










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.61 n.4 San José Oct./Dec. 2013
Population biology of the mangrove crab Ucides cordatus (Decapoda: Ucididae) in an estuary from semiarid Northeastern Brazil
Biología poblacional del cangrejo de manglar Ucides cordatus (Decapoda: Ucididae) en un estuario tropical semiárido
Biología poblacional del cangrejo de manglar Ucides cordatus (Decapoda: Ucididae) en un estuario tropical semiárido
*Dirección para correspondencia:
Abstract
The mangrove crab Ucides cordatus is an important resource of estuarine regions along the Brazilian coast. U. cordatus is distributed from Florida, USA, to the coast of Santa Catarina, Brazil. The species plays an important role in processing leaf litter in the mangroves, which optimizes the processes of energy transfer and nutrient cycling, and is considered a keystone species in the ecosystem. Population declines have been reported in different parts of the Brazilian coast. In the present study we evaluated aspects of the population structure, sex ratio and size at morphological sexual maturity. We analyzed 977 specimens collected monthly over 24 months (2010-2012), in a mangrove of the Jaguaribe River, in the municipality of Aracati on the East coast of Ceará state, Northeastern Brazil. The study area has a mild semiarid tropical climate, with mean temperatures between 26 and 28oC. The area is located within the eco-region of the semiarid Northeast coast, where mangroves occur in small areas and estuaries are affected by mesomareal regimes. The population structure was evaluated by the frequency distribution of size classes in each month, and the overall sex ratio was analyzed using the chi-square test. Size at morphological sexual maturity was estimated based on the allometry of the cheliped of the males and the abdomen width of the females, using the program REGRANS. The size-frequency distribution was unimodal in both sexes. The overall sex ratio (M: F) (1:0.6) was significantly different from 1:1. Analysis of the sex ratio by size class showed that the proportion of males increased significantly from size class 55-60mm upward, and this pattern persisted in the larger size classes. In the smaller size classes the sex ratio did not differ from 1:1. The size at morphological sexual maturity was estimated at a carapace width (CW) of 52mm and 45mm for males and females, respectively. Analysis of the population parameters indicated that the population of U. cordatus in the Jaguaribe River mangrove is stable. However, constant monitoring of the population is required to detect any changes in the population attributes that may affect this stability. Rev. Biol. Trop. 61 (4): 1721-1735. Epub 2013 December 01.
Key words: allometry, population structure, fishery management, sex ratio, size at sexual maturity.
Resumen
El cangrejo Ucides cordatus es un importante recurso pesquero de las regiones de estuarios a lo largo de la costa brasileña. Disminuciones de las poblaciones han sido reportadas en diferentes regiones del país. El presente estudio evaluó aspectos de la estructura poblacional, proporción sexual y talla en la madurez sexual. Se analizó un total de 977 ejemplares capturados mensualmente, por 24 meses, en lo manglar del río Jaguaribe, en el municipio de Aracati, en la costa este de Ceará, nordeste del Brasil. El área de estudio tiene un clima semiárido tropical, con temperaturas medias entre los 26 y 28°C. La estructura poblacional fue evaluada para determinar la distribución de frecuencia de clases de talla. Para analizar diferencias significativas respecto a la proporción sexual se aplicó la prueba Chi-quadrado. La talla de la madurez sexual morfológica fue estimada con base en la alometría del propodito del quelípedo de los machos y de la anchura del abdomen de las hembras, utilizando el programa REGRANS. La distribución de frecuencia de talla fue unimodal para los dos sexos. La proporción sexual total (M:H)(1:0.6), se desvió significativamente de la igualdad. El análisis de la proporción entre sexos, relativo a las clases de tallas mostró que la proporción de machos se tornó significativamente más alta desde la clase de 55-60mm de talla, y este se mantuvo en las clases de talla más grande. No obstante, en las clases más pequeñas la proporción entre los sexos no fue distinta de 1:1. La talla de la madurez sexual morfológica fue de 52mm de ancho del caparazón y 45mm para los machos y hembras, respectivamente. Los resultados de los atributos analizados permitieran deducir que la población de U. cordatus en el manglar del río Jaguaribe se encuentran en una situación de estabilidad. Sin embargo, el monitoreo constante de la población es necesario para detectar alteraciones en los atributos de la población que puedan afectar esta situación de estabilidad.
Palabras clave: alometría, estructura poblacional, manejo pesquero, proporción sexual, talla de madurez sexual
The study of population biology furthers our understanding of the ecological stability of a species in a particular habitat (Castiglioni et al., 2006). Population parameters of brachyuran crabs have been investigated by analyzing the distribution of individuals in size class, sex ratio, range in size at sexual maturity, reproductive cycle, fecundity, and juvenile recruitment (Thurman, 1985; Díaz & Conde, 1989; Flores & Negreiros-Fransozo, 1999; Conde et al., 2000; Cobo, 2005; Litulo, 2005; Benetti et al., 2007; Bezerra & Mathews-Cascon, 2007; Kassuga et al., 2008; Gregati & Negreiros-Fransozo, 2009; Hartnoll et al., 2009; Castiglioni et al., 2011; Araújo et al., 2012).
The size-frequency distribution of a population is a dynamic characteristic that can change during the year, as a result of reproduction and larval recruitment (Thurman, 1985). In crab populations of tropical regions, the distribution of individuals in size classes has been found to be unimodal (Díaz & Conde, 1989; Diele et al., 2005; Litulo, 2005; Fernandes-Góes et al., 2005; Gregati & Negreiros-Fransozo, 2009), suggesting stable populations with constant rates of mortality and recruitment (Hartnoll & Bryant, 1990).
In natural populations, the proportion of males and females at birth tends to be 1:1 (Fisher 1930). After birth, many factors can affect this relationship, such as mortality rates, longevity and growth, predation, migration, utilization of different habitats by two sexes (Wenner, 1972; Hartnoll et al., 2009), and processes related to reproductive events (Gregati & Negreiros-Fransozo, 2009).
The concept of age/size at the onset of sexual maturity is essential for understanding the reproductive strategy and fitness of individuals of a species, as factors determining the reproductive potential (McQuaid et al., 2006; Castiglioni & Coelho, 2011). Sexual maturity is an important regulator of population dynamics, influencing demography and recruitment rates (Gerhart & Bert, 2008).
Physiological and morphological changes that enable the appearance of secondary sexual traits responsible for the transition from the juvenile to adult phase characterize the onset of sexual maturity in crustaceans (Hartnoll, 1978). In this case, early maturity in brachyurans can be detected by changes in the growth proportions (relative or allometric) of structures associated with secondary sexual characteristics such as the cheliped in males and the abdomen width in females (Hartnoll, 1974; Hartnoll, 1982; Pinheiro & Fransozo, 1998). Its onset occurs when an individual reaches a certain size, and can vary greatly both among populations of the same species and among individuals within a population (Fonteles-Filho, 1989). Size at sexual maturity is a key parameter for the development of management plans for commercially exploited species (González-Gurriarán & Freire, 1994; Pinheiro & Fransozo, 1998; McQuaid et al., 2006; Rostant et al., 2008), as it allows the establishment of a minimum size at capture (Lizzárraga-Cubedo et al., 2003; Corgos & Freire, 2006).
The mangrove crab Ucides cordatus is distributed from Florida, USA to the coast of Santa Catarina, Brazil (Melo, 1996). It inhabits the mangroves of the intertidal zone, where it digs burrows in the sediment, each occupied by a single individual (Costa, 1972; Alcântara-Filho, 1978; Nascimento, 1993; Góes et al., 2010). The species plays an important role in processing leaf litter in mangroves, which optimizes the processes of energy transfer and nutrient cycling (Nordhaus & Wolff, 2007). Therefore, where it occurs, U. cordatus is considered a keystone species in the ecosystem (Schories et al., 2003). Besides, this species has a seasonal breeding period, which along the Brazilian coast is between December and April (Ivo & Gesteira, 1999).
U. cordatus has significant economic importance as a fishery resource in estuarine regions, principally in Northeast Brazil, and its exploitation involves an extensive production chain (Ibama, 1994; Ivo & Gesteira, 1999; Glaser & Diele, 2004). The fishery for this crustacean is selective by size and sex. In general, females and specimens with a carapace width less than six centimetres are rejected in favour of larger specimens, preferably males (Botelho et al., 2000; Paiva, 1997; Passos & Di Beneditto, 2005; Leite et al., 2006; Fernandes & Carvalho, 2007). Declines in U. cordatus populations have been reported in many coastal regions of Brazil (Diele et al., 2005, Legat et al., 2005; Mendonça & Pereira, 2009), and currently, this crustacean is included in the list of species threatened by overexploitation (MMA - Ministério do Meio Ambiente, 2004).
In the area evaluated in this study, populations of U. cordatus are potentially subject to impact factors related to dams, shrimp aquaculture and fishing, as well as weather and typical climatic adversities such as scarce and irregular rainfall distribution. These characteristics can potentially affect biological traits of populations (Marins et al., 2003). The study aimed to generate data as a basis for monitoring and conservation of this population, following an approach based on Life History Theory, focusing on size structure, sex ratio and size at sexual maturity. This population biology approach yielded useful information for the implementation of conservation measures for this crab.
Materials and methods
Study site: The study area is located in a mangrove of Jaguaribe River (4°26’15’’ S - 37°48’45’’ W) in the municipality of Aracati on the Rast coast of Ceará state, Northeastern Brazil. The Jaguaribe River basin is divided into five sub-basins, drains a total area of 72.043km2 and is approximately 633km long (Marins et al., 2003). The estuarine zone is covered by mangrove over an area of 11.64km2 (Semace, 2006). The area is located within the eco-region of the semiarid Northeast coast, where mangroves occur in small areas and estuaries are affected by mesomareal schemes (2-4m) (Semace, 2006). The region has a mild semi-arid tropical climate, with mean temperatures between 26 and 28oC (Ipece, 2010). Rainfall is distributed in two distinct seasons, although there are exceptionally dry or wet years (Marins et al., 2003). The period of greatest rainfall is January to May, with a mean maximum of 237.8mm in March. From June to December, mean rainfall does not exceed 47.7mm, with the mean minimum of 2.4mm occurring in September.
Sampling: Specimens of U. cordatus were collected monthly from July 2010 through June 2012, during periods of low tide. Traps called “forjo” were used to capture the crabs. In the Jaguaribe River, the forjo is made out of a 2L plastic bottle. These traps are placed so as to fully close the entrance of the crab burrows, and leaves of the red mangrove Rhizophora mangle are used as bait (Carvalho & Igarashi, 2009). Several areas of the mangrove, used as fishing sites by the local community, were selected for the collection of crabs. At each site, we used 50 forjos. The traps were placed during the afternoon and collected the following morning, after approximately 12h. Individuals collected were killed by temperature shock at 3oC in a container of ice. They were then transported to the laboratory where the sex was recorded by inspecting the morphology of the abdomen (Pinheiro & Fiscarelli, 2001). The length and width of the carapace were measured using a caliper of accuracy 0.05mm.
Population structure: The size structure of the population (males and females) was analyzed as a function of the frequency distribution of individuals, both for the total number of crabs collected during the study period (Litulo, 2005), and grouped by seasons (dry and rainy). The data for the dry season were grouped considering the period from July through December 2010 and 2011, and the rainy season from January through June in 2011 and 2012.
The values for carapace width were allocated to size classes by means of a frequency distribution, in accordance with the method proposed by Sturges (1926). The normality of the frequency-distribution data was assessed using the Kolmogorov-Smirnov test (Zar, 2010). The mean size of males and females was compared using the Mann-Whitney U test. The sex ratio for all specimens, and the variation according to month and size class were compared. Deviation from the expected 1:1 ratio was assessed using the chi-square test (Zar, 2010).
Size at sexual maturity:To estimate size at sexual maturity, morphological criteria were used, i.e., the morphometric relationships between the length of the propodus of the major cheliped (y) and the carapace width (x) in males (PL x CW); and the abdomen width (y) and carapace width (x) in females (AW x CW) (Leite, 2005; Leite et al., 2006). The abdomen of females was measured between the fourth and fifth somite. These measurements were chosen taking into account the importance of the variables PL and AW in the reproductive process of males and females, respectively (Hartnoll, 1982; Castiglioni & Coelho, 2011). Individuals with missing appendages or injuries, or males lacking evident heterochely were excluded from the analysis. The empirical points of the relationships (PL x CW) and (AW x CW) were submitted to a regression analysis, with adjustment by the least-squares method. Several criteria have been suggested to analyze the fit of regression equations to a set of data. This study employed the F test and graphical examination of the standardized residuals (Draper & Smith, 1966).The F test was used to verify whether the projections generated by the regression differed significantly from the empirical data obtained. The hypotheses tested were:
• H0: the data projected by the model do not differ from the empirical data
• H1: the data projected by the model differ from the empirical data
• The graphical examination of the standard residuals was performed by visual inspection (Bussab & Morettin, 2003; Zar, 2010).
Size at morphological maturity of U. cordatus was detected by changes in the pattern of relative growth of the morphometric variables analyzed, where b is the slope of the regression (constant allometry) (Hartnoll, 1982). The allometry patterns were established according to the following criteria: b=1 (isometry), b>1 (positive allometry), b<1 (negative allometry) (Hartnoll, 1982). To detect the b-difference from unity, an interval of 0.90-1.10 was used for b (Kurata, 1962; Kuris et al., 1987; Clayton 1990; Mantelatto & Martinelli, 2001; Fransozo et al., 2003). The inflection point (changes in the slope of the regression lines of the morphometric relationships) was sought iteratively using an algorithm in the REGRANS program (Pezzuto, 1993). The algorithm searches for the carapace width value at which data can be separated into two subsets (immature and mature individuals) (Sant´Ana & Pezzuto, 2009). Thus, the value obtained corresponds to the size at morphological sexual maturity. The difference in the slope of the regressions between immature and mature individuals of both sexes was tested using the Student t test for comparison of angular coefficients (Ivo & Fonteles-Filho, 1997; Zar, 2010). All statistical analyses were performed at the 5% significance level.
Results
Population structure: During the study, 977 crabs were collected: 594 males (60.8%) and 383 females (39.2%). Carapace width (CW) ranged from 20.45 to 80.55mm (mean=57.32±11:51mm) in males and from 26.55mm to 71.12mm (mean=51.62±11.67mm) in females. Males were significantly larger than females (Mann-Whitney U-test=7.49, p<0.0001).
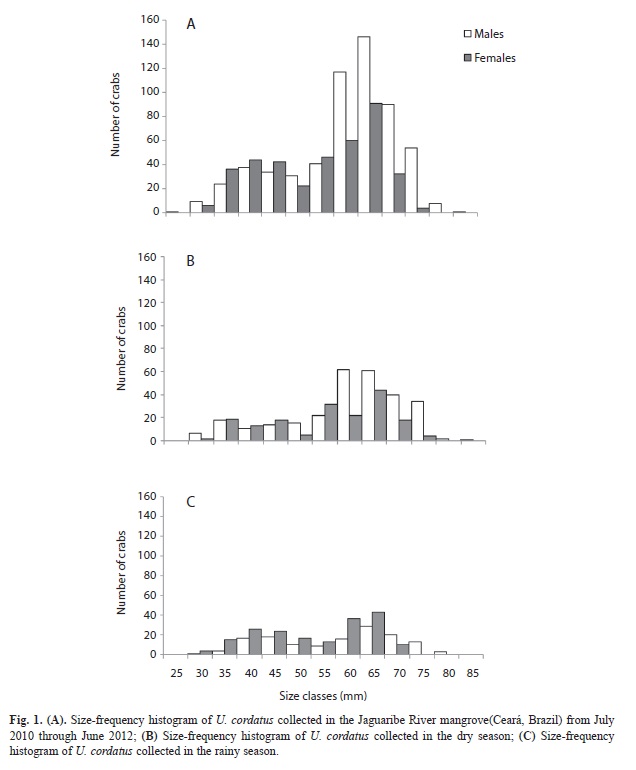
Size did not have a normal frequency distribution, either in males (KS-test=0.13291, p<0.01) or in females (KS-test=0.1094, p<0.01). Size-frequency histograms showed a unimodal distribution. The modal class was found in the range of 60-65mm CW for both sexes (Fig. 1A). In the dry and rainy seasons, the size structure of the population was similar, with only a reduction in the abundances of the modal 60-65mm class for both males and females, in both seasons (Fig. 1B and C).
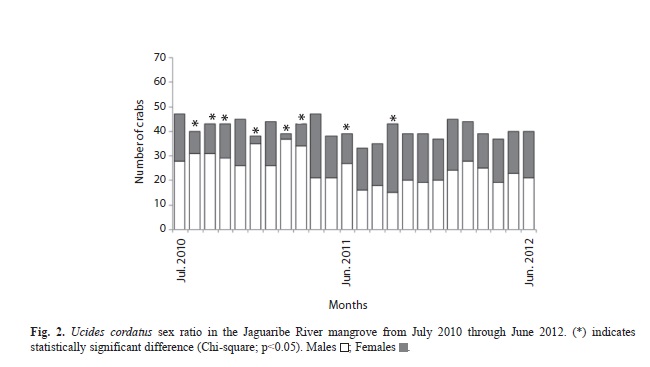
The overall sex ratio (1M: 0.6F) deviated significantly from the expected ratio of 1:1 (Chi-square=45.6, p<0.0001). However, in most months (67%) the ratio did not differ from 1:1. The sex ratio was skewed toward males in 29% of cases and toward females in only 4% (Fig. 2).
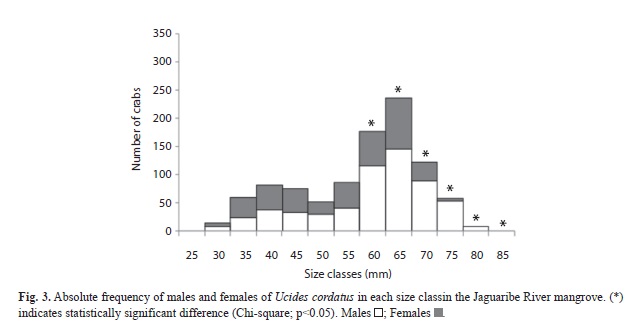
The analysis of sex ratio by size class showed that the ratio of males becomes significantly higher from size class 55-60mm upward, and this pattern persisted in the subsequent larger size classes. However, in the smaller size classes the sex ratio did not differ from 1:1 (Fig. 3).
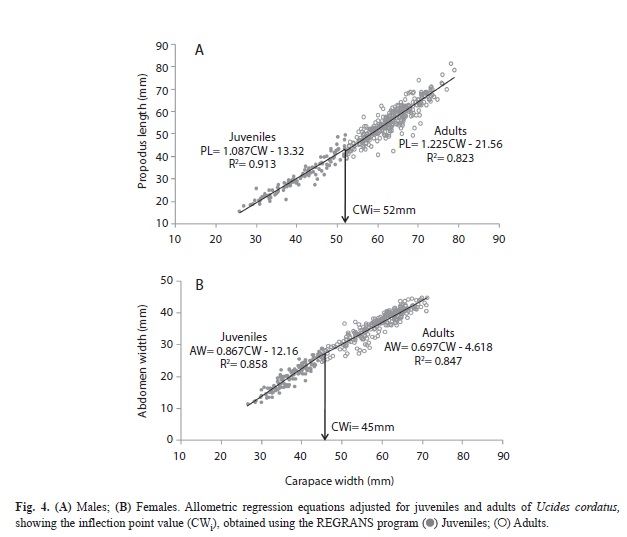
Size at sexual maturity: The inflection points of the carapace width (CW), calculated by the REGRANS program, were 52mm and 45mm for males and females, respectively. These CW values delimited the juveniles (immature, n=118) and adults (mature, n=326), indicating the onset of morphological sexual maturity. The cheliped propodus length (PL) of immature males showed an isometric growth pattern (b=1.087), while adults exhibited positive allometry (b=1.225) (Fig. 4A). In males, the t-test showed significant differences in the slopes of the regression lines between juveniles and adults (t-test=5.35, p<0.0001). Although female juveniles (immature, n=126) and adults (mature, n=251) both showed negative allometry with respect to abdomen width (AW), adults had a significantly lower growth rate (b=0.697) than juveniles (b=0.897) (t-test=-4.78, p<0.0001) (Fig. 4B).
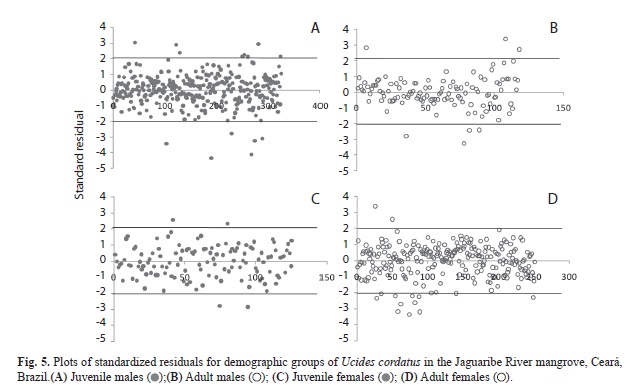
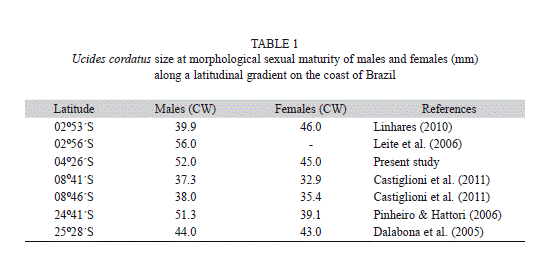
The F test indicated homoscedasticity of the regressions for males (juveniles: Fcal: 0.623; Fcrit: 1.53; df: 117; adults: Fcal: 0.079; Fcrit: 1.00; df: 325) and females (juveniles: Fcal: 0.395; Fcrit: 1.25; gl: 125; adults: Fcal: 0.192; Fcrit: 1.00; df: 250). Thus, the linear model provided the best fit to the data. The plot of standardized residuals showed that in the regression of juvenile males, a total of 14 points were above or below the analysis interval. Thus, the dispersion represented 12% of the total estimates, which means that 88% of the residuals fell within the analysis interval (+2, -2) (Fig. 5A). For adult males, 98% of the residuals fell within the analysis interval (Fig. 5B). For juvenile females, only four points fell above or below the analysis interval, indicating that the data dispersion represented only 3% of the estimated total. Thus, 70% of the residuals were delimited by the interval (Fig. 5C). In adult females, 6% of the data were outside the ranges. Therefore, 94% of the residuals fell within the range analyzed (Fig. 5D). Table 1 illustrates the differences in size at sexual maturity of U. cordatus in relation to a latitudinal gradient.
The size-frequency distribution of both sexes was unimodal. This distribution pattern has been observed for other crab species (Thurman, 1985; Díaz & Conde, 1989; Spivak et al., 1991; Cobo, 2005; Litulo, 2005; Castiglioni et al., 2006; Gregati & Negreiros-Fransozo, 2009; Castiglioni et al., 2011), as well as for U. cordatus (Diele et al., 2005). A unimodal distribution suggests a stable population with constant mortality and recruitment rates (Litulo, 2005). In this study, males and females were most frequently found in the 60-65mm CW size class. The predominance of individuals in intermediate size classes is a function of the decrease in growth rate with age (Diele et al., 2005). According to these authors, in U. cordatus the dominance of individuals of intermediate and larger sizes is common in populations with greater longevity. This pattern has also been described in other crab species (Leme, 2002; Gregati & Negreiros-Fransozo, 2009; Hartnoll et al., 2009).
Mean size was significantly higher in males than in females, indicating the presence of sexual dimorphism. Sexual dimorphism in size can be considered the natural state for U. cordatus populations (Diele & Koch, 2010), as this pattern has been observed in the same species by several investigators along the Brazilian coast (Alcântara-Filho, 1978; Ivo et al., 1999; Vasconcelos et al., 1999; Alves & Nishida, 2004; Leite, 2005; Marques, 2008; Castro et al., 2008). Life-history theory predicts that somatic growth and reproduction are antagonistic processes which compete for limited resources (Gadgil & Bossert, 1970; Calow, 1979; Glazier, 1999; Lika & Kooijman, 2003; Wada et al., 2008). In crustaceans, more resources are directed to reproduction in females, due to processes such as gonad maturation and maintenance of oxygen flow to the egg mass (Hartnoll, 2006; Hartnoll et al., 2009), these resources then become unavailable for growth. Thus, it is suggested that the sexual dimorphism in U. cordatus is due to the negative relationship (trade-off) between reproduction and growth. The occurrence of this pattern indicates that the population analyzed is not under severe fishing pressure (Diele & Koch, 2010).
The sex ratio observed for U. cordatus, including the complete data set, was significantly skewed toward males. Similar observations for this species have been reported by others (Alves & Nishida, 2004; Diele et al., 2005; Castro et al., 2008; Marques, 2008). In general, several factors may contribute to a deviation from a 1:1 ratio, such as migration, differential longevity of each sex, using different areas of the habitat, and differences in growth and mortality rates between sexes (Wenner, 1972; Johnson, 2003). For size classes, males were predominant from the range of 55-60mm upward. Females of some crab species have a tendency to accumulate in the intermediate size classes, because they have a smaller maximum size and a slower growth rate (Diele et al., 2005), caused by the high energy costs of reproduction (Warner, 1967; Hartnoll & Gould, 1988). In contrast, males spend less time in intermediate size classes, having faster growth rates at these size ranges (Diele et al., 2005).
Male-biased selectivity in exploited species such as U. cordatus can also change the sex ratio (Fenberg & Roy, 2007). According to Diele et al. (2005), areas with a longer history of exploitation have a higher proportion of females, and this deviation may indicate that the population has been overfished (Conti & Nalesso, 2010; Sforza et al., 2010). The results obtained here suggest no evidence of overexploitation.
There were significant differences in the proportion of cheliped growth, with juveniles showing isometric growth, and adults showing positive allometry. Differences in the levels of allometry of this appendage during ontogeny have been previously observed for this species (Dalabona et al., 2005; Leite et al., 2006; Pinheiro & Hattori, 2006; Linhares, 2010), as well as for other brachyurans (Clayton & Snowden, 1991; Gardner & Williams, 1992; Pinheiro & Fransozo, 1998; Corgos & Freire, 2006). This cheliped growth pattern has a functional significance for some species of Brachyura, since the chelipeds are used in foraging, territorial defense, courtship and copulation. Besides conferring a selective advantage for mating, they are a necessary requirement for reproductive success (Hartnoll, 1974; Lee, 1995; Pinheiro & Fransozo, 1998). The F test,as well as the examination of residuals, indicated the occurrence of homoscedasticity of the data for males and females, as the residuals were distributed evenly above and below zero (Zar, 2010). This validated the use of a linear model and, therefore, there was no necessity for a logarithmic transformation (Zar, 2010). In some crustacean species, morphometric relationships used to detect the size at sexual maturity by variations in the allometric growth, also have been adjusted by linear models (Streissl & Hödl, 2002; Gerhart & Bert, 2008).
Although both juvenile and adult females exhibited negative allometric growth of the abdomen, a tendency for decreased growth rates was evident in this segment in the adults, in agreement with other studies on this species (Dalabona et al., 2005; Leite 2005; Pinheiro & Hattori, 2006; Linhares 2010). The female abdomen has a functional role in protecting and incubating eggs (Hartnoll, 1974). The trend to a reduction in allometry at the transition to the adult phase is due to this region of the body already having reached adequate proportions for the protection of eggs without compromising female locomotion (Hartnoll, 1982). Therefore, allometric variations during ontogeny may be associated with the reproductive strategies of a species, with these variations reflecting differential investment of resources in growth (Charnov et al., 2001).
Males of U. cordatus reached morphological sexual maturity when they were approximately 15% larger than females, a trend similar to that observed for this species by other investigators (Dalabona et al., 2005; Pinheiro & Hattori, 2006; Castiglioni et al., 2011). Females can mature at smaller sizes than males, possibly due to requirements related to the costs of reproduction (Stearns, 1989; Hartnoll, 2006) that generate a trade-off with somatic growth (Gadgil & Bossert, 1970; Calow, 1979; Glazier, 1999; Lika & Kooijman, 2003; Wada et al., 2008). The maturation of males at a larger size than females may confer a reproductive advantage, as it assists them in manipulating females, with the aid of appendages, during courtship and mating (Corgos & Freire, 2006), as well as enabling copulation with females of a wider size range (Pinheiro & Fiscarelli, 2001). Smaller male than female size at sexual maturity has been reported for some populations of U. cordatus (Mota-Alves, 1975; Linhares, 2010). A reduction in size/age at maturity is considered to be an indicator of stress caused by fishing (Trippel, 1995). Thus, the occurrence of the onset of maturity at a larger size in males than in females indicates that this population may not yet be subject to stress caused by fishing.
Changes in this life-history trait can be explained by considering two hypotheses: the “Compensatory response” (Trippel, 1995) and the “Evolutionary response” (Borisov, 1978; Law & Grey, 1989; Law, 2000). These two hypotheses are not mutually exclusive: changes can have both environmental and genetic components (Heino & Godo, 2002). The first hypothesis predicts that an individual can reach sexual maturity more quickly because of the decrease in competition for resources, which increases their growth rates. Thus, early maturation would be associated with phenotypic plasticity resulting from a response to a change in the environment (Engelhard & Heino, 2004). The second predicts that early maturation is due to genetic changes arising from the effects of selective fishing. Intense selective pressure can alter genetic diversity, causing the loss of some genotypes due to directional selection (Heino & Godo, 2002). Thus, individuals with a tendency to mature when they are younger or smaller would be favoured and contribute more of their genes to the next generation than individuals with a tendency to mature when they are larger or older (Law, 2000; Heino & Godo, 2002). This mechanism of size reduction allows the first reproductive event to occur before the individual becomes vulnerable to factors causing mortality (Stearns, 2000).For purposes of management it is important to understand whether changes in this life-history trait are due to phenotypic plasticity or genetic factors, because in the former case the changes are in principle reversible, but in the latter they are not (Olsen et al., 2005).
Latitudinal variations in size at the onset of morphological sexual maturity were observed in U. cordatus. According to Hines (1989), the causes of geographic variation may be a factor of temperature and its interaction with photoperiod. Density-dependent factors, as well as food availability can cause metabolic changes with effects on growth rates, leading to an advance or delay in size at maturity (Wenner et al., 1974; Hines, 1989). Alterations in this variable can also be attributed to the action of different stressors on populations, such as fishing pressure (McQuaid et al., 2006), or result from variations or inaccuracies in the methods employed (Souza-Carvalho et al., 2011).
The results obtained for the population features analyzed, such as the unimodal size structure, the occurrence of sexual dimorphism, male-biased sex ratio, and larger size at maturity in males, allow us to infer that this population of U. cordatus does not show compelling evidence of overexploitation. This implies that the population status is stable. However, constant monitoring is necessary in order to detect any changes in the population features that may alter this status.
Acknowledgments
The first author thanks CAPES (Coordenação de Aperfeiçoamento de Pessoal de Nível Superior) for providing a PhD scholarship. We are grateful to the fisherman Ronaldo Gonzaga da Silva for his help in fieldwork, and to Kate Ingram for translating the text. All sampling in this study was conducted in compliance with current applicable federal laws (ICMBio 20240-1).
References
Alcântara-Filho, P. (1978). Contribuição ao estudo da biologia do caranguejo-uçá Ucides cordatus cordatus (Linnaeus, 1763) (Crustacea, Decapoda, Brachyura), no manguezal do rio Ceará (Brasil). Arquivos de Ciências do Mar, 18: 1-41. [ Links ]
Alves, R. R. N., & Nishida, A. K. (2004). Population structure of the mangrove crab Ucides cordatus (Crustacea: Decapoda: Brachyura) in the estuary of the Mamanguape River, northeast Brazil. Tropical Oceanography, 32: 23-37. [ Links ]
Araújo, M. S. L., Barreto, A. V., Negromonte, A. O., & Schwamborn, R. (2012). Population ecology of the blue crab Callinectes danae (Crustacea: Portunidae) in a Brazilian tropical estuary. Anais da Academia Brasileira de Ciências, 84: 129-138. [ Links ]
Benetti, A. S., Negreiros-Fransozo, M. L., & Costa, T. M. (2007). Population and reproductive biology of the crab Uca burgesi (Crustacea: Ocypodidae) in three subtropical mangrove forests. Revista de Biología Tropical, 55: 55-70. [ Links ]
Bezerra, L. E. A., & Mathews-Cascon, H. (2007). Population and reproductive biology of the fiddler crab Uca thayeri Rathbun, 1900 (Crustacea: Ocypodidae) in a tropical mangrove from Northeast Brazil. Acta Oecologica, 31: 251-258. [ Links ]
Borisov, V. M. (1978). The selective effect of fishing on the population structure of species with long life cycle. Journal of Ichthyology, 18: 896-904. [ Links ]
Botelho, E. R. O., Santos, M. C. R., & Pontes, A. C. P. (2000). Algumas considerações sobre a redinha na captura do caranguejo-uçá Ucides cordatus (Linnaeus, 1763) no litoral sul de Pernambuco-Brasil. Boletim Técnico Científico CEPENE, 8: 55-71. [ Links ]
Bussab, W. O., & Morettin, P. A. (2003). Estatística Básica. São Paulo: Editora Saraiva. [ Links ]
Calow, P. (1979). The cost of reproduction – a physiological approach. Biological Review, 54: 23-40. [ Links ]
Carvalho, H. R. L., & Igarashi, M. A. (2009). A utilização do forjo na captura do caranguejo-uçá (Ucides cordatus) na comunidade de Tapebas em Fortaleza-CE. Biotemas, 22: 69-74. [ Links ]
Castiglioni, D. S., & Coelho, P. A. (2011). Determinação da maturidade sexual de Ucides cordatus (Decapoda, Brachyura, Ucididae) em duas áreas de manguezal do litoral sul de Pernambuco, Brasil. Iheringia, 101: 138-144. [ Links ]
Castiglioni, D. S., Negreiros-Fransozo, M. L., & Mortari, R. C. (2006). Biologia populacional do caranguejo violinista Uca rapax (Smith, 1870) (Crustacea, Ocypodoidea), proveniente de uma área de manguezal degradado em Paraty, RJ, Brasil. Atlântica, 28: 73-86. [ Links ]
Castiglioni, D. S., Oliveira, P. J. A., Silva, J. S., & Coelho, P. A. (2011). Population dynamics of Sesarma rectum (Crustacea: Brachyura: Grapsidae) in Araquindá River mangrove, north-east of Brazil. Journal of the Marine Biological Association of the United Kingdom, 91: 1394-1401. [ Links ]
Castro, A. C. L., Correia, M. M. F., Nascimento, A. R., Piedade-Júnior, R. N., Gama, L. R. M., Sousa, M. M., Sena, A. C. S., & Sousa, R. C. C. (2008). Aspectos bioecológicos do caranguejo-uçá (Ucides cordatus cordatus L. 1763) (Decapoda, Brachyura) nos manguezais da ilha de São Luís e litoral oriental do Estado do Maranhão, Brasil. Amazônia: Ciência & Desenvolvimento, 3: 17-36. [ Links ]
Charnov, E. L., Turner, T. F., & Winemiller, K. O. (2001).Reproductive constraints and the evolution of life histories with indeterminate growth. Proceedings of the National Academy of Sciences, U.S.A, 98: 9460-9464. [ Links ]
Clayton, D. A., & Snowden, R. J. (1991). Allometric growth in Ilyoplax stevensi (Decapoda, Brachyura, Ocypodidae). Crustaceana, 61: 2-9. [ Links ]
Clayton, J. C. (1990). Crustacean alometric growth: a case for caution. Crustaceana, 58: 570-590. [ Links ]
Cobo, V. J. (2005). Population biology of the spider crab Mithraculus forceps (A. Milne-Edwards, 1875) (Majidae, Mithracinae) on the southeastern Brazilian coast. Crustaceana, 78: 1079-1087. [ Links ]
Conde, J. E., Tognella, M. M. P., Paes, E. T., Soares, M. L. G., Louro, I. A., & Schaeffer-Novelli, Y. (2000). Population and life history features of the crab Aratus pisonii (Decapoda: Grapsidae) in a subtropical estuary. Interciencia, 25: 151-158. [ Links ]
Conti, R. C., & Nalesso, R. C. (2010).Status of the population structure of the mangrove crab Ucides cordatus (Decapoda: Ocypodidae) on the Piraquê-açu river estuary, Espírito Santo, Brazil. Brazilian Journal of Oceanography, 58: 81-82. [ Links ]
Corgos, A., & Freire, J. (2006). Morphometric and gonad maturity in the spider crab Maja brachydactila: a comparison of methods for estimating size at maturity in species with determinate growth. ICES Journal of Marine Science, 63: 851-859. [ Links ]
Costa, R. S. (1972). Fisioecologia do caranguejo-uçá Ucides cordatus (Linnaeus, 1763) – Crustaceo Decápode – no nordeste brasileiro. Tese de Doutorado. Universidade de São Paulo, São Paulo, Brasil. [ Links ]
Dalabona, G., Loyola, J., & Pinheiro, M. A. A. (2005). Size at morphological maturity of Ucides cordatus (Linnaeus, 1763) (Brachyura, Ocypodidae) in the Laranjeiras Bay, Southern Brazil. Brazilian Archives of Biology and Technology, 48: 139-145. [ Links ]
Díaz, H., & Conde, J. E. (1989).Population dynamics and life history of the mangrove crab Aratus pisonii (Brachyura, Grapsidae) in a marine environment. Bulletin of Marine Science, 45: 148-163. [ Links ]
Diele, K., & Koch, V. (2010). Growth and mortality of the exploited mangrove crab Ucides cordatus (Ucididae) in N-Brazil. Journal of Experimental Marine Biology and Ecology, 395: 171-180. [ Links ]
Diele, K., Koch, V., & Saint-Paul, U. (2005). Population structure, catch composition and CPUE of artisanally harvested mangrove crab Ucides cordatus (Ocypodidae) in the Caeté estuary, North Brazil: Indications for overfishing? Aquatic Living Resources, 18: 169-178. [ Links ]
Draper, N. R., & Smith, H. (1966). Applied regressions analysis. New York: John Wiley. [ Links ]
Engelhard, G. H., & Heino, M. (2004).Maturity changes in Norwegian spring herring Clupea harengus: compensatory or evolutionary responses? Marine Ecology Progress Series, 272: 245-256. [ Links ]
Fenberg, P. B., & Roy, K. (2007). Ecological and evolutionary consequences of size-selective harvesting: how much do we know? Molecular Ecology, 17: 209-220. [ Links ]
Fernandes, M. E. B., & Carvalho, M. L. (2007). Bioecologia de Ucides cordatus Linnaeus, 1763 (Decapoda: Brachyura) na costa do estado do Amapá. Boletim do Laboratório de Hidrobiologia, 20: 15-22. [ Links ]
Fernandes-Góes, L. C., Fransozo, A., & Góes, J. M. (2005). Population dynamics of Dardanus insignis (Saussure, 1858), in the Ubatuba region, São Paulo, Brazil. Nauplius, 13: 191-196. [ Links ]
Fisher, R. A. (1930). The genetical theory of natural selection. Oxford: Oxford University Press. [ Links ]
Flores, A. A. V., & Negreiros-Fransozo, M. L. (1999). On the population biology of mottled shore crab Pachygrapsus transversus (Gibbes, 1850) (Brachyura, Grapsidae) in a subtropical area. Bulletin of Marine Science, 65: 59-73. [ Links ]
Fonteles-Filho, A. A. (1989). Recursos pesqueiros: Biologia e dinâmica populacional. Fortaleza: Imprensa Oficial do Ceará [ Links ].
Fransozo, A., Garcia, R. B., & Mantelatto, F. L. M. (2003). Morphometry and sexual maturity of the tropical hermit crab Calcinus tibicen (Crustacea, Anomura) from Brazil. Journal of Natural History, 37: 297-304. [ Links ]
Gadgil, M., & Bossert, W. H. (1970). Life historical consequences of natural selection. American Naturalist, 104: 1-24. [ Links ]
Gardner, C., & Williams, H. (1992). Maturation in the giant crab, Pseudocarcinus gigas, and the potential for sperm limitation in the Tasmanian fishery. Marine Freshwater Research, 53: 661-667. [ Links ]
Gerhart, S. D., & Bert, T. M. (2008). Life-History aspects of the stone crabs (Genus Menippe): size at maturity, growth and age. Journal of Crustacean Biology, 28: 252-261. [ Links ]
Glaser, M., & Diele, K. (2004). Asymmetric outcomes: assessing central aspects of the biological, economic and social sustainability of a mangrove crab fishery, (Ocypodidae) in North Brazil. Ucides cordatusEcological Economics, 49: 361-373. [ Links ]
Glazier, D. S. (1999). Trade-offs between reproductive and somatic (storage) investments in animals a comparative test of the Van Noordwijik and De Jonge model. Evolutionary Ecology, 13: 539-535. [ Links ]
Góes, P., Branco, J. O., Pinheiro, M. A. A., Barbieri, E., Costa, D., & Fernandes, L. L. (2010). Bioecology of the uçá-crab, Ucides cordatus (Linnaeus, 1763), in Vitória Bay, Espírito Santo State, Brazil. Brazilian Journal of Oceanography, 58: 153-163. [ Links ]
González-Gurriarán, E., & Freire, J. (1994).Sexual maturity in the velvet swimming crab Necora puber (Brachyura, Portunidae): morphometric and reproductive analyses. Journal of Marine Science, 51: 133-145. [ Links ]
Gregati, R. A., & Negreiros-Fransozo, M. L. (2009). Population biology of the burrowing crab Neohelice granulata (Crustacea: Decapoda: Varunidae) from a tropical mangrove in Brazil. Zoologia, 26: 32-37. [ Links ]
Hartnoll, R. G. (1974). Variation in growth pattern between some secondary characters in crabs (Decapoda, Brachyura). Crustaceana, 27: 131-136. [ Links ]
Hartnoll, R. G. (1978). The determination of relative growth in Crustacea. Crustaceana, 34: 281-293. [ Links ]
Hartnoll, R. G. (1982). Growth. In D. E. Bliss (Ed.). The biology of Crustacea (p. 11-196). London: Academic Press. [ Links ]
Hartnoll, R. G. (2006). Reproductive investment in Brachyura. Hydrobiologia, 557: 31-40. [ Links ]
Hartnoll, R. G., Broderick, A. C., Godley, B. J., & Saunders, K. E. (2009). Population structure of the land crab Johngarthia lagostoma on Ascension Island. Journal of Crustacean Biology, 29: 57-61. [ Links ]
Hartnoll, R. G., & Bryant, D. (1990). Size-frequency distributions in decapods crustacean – The quick, the dead, and the cast-offs. Journal of Crustacean Biology, 10: 14-19. [ Links ]
Hartnoll, R. G., & Gould, P. (1988). Brachyuran life history strategies and the optimization of egg production. Symposium of the Zoological Society of London, 59: 1-9. [ Links ]
Heino, M., & Godo, O. R. (2002). Fisheries-induced selection pressures in the context of sustainable fisheries. Bulletin Marine Science, 70: 639-656. [ Links ]
Hines, A. H. (1989). Geographic variation in size at maturity in brachyuran crabs. Bulletin of Marine Science, 45: 356-368. [ Links ]
Ibama. (1994). Lagosta, caranguejo-uçá e camarão do nordeste. Coleção Meio Ambiente, série estudos Pesca, Brasília, Brasil. [ Links ]
Ipece. (2010). Plano básico municipal – Aracati. Instituto de Pesquisa e Estratégia Econômica do Ceará, Governo do Estado do Ceará [ Links ].
Ivo, C. T. C., Dias, A. F., & Mota, R. I. (1999). Estudo sobre a biologia do caranguejo-uçá Ucides cordatus cordatus (Linnaeus, 1763), capturado no delta do rio Parnaíba, Estado do Piauí. Boletim Técnico Científico CEPENE, 7: 53-84. [ Links ]
Ivo, C. T. C., & Fonteles-Filho, A. A. (1997). Estatística pesqueira: aplicação em engenharia de pesca. Fortaleza, Brasil: Tom gráfica e editora. [ Links ]
Ivo, C. T. C., & Gesteira, T. V. C. (1999). Sinopses sobre a biologia do caranguejo-uçá Ucides cordatus cordatus (Linnaeus, 1763), capturados em estuários de sua área de ocorrência. Boletim Técnico Científico CEPENE, 7: 9-50. [ Links ]
Johnson, P. T. J. (2003). Biased sex ratios in fiddler crabs (Brachyura, Ocypodidae): a review and evaluation of the influence of sampling method, size class and sex specific mortality. Crustaceana, 76: 559-580. [ Links ]
Kassuga, A. D., Silva-Ferreira, T. C. G., Ostrovski, M. C., & Costa, T. M. M. (2008). Population structure of Munida microphthalma (Crustacea: Decapoda: Galatheidae) from the northern coast of Rio de Janeiro State, Brazil. Pan-American Journal of Aquatic Sciences, 3: 116-121. [ Links ]
Kurata, H. (1962). Studies on the age and growth of Crustacea. Bulletin of Hokkaido Reg. Fishery of Research Laboratory, 24: 1-115. [ Links ]
Kuris, A. M., Ravanan, Z., Sagi, A., & Cohen, D. (1987). Morphotypic differentiation of male Malaysian giant prawns, Macrobrachium rosenbergii. Journal of Crustacean Biology, 7: 219-237. [ Links ]
Law, R. (2000). Fishing, selection and phenotypic evolution. ICES Journal of Marine Science, 57: 659-668. [ Links ]
Law, R., & Grey, D. R. (1989). Evolution yields from populations with age-especific cropping. Evolutionary Ecology, 3: 343-359. [ Links ]
Lee, S. Y. (1995). Cheliped size and structure: the evolution of a multifunctional decapods organ. Journal of Experimental Marine Biology and Ecology, 193: 161-176. [ Links ]
Legat, J. F. A., Puchnick, A. L., Castro, P. F., Pereira, A. M. L., Góes, J. M., & Fernandes-Góes, L. C. (2005). Current fishery status of Ucides cordatus (Linnaeus, 1763) (Brachyura: Ocypodidae) in Parnaíba delta region, Brazil. Nauplius, 13: 65-70. [ Links ]
Leite, M. M. L. (2005). Relações morfométricas para compreensão de aspectos reprodutivos do caranguejo-uçá Ucides cordatus (Linnaeus 1763), no estuário do rio Coreaú-Ceará. (Dissertação de Mestrado) Universidade Federal do Ceará, Fortaleza, Ceará, Brasil. [ Links ]
Leite, M. M. L., A. A. Fonteles-Filho, J. R. F. Silva, & Cardoso, N. S. (2006). Maturidade reprodutiva funcional do caranguejo-uçá, Ucides cordatus (Crustacea, Decapoda), no estuário do rio Coreaú, Camocim, Ceará. Boletim Técnico Científico. CEPENE, 14: 41-49. [ Links ]
Leme, M. H. A. (2002). A comparative analysis of the population biology of the mangrove crab Aratus pisonii and Sesarma rectum (Brachyura, Grapsidae) from North coast of São Paulo State, Brazil. Journal of Crustacean Biology, 22: 553-557. [ Links ]
Lika, K., & Kooijman, S. A. L. M. (2003). Life history implications of allocation to growth versus reproduction in dynamic energy budgets. Bulletin of Mathematical Biology, 65: 809-834. [ Links ]
Linhares, J. C. S. (2010). Estratégias reprodutivas do caranguejo-uçá Ucides cordatus (Crustacea:Brachyura:Ucididae). (Dissertação de Mestrado) Universidade Federal do Ceará, Fortaleza, Brasil. [ Links ]
Litulo, C. 2005. Population structure and reproductive biology of the fiddler crab Uca inversa (Hoffman, 1847) (Brachyura: Ocypodidae). Acta Oecologica, 27: 135-141. [ Links ]
Lizzárraga-Cubedo, H. A., I. Tuck, N. Bayley, G. J. Pierce, & Kinnear, J. A. M. (2003). Comparisons of size at maturity and fecundity of two Scottish populations of the European lobster, Homarus gammarus. Fisheries Research, 65: 137-152. [ Links ]
Mantelatto, F. L. M., & Martinelli, J. M. (2001).Relative growth and sexual dimorphismof the South Atlantic hermit crab Loxopagurus loxochelis (Anomura, Diogenidae) from Ubatuba, Brazil. Journal of Natural History, 35: 429-437. [ Links ]
Marins, R. V., Lacerda, L. D., Abreu, E. V. I., & Dias, F. J. S. (2003). Efeitos da açudagem no rio Jaguaribe. Ciência Hoje, 33: 66-70. [ Links ]
Marques, D. F. (2008). Avaliação do estoque do caranguejo-uçá Ucides cordatus (Linnaeus, 1763) na zona estuarina do rio Pacoti, estado do Ceará. (Dissertação de Mestrado) Universidade Federal do Ceará, Fortaleza, Brasil. [ Links ]
Mantelatto, F. L. M., & Martinelli, J. M. (2001). Relative growth and sexual dimorphismof the South Atlantic hermit crab Loxopagurus loxochelis (Anomura, Diogenidae) from Ubatuba, Brazil. Journal of Natural History, 35: 429-437. [ Links ]
McQuaid, N., Briggs, R. P., & Roberts, D. (2006).Estimation of the size of onset of sexual maturity in Nephrops norvegicus (L.). Fisheries Research, 81: 26-36. [ Links ]
Melo, G. A. (1996). Manual de identificação dos Brachyura (Caranguejos e Siris) do Litoral Brasileiro. São Paulo: Editora Plêiade, FAPESP (Fundação de Amparo à Pesquisa do Estado de São Paulo). [ Links ]
Mendonça, J. T., & Pereira, A. L. C. (2009). Avaliação das capturas de caranguejo-uçá Ucides cordatus no município de Iguape, litoral sul de São Paulo, Brasil. Boletim do Instituto de Pesca, 35: 169-179. [ Links ]
MMA - Ministério do Meio Ambiente. 2004. Instrução normativa n. 5, de 21 de maio de 2004 – Anexo II: Lista Nacional das espécies de invertebrados aquáticos e peixes sobreexplotadas ou ameaçadas de sobreexplotação. [ Links ]
Mota-Alves, M. I. (1975). Sobre a reprodução do caranguejo-uçá Ucides cordatus, em mangues do Estado do Ceará. Arquivos de Ciências do Mar, 15: 85-91. [ Links ]
Nascimento, S. A. (1993). Biologia do caranguejo-uçá (Ucides cordatus). Aracajú: Administração Estadual do Meio Ambiente (ADEMA). [ Links ]
Nordhaus, I., & Wolff, M. (2007). Feeding ecology of the mangrove crab Ucides cordatus (Ocypodidae): food choice, food quality and assimilation efficiency. Marine Biology, 151: 1665-1681. [ Links ]
Olsen, E. M., Lilly, G. R., Heino, M., Morgan, M. J., Brattey, J., & Dickmann, U. (2005). Assessing changes in age and size at maturation in collapsing populations of Atlantic cod (Gadus morhua). Canadian Journal of Fisheries and Aquatic Sciences, 62: 811-823. [ Links ]
Paiva, M. P. 1997. Recursos pesqueiros estuarinos e marinhos do Brasil. Fortaleza: Edições UFC. [ Links ]
Passos, C. A., & Beneditto, A. P. M. (2005). Captura comercial do caranguejo-uçá, Ucides cordatus (L., 1763) no manguezal de Gargaú - R. J. Biotemas, 18: 223 -231. [ Links ]
Pezzuto, R. (1993). Regrans: A “basic” program for an extensive analysis of relative growth. Atlântica, 15: 93-105. [ Links ]
Pinheiro, M. A. A., & Fiscarelli, A. G. (2001). Manual de apoio à fiscalização do caranguejo-uçá (Ucides cordatus). Itajaí: Unesp/Cepsul/Ibama. [ Links ]
Pinheiro, M. A. A., & Fransozo, A. (1998). Sexual maturity of speckeled swimming crab Arenaeus cribarius (Lamarck, 1818) (Decapoda: Brachyura: Portunidae), in Ubatuba littoral, São Paulo State, Brazil. Crustaceana, 71: 434-452. [ Links ]
Pinheiro, M. A. A., & Hattori, G. Y. (2006). Relative growth of the mangrove crab Ucides cordatus (Linnaeus, 1763) (Crustacea, Brachyura, Ocypodidae) at Iguape, São Paulo. Brazilian Archives of Biology and Technology, 49: 813-823. [ Links ]
Rostant, L. V., Alkins-Koo, M., & Maitland, D. P. (2008). Growth and maturity in the manicou crab Eudaniela garmani (Brachyura: Pseudothelphusidae) from Trinidad, West Indies. Journal of Crustacean Biology, 28: 485-493. [ Links ]
Sant´Ana, R., & Pezzuto, P. R. (2009). Sexual maturity of the deep-sea red crab Chaceon notialis Manning & Holthuis 1989 (Brachyura: Geryonidae) in southern Brazil. Latin American Journal of Aquatic Research, 37: 429-442. [ Links ]
Schories, D., Barletta-Bergan, A., Barletta, M., Krumme, U., Mehlig, U., & Rademaker, V. (2003). The keystone role of leaf-removing crabs in mangrove forests of North Brazil. Wetlands Ecology and Management, 11: 243-255. [ Links ]
Semace. 2006. Atlas dos manguezais do nordeste do Brasil: avaliação das áreas de manguezais dos estados do Piauí, Ceará, Rio Grande do Norte, Paraíba e Pernambuco, Fortaleza, Brasil. Fortaleza: Superintendência Estadual do Meio Ambiente. [ Links ]
Sforza, R., Nalesso, C. R., & Joyeux, J. C. (2010). Distribution and population structure of Callinectes danae (Decapoda: Portunidae) in a tropical Brazilian estuary. Journal of Crustacean Biology, 30: 597-606. [ Links ]
Souza-Carvalho, E. A. S., Carvalho, F. L., & Couto, E. C. G. (2011). Maturidade sexual em Callinectes ornatus Ordway, 1863 (Crustacea: Decapoda: Portunidae) no litoral de ilhéus, BA, Brasil. Papéis Avulsos de Zoologia, 51: 367-372. [ Links ]
Spivak, E. D., Gavio, M. A., & Navarro, C. E. (1991). Life history and structure of the world´s southernmost Uca population: Uca uruguayensis (Crustacea: Brachyura) in Mar Chiquita Lagoon (Argentina). Bulletin of Marine Science, 48: 679-688. [ Links ]
Stearns, S. C. (1989). Trade-offs in life-history evolution. Functional Ecology, 3: 259-268. [ Links ]
Stearns, S. C. (2000). Life history evolution: successes, limitations, and prospects. Naturwissenschaften, 87: 476-486. [ Links ]
Streissl, F., & Hödl, W. (2002). Growth, mophometrics, size at maturity, sexual dimorphism and condition index of Austropotamobius torrentium Schrank. Hydrobiologia, 477: 201-208. [ Links ]
Sturges, H. A. (1926). The choice of a class interval. Journal of American Statistical Association, 21: 65-66. [ Links ]
Thurman, C. L. (1985). Reproductive biology and population structure of the fiddler crab Uca subcylindrica (Stimpson). Biological Bulletin, 169: 215-229. [ Links ]
Trippel, E. A. (1995). Age at maturity as a stress indicator in fisheries. Bioscience, 45: 759-771. [ Links ]
Vasconcelos, E. M. S., Vasconcelos, J. S., & Ivo, C. T. C. (1999). Estudo sobre a biologia do caranguejo-uçá Ucides cordatus cordatus (Linnaeus, 1763), capturado no estuário do rio Curimatau (Canguaretama) no Estado do Rio grande do Norte. Boletim Técnico Científico CEPENE, 7: 85-116. [ Links ]
Wada, S., Oba, T., Nakata, K., & Ito, A. 2008. Temporal allocation pattern between reproduction and molt within a breeding season of the hermit crab Pagurus nigrivittatus. Crustacean Research, 37: 67-73. [ Links ]
Warner, G. F. (1967). The life history of the mangrove tree crab Aratus pisonii. Journal of Zoology, 153: 321-335. [ Links ]
Wenner, A. M. (1972). Sex ratio as a function of size in marine Crustacea. American Naturalist, 106, 321-350. [ Links ]
Wenner, A. M., Fusaro, C., & Oaten, A. (1974).Size at onset of sexual maturity and growth rate in crustacean populations. Canadian Journal of Zoology, 52, 1095-1106. [ Links ]
Zar, J. H. (2010). Biostatistical analysis. Upper Saddle River (NJ), USA: Prentice Hall. [ Links ]
*Correspondencia a:
1Marcos de Miranda Leão Leite. Programa de Pós-Graduação em Ecologia e Recursos Naturais, Universidade Federal do Ceará, Fortaleza, Brazil; 1975.mirandaleao@gmail.com
2Carla Ferreira Rezende. Departamento de Biologia, Universidade Federal do Ceará, Campus do Pici, Centro de Ciências Bloco 902, Secretaria do Programa de Pós-Graduação em Ecologia e Recursos Naturais, CEP 60.455-760, Fortaleza, Ceará, Brazil; carlarezende.ufc@gmail.com
2José Roberto Feitosa Silva. Departamento de Biologia, Universidade Federal do Ceará, Campus do Pici, Centro de Ciências Bloco 902, Secretaria do Programa de Pós-Graduação em Ecologia e Recursos Naturais, CEP 60.455-760, Fortaleza, Ceará, Brazil. robertofeitosa@ufc.br
Received 08-II-2013. Corrected 19-VI-2013. Accepted 22-VII-2013.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}