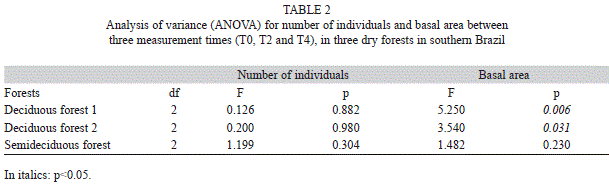{kind=link}
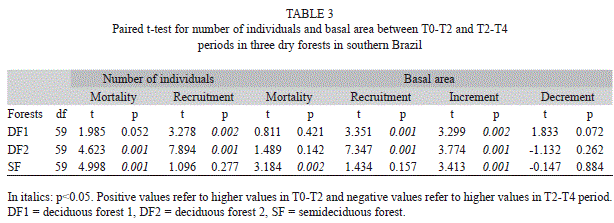{kind=link}
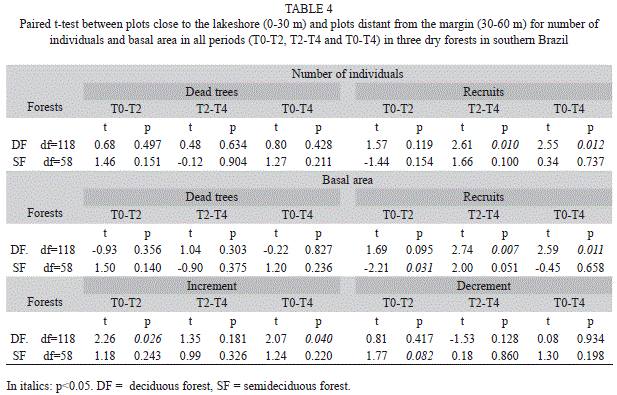{kind=link}