










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.61 n.4 San José Oct./Dec. 2013
The bird community in naturally regenerating Myracrodruon urundeuva (Anacardiaceae) forest in Southeastern Brazil
Comunidad de aves en un bosque de Myracrodruon urundeuva (Anacardiaceae)en regeneración natural del sureste de Brasil
Comunidad de aves en un bosque de Myracrodruon urundeuva (Anacardiaceae)en regeneración natural del sureste de Brasil
*Dirección para correspondencia:
Abstract
Conservation and restoration projects involving abandoned cattle pastures pose several challenges. Restoration ecologists recognize that natural restoration is possible, but few species can successfully colonize degraded pastures. The aim of the present study was to evaluate whether land colonized by the tree Myracrodruon urundeuva increase the amount of habitat available for bird communities in Southeastern Brazil. We used fixed point counts within a 50m radius to compare the richness and composition of the bird community inhabiting naturally regenerated M. urundeuva with that an adjacent native forest remnant and a forest restored with planted Acacia spp. We recorded a total of 68 bird species. The M. urundeuva forest was able to provide valuable habitat for bird species, including more forest species, in comparison with the Acacia planting. These results indicate that M. urundeuva is a useful species for the ecological restoration of degraded pasture land.
Key words: Acacia mangium, A. auriculiformis, passive restoration, forest dependence, habitat strata.
Resumen
La conservación y restauración de pastizales abandonados representan diversos retos. Los ecologistas reconocen que la restauración natural es posible, pero algunas especies pueden colonizar con éxito los pastizales degradados. El objetivo de este estudio fue evaluar si una zona de pastos colonizada por M. urundeuva puede aumentar la cantidad de hábitat disponible para las comunidades de aves en el sureste de Brasil. La metodología utilizada fue la de conteo de puntos en un radio de 50m para comparar la riqueza y composición de comunidades de aves en un pastizal regenerado por M. urundeuva con un remanente de bosque nativo y bosque restaurado mediante la plantación de Acacia spp. Se registró un total de 68 especies de aves. El área colonizada por M. urundeuva representa un hábitat importante para las aves, incluyendo especies forestales, en comparación con la plantación de Acacia spp. Estos resultados indican que M. urundeuva es una especie útil para la restauración ecológica de los pastizales degradados.
Palabras clave: Acacia mangium, A. auriculiformis, restauración pasiva, bosque dependencia, estratos de hábitat.
The conversion of forest to pasture for cattle grazing is by far the most common practice contributing to deforestation and degradation of Latin America (Parsons, 1976; FAO, 2005). This process is common in the Atlantic Forest of Brazil, where the conversion of the original dense forest has produced abandoned and non-productive pastures: due to deforestation, only 10% of the original forest remains in a reasonable approximation of its original state (Ribeiro et al., 2009). As a current initiative with the aim of reducing human damage, efforts are being made to restore this degraded landscape (Choi, 2004; Florentine & Westbrooke; 2004, Rodrigues et al., 2009). In general, restoration practices involve two broad strategies. First, planting trees is a common and successful practice, primarily in small degraded areas. Second, natural succession occurs on its own in areas without further disturbance (Morrison & Lindell, 2011).
Restoration ecologists recognize that passive restoration (i.e., natural regeneration: Clewell et al., 2005) is possible, especially in sites where residual trees remain and where seed banks consist primarily of native species (Aide et al., 2000, Lamb et al., 2005). In abandoned cattle pastures soil degradation and compaction tend to decrease the presence of seed banks and favor the dominance of exotic grasses. These factors impede the establishment of native species and natural regeneration and few species can successfully colonize degraded pasture (Zimmerman et al. 2000; Guariguata & Ostertag, 2001; Holl, 2002). Under these circumstances, few species can successfully colonize degraded pasture. In Vale do Rio Doce region, Minas Gerais State, Southeastern Brazil, abandoned cattle pasture has been colonized by a single species of tree: Myracrodruon urundeuva Fr. All. The M. urundeuva is a native deciduous tree, belonging to the family Anacardiaceae, with anemocoric dispersion and propagation by seeds; it is vegetatively classified as late secondary, but M. urundeuva is a pioneer species in pasture and degraded land (Bertoni & Dickfeldt, 2007). M. urundeuva occurs naturally from Mexico to Paraguay, and in Brazil it is widely distributed in the Northeast, Southeast and Midwest (Santin & Leitão-Filho, 1991; Lorenzi, 1992). The expansion and dominance of M. urundeuva in Vale do Rio Doce region (Minas Gerais State), Southeastern Brazil, on degraded pasture land was a result of land use practices that involved a change from a state marked by an extreme amount of soil degradation resulting from the use of pasture for cattle grazing and a reduction of productivity, as well as economic changes (Oliveira, 2011). However, the colonization of land by M. urundeuva may inhibit the addition of species in later successional stages (Oliveira, 2011). Baruqui et al. (1985) described that during winter, M. urundeuva leaves fall down being rapidly decomposed and not forming a layer of biomass: in such specific sites, the soil is not covered, facilitating erosion and compaction, and suppressing any vegetation growth. For this reason, management interventions may be necessary to increasing structural complexity of land colonized by M. urundeuva (Zimmerman et al., 2000).
In the present study, bird communities were used to provide basic information about the ways in which M. urundeuva regeneration can provide habitats for birds in Southeastern Brazil. Whether the land colonized by M. urundeuva can increase the amount of habitat available for wildlife is the fundamental question. The main objective of the study was to compare the richness and composition of the bird community in naturally regenerating M. urundeuva forest to that of an adjacent remnant of native forest and a forest restored with rapidly-growing exotic trees. The results of this study have useful implication for the ecological restoration of degraded pasture land.
Materials and Methods
Study sites: Our study was conducted in Instituto Terra (19°22’45” S - 41°25’45” W), Aimorés, Minas Gerais State, Southeastern Brazil. Instituto Terra is a 676ha Private Conservation Unit (Reserva Particular do Patrimônio Natural) and its cover was originally composed by semideciduous Atlantic interior forest (Oliveira-Filho & Fontes, 2000). Precipitation year average is 953mm and evaporation year average is 1 224mm (Bastos, 2010). Instituto Terra was founded in 1999 with the goal of promoting ecosystem restoration on this land. Before 1999, the area was occupied by a cattle farm dominated by plains bristlegrass (Setaria vulpiseta (Lam.) Roem. & Schult.) and African grass (Urochloa decumbens (Stapf) R.D.Webster) (which are both exotic grasses), and a small and degraded forest remnant. Since 2000, Instituto Terra has used different restoration techniques to control land degradation, exotic grass expansion and soil erosion.
The site is currently a mosaic of distinct restored secondary forests. For this study, we selected three sites in this mosaic: secondary forest formed by the natural regeneration of M. urundeuva, a natural forest remnant and a forest established by planting Acacia mangium Willd. and A. auriculiformis A. Cunn. ex Benth., hereafter referred to as of M. urundeuva forest, Natural forest and Acacia plantation, respectively. In the first forest types, M. urundeuva showed good development (up to 45 years) in a degraded and sloping terrain of 20ha and has established a dominant canopy (6-9m tall). In 2001, M. urundeuva individuals were subjected to shelterwood cutting to reduce the species dominance, while other native tree species such as Joanesia principles Vell., Handroanthus heptaphyllus (Vell.) Mattos, and Ceiba speciosa (A. St.-Hil.) Ravenna were planted at a spacing of 2m. From 2005 to 2008, M. urundeuva decreased from 58% to 34%, and the Shannon-Wiener index increased from 1.54 to 3.16 (Souza & Vieira, 2010, unpublished data). The exotic plains bristlegrass dominated the understory and was grazed and mowed extensively. Natural forest consisted of a small remnant of 25ha that was degraded by selective logging in the past but has not been exploited for 35-40 years. Despite its degradation, this site is characterized by a complex structure with canopy and understory cover and is the best reference site to represent the natural forest. The Shannon-Wiener index for NF was 3.34 (Souza & Vieira 2010, unpublished data). In the Acacia plantation, Acacia mangium and A. auriculiformis were planted at a spacing of 3m on a 40ha area of pasture land in 2002. Acacia spp. constituted 29.3% of the trees in this site. The shrub layer was scarce, and the understory was dominated by plain bristlegrass that was continuously cut (Souza & Vieira, 2010, unpublished data; Bastos, 2010). M. urundeuva forest is located adjacent to Natural forest and at least 1km apart from Acacia plantation.
Bird sampling: In each of the three study sites, the bird censuses were conducted using fixed 50m radius point-counts (Bibby et al., 1992). We located six points, each 200m apart, in each study site. The points in each study site were sampled consecutively, starting with the first point, in the early morning when the vocal activities of diurnal birds began (about 6:00am-9:00am). Six points were sampled each morning, for a total of one site per day. The selected study sites were sampled on consecutive days. At each point, we recorded all bird species that were seen and/or heard during a 15min period, with a 15min break between the points. At a single sampling point, each pair or flock was counted as one detection. Precautions were taken to avoid counting the same pair or group more than once, particularly in the case of highly mobile species. Birds flying overhead were noted separately but not included in the analyses. Poor weather conditions such as rain, strong wind and fog were avoided. The point-counts were performed from August 2010 to June 2011, with a total of five surveys at each study site.
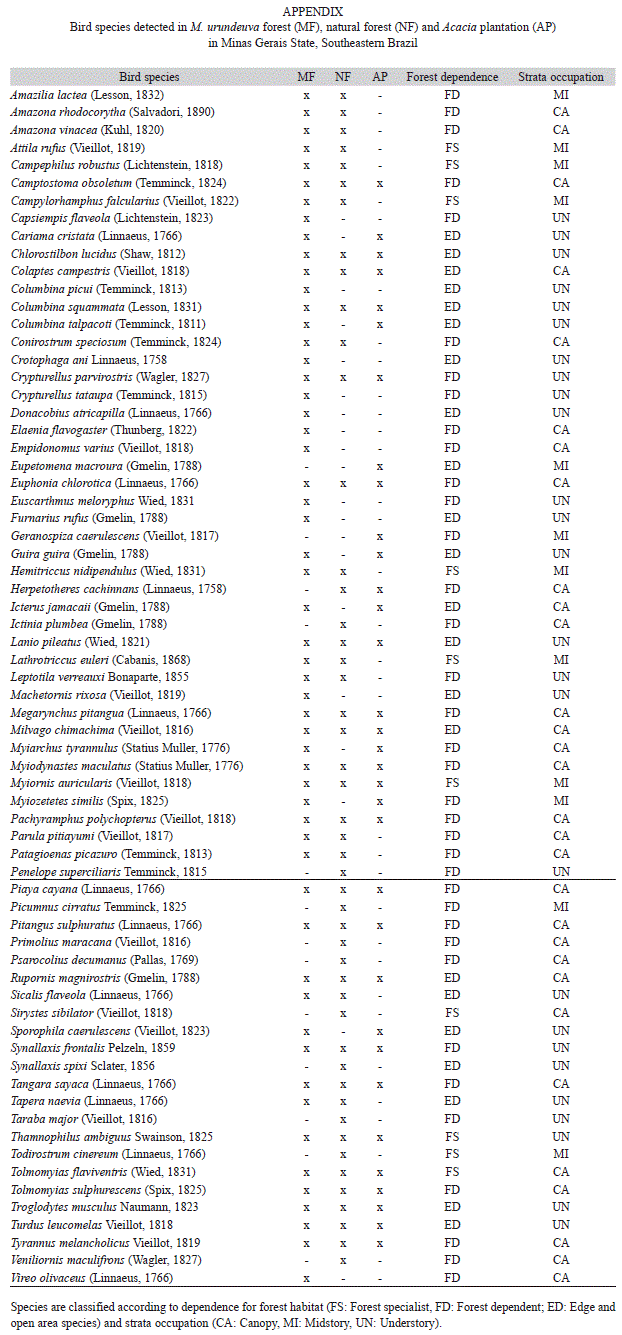
We calculated the Chao 1 index with the 95% confident intervals using EstimateS8.0 (Colwell, 2006) to compare bird richness between the three sites. A non-overlapping confidence interval implies significant differences in bird richness. We used Sorenson index to estimate similarities of bird community among selected sites types (Magurran, 2004). Species classification was conducted according to the Cômite Brasileiro de Registros Ornitológicos (CBRO, 2011). Based on field observations and the literature (Zurita et al., 2006; Volpato et al., 2010), we classified species into three groups according to their forest dependence: (1) forest specialist, found primarily in the interior of native forests subjected to little disturbance; (2) forest generalist, found in many forest types, including disturbed and secondary forests; and (3) edge and open areas species, found primarily along borders and in non-forest habitats, but rarely found in forests.
We also divided species into three groups, according to their usual occupation in the vertical strata (Parker et al., 1996): (1) canopy species, which are mostly forage and reproduce in the canopy; (2) midstory species, which are mostly found below the canopy and 2m or more above the ground; and (3) understory species, which live and breed on the ground and in vegetation up to 2m above the ground.
We evaluated differences in the mean number of bird species detected by point-count between three study sites, using repeated-measures ANOVA. In cases where the ANOVA was significant, LSD post hoc tests were used. Data were log-transformed to meet the assumptions of the ANOVA. We also evaluated differences in the number of bird species segregated by forest dependence and strata occupation. All analyses were conducted with Statistica 7.0 software (StatSoft Inc., Tulsa, OK, USA).
The composition of the bird communities was compared among bird census points at three study sites (six points at each site) with a Principal Component Analysis (PCA) in PC-ORD4 software (McCune & Mefford, 1999). To avoid distortion, 19 species were removed because they were recorded at low frequencies (one or two points). A total of 49 species were included in the analysis.
Results
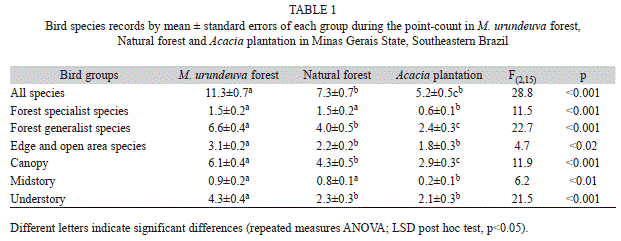
We recorded a total of 68 bird species: 55 in M. urundeuva forest, 48 in Natural forest and 33 in Acacia plantation. The Chao 1 method estimated a total of 72.5 (confidence interval 60.1-114.8) species in M. urundeuva forest, 50.8 (48.5-62.5) in natural forest and 42 (35-73.4) in Acacia plantation. Comparisons from 95% confident intervals showed greater species richness in M. urundeuva forest than in Natural forest and Acacia plantation.
The Sorenson similarity index showed that the species composition was different especially between Natural forest and Acacia plantation (0.59: 25 species common to both sites) and M. urundeuva forest and Acacia plantation (0.68: 30 species common to both sites). The similarity index was higher between M. urundeuva forest and Natural forest (72%: 37 species common to both sites. Of the 68 recorded species, 23 were found exclusively in a specific study type with 12 species in M. urundeuva forest, nine in Natural forest and two in Acacia plantation (Appendix).
The average number of bird species (per point) in M. urundeuva forests was twice the recorded number of species in Acacia plantation (Table 1). This result was similar for forest generalist and edge and open area bird species. Number of generalist and edge and open area bird species was significantly higher in M. urundeuva forest (Table 1). The number of forest specialist birds was higher in Natural forest than in Acacia plantation, but the number in natural forest did not differ significantly from that in M. urundeuva forest (Table 1).
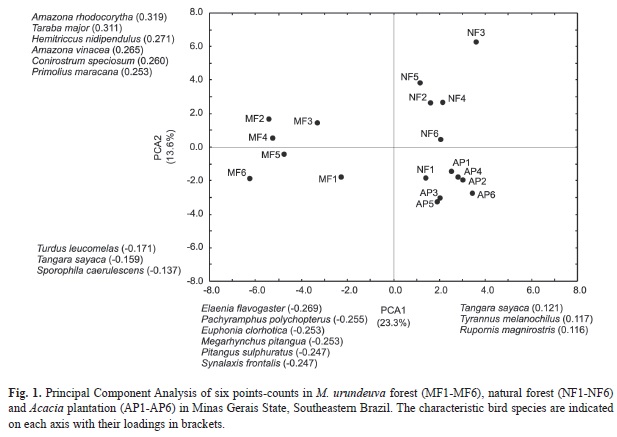
The analyses for foraging strata showed that the number of canopy and understory species was significantly higher in M. urundeuva forest than in Natural forest and Acacia plantation (Table 1). The number of midstory species was significantly lower in Acacia plantation than in M. urundeuva forest and natural forest (Table 1). The sampling site plot of the two first axes of the PCA is shown in Fig. 1. The first ordination axis (explaining 23.3% of the total variance) clearly separated M. urundeuva forest from Acacia plantation and natural forest. The species with higher negative loadings (Fig. 1, left area of the graph) were five canopy generalist forest birds (Elaenia flavogaster, Pachyramphus polychopterus, Euphonia chlorotica, Megarynchus pitangua and Pitangus sulphuratus and one bird that lives and breeds in the understory (Synallaxis frontalis). All three bird species with higher positive loadings (Fig. 1, right area of the graph) were canopy species, but two were forest generalist species (Tangara sayaca and Tyrannus melancholicus) and one was an edge species (Rupornis magnirostris). The second axis explained an additional 13.6% of the total variance and primarily separated Acacia plantation from five plots of natural forest. One point in natural forest (NF1) was located among the Acacia plantation points. The three species with higher negative loadings (Fig. 1, left bottom area of the graph) were two birds that live and breed in the understory (Turdus leucomelas and Sporophila caerulescens) and one canopy generalist forest bird (T. sayaca). Among the six species with higher positive loadings (Fig. 1, left up area of the graph) and consequently a stronger association with the natural forest plots, one specialist forest birds that live and breed in the midstory (Hemitriccus nidipendulus) and where generalist forest bird that forage and reproduce in the understory (Taraba major), and four species were canopy generalist forest birds (Amazona rhodocorytha, A. vinacea, Conirostrum speciosum and Primolius maracana).
Discussion
Our study showed that avian species richness in the M. urundeuva forest was greater than the one found in the natural forest and in the Acacia plantation. The highest species number for all ecological groups was found at this site. Furthermore, the PCA analyses showed that the M. urundeuva forest included a bird community that was distinct from those of the other study sites. The ecological restoration techniques applied in the M. urundeuva forest, including the logging of this tree species and native tree planting in the past, appeared to have produced a higher level of bird richness in this managed forest. This improvement can be attributed to the efforts to remove the dominance of M. urundeuva and to improve the structure of the vegetation (Souza & Vieira 2010, unpublished data). Most previous studies have shown that bird richness in planted or naturally regenerated forests is related to the structure and composition of the habitat. For example, in Australia, Munro et al. (2011) showed that the number of bird species increased with the amount of structurally complex vegetation in restoration planting. In contrast, a sparse understory is the principal reason for a lower level of bird richness in tree plantations in Argentina (Zurita et al., 2006) and Southeastern Brazil (Marsden et al., 2001). For this reason, the management of the M. urundeuva forest allows the coexistence of many bird species and favors an increase in bird richness, including the richness of forest species (specialist and generalist forest birds). These same characteristics can favor occupancy by the sub-canopy species (species that occupy the midstory and the understory). However, the higher richness of understory birds in the M. urundeuva forest, due primarily to the edge species, suggests that certain characteristics of the vegetation are attractive for these birds, which are often present in open and disturbed sites (species such as Guira guira, Crotophaga ani, Furnarius rufus, Machetornis rixosa and Columbina talpacoti, Appendix). Previous studies have shown that structural complexity tends to increase with age (Munro et al., 2009; Munro et al., 2011). Thus, changes in bird community composition with increasing age and structural complexity can be expected in M. urundeuva forest, resulting in a decrease in edge species and an increase in specialist and generalist birds.
Bird richness in the natural forest was lower than in the M. urundeuva forest, although specialist forest species did not show significant differences between these two sites. However, PCA analyses showed that community composition were distinct between the study sites. The second axis separated the Natural forest points from M. urundeuva forest and Acacia plantation that were characterized by a set of canopy forest species, including two bird species (A. rhodocorytha and A. vinacea) classified as “Endangered”, and one (P. maracana) classified as “Nearly Threatened”, according the International Union for the Conservation of Nature (IUCN, 2012). Furthermore, the second axis of the PCA did not isolate one point count (NF1) from the points for the Acacia plantation (AP1-AP6). This point (NF1) was the nearest to the forest edge. This location could explain the occurrence of generalist forest species (T. sayaca and T. melancholicus) and edge and open area species (R. magnirostris and T. leucomelas) in this site.
The most common strategy for the restoration of degraded areas is to plant tree seedlings (Lamb et al., 2005). In abandoned pastures, restoration by planting rapidly growing trees may be the preferred choice due to their effectiveness in the rehabilitation of the soil (D’Antonio & Meyerson, 2002), their rapid production of a canopy cover and their ability to reduce soil degradation and catalyze ecological succession (Parrotta et al., 1997; Parrotta & Knowles, 1999; Ewel & Putz, 2004; Lee et al., 2005; Schlaepfer et al., 2011). Exotic tree species such as Acacia have been planted in restoration areas throughout the tropics (Parrotta & Knowles, 1999; Passell, 2000; Gebrekirstos et al., 2006) due to their ability to survive in degraded soil and to rapidly produce a canopy cover. Despite the improvement of the canopy layer by the Acacia plantation, Bastos (2010) showed that regenerated vegetation was scarce in this plantation and that the understory was dominated by plains bristlegrass. In our study, the lower richness found for sub-canopy bird species (midstory and understory birds) appears to be a response to this vegetation structure, as previously observed by Kwok & Corlett (2000) in Lophostemon confertus (R.Br.) Peter G. Wilson & J. T. Waterh. plantation and by Zurita et al. (2006) in a Pinus plantation. In contrast, Volpato et al. (2010) observed that the dense midstory in an exotic tree plantation (Pinus spp.) favored occupancy by sub-canopy bird species.
Our results represent the first evaluation of the natural restoration by M. urundeuva in degraded pasture land. The habitat value of M. urundeuva forest could provide valuable habitat for several bird species, including forest species (specialist and generalist birds). These results indicate that M. urundeuva is certainly a useful species for weakening the barriers of exotic pastures to tree regeneration. In abandoned cattle pastures, soil degradation and compaction, a decrease in soil seed banks and exotic grass dominance usually represent barriers to the establishment of native species and natural regeneration (Guariguata & Ostertag, 2001; Holl, 2002; Zimmerman et al., 2000). Under these circumstances, few species can successfully colonize degraded pasture. Previous studies have demonstrated the ability of certain species to colonize abandoned and degraded land, and evaluating their relevance for restoration (Zimmerman et al., 2000; Aide et al., 2000; Griscom et al., 2009; Griscom & Ashton, 2011). Bertoni & Dickfeldt (2007) demonstrated that M. urundeuva tree seedlings represent an efficient approach to the recovery of degraded areas. Due to its tolerance of low soil conditions, broadcast sowing and ability to foster the recovery of land from harsh soil erosion, M. urundeuva is one attractive option for restoration application in areas that have suffered extreme soil degradation. Tree species that regenerate naturally in degraded land, such as M. urundeuva, should be managed to produce an efficient and inexpensive method of ecological restoration. In addition, considering the expansion and dominance of M. urundeuva in some degraded land in Southeastern Brazil, further investigation is required to evaluate how homogeneous M. urundeuva forests can be managed to resemble the composition of natural forest and increase the amount of available habitat for wildlife.
Acknowledgments
We thank the Instituto Terra for permissions to conduct research in the RPPN Fazenda Bulcão, Aimorés, MG, Brazil and especially for Jaeder Vieria. We also thank Jovanilson do Nascimento, Josenilto do Nascimento, Jardel Rezende, Márcia Oliveira, Sérgio Alcantara and Ruth Severino helped during the field work. Post-doc grants for the first author were provided by FAPEMIG (The Minas Gerais State Research Foundation) and a productivity grant for the second author was provided by CNPq (Brazilian Council for Development of Science and Technology, Brasilia).
References
Aide, T. M., Zimmerman, J. K., Pascarella, J. B., Rivera, L. W., & Marcano-Vega, H. (2000). Forest regeneration in a chronosequence of tropical abandoned pastures: Implications for restoration ecology. Restoration Ecology, 8, 328-338. [ Links ]
Bastos, S. C. (2010). Aplicação de indicadores de avaliação e monitoramento em um projeto de restauração florestal, Reserva Particular do Patrimônio Natural - RPPN Fazenda Bulcão, Aimorés, MG. (Unpublished master’s thesis). Universidade Federal de Viçosa, Brazil. [ Links ]
Bertoni, J. E. A., & Dickfeldt, E. P. (2007). Plantio de Myracrodruon urundeuva fr. all. (aroeira) em área alterada de floresta: desenvolvimento das mudas e restauração florestal. Revista do Instituto Florestal, 19, 31-38. [ Links ]
Baruqui, F. M., Resende, M., & Figueiredo, M. S. (1985). Causas da degradação e possibilidades de recuperação das pastagens em Minas Gerais (Zona da Mata e Rio Doce). Informe Agropecuário, 11, 27-37. [ Links ]
Bibby, C. J., Burgess, N. D., & Hill, D. A. (1992). Bird census techniques. London: Academic Press. [ Links ]
CBRO. (2011). Comitê Brasileiro de Registros Ornitológicos. Listas das aves do Brasil. 10ª Edição. Retrieved from http://www.cbro.org.br. [ Links ]
Choi, Y. D. (2004). Theories for ecological restoration in changing environment: Toward ‘futuristic’ restoration. Ecological Restoration, 19, 75-81. [ Links ]
Clewell, A., Rieger, J., & Munro, J. (2005). Guidelines for Developing and Managing Ecological Restoration Projects. Society for Ecological Restoration International - SER. Society for Ecological Restoration, Tucson, 2nd Edition. Retrieved from http://ww.ser.org. [ Links ]
Colwell, R. K. (2006). EstimateS: Statistical estimation of species richness and shared species from samples. Version 8.0.0. Retrieved from http:// purl.oclc.org/estimates. [ Links ]
D’Antonio, C., & Meyerson, L. A. (2002). Exotic Plant Species as Problems and Solutions in Ecological Restoration: A Synthesis. Restoration Ecology, 10, 703-713. [ Links ]
Ewel, J. J., & Putz, F. E. (2004). A place for alien species in ecosystem restoration. Frontiers in Ecology and the Environment, 2, 354-360. [ Links ]
FAO. (2005). Global Forest Resources Assessment 2005, Progress towards sustainable forest management. Vol. 147 of FAO Forestry Paper. Rome: Food and Agriculture Organization of the United Nations. [ Links ]
Florentine, S. K., & Westbrooke, M. E. (2004). Restoration on abandoned tropical pasturelands-do we know enough? Journal for Nature Conservation, 12, 85-94. [ Links ]
Gebrekirstos, A., Teketay, D., Fetene, M., & Mitlohner, R. (2006). Adaptation of five co-occurring tree and shrub species to water stress and its implication in restoration of degraded lands. Forest Ecology and Management, 229, 259-267. [ Links ]
Griscom, H. P., & Ashton, M. S. (2011). Restoration of dry tropical forests in Central America: A review of pattern and process. Forest Ecology and Management, 261, 1564-1579. [ Links ]
Griscom, H. P., Griscom, B. W., & Ashton, M. S. (2009). Forest Regeneration from Pasture in the Dry Tropics of Panama: Effects of Cattle, Exotic Grass, and Forested Riparia. Restoration Ecology, 17, 117-126. [ Links ]
Guariguata, M. R., & Ostertag, R. (2001). Neotropical secondary forest succession: changes in structural and functional characteristics. Forest Ecology and Management, 148, 185-206. [ Links ]
Holl, K. D. (2002). Effect of shrubs on tree seedling establishment in abandoned tropical pasture. Journal of Ecology, 90, 179-187. [ Links ]
IUCN. (2012). Red List of Threatened Species. Version 2012.1. Retrieved from http://www.iucnredlist.org. [ Links ]
Kwok, H. K., & Corlett, R. T. (2000). The bird community of a natural secondary forest and a Lophostemon confertus plantation in Hong Kong, South China. Forest Ecology and Management, 130, 227-234. [ Links ]
Lamb, D., Erskine, P. D., & Parrotta, J. A. (2005). Restoration of degraded tropical forest landscapes. Science, 310, 1628-1632. [ Links ]
Lee, E. W. S., Hau, B. C. H., & Corlett, R. T. (2005). Natural regeneration in exotic tree plantations in Hong Kong, China. Forest Ecology and Management, 212, 358-366. [ Links ]
Lorenzi, H. (1992). Árvores brasileiras: manual de identificação e cultivo de plantas arbóreas nativas do Brasil. Nova Odessa: Plantarum. [ Links ]
Magurran, A. E. (2004). Measuring biological diversity. Oxford: Blackwell Science. [ Links ]
Marsden, S. J., Whiffin, M., & Galetti, M. (2001). Bird diversity and abundance in forest fragments and Eucalyptus plantations around a Brazilian Atlantic Forest reserve. Biodiversity and Conservation, 10, 737-751. [ Links ]
McCune, B., & Mefford, M. J. (1999). PC-ORD - Multivariate analysis of ecological data, Version 4. Gleneden Beach, MjM Software Design. [ Links ]
Morrison, E. B., & Lindell, C. A. (2011). Active or Passive Forest Restoration? Assessing Restoration Alternatives with Avian Foraging Behavior. Restoration Ecology, 19, 170-177. [ Links ]
Munro, N. T., Fischer, J., Barrett, G., Wood, J., Leavesley, A., & Lindenmayer, D. B. (2011). Bird’s response to revegetation of different structure and floristics. Are restoration plantings restoring bird communities? Restoration Ecology, 19, 223-235. [ Links ]
Munro, N. T., Fischer, J., Wood, J., & Lindenmayer, D. B. (2009). Revegetation in agricultural areas: the development of structural complexity and floristic diversity. Ecological Application, 19, 1197-1210. [ Links ]
Oliveira, F. P. (2011). Monodominância de aroeira: Fitossociologia, relações pedológicas e distribuição espacial em Tumiritinga, MG. (Unpublished master’s thesis). Universidade Federal de Viçosa, Brazil. [ Links ]
Oliveira-Filho, A. T., & Fontes, M. A. L. (2000). Patterns of floristic differentiation among Atlantic Forests in Southeastern Brazil and the influence of climate. Biotropica, 32, 793-810. [ Links ]
Parker, T. A. III., Stotz, D. F., & Fitzpatrick, J. W. (1996). Ecological and Distributional Databases for Neotropical Birds. In T. A. Parker III, D. F. Stotz, J. W. Fitzpatrick, & D. K. Moskovits (Eds.), Neotropical birds: Ecology and Conservation (pp. 114-436). Chicago: University of Chicago Press. [ Links ]
Parrotta, J. A., Knowles, O. H., & Wunderle Jr., J. M. (1997). Development of floristic diversity in 10-year-old restoration Forest on a bauxite mined site in Amazonia. Forest Ecology and Management, 99, 21-42. [ Links ]
Parrotta, J. A., & Knowles, O. H. (1999). Restoration of tropical moist forests on bauxite mined lands in the Brazilian Amazon. Restoration Ecology, 7, 103-116. [ Links ]
Parsons, J. J. (1976). Forest to pasture: development or destruction? Revista de Biología Tropical, 24, 121-138. [ Links ]
Passell, H. D. (2000). Recovery of Bird Species in Minimally Restored Indonesian Tin Strip Mines. Restoration Ecology, 8, 112-118. [ Links ]
Ribeiro, M. C., Metzger, J. P., Martensen, A. C., Ponzoni, F. J., & Hirota, M. M. (2009). The Brazilian Atlantic Forest: How much is left, and how is the remaining forest distributed? Implications for conservation. Biological Conservation, 142, 1141-1153. [ Links ]
Rodrigues, R. R., Lima, R. A. F., Gandolfi, S., & Nave, A. G. (2009). On the restoration of high diversity forests: 30 years of experience in the Brazilian Atlantic Forest. Biological Conservation, 142, 1242-1251. [ Links ]
Santin, D. A., & Leitão-Filho, H. F. (1991). Restabelecimento e revisão taxonômica do gênero Myracrodruon Freire Allemão (Anacardiaceae). Revista Brasileira de Botânica, 14, 133-145. [ Links ]
Schlaepfer, M. A., Sax, D. F., & Olden, J. D. (2011). The potential conservation Value of Non - Native Species. Conservation Biology, 25, 428-437. [ Links ]
Volpato, G. H., Prado, V. M., & Anjos, L. (2010). What can tree plantations do for forest birds in fragmented forest landscapes? A case study in southern Brazil. Forest Ecology and Management, 260, 1156-1163. [ Links ]
Zimmerman, J. K., Pascarella, J. B., & Aide, T. M. (2000). Barriers to forest regeneration in an abandoned pasture in Puerto Rico. Restoration Ecology, 8, 350-360. [ Links ]
Zurita, G. A., Rey, N., Varela, D. M., Villagra, M., & Bellocq, M. I. (2006). Conversion of the Atlantic Forest into native and exotic tree plantations: Effects on bird communities from the local and regional perspectives. Forest Ecology and Management, 235, 164-173. [ Links ]
*Correspondencia a:
1Graziele Hernandes Volpato. Programa de Pós-Graduação em Ciência Florestal, Laboratório de Restauração Florestal, Departamento de Engenharia Florestal, Universidade Federal de Viçosa, Viçosa, MG; gravolpato@yahoo.com.br Corresponding author
1Sebastião Venâncio Martins. Programa de Pós-Graduação em Ciência Florestal, Laboratório de Restauração Florestal, Departamento de Engenharia Florestal, Universidade Federal de Viçosa, Viçosa, MG; venancio@ufv.br
Received 10-I-2013. Corrected 13-V-2013. Accepted 13-VI-2013


{kind=link}
{kind=link}
{kind=link}
{kind=link}