










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.61 n.4 San José Oct./Dec. 2013
Acclimation of seedlings of Gnetum leyboldii Tul. (Gnetaceae) to light changes in a tropical rain forest
Aclimatación de plántulas Gnetum leyboldii Tul. (Gnetaceae) a los cambios de la luz en un bosque lluvioso tropical
Aclimatación de plántulas Gnetum leyboldii Tul. (Gnetaceae) a los cambios de la luz en un bosque lluvioso tropical
*Dirección para correspondencia:
Abstract
The neotropical liana Gnetum leyboldii (Gnetaceae) is a gymnosperm that resembles angiosperms in wood anatomy, overall morphology, and seed dispersal mechanism. Like other woody lianas, seedlings germinate in the shaded forest understory and start climbing towards the canopy, being exposed to sites with extreme differences in light conditions. However, the extent of physiological and structural adjustment to contrasting light conditions in the early regeneration stages of Gnetum is unknown. To answer this question, we analyzed seedling growth and photosynthetic responses using a common garden experiment with two light regimes: full sun and low light (20% of full sun) at La Selva Biological Station, Costa Rica. We also characterized the germination pattern of this species. We monitored one and half-month old seedlings for four months. Leaf structure finely adapted to light treatments, but gas exchange properties were buffered by large seed reserves, which dominated biomass distribution (about 50% of the total biomass), followed by stem (27%), leaf (16%) and root biomass (6%) across light conditions. The presence of large seeds and the low photosynthetic rates of seedlings in both environments show that G. leyboldii is specialized to exploit deep shade. More research is needed to determine if the patterns found in G. leyboldii are typical of similar lianas that initially exploit deep-shaded understories in their ascension to the canopy.
Key words: Gnetum leyboldii, lianas, photosynthetic acclimation, biomass allocation, germination, Costa Rica.
Resumen
La liana neotropical Gnetum leyboldii (Gnetaceae) es una gimnosperma que se asemeja a las angiospermas en la anatomía de la madera, morfología general de la planta y mecanismo de dispersión de semillas. Al igual que otras lianas leñosas, las plántulas se regeneran en el sotobosque bajo dosel cerrado y eventualmente ascienden hacia el dosel, explotando sitios con diferencias extremas en condiciones lumínicas. Se desconoce el grado de ajuste fisiológico a condiciones lumínicas contrastantes en las primeras fases de regeneración de Gnetum. Para contestar esta pregunta, analizamos las respuestas de crecimiento de las plántulas a ambientes contrastantes de luz de sol y sombra en un jardín común con condiciones de alta (cielo abierto) y baja luminosidad (20% del ambiente de sol) en la Estación Biológica La Selva, Costa Rica. También caracterizamos su patrón de germinación. Monitoreamos plántulas de 1.5 meses de edad por 4 meses. La estructura foliar mostró una fina adaptación a los tratamientos de luz, pero las propiedades de intercambio gaseoso no cambiaron sino que fueron amortiguadas por las reservas de las semillas grandes, las cuales dominaron la distribución de biomasa (aproximadamente 50% de la biomasa total) seguidas por el tallo (27%), la hoja (16%) y raíces (6%). El tener semillas grandes y plántulas con bajas tasas fotosintéticas muestra que G. leyboldii en su etapa de plántula está adaptado para explotar la sombra profunda. Se requiere más investigación para determinar si los patrones encontrados en G. leyboldii son típicos de otras lianas que inicialmente explotan la sombra profunda en su ascensión al dosel.
Palabras clave: Gnetum leyboldii, lianas, aclimatación fotosintética, adaptación a la sombra, distribución de biomasa, germinación criptocotilar, Costa Rica.
Lianas represent a conspicuous structural element of tropical forests. The great abundance and distribution of lianas distinguishes tropical from temperate forests (Gentry, 1991; Putz, 1984; Schnitzer & Bongers, 2002). In their ascension to the forest canopy lianas develop a strategy of biomass allocation that enhances mobility and increases light interception, using external substrates for mechanical support while expressing a variety of climbing mechanisms and flexible biomass allocation, targeting sites with higher opportunities for carbon gain (Avalos & Mulkey, 1999a; Toledo-Aceves & Swaine, 2008). Lianas that germinate and survive in the shade show clear signs of shade tolerance (Nabe-Nielsen, 2002; Sanches & Válio, 2008), despite that many species are considered typical pioneers in terms of their physiological and morphological responses to light changes as adults (Avalos & Mulkey, 1999a; Avalos, Mulkey, Kitajima & Wright, 2007; Cai, Poorter, Cao & Bongers, 2007; Sanches & Válio, 2002).
Compared to trees, lianas invest more biomass in leaves (Castellanos, 1991; Putz, 1984) and roots (Toledo-Aceves & Swaine, 2008), and concentrate foliage on top of the canopy (Avalos &Mulkey, 1999b; Avalos et al., 2007). These differences in biomass allocation facilitate a rapid liana response to light changes, and faster adjustment to heterogeneous light conditions (Avalos & Mulkey, 1999a; Avalos et al., 2007), favoring their potential increase under global warming scenarios (Granados & Körner, 2002; Schnitzer & Bongers, 2011). Their well-developed capacity for clonal growth, allows them to colonize the entire forest profile and simultaneously exploit sites with extreme differences in light (Avalos & Mulkey, 1999a; Avalos et al., 2007). Their physiological flexibility, abundance and dominance of the canopy leaf area make lianas a highly dynamic element affecting the growth, structure and diversity of tropical plant species (Phillips, Vásquez-Martínez, Monteagudo-Mendoza, Baker & Núñez-Vargas, 2005; Schnitzer & Carson, 2010).
Lianas of the genus Gnetum are broad-leaved gymnosperms with more than 30 species of pantropical distribution (Rodin & Kapil, 1969). Among non-flowering vascular plants, Gnetum is most similar to angiosperms in vegetative structure. The anatomical organization of leaves and woody tissues of Gnetum is very similar to those of dicotyledoneous plants (Fisher & Ewers, 1995) to the extent that Gnetales is considered a phylogenetic link between Gymnosperms and Angiosperms (Shutov, Braun, Chesnokov, Horstmann, Kakhovskaya & Bäumlein, 1998). Most of the research on this genus has focused on anatomical (C. R. B., 1908; Coulter, 1908; J. M. C., 1915, 1916a, 1916b; Thompson, 1916; Haining, 1920; Johnson, 1950; Muhammad & Sattler, 1982; Fisher & Ewers, 1995; Carlquist, 1996) and genetic questions (Frohlich & Meyerowitz, 1997). The study of Feild and Balun (2008) is the only one that has examined physiological responses in this genus. These authors considered adults of four species (the only two tree species in the genus, and two liana species) growing under full sun in the rainforests of Papua New Guinea, and expected high photosynthetic rates similar to woody tropical pioneers due to the climbing habit (being the only gymnosperm with this growth form), wide vessels, broad pinnate-veined leaves and abundance in productive lowland rainforest. In contrast, all Gnetum species showed physiological responses similar to shade tolerant angiosperms (Field & Balun, 2008). Despite of this contribution, there is a dearth of ecophysiological studies on Gnetum, especially on its early regeneration requirements, and how their responses compare with those of other lianas, especially those that start in the shade (i.e., Nabe-Nielsen, 2002; Sanches &Válio, 2008).
In this study, we examined the biomass allocation and the photosynthetic performance of seedlings of Gnetum leyboldii Tul. to contrasting conditions of sun and shade environments. We also characterized its germination pattern. Seedlings of this liana established under full shade, but adults display foliage on top of the canopy. As G. leyboldii is a gymnosperm with many angiosperm-like morphological traits, the analysis of its light acclimation and biomass allocation strategies at the seedling stage is crucial to understand the diversity of the early regeneration niche of lianas (and tropical plants in general) within the context of the light-limited forest understory.
Materials and Methods
Study site: We conducted the research at La Selva Biological Station, Costa Rica (10°25’58’’ N - 84°00’06’’ W, 30-150m). La Selva is a forest reserve of 1 600ha managed by the Organization for Tropical Studies, located in the NE Caribbean lowlands at the base of the Central Volcanic Mountain Slope. Hartshorn & Peralta, (1988) classifies La Selva as a tropical wet forest, according to the Holdridge life zone system. La Selva has a mean liana density of 1 493 stems/ha and a mean species richness of 23 species per 864m2 (Marasco, Schnitzer & Carson, 2004). The incident mean daily solar radiation at this site is 14.9M/Jm2/d, with a range of 0.4 to 31.3M/Jm2/d (1993-1998), and the mean annual temperature (1982-1998) is 24.6°C (Loescher, Oberbauer, Gholz & Clark, 2003). Mean annual precipitation is 4 000mm (1963-1991), with a short dry period from December to late May, although no month receives less than 100mm (Sanford, Paaby, Luvall & Phillips, 1994).
Study species: The dioecious liana Gnetum leyboldii Tul. (Gnetaceae) can reach heights of up to 30m above the ground, with stems of 15-20cm in DBH. Leaves are simple and opposite, decussate, coriaceous, broadly ovate, 9-15cm long and 5-10cm wide. Seeds are 4.5-5.0cm long and 2cm wide, oblong, with an external reddish tissue, which is moderately fleshy resembling a drupe, and about 1mm thick (Grayum, 2003; Fig. 1). Seeds have an average weight of 12.86±0.16g (n=125).
Germination and seedling growth experiments: Seed germination and seedling growth experiments started in June 2003. We collected seeds of G. leyboldii from two sites: one along the Holdridge Trail “SHO” (23 seeds collected on June 6, and 42 on June 27), and the other along the Experimental Trail “CES” (71 seeds collected on June 6, and 41 on June 18). We put seeds to germinate under moderate sun conditions in flats filled with filter paper soaked in water, and watered using the natural precipitation of the La Selva greenhouse. After one week, we transplanted all seeds to a flexible plastic bag (226cm3) filled with soil taken from the field in an area close to the common garden (see below). The bags were sufficiently large to prevent the adverse effects of root binding, and provided appropriate conditions prior to transplant into the common garden. We monitored each individual seedling for changes in leaf production and stem elongation every two weeks.
Common garden experiment: We established a common garden in a large gap (250m2) without crown interference from surrounding trees. We removed plant debris from an area of 13.5x7.5m, and randomly assigned each seedling to four contiguous subplots of 5x2.5m (two sun and two shade plots), leaving a distance of 50cm between seedlings using a total of 15 individuals per plot. We exposed seedlings under the high light treatment (or “sun seedlings”) to direct sun and enclosed them with chicken wire held up with bamboo poles to prevent large and medium sized mammals from entering the garden. We exposed seedlings under low light (or “shade seedlings”) to 20% of the natural light using neutral greenhouse shade cloth put over shade houses made of 2m tall bamboo poles. We verified these light intensities using two quantum light sensors (LI-190SZ, LI-COR Inc., Nebraska, USA) connected to a data logger (LI-1400, LI-COR Inc., Nebraska, USA). The understory at La Selva under deep canopy cover in primary forests usually receives 1-5% of full sun (Chazdon & Fetcher, 1984). Although we did not replicate these light levels, the ones used in this experiment (full sun vs. 20% of full sun) presented sufficient differences to trigger a response in seedlings. After four months (October to February 2003) we harvested all seedlings and partitioned their biomass into leaves, stems and petioles, roots, and embryonic cotyledons (which were included as part of the seed biomass). Seedling age (days after germination) ranged from 131-212 days at the date of harvest.
Adjustment to different light conditions: We monitored changes in leaf structure, photosynthetic responses, and biomass allocation of seedlings in each light treatment. Physiological measurements were restricted to leaves fully expanded under sun or shade. We took care to distinguish new leaves produced after the transfer from the shade house into the common garden by numbering leaves with plastic tags. We measured the proportion of total biomass distributed to roots, stems and petioles, and leaves from dry weights of at least 30 replicates per treatment. We also calculated the leaf area ratio (LAR), the leaf weight ratio (LWR), and the leaf mass per unit of area per plant after adding all leaves in a seedling (LMAP). Total leaf area (cm2) of all leaves in a seedling was calculated by adding up the area of individual leaves after fitting a linear regression between leaf width and length versus actual leaf area measured with a LI-COR leaf area meter. We stored and dried all plant material used for biomass allocation in an oven at 60oC for two days until constant weight prior to measurement.
Photosynthetic capacity: To characterize the photosynthetic capacity of seedlings, we measured fully expanded leaves using a portable in situ photosynthesis system (LI-6400, LI-COR Inc., Nebraska, USA), an ambient CO2 concentration of 386µmol CO2 a water vapor concentration of 24-28mol H2O and a light intensity of 1500µmol photons/m2/s (PFD).
The following analyses were performed:
Germination: We used failure-time analysis to compare germination trajectories for each site and collection date (Fox, 2001). This method consists of a univariate survival analysis, which estimates the survival functions using the product-limit (Kaplan-Meier) method for one or more groups (site and collection date). The survival data contain duration times until the occurrence of a specific event, in this case the time until seed germination. We used a log-rank c2 statistic to test for homogeneity between groups (SAS 1994).
Biomass allocation among light treatments: We used a Principal Component Analysis (PCA, Quinn & Keough, 2002) to consolidate the main relationships between biomass allocation variables among light treatments. PCA removes the colinearity among the five variables included in this analysis (root, shoot, leaf and seed mass, and leaf area), which are functionally related. We used a one-way MANOVA (light treatment as the main factor) to compare the mean value of the two principal components for each light condition.
Assessment of differences in leaf structure and photosynthetic responses among light treatments: We used a one-way MANOVA with light treatment as the main factor to compare differences in LAR, LWR and LMAP after consolidating these variables using a PCA. We used one-way ANOVA to determine physiological differences among light conditions in terms of photosynthetic rates and stomatal conductance. In all instances, we adjusted F-values with a sequential Bonferroni correction following Rice (1988). All statistical analyses were done using the JMP statistical package (SAS, 1994).
Results
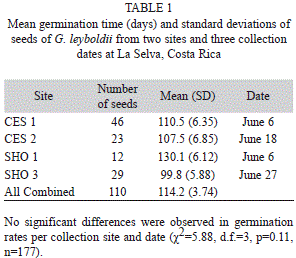
Germination: The germination of G. leyboldii was cryptocotylar and hypogeal, and the embryo had opposite cataphylls. Germination time ranged from nine to 166 days. There were no significant differences in the proportion of germinating seeds per collection site and collection date (χ2=5.88, d.f.=3, p=0.11; Fig. 2). Overall, 62% of the seeds had a mean germination time of 114.2±3.74 days (n=177; Table 1).
Leaf production and leaf loss: Leaf production in the common garden was higher under shade conditions (average new leaves per seedling=11.1±0.7, n=20) as compared to sun seedlings (average=9.4±1.1, n=20). Leaf loss was higher under sun (average leaves lost per seedling=1.00±0.05, n=20) relative to shade conditions (average=0.65±0.05, n=20). In both cases, there was high variation among seedlings.
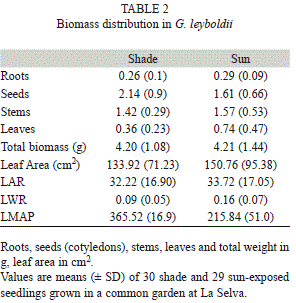
Effects of light treatments on biomass allocation and leaf structure: Biomass distribution maintained similar patterns across light treatments, although shade seedlings had higher seed biomass relative to sun seedlings, and sun seedlings had more biomass allocated to leaves (despite of lower leaf numbers in this environment, Table 2). This is consisting with a slightly higher LWR in the sun and lower LMAP in the shade. Sun seedlings have considerably more weight in leaves relative to leaf area than shade seedlings.
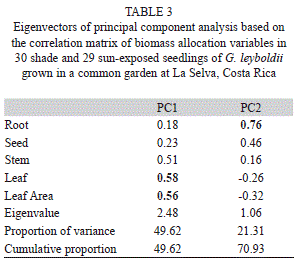
The first principal component explained 49.62% of the total variation in overall biomass allocation, having leaf biomass as the dominating factor. Root and seed biomass dominated the second principal component, which explained 21.31% of the variation (Table 3). The first component was normally distributed, whereas the second component was transformed using the Box-Cox procedure to satisfy the requirements of MANOVA, which used these components as response variables and light treatment as predictor variable. It showed lack of significant differences between sun and shade treatments in overall biomass allocation (F1,57=0.01, p=0.42). Patterns of biomass distribution were similar for sun and shade seedlings, being dominated by seed mass (about half of total biomass in all cases) followed by shoots, leaves and roots.
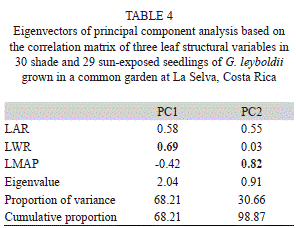
In terms of leaf structure, the first principal component explained 68.21% of the total variation, whereas the second component explained 30.66%. LWR dominated the first component, followed by LMAP in the second component (Table 4). LWR almost doubled in the sun, whereas LMAP was 60% higher in the shade. Minor changes in leaf structure facilitated the adaptation to light conditions under sun and shade environments in G. leyboldii.
Effects of light treatments on photosynthetic responses: The photosynthetic rate and stomatal conductance for shade and sun seedlings were low, and we did not observe significant differences between light environments (F2,18=0.37, p=0.6; Table 5).
Discussion
The growth and photosynthetic responses of seedlings of G. leyboldii conformed to a shade-adaptive character syndrome (Mulkey, Wright & Smith, 1993). Plants that conform to this syndrome are able to not only tolerate long periods in the shade, but also grow, survive and reproduce under a closed canopy (Valladares & Niinemets, 2008). A shade-tolerant plant will survive, grow and eventually move into a different life stage under shade. The ability to gather enough resources while avoiding losses determines this shade-adaptive character syndrome as an integrated life-history strategy, combining whole-plant level responses, including biomass allocation, morphological adjustment, plant architecture and leaf-level physiological changes (Foster & Janson, 1985; Callaway, 1992; Osunkoya, Ash, Hopkins & Graham, 1994). Shade-tolerants have lower phenotypic plasticity and slow growth over short time scales (e.g., limited biomass increases in low light), but can show high plasticity for morphological features that optimize light capture, as well as survival in the shade. The presence of recalcitrant seeds in this species is congruent with this syndrome.
The main adjustment to contrasting light environments was effected through changes in leaf packaging after leaf expansion. There was an increase of over 50% in leaf biomass and LWR in the sun, with a consequent decrease in LMAP of a similar magnitude. Leaves in the sun had a smaller area per unit of leaf weight, and overall more biomass in leaves relative to the overall plant weight. More leaves in the shade of a relatively larger area per unit of leaf weight target light interception. These differences were subtle and were not reflected in gas exchange properties, and were potentially buffered by large seed reserves still present under both environments and in seedlings that started to climb after 4 months. The changes in leaf structure observed here are consistent with the expected direction of differences under sun and shade environments and are not unique to lianas (i.e., Poorter, 2001).
Compared to other tropical species, G. leyboldii has large seeds (e.g. Foster & Janson, 1985; Hammond & Brown, 1995; Kitajima, 1996a, 2002) and large nutrient reserves, consistent with the shade-adaptive character syndrome. Large seeds are favored when carbon deficits are likely to occur early in life due to deep shade (Armstrong & Westoby, 1993; Harms & Dalling, 1997; Kitajima, 2003). The fact that seedlings were still attached to their cotyledons after four months, and that seed mass represented the highest allocation compartment (nearly 50% of total weight) in seedlings with stem lengths of over 1.5m and a relatively well-developed leaf area, may explain the lack of physiological differences among light treatments, although differences in light availability were significant. Seed reserves buffered the effects of environmental differences, and created a seedling phenotype with a similar performance under sun and shade environments. The long connection period between seedlings and seed reserves may provide G. leyboldii with enough time to establish under a wide variety of light conditions, and even start a necessary ecto-mycorrhizic association (Onguene & Kuyper, 2001). The low photosynthetic rates observed here are also characteristic of shade-adapted species (Kitajima, 1996b), and are consistent with the patterns reported by Feild & Balun (2008) for adult Gnetum plants, although from different species.
In contrast to our data, Gómez (1983) reported a germination period of 25-35 days. This difference is due to a lower seed number used in that study, and that seeds were on the ground for some time before being put to germinate. Another characteristic associated with delayed germination is the presence of cryptocotylar cotyledons (Duke, 1969; Flores & Rivera, 1983) also present in G. leyboldii. What really determines the time of seed germination is yet to be determined, and more research is needed to fully understand this pattern.
Finally, we expect the preferred light strategy of G. leyboldii to shift from shade tolerance in seedlings to light demanding in adults, which display their foliage on top of the canopy. This expectation emerges from the evidence of Fetcher, Oberbauer and Chazdon (1994), who reported changes in leaf structure, with adult lianas doubling leaf thickness relative to seedlings grown in full shade, whereas in seedlings there was a sustained increase in leaf thickness with increasing light. Seedlings adapt to heterogeneous light conditions by changing leaf structure, as shown in our study. Ontogenetic niche shifts along a vertical gradient are common in canopy trees (Poorter, Bongers, Sterck & Wöll, 2005) and should be the norm in lianas that regenerate in the shade but ascend to the canopy, as in this case. The results of Feild and Balun (2008) for adult Gnetum plants challenge this expectation by showing that adult species of Gnetum were shade tolerants. More research is needed to proof ontogenetic niche shifts in G. leyboldii. To do this, it will be necessary to couple the examination of physiological responses with the analysis of foliage distribution along the forest profile in adults of this species.
In conclusion, the large seeds, slow germination time, long permanence of seed reserves during initial seedling growth, low photosynthetic rates, and fine-tuning adjustment of leaf structure among light treatments of sun and shade show that G. leyboldii initially regenerates as a specialized shade-adapted species (Poorter & Rose, 2005). More time under different light conditions may be needed to characterize the ontogenetic niche shift of G. leyboldii when seed resources are completely depleted and the liana climbs into the canopy. Since lianas are increasing in abundance and biomass across the tropics (Schnitzer & Bongers, 2011) the understanding of the functional basis of this shade tolerance is crucial to increase our knowledge on the range of physiological responses associated with lianas especially under global warming scenarios. Within this context, G. leyboldii represents an excellent model organism.
Acknowledgments
The authors acknowledge the support of the Organization for Tropical Studies (OTS) and the CR-USA Foundation through the OTS-REU-CR Program. The organization IDEAWILD provided the datalogger and light sensors. The TOWERS team, Eugenia Flores, Luis Diego Gómez, Evan Notman, Karin Gastreich, Marcela Fernández, Harlyn Ordóñez, Juanita Zeledón, Enrique Castro, Antonio Trabucco, Reinaldo Aguilar, and Rocío Fernández kindly provided comments that improved the manuscript.
References
Armstrong, D. P. & Westoby, M. (1993). Seedlings from large seeds tolerate defoliation better: A test using phylogenetically independent contrasts. Ecology, 74(4), 1092-1100. [ Links ]
Avalos, G. & Mulkey, S. S. (1999a). Photosynthetic acclimation of the liana Stigmaphyllon lindenianum (Malpighiaceae) to light changes in a tropical dry forest canopy. Oecologia, 120(4), 475-484. [ Links ]
Avalos, G. & Mulkey, S. S. (1999b). Seasonal changes in liana cover in the upper canopy of a neotropical dry forest. Biotropica, 31(1), 186-192. [ Links ]
Avalos, G., Mulkey, S. S., Kitajima, K. & Wright, S. J. (2007). Colonization strategies of two liana species in a tropical dry forest canopy. Biotropica, 39(3), 393-399. [ Links ]
Cai, Z. Q., Poorter, L., Cao, K. F. & Bongers, F. (2007). Seedling growth strategies in Bauhinia species: comparing lianas and trees. Annals of Botany, 100(4), 831-838. [ Links ]
Callaway, R. M. (1992). Morphological and physiological responses of three California Oak species to shade. International Journal of Plant Sciences, 153(3), 434-441. [ Links ]
Carlquist, S. (1996). Wood, bark and stem anatomy of New World species of Gnetum. Botanical Journal of the Linnean Society, 120(1), 1-19. [ Links ]
Castellanos, A. E. (1991). Photosynthesis and gas exchange of vines. In F. E. Putz & H. A. Mooney (Eds.), The biology of vines (pp. 181-204). Cambridge: Cambridge University Press. [ Links ]
Chazdon, R. L. & Fetcher, N. (1984). Photosynthetic light environments in a lowland tropical rainforest in Costa Rica. Journal of Ecology, 72(2), 553-564. [ Links ]
Coulter, J. M. (1908).The Embryo sac and embryo of Gnetum gnemon. Botanical Gazette, 46(1), 43-49. [ Links ]
C. R. B. (1908). Adventitious buds in leaves of Gnetum. Botanical Gazette, 45(5), 357. [ Links ]
Duke, J. A. (1969). On tropical tree seedlings. I. seeds, seedlings, systems, and systematics. Annals of the Missouri Botanical Garden, 56(2), 125-161. [ Links ]
Feild, T. S. & Balun, L. (2008). Xylem hydraulic and photosynthetic function of Gnetum (Gnetales) species from Papua New Guinea. New Phytologist, 177(3), 665-675. [ Links ]
Fetcher, N., Oberbauer, S. F. & Chazdon, R. L. (1994). Physiological Ecology of Plants. In L. A. McDade, K. S. Bawa, H. A. Hespenheide & G. S. Hartshorn (Eds.), La Selva: Ecology and Natural History of a Neotropical Rain Forest (pp. 128-141). Chicago: University of Chicago Press. [ Links ]
Fisher, J. B. & Ewers, F. W. 1995. Vessel dimensions in liana and tree species of Gnetum (Gnetales). American Journal Botany, 82(11), 1350-1357. [ Links ]
Flores, E. M. & Rivera, D. I. (1983). Criptocotilia en algunas dicotiledóneas tropicales. Brenesia, 32, 19-26. [ Links ]
Foster, S. A. & Janson, C. H. (1985). The relationship between seed size and establishment conditions in tropical woody plants. Ecology, 66(3), 773-780. [ Links ]
Fox, G. A. (2001). Failure-time Analysis: Studying times to events and rates at which events occur. In S. A. Scheiner & J. Gurevitch (Eds.), Design and Analysis of Ecological Experiments (pp. 235-266). New York: Oxford University Press. [ Links ]
Frohlich, M. W. & Meyerowitz, E. M. (1997). The search for flower homeotic gene homologs in basal Angiosperms and Gnetales: A potential new source of data on the evolutionary origin of flowers. International Journal of Plant Sciences, 158(6), S131-S142. [ Links ]
Gentry, A. H. (1991). The distribution and evolution of climbing plants. In F. E. Putz & H. A. Mooney (Eds.), The Biology of Vines (pp. 3-42).Cambridge: Cambridge University Press. [ Links ]
Gómez, L. D. (1983). Plantae Mesoamericana Novae, VIII. Phytologia, 53(2), 97-100. [ Links ]
Granados, J. & Körner, C. (2002). In deep shade, elevated CO2 increases the vigor of tropical climbing plants. Global Change Biology, 8(11), 1109-1117. [ Links ]
Grayum, M. H. (2003). Gnetaceae. In B. F. Hammel, M. H. Grayum & N. Zamora (Eds.), Manual de Plantas de Costa Rica Vol II (pp. 7-8). Missouri: Missouri Botanical Garden Press. [ Links ]
Haining, H. I. (1920). Development of embryo of Gnetum. Botanical Gazette 70(6), 436-445. [ Links ]
Hammond, D. S. & Brown, V. K. (1995). Seed size of woody plants in relation to disturbance, dispersal, soil type in wet neotropical forest. Ecology, 76(8), 2544-2561. [ Links ]
Harms, K. E. & Dalling, J. W. (1997). Damage and herbivory tolerance through resprouting as an advantage of large seed size in tropical trees and lianas. Journal of Tropical Ecology, 13(4), 617-621. [ Links ]
Hartshorn, G. S. & Peralta, R. (1988). Preliminary description of primary forests along the La Selva-Volcan Barva altitudinal transect, Costa Rica. In F. Almeida & C. Pringle (Eds.), Tropical Rainforests: Diversity and Conservation (pp. 281-295). San Fransisco: California Academy of Science. [ Links ]
J. M. C. (1915). Morphology of Gnetum. Botanical Gazette, 60(4), 335. [ Links ]
J. M. C. (1916a). Morphology of Gnetum. Botanical. Gazette, 62(1) 85. [ Links ]
J. M. C. (1916b). Morphology of Gnetum. Botanical Gazette, 62(3), 246. [ Links ]
Johnson, M. A. (1950). Growth and development of the shoot of Gnetum gnemon L. I. The shoot apex and pith. Bulletin of the Torrey Botanical Club 77(5), 354-367. [ Links ]
Kitajima, K. (1996a). Cotyledon functional morphology, patterns of seed reserve utilization and regeneration niches of tropical tree seedlings. In M. D. Swaine (Ed.), The Ecology of Tropical Forest Tree Seedlings (pp. 193-210). New York: UNESCO. [ Links ]
Kitajima, K. (1996b). Ecophysiology of tropical tree seedlings. In S. S. Mulkey, R. L. Chazdon & A. P. Smith (Eds.), Tropical forest plant ecophysiology (pp. 559-596). NewYork: Chapman & Hall. [ Links ]
Kitajima, K. (2002). Do shade-tolerant tropical tree seedlings depend longer on seed reserves? Functional growth analysis of the Bignoniaceae species. Functional Ecology, 16, 433-444. [ Links ]
Kitajima, K. (2003). Impact of cotyledon and leaf removal on seedling survival in three tree species with contrasting cotyledon functions. Biotropica, 35(3), 429-434. [ Links ]
Loescher, H. W., Oberbauer, S. F., Gholz, H. L. & Clark, D. B. (2003). Environmental controls on net ecosystem-level carbon exchange and productivity in a Central American tropical wet forest. Global Change Biology, 9(3), 396-412. [ Links ]
Marasco, J., Schnitzer, S. A. & Carson, W. P. (2004). Liana diversity, abundance, and mortality in a tropical wet forest in Costa Rica. Forest Ecology and Management, 190, 3-14. [ Links ]
Muhammad, A. F. & Sattler, R. (1982). Vessel structure of Gnetum and the origin of Angiosperms. American Journal of Botany, 69(6), 1004-1021. [ Links ]
Mulkey, S. S., Wright, S. J. & Smith, A. P. (1993). Comparative physiology and demography of three Neotropical forest shrubs: alternative shade-adaptive character syndromes. Oecologia, 96(4), 526-536. [ Links ]
Nabe-Nielsen, J. (2002). Growth and mortality rates of the liana Machaerium cuspidatum in relation to light and topographic position. Biotropica, 34(2), 319-322. [ Links ]
Osunkoya, O. O., Ash, J. E., Hopkins, M. S. & Graham, A. W. (1994). Influence of seed size and seedling ecological attributes on shade-tolerance of rain-forest tree species in Northern Queensland. Journal of Ecology, 82(1), 149-163. [ Links ]
Onguene, N. A. & Kuyper, T. W. (2001). Mycorrhizal associations in the rain forest of South Cameroon. Forest Ecology andManagement, 140(3), 277-287. [ Links ]
Phillips, O. L., Vásquez-Martínez, R., Monteagudo-Mendoza, A., Baker, T. R. & Núñez-Vargas, P. (2005). Large lianas as hyperdynamic elements of the tropical canopy. Ecology, 86(5), 1250-1258. [ Links ]
Poorter, L. (2001). Light-dependent changes in biomass allocation and their importance for growth of rain forest tree species. Functional Ecology, 15(1), 113-123. [ Links ]
Poorter, L., Bongers, F., Sterck, F. J. & Wöll, H. (2005). Beyond the regeneration phase: differentiation of height-light trajectories among tropical tree species. Journal of Ecology, 93(2), 256-267. [ Links ]
Poorter, L. & Rose, S. A. (2005). Light-dependent changes in the relationship between seed mass and seedling traits: ameta-analysis for rain forest tree species. Oecologia, 142(3), 378-387. [ Links ]
Putz, F. E. (1984). The natural history of lianas on Barro Colorado Island, Panama. Ecology, 65(6), 1713-1724. [ Links ]
Quinn, G. P. & Keough, M. J. (2002). Experimental design and data analysis for biologists. Cambridge: Cambridge University Press. [ Links ]
Rice, W. R. (1988). Analyzing tables of statistical tests. Evolution, 43(1), 223-225. [ Links ]
Rodin, R. J. & Kapil, R. N. (1969). Comparative anatomy of the seed coats of Gnetum and their probable evolution. American Journal of Botany, 56(4), 420-431. [ Links ]
Sanches, M. C. & Válio, I. F. M. (2002). Seedling growth of climbing species from a southeast Brazilian tropical forest. Plant Ecology, 159(1), 51-59. [ Links ]
Sanches, M. C. & Válio, I. F. M. (2008). Photosynthetic response of two tropical liana species grown under different irradiances. Photosynthetica, 46(4), 557-566. [ Links ]
Sanford, R., Paaby, P., Luvall, J. C. & Phillips, E. (1994). Climate, geomorphology and aquatic systems. In L. A. McDade, K. S. Bawa, H. A. Hespenheide & G. S. Hartshorn (Eds.), La Selva: Ecology and Natural History of a Neotropical Rain Forest (pp. 19-33). Chicago: University of Chicago Press. [ Links ]
SAS. (1994). JMP user’s guide. Cary, North Carolina: SAS Institute. [ Links ]
Schnitzer, S. A. & Bongers, F. (2002). The ecology of lianas and their role in forests. Trends in Ecology and Evolution, 17(5), 223-230. [ Links ]
Schnitzer, S. A. & Bongers, F. (2011). Increasing liana abundance and biomass in tropical forests: emerging patterns and putative mechanisms. Ecology Letters, 14(4), 397-406. [ Links ]
Schnitzer, S. A. & Carson, W. P. (2010). Lianas suppress tree regeneration and diversity in treefall gaps. Ecology Letters, 13(7), 849-857. [ Links ]
Shutov, A. D., Braun, H., Chesnokov, Y. V., Horstmann, Ch., Kakhovskaya, I. A. & Bäumlein, H. (1998). Sequence peculiarity of Gnetalean legumin-like seed storage proteins. Journal of Molecular Evolution, 47(4), 486-492. [ Links ]
Thompson, W. P. (1916). The morphology and affinities of Gnetum. American Journal of Botany, 3(4), 135-184. [ Links ]
Toledo-Aceves, T. & Swaine, M. D. (2008). Biomass allocation and photosynthetic responses of lianas and pioneer tree seedlings to light. Acta Oecologica, 34(1), 38-49. [ Links ]
Valladares, F. & Niinemets, Ü. (2008). Shade tolerance, a key plant feature of complex nature and consequences. Annual Review of Ecology, Evolution, and Systematics, 39(1), 237-257. [ Links ]
*Correspondencia: Gerardo Celis: School of Natural Resources and Environment, University of Florida,103 Black Hall, P.O. Box 116455, Gainesville, FL 32611 USA; celis@ufl.edu
Gerardo Avalos: Escuela de Biología, Universidad de Costa Rica, 11501-2060San Pedro, San José, Costa Rica; The School for Field Studies, Center for Sustainable Development Studies, 100 Cummings Center Suite 534-G, Beverly, Massachusetts 01915-6239 USA gerardo.avalos@ucr.ac.cr
1. School of Natural Resources and Environment, University of Florida,103 Black Hall, P.O. Box 116455, Gainesville, FL 32611 USA; celis@ufl.edu
2. Escuela de Biología, Universidad de Costa Rica, 11501-2060San Pedro, San José, Costa Rica; gerardo.avalos@ucr.ac.cr
3. The School for Field Studies, Center for Sustainable Development Studies, 100 Cummings Center Suite 534-G, Beverly, Massachusetts 01915-6239 USA
Received 26-XI-2012. Corrected 20-VI-2013. Accepted 22-VII-2013.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}