










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.61 n.4 San José Oct./Dec. 2013
Synchronism between Aspidosperma macrocarpon (Apocynaceae) resources allocation and the establishment of the gall inducer Pseudophacopteron sp. (Hemiptera: Psylloidea)
Sincronismo de Aspidosperma macrocarpon (Apocynaceae) entre la asignación de recursos y el establecimiento del inductor de agallas Pseudophacopteron sp. (Hemiptera: Psylloidea)
Sincronismo de Aspidosperma macrocarpon (Apocynaceae) entre la asignación de recursos y el establecimiento del inductor de agallas Pseudophacopteron sp. (Hemiptera: Psylloidea)
*Dirección de correspondencia:
Abstract
The joint interpretation of phenology and nutritional metabolism provides important data on plant tissues reactivity and the period of gall induction. A population of Aspidosperma macrocarpon (Apocynaceae) with leaf galls induced by a Pseudophacopteron sp. (Psylloidea) was studied in Goiás state, Brazil. Assuming the morphological similarity between host leaves and intralaminar galls, a gradient from non-galled leaves towards galls should be generated, establishing a morpho-physiological continuum. The phenology, infestation of galls, and the carbohydrate and nitrogen contents were monthly evaluated in 10-20 individuals, from September 2009 to September 2010. Our objective was to analyze the nutritional status and the establishment of a physiological continuum between the galls and the non-galled leaves of A. macrocarpon. The period of leaf flushing coincided with the highest levels of nitrogen allocated to the new leaves, and to the lowest levels of carbohydrates. The nutrients were previously consumed by the growing leaves, by the time of gall induction. The levels of carbohydrates were higher in galls than in non-galled leaves in time-based analyses, which indicateed their potential sink functionality. The leaves were infested in October, galls developed along the year, and gall senescence took place from March to September, together with host leaves. This first senescent leaves caused insect mortality. The higher availability of nutrients at the moment of gall induction was demonstrated and seems to be important not only for the establishment of the galling insect but also for the responsiveness of the host plant tissues.
Key words: carbohydrates, continuum, insect-plant interaction, leaf flushing, nitrogen, reactive tissues, vegetative phenology.
Resumen
La interpretación conjunta de la fenología y el metabolismo nutricional proporciona datos importantes sobre la reacción de tejidos vegetales y el período de inducción de agallas. En el estado de Goiás, Brasil se estudió una población de Aspidosperma macrocarpon (Apocynaceae) con agallas foliares inducidas por un Pseudophacopteron sp. (Psylloidea). Suponiendo que la similitud morfológica entre las hojas del hospedero y las agallas intralaminares genera un gradiente de hojas sin agallas hacia hojas con agallas, se establece un continuo morfo-fisiológico. La fenología, la infestación de las agallas y el contenido de carbohidratos y nitrógeno se evaluaron mensualmente en 10-20 individuos, de septiembre 2009 a septiembre 2010. El objetivo fue analizar el estado nutricional y el establecimiento de un continuo fisiológico entre las hojas que producen agallas y las hojas sin agallas de A. macrocarpon. El período de emergencia de hojas coincidió con los niveles más altos de nitrógeno en las hojas nuevas, y los niveles más bajos de carbohidratos.Los nutrientes se consumieron previamente por las hojas en crecimiento, en el momento de inducción de las agallas. Los niveles de carbohidratos fueron más altos en las hojas con agallas que en aquellas sin agallas, en análisis basados en tiempo, lo que indica su ciclo de funcionalidad potencial. Las hojas hospederas se infestaron en octubre, el desarrollo y la senescencia de las agallas se llevó a cabo entre marzo y septiembre, en las hojas hospederas. Estas primeras hojas senescentes causaron mortalidad de insectos. Hay mayor disponibilidad de nutrientes en el momento de la inducción de las agallas y parece ser importante no sólo para el establecimiento del insecto que produce agallas, sino también para la capacidad de respuesta de los tejidos de la planta hospedera.
Palabras clave: carbohidratos, continuo, interacción insecto-planta, emergencia de hojas, nitrógeno, tejido reactivo, fenología vegetativa.
Gall-inducing insects are sensitive to several chemical, physiological and phenological plant modifications (Fernandes & Price, 1992; Campos, Costa, Isaias, Moreira, Oliveira & Lemos- Filho, 2010; Oliveira, Mendonça, Moreira, Lemos-Filho & Isaias, 2012). Variations on climatic conditions and water availability affect the host plant phenology, and could therefore alter the life cycle and the distribution of galling insects as demonstrated by Oliveira et al. (2012) for Copaifera langsdorffii (Fabaceae)-Cecidomyiidae system where low water potential was followed by an increase in galling richness. Evaluations of the abundance of galls and host plant phenology throughout one-year cycle provide information on the synchronism between the behavior of the gall inducer and the nutritional status of host plant tissues (Campos et al., 2010). This synchronism may be crucial to determine the abundance of individuals (Yukawa, 2000), and the fitness of the galling herbivores (Oliveira et al., 2012).
The adjustment of the different phenophases among host plant individuals in a population may be related to their endogenous characteristics and environmental variables. The abiotic factors can influence plant physiology, and determine or restrict the period of occurrence of each plant phenophase (Bencke & Morellato, 2002; Pedroni, Sanchez & Santos, 2002). In the interaction with galling herbivores, the phenological analysis may be used to test the nutritional hypothesis, proposed by Price, Waring & Fernandes (1986, 1987). This hypothesis states that the gall is a strong sink of nutrients during its development. Consequently, the nutrients accumulated can either nourish the herbivore or maintain the cellular machinery of the gall (Oliveira, Christiano, Soares & Isaias, 2006; Moura, Soares & Isaias, 2008; Oliveira & Isaias, 2010). The nutritional status of plant tissues in arthropod-induced galls is quite interesting for the herbivores, and may have a substantial impact on carbon partitioning mechanisms within the proper host plants (Inbar, Eshel & Wool, 1995). The presence of galling herbivores within the host plant cells modify the balance between source tissues, responsible for photosynthesis, and the sink ones, the primary destiny of the photoassimilates (Dorchin, Cramer & Hoffmann, 2006). The alterations in the source-sink relationships are particularly evidenced in the tissues close to the gall, leading to the accumulation of carbohydrates and nitrogen (Hartley, 1998; Castro, Oliveira, Moreira, Lemos-Filho & Isaias, 2012).
The phenological studies of interactions of Aspidosperma spp. (Apocynaceae) with their galling herbivores started with Campos et al. (2010), who characterized the physiological impacts of two distinct taxa of insects on leaves of A. australe Müell. Arg. and of A. spruceanum Benth. ex Müell. Arg. Currently, we propose phenological analysis associated with the evaluation of the nutritional status of plant tissues as a tool to access the developmental patterns in galling herbivores-host plant systems, specially increasing the knowledge on the interactions within the Aspidosperma genus. It is proposed that the life cycle of the galling insect (Pseudophacopteron sp.) is related to the host plant phenology on Aspidosperma macrocarpon Mart., and that the levels of infestation of galls coincide with the greater availability of nutrients in host plant tissues. Moreover, assuming the morphological similarity between host leaves and galls in this system, a gradient from non-galled leaves, through non-galled portions of galled leaves, and towards galls should be generated, establishing a morpho-physiological continuum. Based on these premises, this study aims to answer the following questions: (1) does leaf flushing, i.e., the time when there are more plant reactive tissues, favor gall induction? (2) Is the nutritional status of the host plant tissues related to the infestation index of galls? And (3) is there a nutritional continuum from the non-galled leaves through galled ones towards gall sites?
Material and Methods
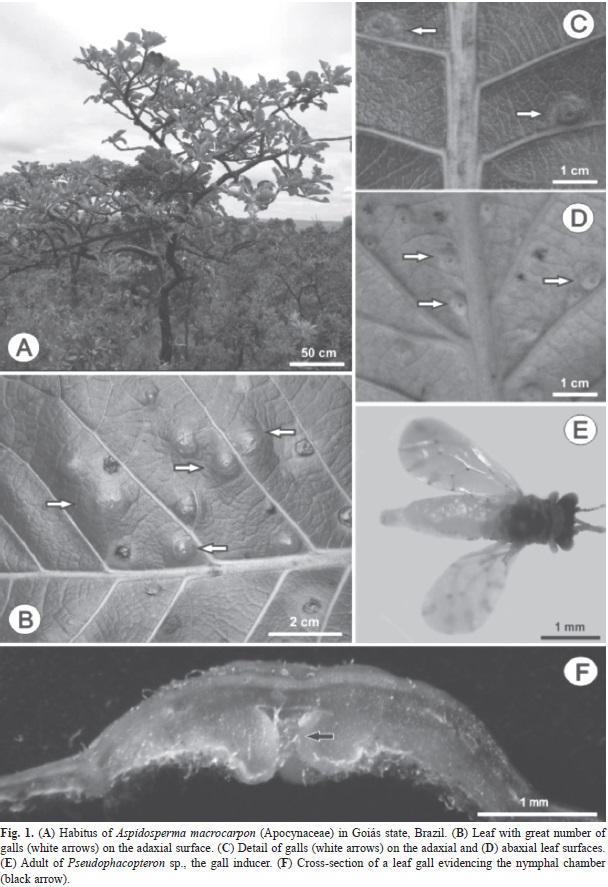
Studied area and plant-insect system: Aspidosperma macrocarpon Mart. (Apocynaceae) is a woody tree, approximately 3m high, with characteristic sympodial growth (Fig. 1A), native of North, Northeast, Middle-West, and Southeast regions of Brazil, and Amazonian Cerrado (Forzza et al., 2010). The leaves are 30cm in length, with an average of 18.1 (SD=14.6, n=40) galls per leaf (Fig. 1B). Mature galls are green, intralaminar, and slightly protruding to the adaxial leaf surface (Fig. 1C) even though it is induced on the abaxial surface (Fig. 1D) by the feeding action of the first-instar nymphal of Pseudophacopteron sp. (Hemiptera: Psylloidea) (Fig. 1E). The nymphal chamber is unique and assumes a central position (Fig. 1F).
Individuals were randomly marked in a population at the Reserva Biológica Prof. José Angelo Rizzo - Serra Dourada (16°22’-15°48’S - 50°44’-49°55’W, altitudes above 700m) in Goiás state, Brazil. The area is subject to two definite seasons: the dry season that lasts from April to September and the rainy season beginning in October, with periods of increased precipitation from November to March (Rizzo, 1970), and annual precipitation around 450mm (INMET 1961-2010). The vegetation comprises the rupestrian Cerrado (Rizzo, 1970), characterized by trees and shrubs over rocky soils (Oliveira & Marquis, 2002).
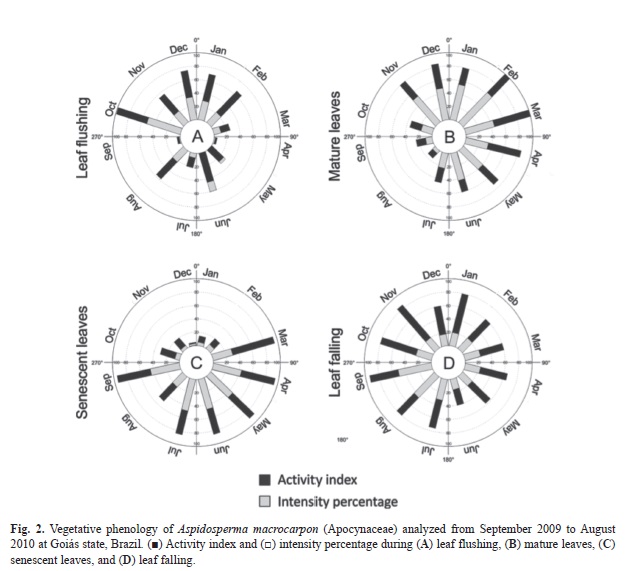
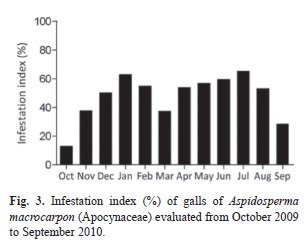
Host plant vegetative phenology and infestation of galls: For the evaluation of host plant phenology, the sampling followed the model suggested by Fournier & Charpantier (1975) in which 20 adult individuals were selected in order of appearance within the plot. The phenological observations were carried out twice a month, from September 2009 to August 2010, registering the vegetative characteristics, herein referred as leaf flushing, mature leaves, senescent leaves and leaf falling. The activity index is a qualitative measure that reveals the absence or presence of a specific phase, its beginning and end, and allows estimating synchrony among individuals in a population (Morellato, Leitão Filho, Rodrigues & Joly, 1990; Bencke & Morellato, 2002). To determine the intensity percentage or index of Fournier (1974), i.e., to quantify the intensity of the phenophase on the individual, phenological phases were recorded on a scale of categories based on the percentage of canopy cover (Bencke & Morellato, 2002). This semiquantitative method ranks the phenophase in five categories: (0) represents the absence of the phenophases, (1) 1-25% of the phenophases, (2) 26-50%, (3) 51-75%, and (4) 76-100%. The leaf flushing was considered from the time the buds reached 0.5cm until unfolding of the leaves (Pedroni et al., 2002).The infestation index was calculated as: (G x 100)/T, where G=number of galled leaves and T=total number of leaves. The data were plotted in the softwares Oriana©4.01 (Kovach Computing Services 1994-2012) and Graphpad Prism® 5.0 (GraphPad Software, Inc. 1992-2009).
Galls were divided in three stages of development based on the average of height versus diameter and the presence of the gall inducer. Young (1.8mm SD=1.1x3.9mm SD=1.0, n=88), mature (1.3mm SD=0.2x5.3mm SD=1.5, n=64), and senescent galls (with no gall inducer) (2.3mmSD=0.3x4.7mm SD=1.0, n=26), were collected and dissected under a stereomicroscope (Zeiss®Stemi 2000-C). The insects were fixed in 70% ethanol and sent to Professor Daniel Burckhardt (Naturhistorisches Museum, Basel, Switzerland) for taxonomical identification.
Dosage of nitrogen and carbohydrates: In order to detect the periods of increased nutrients availability (nitrogen and carbohydrates), non-galled and galled leaves (n=10) from 10 individuals were collected monthly from October 2009 to September 2010. Three different samples were considered: the non-galled leaves (NGL), the galls (G), and the non-galled portions of galled leaves (PGL). Analyses of nitrogen and carbohydrates were performed throughout one year and the comparison among the samples (NGL, PGL and G) was taken as an average value for each trimester.
The nitrogen content calculation was based on the Kjeldahl method (Tedesco, Gianello, Bissani, Bohnen & Volkweiss, 1995) using plant material dried in an oven of forced circulation at 50°C for 48 hours. The samples of NGL, PGL and G were ground in a mortar, and 250mg of dry mass (DM) of each sample were submitted to the sulfuric digestion. Later on, the material was submitted to distillation process by steam distillation (distiller Tecnal TE-0363) where the ammonia released in the form of NH4OH was trapped in 2% boric acid, and titrated against standardized 0.02N hydrochloric acid.
For carbohydrates determination, samples of NGL, PGL and G were treated in a microwave oven for 30s (Marur & Sodek, 1995), dried at 50°C for 48 hours, and ground in a microgrounder Willye TE-648. The extraction of total soluble sugars (TSS) was performed in a solution of methanol: chloroform: water (12:5:3 v/v) (Bielski &Turner, 1966).The content of water-soluble polysaccharides (WSP) was obtained by resuspending the residue in 10% ethanol (Shannon, 1968). A new extract obtained from the pellet resuspension in 30% perchloric acid (McCready, Guggolz, Silveira & Owens, 1950) was taken to measure the content of starch. The dosage was performed in triplicate sampling by colorimetric analysis using the phenol-sulfuric acid method (Dubois, Gilles, Hamilton, Rebers & Smith, 1956) modified by Chow & Landhaüsser (2004) using glucose as standard.
The nutrient data were submitted to statistical analysis of variance by ANOVA, Shapiro-Wilk normality test, comparison of means by Tukey test for parametric analyses, and Kruskal-Wallis and Dunn’s tests for non-parametric ones on GraphPad Prism® 5.0 (GraphPad Software, Inc. 1992-2009).
Results
Host plant phenological features and gall infestation: There were two peaks of leaf flushing in October and June 2010 (Fig. 2A). During this phenophase, the rates of flushing fluctuated with low synchronism, except in October, when 100% of the population was sprouting. Mature leaves were observed all over the year with the greatest occurrence in February-March 2010 (Fig. 2B). The senescent leaves were almost absent in December, and were observed in over 90% of the individuals in March-July (Fig. 2C). Leaf falling also occurred all over the year with a peak in August-September 2009 (Fig. 2D).
The infestation index along the year of evaluation revealed two peaks, one in January (62.8%) and another in July (65.2%) (Fig. 3). The increase of infestation index occurred just when there was a peak of leaf flushing (Fig. 2A and Fig. 3).
Host plant and gall nutritional status: The nitrogen and carbohydrate contents (TSS and WSP) slightly varied throughout the period of study. In general, the nitrogen content reduced from January-March to July-September in all the samples (Table 1), contrasting to the TSS and WSP, which increased from January to September (Table 2). The starch content did not vary (Table 2).
Comparing non-galled leaves (NGL), non-galled portions of galled leaves (PGL), and galls (G), the nitrogen content did not vary throughout the year, except in October-December, when the contents in PGL were higher than in NGL (Table 1). The contents of carbohydrates were usually constant among the samples, but demonstrated a tendency to a higher accumulation in galls in time-based analyses. From January to March, galls accumulated either more WSP or starch, while from July to September, they accumulated TSS and starch (Table 2).
Discussion
The peak of leaf flushing observed in October in the population of A. macrocarpon favored gall induction due to the availability of responsive tissues on host plants. The relation between the responsiveness of host tissues and gall induction has been proposed by Weis, Walton & Crego (1988) and Rohfritsch (1992), and it was proved in Rollinia laurifolia (Annonaceae)-Pseudotectococcus rolliniae (Eriococcidae) (Gonçalves, Isaias, Vale & Fernandes, 2005; Gonçalves, Moreira & Isaias, 2009), and Copaifera langsdorffii-Cecidomyiidae (Oliveira & Isaias, 2009) systems. As well, Campos et al. (2010) and Oliveira et al. (2012) connected gall induction with leaf flushing in A. australe-Pseudophacopteron sp., and in Copaifera langsdorffii-multiple Cecidomyiidae systems. Moreover, the increase of infestation occurred from October to January in the A. macrocarpon-Pseudophacopteron sp. system coincided with leaf flushing and maturation. These results corroborate Weis’ hypothesis.
The peak of leaf falling occurred in July-September with more than 50% of activity index. However, leaf falling continued throughout the year, indicating that A. macrocarpon in Serra Dourada has lost and replaced leaves more than once a year. However, Pirani, Sanchez & Pedroni (2009) had previously classified the species in Mato Grosso state, Brazil, as deciduous, with leaf falling occurring from July to September, and leaf flushing in October and November. These data indicated that some populations of A. macrocarpon may be semi-deciduous, remaining with both young and mature leaves throughout the year. This dissimilarity may be caused by environmental influences (Pedroni et al., 2002), or by endogenous characteristics (Bencke & Morelatto, 2002).
During leaf flushing, the intensity percentage could exceed the index of activity, indicating that a few plants were at this phenophase, but each one had a great amount of new leaves. This phenological peculiarity can be explained by the phenological complexity presented in tropical systems, as already reported by Bencke & Morellato (2002). The activity index detected no significant synchrony in leaf flushing among individuals, because only in October the whole population presented the emergence of new leaves. However, even in this case, 50% of the canopy was occupied with young leaves that associated to the constant presence of mature leaves, indicated the semi-deciduous behavior of the population. The percentage of young leaves was around 30% over the period of study, showing that maturation occurred gradually, a fact corroborated by the percentage of mature leaves.
The highest level of infestation coincided with the best nutritional status of the host plant tissues of A. macrocarpon based on their nitrogen content. The content of this nutritional compound was higher in all samples (NGL, PGL and G) by the time of leaf flushing, when plant tissues were generally nitrogen rich, as for Copaifera langsdorffii system (Castro et al., 2012). Throughout the year, the nitrogen content decreased probably by its relocation to other plant parts during leaf senescence (Fischer, 2007). The decrease in the percentage of nitrogen observed from April on reflects the period of maturation, senescence and leaf abscission, what was previously reported by Leitão & Silva (2004) in the evaluation of mineral compartmentalization in the phenophases of Ouratea spectabilis (Mart.) Engl. (Ochnaceae) in a Cerrado area.
The tendency to a higher accumulation of carbohydrates in galls in time-based analysis was related to the dynamics of gall development as reported by Marmit & Sharma (2008) on Mangifera indica (Anacardiaceae). The TSS accumulated as a source of energy for future investment in leaf sprouting and gall development. Moreover, the carbohydrates were readily consumed during leaf growth and gall development, preventing its accumulation in detectable levels. Similarly to the observed in the months of leaf flushing of A. macrocarpon, Dantas, Ribeiro & Pereira (2007) observed low levels of TSS in young leaves of the grapevine ‘Syrah’. The galls on A. macrocarpon do not seem to function as strong sinks like the horn-shaped gall on C. langsdorffii, which has higher mass and accumulates high amounts of nutrients (Castro et al., 2012). Nevertheless, their levels of nutrients can confirm not only that the gall is an adaptation to improve feed quality for the galling herbivore but also for the maintenance of gall metabolism.
The higher levels of WSP in galls in July-September denotes the restructuring of the quantity and even the quality of pectins (Wolf & Greiner, 2012) during the maturation of galls, which represent new organs where tissues with distinct functions need to be redifferentiated (sensu Lev-Yadun, 2003). The accumulation of starch on host tissues of A. macrocarpon did not vary either on time-bases or within the samples except for the gall tissues in January-March and in July-September. Carbohydrates have been drained to the gall during its development and accumulated as starch grains.
As proposed by Dreger-Jauffret & Shorthouse (1992), galling insects are unique in controlling the within-plant movement of photoassimilates and tissue morphogenesis. The nutritional analyses of galled together with its neighboring tissues in A. macrocarpon revealed a continuum gradient of carbohydrates towards gall sites. This gradient slightly fluctuated in relation to the host plant phenological phase, mainly by the time of leaf flushing. Furthermore, the nutritional status of host plant tissues favors the induction and the establishment of the insect gall, and also the maintenance of host plant cells.
Our analyses led us to the conclusion that the host nutritional traits are directed towards leaf flushing and development. The gall inducer presents a temporal synchronism to maximize the use of host plant resources to its own benefit. In A. macrocarpon-Pseudophacopteron sp. system, the “window of opportunity” is restrict, differently from other gall systems where the inducer can oviposit on leaves in all stages of development or in other plant organs, like shoot and flowers. The high level of nitrogen by the time of leaf flushing favors the establishment of the galling inducer. From March on, some endogenous features of the host plant triggers leaf falling which denotes a mechanism that controls the population of galling herbivore. As a univoltine insect, the beginning of leaf falling causes the interrupting of its life cycle by premature abscission of immature galls.
Acknowledgments
The authors thank Daniel Burckhardt for insects’ identification, Andre R.T. Nascimento for helping with statistics, Taiza M. Silva for field support, and CAPES, CNPq and FAPEMIG for financial support.
References
Bencke, C. S. C., & Morellato, L. P. C. (2002). Comparação de dois métodos de avaliação da fenologia de plantas, sua interpretação e representação. Revista Brasileira de Botânica, 25, 269-275. [ Links ]
Bielski, L. R., & Turner, L. A. (1966). Separation and estimation of amino acids in crude plant extracts by thin-layer electrophoresis and chromatography. Analytical Biochemistry, 17, 278-293. [ Links ]
Campos, P. T., Costa, M. C. D., Isaias, R. M. S., Moreira, A. S. F. P., Oliveira, D. C., & Lemos-Filho, J. P. (2010). Phenological relationships between two insect galls and their host plants: Aspidosperma australe and A. spruceanum (Apocynaceae). Acta Botanica Brasilica, 24(3), 727-733. [ Links ]
Castro, A. C., Oliveira, D. C., Moreira, A. S. F. P., Lemos-Filho, J. P., & Isaias, R. M. S. (2012). Source–sink relationship and photosynthesis in the horn-shaped gall and its host plant Copaifera langsdorffii Desf. (Fabaceae). South African Journal of Botany, 83, 121-126. Retrieved from http://dx.doi.org/10.1016/j.sajb.2012.08.007. [ Links ]
Chow, P. S., & Landhäusser, S. M. (2004). A method for routine measurements of total sugar and starch content in woody plant tissues. Tree Physiology, 24, 1129-1136. [ Links ]
Dantas, B. F., Ribeiro, L. S., & Pereira, M. S. (2007). Teor de açúcares solúveis e insolúveis em folha de videiras, cv. syrah, em diferentes posições no ramo e épocas do ano. Revista Brasileira de Fruticultura, 29(1), 42-47. [ Links ]
Dorchin, N., Cramer, M. D., & Hoffmann, J. H. (2006). Photosynthesis and sink activity of wasp-induced galls in Acacia pycnantha. Ecology, 87, 1781-1791. Retrieved from http://treephys.oxfordjournals.org/. [ Links ]
Dreger-Jauffret, F., & Shorthouse, J. D. (1992). Diversity of gall-inducing insects and their galls, In J. D. Shorthouse & O. Rohfritsch (Eds.), Biology of insect induced galls (pp. 8-33). Oxford: Oxford University. [ Links ]
Dubois, M., Gilles, K. A., Hamilton, J. K., Rebers, P. A., & Smith, F. (1956). Colorimetric method for determination of sugars and related substances. Analytical Chemistry, 28, 350-356. [ Links ]
Fernandes, G. W., & Price, P. W. (1992). The adaptive significance of insect gall distribution: survivorship of species in xeric and mesic habitats. Oecologia, 90, 14-20. [ Links ]
Fischer, A. (2007). Nutrient remobilization during leaf senescence. In S. Gan. (Ed.), Senescence Processes in Plants - Annual Plant Reviews (pp. 87-102). New York: Cornell University. [ Links ]
Forzza, R. C., Stehmann, J. R., Nadruz, M., Filardi, R. P. F., Costa, A., Jr. Carvalho, A. A., Peixoto, A. L., Walter, B. M. T., Bicudo, C., Moura, C. W. N., Zappi, D., Costa, D. P., Lleras, E., Martinelli, G., Lima, H. C., Prado, J., Baumgratz, J. F. A., Pirani, J. R., Sylvestre, L. S., Maia, L. C., Lohmann, L. G., Paganucci, L., Alves, M. V. S., Silveira, M., Mamede, M. C. H., Bastos, M. N. C., Morim, M. P., Barbosa, M. R., Menezes, M., Hopkins, M., Evangelista, P. H. L., Goldenberg, R., Secco, R., Rodrigues, R. S., Cavalcanti, T., & Souza, V. C. (2010). Catalogo de Plantas fungos e do Brasil, v.1. Río de Janeiro: Andrea Jakobsson Estúdio. [ Links ]
Fournier, O. L. A. (1974). Un método cuantitativo para la medición de características fenológicas en árboles. Turrialba, 24, 422-423. [ Links ]
Fournier, O. L. A., & Charpantier, C. (1975). El tamaño de la muestra y la frecuencia de las observaciones en el estudio de las características fenológicas de los árboles tropicales. Turrialba, 25, 45-48. [ Links ]
Gonçalves, S. J. M. R., Isaias, R. M. S., Vale, F. H. A., & Fernandes, G. W. (2005). Sexual dimorphism of Pseudotectococcus rolliniae Hodgson & Gonçalves 2004 (Hemiptera Coccoidea Eriococcidae) influences gall morphology on Rollinia laurifolia Schltdl. (Annonaceae). Tropical Zoology, 18, 161-169. [ Links ]
Gonçalves, S. J. M. R., Moreira, G. R. P., & Isaias, R. M. S. (2009). A unique seasonal cycle in a leaf gall-inducing insect: the formation of stem galls for dormancy. Journal of Natural History, 43, 843-854. doi: 10.1080/00222930802615690. [ Links ]
Hartley, S. E. (1998). The chemical composition of plant galls: Are level of nutrients and secondary compounds controlled by the gall former? Oecologia, 113, 492-501. [ Links ]
Inbar, M., Eshel, A., & Wool, D. (1995). Interspecific competition among phloem-feeding insects mediated by induced host-plant sinks. Ecology, 76, 1506-1515. Retrieved from http://www.jstor.org/stable/1938152. [ Links ]
Leitão, A. C., & Silva, O. A. (2004). Variação sazonal de macronutrientes em uma espécie arbórea de cerrado, na Reserva Biológica e Estação Experimental de Mogi-Guaçu, estado de São Paulo, Brasil. Rodriguésia, 55(84), 127-136. Retrieved fom http://rodriguesia.jbrj.gov.br/FASCICULOS/Rodrig55_84/85584.pdf. [ Links ]
Lev-Yadun, S. (2003). Stem cells in plants are differentiated too. Current Topics in Plant Biology, 4, 93-102. [ Links ]
Marmit, K. S., & Sharma, S. L. (2008). Quantitative estimation of some metabolites and enzymes in insect induced leaf galls of Mangifera indica. Asian Journal of Experimental Sciences, 22(3), 343-346. [ Links ]
Marur, C., & Sodek, L. (1995). Microwave drying of plant material for biochemical analysis. Revista Brasileira de Fisiologia Vegetal, 7(1), 111-114. [ Links ]
McCready, R. M., Guggolz, J., Silveira, V., & Owens, H. S. (1950). Determination of starch and amylase in vegetables. Application to peas. Analytical Chemistry, 22, 1156-1158. [ Links ]
Morellato, L. P. C., Leitão Filho, H. F., Rodrigues, R. R., & Joly, C. A. (1990). Estratégias fenológicas de espécies arbóreas em floresta de altitude na Serra do Japi, Jundiaí São Paulo. Revista Brasileira de Biologia, 50, 149-162. [ Links ]
Moura, M. Z. D., Soares, G. L. G., & Isaias, R. M. S. (2008). Species-specific changes in tissue morphogenesis induced by two arthropod leaf gallers in Lantana camara L. (Verbenaceae). Australian Journal of Botany, 56, 153-160. doi: 10.1071/BT07131 0067-1924/08/020153. [ Links ]
Oliveira, D. C., & Isaias, R. M. S. (2009). Influence of leaflet age in anatomy and possible adaptive values of the midrib gall of Copaifera langsdorffii (Fabaceae: Caesalpinoidae). Revista de Biología Tropical, 57, 293-302. Retrieved from http://www.biologiatropical.ucr.ac.cr/attachments/volumes/vol57-1-2/26-Oliveira-Copaifera%20langsdorffii.pdf. [ Links ]
Oliveira, D. C., & Isaias, R. M. S. (2010). Redifferentiation of leaflet tissues during midrib gall development in Copaifera langsdorffii (Fabaceae). South African Journal of Botany, 76(2), 239-248. doi:10.1016/j.sajb.2009.10.011. [ Links ]
Oliveira, P. S., & Marquis, R. J. (2002). The Cerrados of Brazil. New York: Columbia University Press. [ Links ]
Oliveira, D. C., Christiano, J. C. S., Soares, G. L. G., & Isaias, R. M. S. (2006). Reações de defesas químicas e estruturais de Lonchocarpus muehlbergianus Hassl. (Fabaceae) à ação do galhador Euphalerus ostreoides Crawf. (Hemiptera: Psyllidae). Revista Brasileira de Botânica, 29, 657-667. [ Links ]
(Fabaceae) and multiple galling insects: formation of seasonal patterns. Oliveira, D. C., Mendonça Jr., M. S., Moreira, A. S. F. P., Lemos-Filho, J. P., & Isaias, R. M. S. (2012). Water stress and phenological synchronism between Copaifera langsdorffiiJournal of Plant Interaction, 1-9. Retrieved from http://dx.doi.org/10.1080/17429145.2012.705339. [ Links ]
Pedroni, F., Sanchez, M., & Santos, F. A. M. (2002). Fenologia da copaíba (Copaifera langsdorffii Desf. – Leguminosae, Caesalpinioideae) em uma floresta semidecídua no sudeste do Brasil. Revista Brasileira de Botânica, 25(2), 183-194. [ Links ]
Pirani, F. R., Sanchez, M., & Pedroni, F. (2009). Fenologia de uma comunidade arbórea em cerrado sentido restrito, Barra do Garças, MT, Brasil. Acta Botanica Brasilica, 23(4), 1096-1109. [ Links ]
Price, P. W., Waring, G. L., & Fernandes, G. W. (1986). Hypotheses on the adaptive nature of galls. Proceedings of the Entomological Society of Washington, 88, 361-363. [ Links ]
Price, P. W., Fernandes, G. W., & Waring, G. L. (1987). Adaptive nature of insect galls. Environmental Entomology, 16, 15-24. [ Links ]
Rizzo, J. A. (1970). Contribuição ao conhecimento da Flora de Goiás-área na Serra Dourada. Tese em Livre Docência. Universidade Federal de Goiás, Goiânia, GO. [ Links ]
Rohfritsch, O. (1992). Patterns in gall development. In J. D. Shorthouse & O. Rohfritsch (Eds.), Biology of insect induced galls (pp. 60-86). Oxford: Oxford University. [ Links ]
Shannon, J. C. (1968). Carbon-14 distribution in carbohydrates of immature Zea mays kernels following 14CO2 treatment of intact plants. Plant Physiology, 43, 1215-1220. [ Links ]
Tedesco, M. J., Gianello, C., Bissani, C. A., Bohnen, H., & Volkweiss, S. J. (1995). Análise de solo, plantas e outros materiais. Porto Alegre: Universidade Federal do Rio Grande do Sul. [ Links ]
Weis, A. E., Walton, R., & Crego, C. L. (1988). Reactive plant tissue site and the population biology of gall makers. Annual Review of Entomology, 3, 467-486. doi: 10.1146/annurev.en.33.010188.002343. [ Links ]
Wolf, S., & Greiner, S. (2012). Growth control by cell wall pectins. Protoplasma. doi: 10.1007/s00709-011-0371-5. [ Links ]
Yukawa, J. (2000). Synchronization of gallers with host plant phenology. Population Ecology, 42, 105-13. [ Links ]
Bielski, L. R., & Turner, L. A. (1966). Separation and estimation of amino acids in crude plant extracts by thin-layer electrophoresis and chromatography. Analytical Biochemistry, 17, 278-293. [ Links ]
Campos, P. T., Costa, M. C. D., Isaias, R. M. S., Moreira, A. S. F. P., Oliveira, D. C., & Lemos-Filho, J. P. (2010). Phenological relationships between two insect galls and their host plants: Aspidosperma australe and A. spruceanum (Apocynaceae). Acta Botanica Brasilica, 24(3), 727-733. [ Links ]
Castro, A. C., Oliveira, D. C., Moreira, A. S. F. P., Lemos-Filho, J. P., & Isaias, R. M. S. (2012). Source–sink relationship and photosynthesis in the horn-shaped gall and its host plant Copaifera langsdorffii Desf. (Fabaceae). South African Journal of Botany, 83, 121-126. Retrieved from http://dx.doi.org/10.1016/j.sajb.2012.08.007. [ Links ]
Chow, P. S., & Landhäusser, S. M. (2004). A method for routine measurements of total sugar and starch content in woody plant tissues. Tree Physiology, 24, 1129-1136. [ Links ]
Dantas, B. F., Ribeiro, L. S., & Pereira, M. S. (2007). Teor de açúcares solúveis e insolúveis em folha de videiras, cv. syrah, em diferentes posições no ramo e épocas do ano. Revista Brasileira de Fruticultura, 29(1), 42-47. [ Links ]
Dorchin, N., Cramer, M. D., & Hoffmann, J. H. (2006). Photosynthesis and sink activity of wasp-induced galls in Acacia pycnantha. Ecology, 87, 1781-1791. Retrieved from http://treephys.oxfordjournals.org/. [ Links ]
Dreger-Jauffret, F., & Shorthouse, J. D. (1992). Diversity of gall-inducing insects and their galls, In J. D. Shorthouse & O. Rohfritsch (Eds.), Biology of insect induced galls (pp. 8-33). Oxford: Oxford University. [ Links ]
Dubois, M., Gilles, K. A., Hamilton, J. K., Rebers, P. A., & Smith, F. (1956). Colorimetric method for determination of sugars and related substances. Analytical Chemistry, 28, 350-356. [ Links ]
Fernandes, G. W., & Price, P. W. (1992). The adaptive significance of insect gall distribution: survivorship of species in xeric and mesic habitats. Oecologia, 90, 14-20. [ Links ]
Fischer, A. (2007). Nutrient remobilization during leaf senescence. In S. Gan. (Ed.), Senescence Processes in Plants - Annual Plant Reviews (pp. 87-102). New York: Cornell University. [ Links ]
Forzza, R. C., Stehmann, J. R., Nadruz, M., Filardi, R. P. F., Costa, A., Jr. Carvalho, A. A., Peixoto, A. L., Walter, B. M. T., Bicudo, C., Moura, C. W. N., Zappi, D., Costa, D. P., Lleras, E., Martinelli, G., Lima, H. C., Prado, J., Baumgratz, J. F. A., Pirani, J. R., Sylvestre, L. S., Maia, L. C., Lohmann, L. G., Paganucci, L., Alves, M. V. S., Silveira, M., Mamede, M. C. H., Bastos, M. N. C., Morim, M. P., Barbosa, M. R., Menezes, M., Hopkins, M., Evangelista, P. H. L., Goldenberg, R., Secco, R., Rodrigues, R. S., Cavalcanti, T., & Souza, V. C. (2010). Catalogo de Plantas fungos e do Brasil, v.1. Río de Janeiro: Andrea Jakobsson Estúdio. [ Links ]
Fournier, O. L. A. (1974). Un método cuantitativo para la medición de características fenológicas en árboles. Turrialba, 24, 422-423. [ Links ]
Fournier, O. L. A., & Charpantier, C. (1975). El tamaño de la muestra y la frecuencia de las observaciones en el estudio de las características fenológicas de los árboles tropicales. Turrialba, 25, 45-48. [ Links ]
Gonçalves, S. J. M. R., Isaias, R. M. S., Vale, F. H. A., & Fernandes, G. W. (2005). Sexual dimorphism of Pseudotectococcus rolliniae Hodgson & Gonçalves 2004 (Hemiptera Coccoidea Eriococcidae) influences gall morphology on Rollinia laurifolia Schltdl. (Annonaceae). Tropical Zoology, 18, 161-169. [ Links ]
Gonçalves, S. J. M. R., Moreira, G. R. P., & Isaias, R. M. S. (2009). A unique seasonal cycle in a leaf gall-inducing insect: the formation of stem galls for dormancy. Journal of Natural History, 43, 843-854. doi: 10.1080/00222930802615690. [ Links ]
Hartley, S. E. (1998). The chemical composition of plant galls: Are level of nutrients and secondary compounds controlled by the gall former? Oecologia, 113, 492-501. [ Links ]
Inbar, M., Eshel, A., & Wool, D. (1995). Interspecific competition among phloem-feeding insects mediated by induced host-plant sinks. Ecology, 76, 1506-1515. Retrieved from http://www.jstor.org/stable/1938152. [ Links ]
Leitão, A. C., & Silva, O. A. (2004). Variação sazonal de macronutrientes em uma espécie arbórea de cerrado, na Reserva Biológica e Estação Experimental de Mogi-Guaçu, estado de São Paulo, Brasil. Rodriguésia, 55(84), 127-136. Retrieved fom http://rodriguesia.jbrj.gov.br/FASCICULOS/Rodrig55_84/85584.pdf. [ Links ]
Lev-Yadun, S. (2003). Stem cells in plants are differentiated too. Current Topics in Plant Biology, 4, 93-102. [ Links ]
Marmit, K. S., & Sharma, S. L. (2008). Quantitative estimation of some metabolites and enzymes in insect induced leaf galls of Mangifera indica. Asian Journal of Experimental Sciences, 22(3), 343-346. [ Links ]
Marur, C., & Sodek, L. (1995). Microwave drying of plant material for biochemical analysis. Revista Brasileira de Fisiologia Vegetal, 7(1), 111-114. [ Links ]
McCready, R. M., Guggolz, J., Silveira, V., & Owens, H. S. (1950). Determination of starch and amylase in vegetables. Application to peas. Analytical Chemistry, 22, 1156-1158. [ Links ]
Morellato, L. P. C., Leitão Filho, H. F., Rodrigues, R. R., & Joly, C. A. (1990). Estratégias fenológicas de espécies arbóreas em floresta de altitude na Serra do Japi, Jundiaí São Paulo. Revista Brasileira de Biologia, 50, 149-162. [ Links ]
Moura, M. Z. D., Soares, G. L. G., & Isaias, R. M. S. (2008). Species-specific changes in tissue morphogenesis induced by two arthropod leaf gallers in Lantana camara L. (Verbenaceae). Australian Journal of Botany, 56, 153-160. doi: 10.1071/BT07131 0067-1924/08/020153. [ Links ]
Oliveira, D. C., & Isaias, R. M. S. (2009). Influence of leaflet age in anatomy and possible adaptive values of the midrib gall of Copaifera langsdorffii (Fabaceae: Caesalpinoidae). Revista de Biología Tropical, 57, 293-302. Retrieved from http://www.biologiatropical.ucr.ac.cr/attachments/volumes/vol57-1-2/26-Oliveira-Copaifera%20langsdorffii.pdf. [ Links ]
Oliveira, D. C., & Isaias, R. M. S. (2010). Redifferentiation of leaflet tissues during midrib gall development in Copaifera langsdorffii (Fabaceae). South African Journal of Botany, 76(2), 239-248. doi:10.1016/j.sajb.2009.10.011. [ Links ]
Oliveira, P. S., & Marquis, R. J. (2002). The Cerrados of Brazil. New York: Columbia University Press. [ Links ]
Oliveira, D. C., Christiano, J. C. S., Soares, G. L. G., & Isaias, R. M. S. (2006). Reações de defesas químicas e estruturais de Lonchocarpus muehlbergianus Hassl. (Fabaceae) à ação do galhador Euphalerus ostreoides Crawf. (Hemiptera: Psyllidae). Revista Brasileira de Botânica, 29, 657-667. [ Links ]
(Fabaceae) and multiple galling insects: formation of seasonal patterns. Oliveira, D. C., Mendonça Jr., M. S., Moreira, A. S. F. P., Lemos-Filho, J. P., & Isaias, R. M. S. (2012). Water stress and phenological synchronism between Copaifera langsdorffiiJournal of Plant Interaction, 1-9. Retrieved from http://dx.doi.org/10.1080/17429145.2012.705339. [ Links ]
Pedroni, F., Sanchez, M., & Santos, F. A. M. (2002). Fenologia da copaíba (Copaifera langsdorffii Desf. – Leguminosae, Caesalpinioideae) em uma floresta semidecídua no sudeste do Brasil. Revista Brasileira de Botânica, 25(2), 183-194. [ Links ]
Pirani, F. R., Sanchez, M., & Pedroni, F. (2009). Fenologia de uma comunidade arbórea em cerrado sentido restrito, Barra do Garças, MT, Brasil. Acta Botanica Brasilica, 23(4), 1096-1109. [ Links ]
Price, P. W., Waring, G. L., & Fernandes, G. W. (1986). Hypotheses on the adaptive nature of galls. Proceedings of the Entomological Society of Washington, 88, 361-363. [ Links ]
Price, P. W., Fernandes, G. W., & Waring, G. L. (1987). Adaptive nature of insect galls. Environmental Entomology, 16, 15-24. [ Links ]
Rizzo, J. A. (1970). Contribuição ao conhecimento da Flora de Goiás-área na Serra Dourada. Tese em Livre Docência. Universidade Federal de Goiás, Goiânia, GO. [ Links ]
Rohfritsch, O. (1992). Patterns in gall development. In J. D. Shorthouse & O. Rohfritsch (Eds.), Biology of insect induced galls (pp. 60-86). Oxford: Oxford University. [ Links ]
Shannon, J. C. (1968). Carbon-14 distribution in carbohydrates of immature Zea mays kernels following 14CO2 treatment of intact plants. Plant Physiology, 43, 1215-1220. [ Links ]
Tedesco, M. J., Gianello, C., Bissani, C. A., Bohnen, H., & Volkweiss, S. J. (1995). Análise de solo, plantas e outros materiais. Porto Alegre: Universidade Federal do Rio Grande do Sul. [ Links ]
Weis, A. E., Walton, R., & Crego, C. L. (1988). Reactive plant tissue site and the population biology of gall makers. Annual Review of Entomology, 3, 467-486. doi: 10.1146/annurev.en.33.010188.002343. [ Links ]
Wolf, S., & Greiner, S. (2012). Growth control by cell wall pectins. Protoplasma. doi: 10.1007/s00709-011-0371-5. [ Links ]
Yukawa, J. (2000). Synchronization of gallers with host plant phenology. Population Ecology, 42, 105-13. [ Links ]
*Correspondencia a:
Ariane C. Castro: Laboratório de Anatomia Vegetal, Universidade Federal de Minas Gerais, Av. Antônio Carlos 6627, Campus Pampulha, Belo Horizonte, Minas Gerais, Brasil; arianechc@yahoo.com.br
Denis C. Oliveira: Laboratório de Ecologia e Desenvolvimento de Galhas, Universidade Federal de Uberlândia, Rua Ceará s/n, Campus Umuarama, Minas Gerais, Brasil; denisoliveira@inbio.ufu.br
Ana Silvia F. P. Moreira: Laboratório de Fisiologia Vegetal, Universidade Federal de Uberlândia, Rua Ceará s/n, CampusUmuarama, Minas Gerais, Brasil; anasilvia@inbio.ufu.br
Rosy M.S. Isaias: Laboratório de Anatomia Vegetal, Universidade Federal de Minas Gerais, Av. Antônio Carlos 6627, Campus Pampulha, Belo Horizonte, Minas Gerais, Brasil;rosy@icb.ufmg.br
1.Laboratório de Anatomia Vegetal, Universidade Federal de Minas Gerais, Av. Antônio Carlos 6627, Campus Pampulha, Belo Horizonte, Minas Gerais, Brasil; arianechc@yahoo.com.br; rosy@icb.ufmg.br Correspondence
2.Laboratório de Ecologia e Desenvolvimento de Galhas, Universidade Federal de Uberlândia, Rua Ceará s/n, Campus Umuarama, Minas Gerais, Brasil; denisoliveira@inbio.ufu.br
3.Laboratório de Fisiologia Vegetal, Universidade Federal de Uberlândia, Rua Ceará s/n, CampusUmuarama, Minas Gerais, Brasil; anasilvia@inbio.ufu.br
Received 15-XI-2012. Corrected 10-II-2013. Accepted 13-III-2013.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}