










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.61 n.3 San José Sep. 2013
Photosynthesis of seedlings of Otoba novogranatensis (Myristicaceae) and Ruagea glabra (Meliaceae) in abandoned pasture, secondary forest and plantation habitats in Costa Rica
Fotosíntesis de plántulas de Otoba novogranatensis (Myristicaceae) y Ruagea glabra (Meliaceae) en pastizales abandonados, bosques secundarios y hábitats de plantaciones en Costa Rica
Fotosíntesis de plántulas de Otoba novogranatensis (Myristicaceae) y Ruagea glabra (Meliaceae) en pastizales abandonados, bosques secundarios y hábitats de plantaciones en Costa Rica
*Dirección para correspondencia:
Abstract
Enrichment planting in naturally recovering secondary forests or in tree plantations is increasingly being used as strategy to restore later-successional, large-seeded tropical forest trees. We seeded two tree species (Otoba novogranatensis and Ruagea glabra) in three agricultural sites in Southern Costa Rica: abandoned pastures, eight to ten year old secondary forests and three year old tree plantations (containing two N-fixing of four total tree species). We measured micrometeorological conditions, soil water content, plant water potential, leaf area, foliar C and N, and photosynthesis to better understand mechanistic responses of seedlings to conditions in the different successional habitats. Micrometeorological conditions, soil water content, and plant water potential were generally similar across habitats. Certain aspects of leaves (such as Specific Leaf Area and foliar N content), and photosynthesis (e.g. quantum yield and electron transport rate) were highest in the plantations, intermediate in the secondary forests, and lowest in abandoned pastures. Enhanced rates of photosynthetic biochemistry (such as Vcmax and Jmax) and Photosystem II efficiency (e.g. thermal energy dissipation) occurred in leaves from the plantations compared to the abandoned pastures, which may be related to higher leaf %N content. Results suggest that foliar N may be of greater importance than soil water content and micrometeorological factors in driving differences in photosynthetic processes across planting habitats. Planting seeds of these two species in plantations containing three year old trees (including two N-fixing species) enhances certain aspects of their photosynthesis and growth, compared to seedlings in abandoned pastures with non-native grasses, and thus can help increase forest recovery on abandoned agricultural lands.
Key words: chlorophyll a fluorescence, forest restoration, gas exchange, leaf N content, Photosystem II, tropical secondary forest.
Resumen
El enriquecimiento de bosques secundarios o plantaciones forestales en proceso de regeneración natural por medio de la siembra de plántulas es una práctica cada vez más utilizada para restaurar bosques tropicales en estado de sucesión tardía. Sembramos dos especies de árboles (Otoba novogranatensis y Ruagea glabra) en pastizales abandonados, bosques secundarios de ocho a diez años de edad y plantaciones forestales de tres años de edad (con dos especies fijadoras de Nitrógeno de un total de cuatro especies) en tres sitios agrícolas en el Sur de Costa Rica. Medimos condiciones micrometeorológicas, contenido de agua del suelo, potencial hídrico de las plantas, área foliar, C y N foliar, y fotosíntesis para entender de una mejor manera las respuestas funcionales de las plántulas ante condiciones de distintos estadíos sucesionales. Las condiciones micrometeorológicas, contenido hídrico del suelo y el potencial hídrico de las plantas fueron mayoritariamente similares entre hábitats. Algunos aspectos de las hojas (como Área Foliar Específica y contenido de N foliar) y fotosíntesis (ej.: rendimiento cuántico y tasa de transporte de electrones) presentaron valores mayores en las plantaciones, intermedios en los bosques secundarios y menores en los pastizales abandonados. Se obtuvo un aumento en las tasas fotosintéticas bioquímicas (como Vcmax, Jmax) y la eficiencia del Fotosistema II (ej.: disipación de energía térmica) en hojas provenientes de las plantaciones comparado a las de los pastizales, posiblemente relacionado a un mayor %N foliar. Los resultados sugieren que el N foliar puede ser más importante que el contenido de agua del suelo y que los factores micrometeorológicos para marcar diferencias en los procesos fotosintéticos entre hábitats. Las plántulas de estas dos especies en las plantaciones con árboles de tres años de edad (incluyendo dos fijadoras de N) incrementaron ciertos aspectos de su fotosíntesis y crecimiento comparado a las plántulas en los pastizales abandonados de especies exóticas, por lo tanto, esta práctica puede ayudar a incrementar la recuperación de los bosques en áreas agrícolas abandonadas.
Palabras clave: fluorescencia de clorofila a, restauración de bosques, intercambio gaseoso, contenido de N foliar, fotosistema II, bosque tropical secundario.
Efforts to restore tropical forests have increased substantially over the past decade. In some cases tropical forests regenerate quickly when agriculture is ceased, whereas in heavily-used sites located far from seed sources, recovery is often sufficiently slow that intervention is necessary to accelerate forest recovery (Lamb et al. 2005, Chazdon 2008). The most common strategy for restoring tropical forest is to plant a mix of relatively fast-growing tree species that establish a canopy, ameliorate stressful microclimate conditions, improve nutrient availability, shade out light demanding grasses, and attract seed dispersers (Holl 2002, Lamb et al. 2005, Chazdon 2008). The assumption is that these species will facilitate the subsequent establishment of a wide suite of forest plants. An increasing number of studies, however, have shown that colonization of larger-seeded and later-successional tree species can be extremely slow, even in cases where there is remnant forest nearby (Martínez-Garza & Howe 2003, Zimmerman et al. 2007, Cole et al. 2010).
One strategy used to increase forest diversity is to “enrich” these sites by seeding or planting later successional species once an initial tree canopy has been established (Lamb et al. 2005, Piotto 2007). Several studies have shown that medium- to large-seeded mature forest trees can be seeded successfully into either secondary forests or tree plantations at a relatively low cost (Engel & Parrotta 2001, Camargo et al. 2002, Bonilla-Moheno & Holl 2010, Cole et al. 2011). However, there has been relatively little consideration of how the composition of the initially planted species affects the growth of species planted in later years.
Our past research shows that seedling growth, relative allocation to above-ground biomass, and foliar N concentrations were higher in seedlings growing in the understory of mixed species plantations with two N-fixing species, compared to secondary forests or abandoned pastures in Southern Costa Rica (Cole et al. 2011). For example, seedling survival was 20% higher, seedling height 33% higher and seedling biomass 2-fold greater in plantations compared to secondary forests and abandoned pastures. Foliar N content was almost twice as high in the plantations as in secondary forest and abandoned pasture habitats. Moreover, litterfall N concentration was higher in plantations than secondary forests, whereas concentrations of several other nutrients (e.g. Ca, Mg, K, Zn and Mn) were higher in secondary forest litter (Celentano et al. 2011). Foliar P concentration was somewhat higher in the plantations and did not differ between abandoned pastures and secondary forests (Cole et al. 2011); surprisingly, litter-fall P and C:P did not differ across the three habitats (Celentano et al. 2011). These results, as well as other studies (Cusack & Montagnini 2004, Siddique et al. 2008), suggest that the initial species can strongly influence nutrient dynamics and growth, and in turn, the successional trajectory in a site.
Most past studies of tree planting to restore tropical forest have primarily measured seed-ling survival and growth, despite the fact that well-replicated, manipulative restoration treatments provide an excellent opportunity to better understand the physiological mechanisms explaining successional patterns of forest seedling dynamics and further our knowledge of tropical seedling physiology. Moreover, knowledge of the particular abiotic factors (e.g. water, light, or nutrient availability) that most limit growth can help inform tropical restoration plans (Loik & Holl 1999), and how to best allocate the limited resources available to restore the extensive areas of degraded tropical lands (Holl & Aide 2011). The physical environment may limit survival and growth of seedlings used in restoration efforts by reducing intrinsic photosynthetic capacity through stomatal limitations to carbon uptake, by reducing the efficiency of light capture and electron transport within Photosystem II (PSII), or by reducing the rate of biochemical processes associated with carbon fixation, utilization of triose phosphates, phloem loading, and sink strength (Pammenter et al. 1993, Raines 2003, Amiard et al. 2005, Minchin & Lacointe 2005, Demmig-Adams & Adams 2006, Anderson et al. 2008).
The goal of this study was to better understand the physiological mechanisms underlying differences in growth for seedlings direct-seeded in abandoned pastures, secondary forests, and plantations that contain two N-fixing trees to restore premontane tropical forest in Southern Costa Rica. In this study, we measured a variety of structural (e.g. leaf area, %C, %N, δ13C, δ15N) and physiological (e.g. water relations and photosynthesis) characteristics for two common tree species within the three habitats at three sites. We tested whether PSII efficiency, photosynthetic capacity, and biochemistry differed within the tree plantations, abandoned pasture, and secondary forest habitats.
Materials and Methods
Study area: The study was carried out in three habitats: an abandoned pasture, a secondary forest, and a plantation (see descriptions below), at three sites locally named Bambu (BB, 8°44’36” N 82°58’04” W), Finca Loma Linda (LL, 8°44’21”N 82°55’44” W) and San Gabriel (SG, 8°45’28” N 82°57’28” W), separated by >1km, and located near the town of Agua Buena, in Southern Costa Rica. Study sites ranged from 1 110 to 1 290m above sea level, with slopes of 1535º and are located in tropical premontane rain forest (Holdridge et al. 1971). All pasture and plantation plots were used for a mixture of coffee and cattle pasture, the typical landuse in this region, for at least 30 years prior to cessation of active management in 2004 (Holl et al. 2011 Table 1 for a detailed land use history of these sites). The soils are a mix of ultisols and andisols; they are moderately acidic with high percent organic matter and relatively low P. The only soil nutrient that varied among habitat types was Ca, which was higher in plantations and secondary forests than in pastures (Cole et al. 2011).
Species description: We direct-seeded five primary forest tree species that have large seeds, as part of a study on enrichment planting (Cole et al. 2011). We selected two species, Otoba novogranatensis Moldenke (Magnoliales, Myristicaceae) and Ruagea glabra Triana & Planch (Sapindales, Meliaceae), that had sufficient germination and leaf area to conduct physiological measurements. Both species are locally-common, canopy trees with large, animal-dispersed, recalcitrant seeds (mean fresh seed weight - O. novogranatensis: 4.6±0.8g; R. glabra: 2.7±0.5g).
Experimental design: We direct-seeded into each of three habitat types: experimentally established tree plantations (three years since tree planting); abandoned pastures (one year since grazing or clearing, hereafter referred to as ‘pasture’), and secondary forests (eight to ten years since agriculture or grazing). Plantation and pasture plots (50×50m) were established in 2004 as part of a large-scale restoration research project to test the effects of different tree planting strategies on forest recovery (described in detail in Holl et al. 2011). We selected secondary forests that had eight to ten years of regrowth and similar previous land use to the other treatments. The study was set up as a randomized block design with site (n=3) as the blocking factor and each of the three habitats at a site located within 10-200m of each other.
The tree plantations included a mixture of four tree species: two native species with com-mercially valuable timber, Terminalia amazonia (J.F. Gmel.) Exell (Combretaceae) and Vochysia guatemalensis Donn. Sm. (Vochysia-ceae), and two fast-growing, naturalized species, Erythrina poeppigiana (Walp.) O.F. Cook and Inga edulis Mart. (both are N-fixers in the Fabaceae), that are commonly used as shade trees in agriculture in the region. Trees were planted at a density of 1 252/ha (separated by 2.8m). At the time of direct seeding, plantations had developed closed canopies that were four to five meters tall; the canopy cover and litter-fall of the sites was almost entirely comprised of the planted species (Celentano et al. 2011), given that the few seedlings that had naturally recruited were quite small (<2m). Pasture plots had almost no tree canopy cover and were dominated by ruderal vegetation (1-1.5m tall) comprised of introduced forage grasses (primarily Axonopus scoparius (Flüggé) Kuhlm., Pennisetum purpureu Schumach. and Urochloa brizantha (Hochst. Ex. A. Rich.) R.D. Webster); a variety of ruderal herbs (primar-ily in the Asteraceae) and the fern Pteridium arachnoideum (Kaulf.).
We marked a 50×50m plot in the center of each secondary forest (total forest size ~1-3ha). Canopy height ranged from 5-15m. The mean stem density (stems >2cm dbh/ha) for the three sites for this study were 3 698 (Bambu), 2 240 (Finca Loma Linda), and 2 708 at San Gabriel. Dominant tree species included Cecropia obtusifolia Bertol., Conostegia xalapensis (Bonpl.) D. Don ex DC., Croton draco Schltdl., Heliocarpus appendiculatus Turcz., and Psidium guajava L (Cole, unpublished data). We did not find any large N-fixing trees in secondary forest plots.
Within each 50×50m plot in each habitat type (abandoned pasture, plantation, and secondary forest), we established three 5×8m subplots separated by over 10m (so that seeding manipulations would not confound measurements of natural recruitment in other areas of the plot); data from the three subplots were combined. Within each subplot seeds were planted in single-species rows at a density of eight seeds per square m and a depth of ~3cm. Grasses and herbaceous vegetation in the pastures were cut immediately prior to seeding to facilitate experimental set up, but quickly grew back.
Freshly fallen seeds were collected during the peak of the fruiting season in March (R. glabra) and April (O. novogranatensis) 2007 from a minimum of four trees located in forest fragments within a ten km radius of planting sites. Seeds were mixed and stored for less than three days before being soaked for 12-24h to imbibe water. Seeds that showed signs of damage or floated in water were discarded (Schatral & Fox 1994). At the time that the current study was conducted (February-March 2008), seedlings were nine to eleven months old and ~15-25cm in height.
Seedling measurements (described below) were generally taken on five individuals in each habitat × site combination, although there were no individuals of R. glabra surviving in the pasture treatment at one site, so n=10-15 individuals of each species in each treatment for most measurements. Because many of the surviving seedlings in the pasture habitat were apparently in negative carbon balance at the time of the A-Q curves (described below), we have data for fewer individuals (n=4 or 7) in the pastures.
Existing vegetation and meteorological conditions: We measured existing overstory and understory vegetation cover at four points per subplot (12 points total) in March 2008. We estimated percent grass cover, forb cover, and bare ground in 1×1m quadrats using a ranking system: 0, 1-5, 5-10, 10-25, 25-50, 50-75, 75-95, and 95-100%. We used a spherical densiometer to measure canopy cover in four directions at one meter height, and averaged the values.
Rainfall and temperature data during the study period were obtained from the Organization for Tropical Studies Las Cruces Biological Station (~8km from the study sites). Micrometeorological conditions within the three habitats were monitored during the period of plant physiological measurements using LI-COR Model 1 000 data loggers. A LI-COR 1 000-102 soil thermistor was inserted at an angle to the surface to a depth of 1cm. A Vaisala INTERCAMP HMP integrated air temperature and relative humidity sensor was suspended within a 4cm diameter piece of pvc capped at one end, which served as a radiation shield. The radiation shield was mounted to rebar with the open end facing the soil surface so that the temperature/RH sensor was at seedling leaf height. A LI-COR LI-190 quantum sensor was mounted on top of the radiation shield to measure photosynthetic Photon Flux Density (PFD; 400-700nm) at the height of the top of seedlings. Conditions were measured using three logger-sensor assemblies simultaneously at one randomly-selected location within each habitat type per site for three consecutive days. Each three day period was uniform in terms of morning temperature, the timing of the onset of cloudiness, and afternoon rainfall. Although February is part of the dry season at this latitude, 2008 had a wetter-than-usual dry season.
Soil and plant water relations: Soil volumetric water content was measured on the days of gas exchange measurements using a HydroSense portable TDR with 20cm long probes (Campbell Scientific, Logan, UT, USA). Probes were inserted into the soil in a grid of nine locations in each plot.
Leaf water potential (?) was measured using a Scholander-type pressure chamber (PMS Instruments, Corvallis, OR, USA) during the mid-morning gas exchange measurement period on five leaves per species × habitat × site. Measurements were made in situ for leaf blades combined with ca. 5cm long petiole segments that were removed with a scissors, and then immediately inserted into the pressure chamber.
Photosynthetic gas exchange and chlorophyll a fluorescence from Photosystem II: Photosynthetic CO2 assimilation (A) was measured from approximately 0700 to 1200 hours local time (before onset of cloudiness and daily rainfall) using a portable open-mode infrared gas analyzer (Model 6400, LI-COR, Inc., Lincoln, NE, USA) with an integrated Leaf Chamber Fluorometer (LCF). Air temperature and vapor pressure deficit (VPD) within the leaf chamber were maintained at ambient levels. Leaf temperatures were recorded with a chromel-constantan thermocouple appressed to the abaxial surface of the leaf within the leaf chamber. The CO2 concentration within the leaf measurement chamber was maintained by scrubbing the incoming airstream with soda lime, and the subsequent addition of 385μmol/ mol of CO2 via injection from an external cartridge. PFD within the chamber was maintained using the LCF Light Emitting Diodes (LEDs). For all measurements, the 2cm2 leaf chamber of the LCF was used. Distal, fully mature leaves of five randomly-selected plants of each species in each plot were inserted into the leaf chamber at their natural branch orientation by mounting the leaf chamber on a tripod. Photosynthetic measurements were recorded when all stability criteria were met when the coefficient of variation for A and stomatal conductance to water vapor (gs) combined was <0.5%.
In order to characterize potential stress within PSII and carbon gain in response to variation in PFD, we simultaneously measured photosynthetic gas exchange and chlorophyll a fluorescence light response curves across planting habitats. Upon enclosure within the chamber, each leaf was allowed to dark-adapt for 20min, followed by a measurement of the maximal quantum yield of chlorophyll a fluorescence from PSII in the dark-adapted state (FV/FM; see next paragraph for fluorescence measurement settings). A software script that combined gas exchange and chlorophyll a fluorescence measurements was utilized to generate the A-Q (CO2 assimilation vs. PFD) response and the responses of the quantum yield of PSII (ΦPSII vs. PFD), the proportion of open PSII reaction centers (qP vs. PFD; nomenclature of Vankooten & Snel 1990), electron transport rate through PSII (ETR vs. PFD), and the engagement of energy dissipation processes within PSII (D vs. PFD). Following determination of FV/FM, the rate of CO2 assimilation was recorded and the software script altered the fluorescence measurement and flash conditions (described below). PFD was then increased stepwise to 1 200μmol/m2.s, with measurements at 1 200, 700, 500, 300, 100, 60 and 0μmol/m2.s. PFD was measured with a gallium-arsenide-phosphide sensor within the leaf chamber. Preliminary experiments showed that stomata responded quickly (<5min) to increases in PFD. At a particular PFD level, measurements were recorded at 4s intervals, and PFD levels were changed to the next quantity when CO2 uptake stabilized (three stability criteria met for CO2 and H2O concentrations and flow rate, with a total CV of 0.1%).
For the determination of FV/FM, the measurement software was set for the LCF to operate at an intensity of one, a modulation of 0.25kHz, a filter setting of one, and a gain of ten; the saturating flash duration was 0.8s at an intensity of seven, with a modulation of 20kHz with a filter of 50kHz. For the light-adapted determination of ΦPSII, the measurement intensity was increased to five and the modulation increased to 20kHz, and the flash duration was 0.8s. For fluorescence parameters that required measurement of Fo’, PSII was oxidized by preferentially exciting Photosystem I (PSI); the actinic light source was turned off for six seconds, and the far red source intensity was set to eight. The far red source was turned on one second before the actinic turned off, and then the far red turned off one second after the actinic was turned off. The modulation was 0.25kHz, and the filter was one Hz.
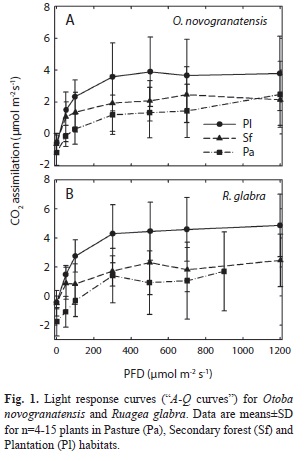
Based on the A-Q curve data, dark respiration (Rd), the apparent quantum yield (Φ), light compensation point (LCP), PFD at incipient saturation of CO2 assimilation (PFDsat), and maximal photosynthetic rate (Amax) were estimated from each light response curve using a non-rectangular hyperbola model (Sims & Pearcy 1991). These measurements may overestimate respiration due to the gasket effect on CO2 diffusion while measuring low rates of gas exchange (Pons & Welschen 2002). We were not able to utilize bottled oxygen to minimize the Kok Effect on dark respiration.
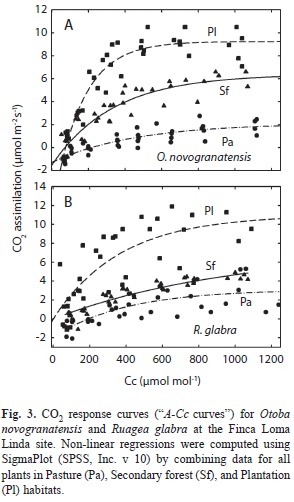
At one site (LL), we compared photosynthetic capacity at high PFD levels by measuring rates of A in relation to varying internal leaf CO2 concentration (cc), or instantaneous A-Ci curves. During all measurements, PFD was held at the median daily maximum value of 1 500mmol/m2.s using the red-blue LEDs, ambient temperatures were ~18-25°C, and VPD was generally <2.50kPa. Photosynthesis was measured and Cc was calculated at each of the following leaf chamber pCO2 target values: 100, 200, 300, 400, 600, 800, 1 000 and 1 200μmol/mol. Photosynthetic capacity under saturating light and optimal ambient conditions was calculated using a non-linear regression between A and leaf chamber pCO2. The maximum rates of carboxylation (Vcmax) and electron transport (Jmax), and Triose Phosphate Utilization for carbohydrate transport across the chloroplast membrane (TPU) were calculated from the A-Ci curves, and were corrected to a common temperature of 25°C (Long & Bernacchi 2003).
Leaf structure and composition: Leaf area (blade only) was measured using a LI-COR Model LI-3100 leaf area meter. The leaves that were used for water potential measurements were returned to the laboratory, dried in a forced draft oven at 40ºC for three days, and weighed. We compared the intrinsic water use efficiency (δ13C) and potential source of N from biological fixation (δ15N) for these leaves. Following drying, leaves were ground in a ball mill and packed in tin capsules. Percent C, %N, δ13C, and δ15N were determined on a continuous-flow elemental analyzer (Carlo Erba Model 1108, CE Elantech, Inc., Lakewood, NJ, USA) coupled to an isotope ratio mass spectrometer (ThermoFinnigan Delta Plus XP, Thermo Fisher Scientific, Waltham, MA) at the UC Santa Cruz Stable Isotope Laboratory.
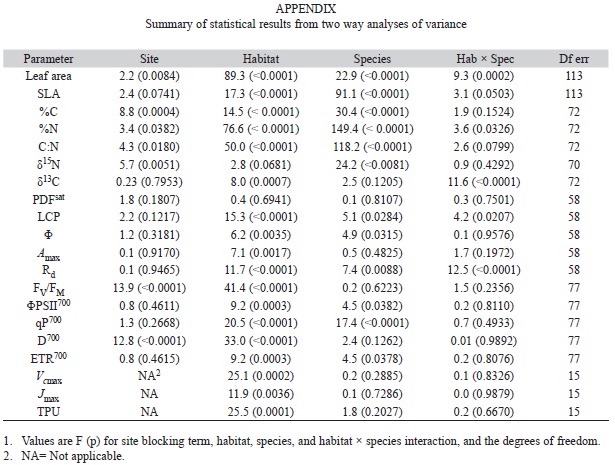
We analyzed the data using a randomized-block, two-way analysis of variance (ANOVA) to compare the effects of habitat and species and their interactions on most plant response variables (e.g. fluorescence, gas exchange, isotopes, water potential) with site as the blocking factor. A-Ci curves were only conducted at a single site so they were analyzed using a two-way analysis of variance without a blocking factor.
The model residuals for most variables met assumptions of normality, but in a few cases variables were arcsine square root transformed (percentages) or log + 1 transformed to normalize data. We used Tukey’s HSD test (p=0.05) for post hoc analyses for pair-wise habitat comparisons. We report means ± 1 SE throughout. All analyses were conducted using SAS version 9.2.
Results
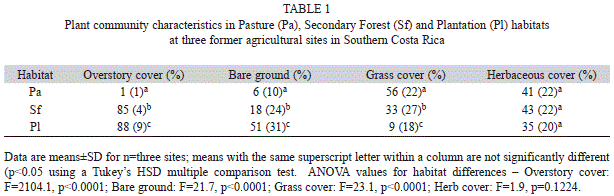
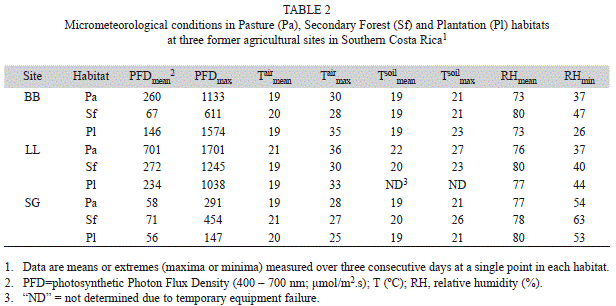
Overstory tree cover was >80% in plantations and secondary forests, and non-existent in pastures (Table 1). Grass cover was highest in pastures, intermediate in secondary forests, and lowest in plantations with the reverse pattern for bare ground; herbaceous cover was similar across habitats (Table 1). Both maximum and mean PFD at seedling height (~0.20m) were highly variable across sites and habitats (Table 2), reflecting variation in understory and overstory structure across sites. Mean air and soil temperatures and RH were relatively similar and did not show a consistent trend across habitat types (Table 2). Total precipitation for the 12 month period from seed planting to seedling physiology measurements was 4 684mm, which is 142% of the 3 294±712mm annual average for the prior 34 years. For 2008, precipitation for January through March (the dry season in this region) was 190mm (inter-annual range 167-263mm).
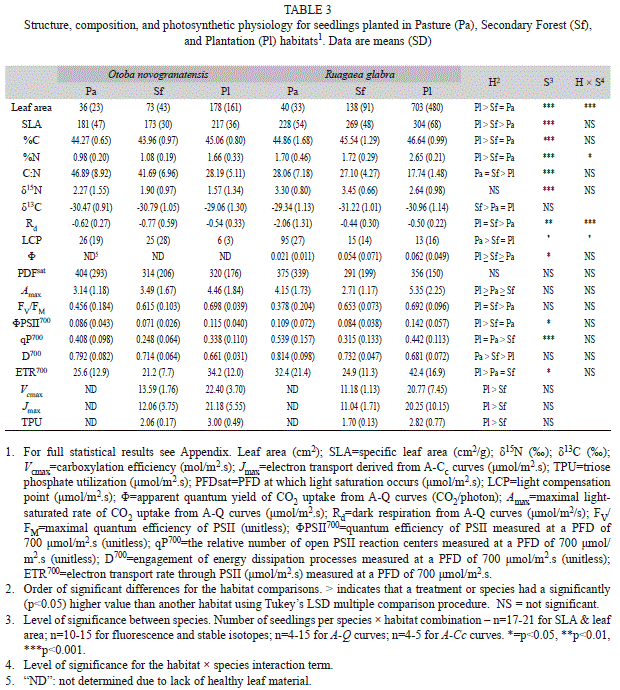
In general, structural, biochemical and physiological characteristics of leaves differed significantly between plantations and pastures with varying or intermediate responses in secondary forests (Table 3, Appendix). Both leaf area and specific leaf area (SLA) were highest in the plantations, intermediate in the secondary forests, and lowest in the pastures (Table 3, Appendix); there was a significant species × habitat interaction term for leaf area, as this trend was more pronounced for R. glabra. Percent C and N in leaves were both significantly higher in plantation habitats planted with N-fixing overstory trees than secondary forests and pastures, with the opposite pattern for C:N. δ15N was higher in R. glabra, but showed only a marginally significant habitat effect (F=2.8, p=0.0651) with a trend towards lower values in the plantation than the pasture sites.
Seedlings in plantations had the highest responses of photosynthetic CO2 assimilation to varying PFD for both species (Fig. 1, Table 3). Based on the light response curves, dark respiration (Rd) was lowest (i.e. least negative) for R. glabra in the plantation habitats, but was similar across habitats for O. novogranatensis (significant habitat × species interaction). The light compensation point (LCP) was lowest in the plantation and highest in the pasture for both species, but the secondary forest aver-age for R. glabra was similar to that in the plantations, whereas for O. novogranatensis it was similar to the pastures. Apparent quantum yield (Φ) was highest in plantations compared to secondary forests and pastures for R. glabra (the model analysis did not converge on a solution for Φ for O. novogranatensis). The light-saturated maximal rate of photosynthetic CO2 uptake (Amax) was higher in plantations than secondary forests, and was variable across species in pastures, likely due to the small sample size of surviving seedlings.
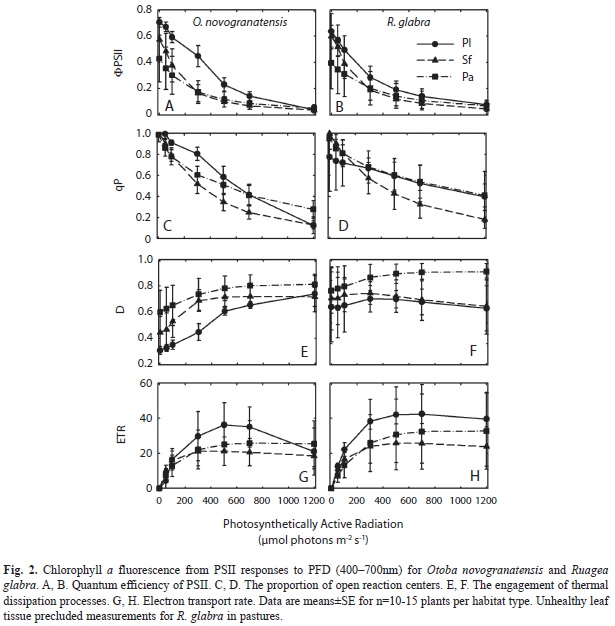
In general, PSII function decreased and thermal dissipation increased as a function of PFD for both species (Fig. 2). The apparent quantum efficiency of PSII for dark-adaptedleaves (FV/FM) was highest for seedlings in plantations and secondary forests, as compared to pastures (Table 3, Appendix). The quantum efficiency of PSII at a PFD of 700μmol/m2.s (ΦPSII700) was highest in plantations. The proportion of open PSII reaction centers (qP700) was highest in pastures and lowest in secondary forests. Thermal dissipation (D700) was highest in pasture habitats. The electron transport rate determined from PSII fluorescence (ETR700) was significantly higher in plantations and in R. glabra.
Based on the A-Ci curves (Fig. 3), both species had a higher photosynthetic capacity (Vcmax, Jmax, and TPU) in plantations than in secondary forests (Table 3, Appendix). Carboxylation efficiency (Vcmax) was 1.9 (R. glabra) and 1.6 fold (O. novogranatensis) higher in plantations compared to secondary forests. Seedlings in the pasture had small and yellowed leaves that did not provide A-Ci curves that could be analyzed by the software.
Discussion
Past studies have suggested that enrichment planting is an effective strategy for introducing later successional species into degraded tropical forest ecosystems (Peña-Claros et al. 2002, Romell et al. 2008, Doucet et al. 2009). Prior research at our study sites showed that seedlings were taller, had lower root:shoot ratios, and higher leaf N content when they were planted in tree plantations, compared to secondary forests or abandoned pastures (Cole et al. 2011). Our current results show that in this moist forest ecosystem, planting seedlings in the same plantations (which contain some N-fixing tree species) can enhance certain aspects of photosynthetic gas exchange, biochemistry, and energy processing within PSII for seedlings of the two species we examined and that micrometeorological conditions may play a lesser role in explaining their growth differences.
Planting seeds under plantation trees resulted in seedlings with greater aboveground growth. The δ15N results suggest that some of the N in leaves of seedlings in plantations is from biological N-fixation by the trees, although we note that this requires further study given that we found marginally (p=0.0615) lower δ15N in abandoned pastures. Higher leaf %N for seedlings in plantations is consistent with results showing more favorable biochemical characteristics associated with photosynthetic capacity and chlorophyll a fluorescence from Photosystem II (Bungard et al. 2002, Kitao et al. 2006, Cernusak et al. 2008, Reich et al. 2009). Moreover, the quantum efficiency of PSII in darkness (FV/FM) and at 700μmol/ m2.s (ΦPSII700), the rate of electron transport (ETR700), and the degree of thermal dissipation (D700) are all consistent with greater leaf N in the plantation habitats. By contrast, leaves of seedlings in the pasture showed signs of chronic photoinhibition. It should be noted that previous research shows that not all species exhibit a causal relationship between leaf N and photosynthetic assimilation (Aragao et al. 2005), and leaf P and other nutrients affect carbon gain as well (Meir et al. 2001, Reich et al. 2009), which may have affected some of the growth differences observed by Cole et al. (2011).
For both species, in plantations there was greater carboxylation efficiency by activated Rubisco, electron transport to drive regenera-tion of the carboxylation substrate RuBP, and triose-phosphate utilization, as determined by the A-Ci curves. These patterns are generally associated with an up-regulation of photosynthetic capacity in the presence of higher foliar N content (Coomes & Grubb 1998, Bungard et al. 2002, Coste et al. 2005, Reich et al. 2009), which could drive greater carbon gain, survival, and growth (Cole et al. 2011). Some aspects of the A-Q response (i.e. less-negative Rd, lower LCP, and higher Amax) were consistent with higher photosynthetic capacity for seedlings from the tree plantations in comparison to pasture and secondary forest habitats (Riddoch et al. 1991, Johnson et al. 1997, Ellis et al. 2000, Posada et al. 2009). When Rd and Amax are considered together, seedlings of both species in the pasture habitats had a lower net C budget compared to leaves from plants in plantations and secondary forests.
Higher N in seedlings in the plantations also appears to be associated with a greater flexibility to adjust PSII light harvesting. As expected with increasing light (PFD) levels, ΦPSII and qP decreased, and D and ETR increased. However, the magnitude of these effects differed across species, as determined by ΦPSII700, qP700 and ETR700. The apparent quantum yield for chlorophyll fluorescence from PSII (FV/FM) was significantly higher in the plantations and secondary forest habitats compared to the pastures. These species may differ across habitats in terms of their ability to undergo flexible thermal energy dissipation via a trans-thylakoid ∆pH and PsbS protein activity (Demmig-Adams & Adams 2006). The number of open PSII reaction centers was similar in the pastures and plantations, and lowest in the secondary forest habitats, which is more difficult to explain. Possibly the higher D in pastures allowed some PSII reaction centers to remain open, while higher leaf N in plantations accounted for higher ETR, with seedlings in secondary forests subject to lower leaf N and intermediate amounts of D. In other studies, some species show a down regulation of PSII efficiency under limited N (Verhoeven et al. 1997). It is possible that our results for ΦPSII, ETR and D across habitats may be due to varying levels of foliar N content that can support production of violaxanthin de-epoxidase, PsbS protein, and other components of PSII (Li et al. 2002, Niyogi et al. 2005, Logan et al. 2008) beyond simply affecting activated Rubisco quantity.
Our results suggest that soil water content and micrometeorological factors may be of lesser importance than foliar N for driving differences in photosynthetic processes across planting habitats. Although overstory cover was much higher and understory cover lower in plantation and secondary forest habitats, micro-meteorological conditions (PFD, Tair, Tsoil and RH) at seedling height were highly variable across sites and planting habitats, and did not show consistent patterns. The lack of micro-meteorological differences across treatments at seedling height (~0.2m) can be explained partly by the vegetation structure. Seedlings were shaded by the tree canopy in plantation and secondary forest sites and by grass cover (1-1.5m height) in pastures, which has been shown to result in relatively similar microclimatic conditions (Holl 1999). Water deficit was also not an apparent factor in the survival and growth of seedlings across sites and treatments, given the lack of habitat differences in soil moisture, plant water potential, or integrated WUE over the lifetime of the leaf, as determined by δ13C.
In some cases, physiological responses of seedlings in the secondary forest habitats were similar to seedlings in pastures (e.g. for leaf area, %C, %N, ΦPSII700, ETR700), whereas for other parameters responses in secondary forests were intermediate between those from pastures and plantations (e.g. SLA and D700), and in a few cases secondary forest values were similar to plantations (e.g. Rd, LCP, FV/FM). The reason for these patterns is not clear, but we offer a few potential possibilities. First, previous work at this study site has shown that while leaf litter N content is higher in the plantations compared to pastures and secondary forests, annual leaf litterfall inputs of K, Ca, Mg and Zn are higher in secondary forests, likely due to the higher diversity of tree species (Celentano et al. 2011). Higher inputs of these other nutrients may counteract the effects of lower N for certain photosynthetic processes. Second, grass cover and bare ground in secondary forests were intermediate to plantations and pastures, so differential competition with understory species may have affected our seedling responses, although experiments directly manipulating understory competition would be necessary to test this hypothesis. Third, our single-point microclimatic measurements may not have adequately characterized the high spatial complexity of the physical conditions in the three habitats.
Overall, our results show that enrichment planting under a mixed-species canopy is a successful strategy for introducing these two later successional tree species as part of tropical forest restoration efforts. Our results suggest that N-fixing trees in plantations may enhance the photosynthetic capacity and PSII efficiency for seedlings of O. novogranatensis and R. glabra and perhaps other tropical forest tree species. But, the role of N-fixation by trees for facilitating seedlings in this system is still not clear. These results highlight the importance of the question of which species to include as part of tropical forest restoration plantings in order to maximize the return on the time and finances invested to restore habitat diversity and functions.
Acknowledgments
We thank R.A. Zahawi and staff of the Las Cruces Biological Station for logistical assistance. We appreciate field assistance from R. Gomez, T. Kehoe and J.A. Rosales. D. Andreason performed the stable isotope analyses.
References
Amiard, V., K.E. Mueh, B. Demmig-Adams, V. Ebbert, R. Turgeon & W.W. Adams. 2005. Anatomical and photosynthetic acclimation to the light environment in species with differing mechanisms of phloem loading. Proc. Nat. Acad. Sci. USA 102: 12968-12973. [ Links ]
Anderson, J.M., W.S. Chow & J. De Las Rivas. 2008. Dynamic flexibility in the structure and function of photosystem II in higher plant thylakoid membranes: the grana enigma. Photosynth. Res. 98: 575-587. [ Links ]
Aragao, D.V., L.B. Fortini, S. Mulkey, D.J. Zarin, M.M. Araujo & C.J.R. De Carvalho. 2005. Correlation but no causation between leaf nitrogen and maximum assimilation: The role of drought and reproduction in gas exchange in an understory tropical plant Miconia ciliata (Melastomataceae). Am. J. Bot. 92: 456-461. [ Links ]
Bonilla-Moheno, M. & K.D. Holl. 2010. Direct seeding to restore tropical mature-forest species in areas of slash-and-burn agriculture. Restor. Ecol. 18: 438-445. [ Links ]
Bungard, R.A., S.A. Zipperlen, M.C. Press & J.D. Scholes. 2002. The influence of nutrients on growth and photosynthesis of seedlings of two rainforest dipterocarp species. Funct. Plant Biol. 29: 505-515. [ Links ]
Camargo, J.L.C., I.D.K. Ferraz & A.M. Imakawa. 2002. Rehabilitation of degraded areas of central Amazonia using direct sowing of forest tree seeds. Restor. Ecol. 10: 636-644. [ Links ]
Celentano, D., R.A. Zahawi, B. Finegan, R. Ostertag, R.J. Cole & K.D. Holl. 2011. Litterfall dynamics under different tropical forest restoration strategies. Biotropica 43: 279-287. [ Links ]
Cernusak, L.A., K. Winter, J. Aranda & B.L. Turner. 2008. Conifers, angiosperm trees, and lianas: Growth, whole-plant water and nitrogen use efficiency, and stable isotope composition (delta C-13 and delta O-18) of seedlings grown in a tropical environment. Plant Physiol. 148: 642-659. [ Links ]
Chazdon, R.L. 2008. Beyond deforestation: Restoring forests and ecosystem services on degraded lands. Science 320: 1458-1460. [ Links ]
Cole, R.J., K.D. Holl & R.A. Zahawi. 2010. Seed rain under tree islands planted to restore degraded lands in a tropical agricultural landscape. Ecol. App. 20: 1255-1269. [ Links ]
Cole, R.J., C. Keene, R.A. Zahawi & K.D. Holl. 2011. Direct seeding of late successional trees to restore tropical montane forest. For. Ecol. Manage. 261: 1590-1597. [ Links ]
Coomes, D.A. & P.J. Grubb. 1998. Responses of juvenile trees to above- and belowground competition in nutrient-starved Amazonian rain forest. Ecology 79: 768-782. [ Links ]
Coste, S., J.C. Roggy, P. Imbert, C. Born, D. Bonal & E. Dreyer. 2005. Leaf photosynthetic traits of 14 tropical rain forest species in relation to leaf nitrogen concentration and shade tolerance. Tree Physiol. 25: 1127-1137. [ Links ]
Cusack, D. & F. Montagnini. 2004. The role of native species plantations in recovery of understory woody diversity in degraded pasturelands of Costa Rica. For. Ecol. Manage. 188: 1-15. [ Links ]
Demmig-Adams, B. & W.W. Adams. 2006. Photoprotection in an ecological context: the remarkable complexity of thermal energy dissipation. New Phytol. 172: 11-21. [ Links ]
Doucet, J.L., Y.L. Kouadio, D. Monticelli & P. Lejeune. 2009. Enrichment of logging gaps with moabi (Baillonella toxisperma Pierre) in a Central African rain forest. For. Ecol. Manage. 258: 2407-2415. [ Links ]
Ellis, A.R., S.P. Hubbell & C. Potvin. 2000. In situ field measurements of photosynthetic rates of tropical tree species: a test of the functional group hypothesis. Can. J. Bot. 78: 1336-1347. [ Links ]
Engel, V.L. & J.A. Parrotta. 2001. An evaluation of direct seeding for reforestation of degraded lands in central São Paulo state, Brazil. For. Ecol. Manage. 152: 169-181. [ Links ]
Holdridge, L.R., W.C. Grenke, W.H. Hatheway, T. Liang & J.A. Tosi Jr. 1971. Forest environments in tropical life zones. Pergamon, Oxford. [ Links ]
Holl, K.D. 1999. Factors limiting tropical rain forest regeneration in abandoned pasture: seed rain, seed germination, microclimate, and soil. Biotropica 31: 229-241. [ Links ]
Holl, K.D. 2002. Tropical moist forest, p. 539-558. In M.R. Perrow & A.J. Davy (eds.). Handbook of ecological restoration. Cambridge University, Cambridge, United Kingdom. [ Links ]
Holl, K.D. & T.M. Aide. 2011. When and where to actively restore ecosystems? For. Ecol. Manage. 261: 1588-1563. [ Links ]
Holl, K.D., R.A. Zahawi, R.J. Cole, R. Ostertag & S. Cordell. 2011. Planting seedlings in plantations versus tree islands as a large-scale tropical forest restoration strategy. Restor. Ecol. 19: 470-479. [ Links ]
Johnson, J.D., R. Tognetti, M. Michelozzi, S. Pinzauti, G. Minotta & M. Borghetti. 1997. Ecophysiological responses of Fagus sylvatica seedlings to changing light conditions. The interaction of light environment and soil fertility on seedling physiology. Physiol. Plantarum 101: 124-134. [ Links ]
Kitao, M., R. Yoneda, H. Tobita, Y. Matsumoto, Y. Maruyama, A. Arifin, A.M. Azani & M.N. Muhamad. 2006. Susceptibility to photoinhibition in seedlings of six tropical fruit tree species native to Malaysia following transplantation to a degraded land. Trees-Struct. Funct. 20: 601-610. [ Links ]
Lamb, D., P.D. Erskine & J.D. Parrotta. 2005. Restoration of degraded tropical forest landscapes. Science 310: 1628-1632. [ Links ]
Li, X.P., P. Muller-Moule, A.M. Gilmore & K.K. Niyogi. 2002. PsbS-dependent enhancement of feedback de-excitation protects Photosystem II from photoinhibition. Proc. Nat. Acad. Sci. USA 99: 15222-15227. [ Links ]
Logan, B.A., S.G. Terry & K.K. Niyogi. 2008. Arabidopsis genotypes with differing levels of psbS expression differ in Photosystem II quantum yield, xanthophyll cycle pool size, and aboveground growth. Int. J. Plant Sci. 169: 597-604. [ Links ]
Loik, M.E. & K.D. Holl. 1999. Photosynthetic responses to light for rainforest seedlings planted in abandoned pasture, Costa Rica. Restor. Ecol. 7: 382-391. [ Links ]
Long, S.P. & C.J. Bernacchi. 2003. Gas exchange measurements, what can they tell us about the underlying limitations to photosynthesis? Procedures and sources of error. J. Exp. Bot. 54: 2393-2401. [ Links ]
Martínez-Garza, C. & H.F. Howe. 2003. Restoring tropical diversity: beating the time tax on species loss. J. App. Ecol. 40: 423-429. [ Links ]
Meir, P., J. Grace & A.C. Miranda. 2001. Leaf respiration in two tropical rainforests: constraints on physiology by phosphorus, nitrogen and temperature. Funct. Ecol. 15: 378-387. [ Links ]
Minchin, P.E.H. & A. Lacointe. 2005. New understanding on phloem physiology and possible consequences for modelling long-distance carbon transport. New Phytol. 166: 771-779. [ Links ]
Niyogi, K.K., X.P. Li, V. Rosenberg & H.S. Jung. 2005. Is PsbS the site of non-photochemical quenching in photosynthesis? J. Exp. Bot. 56: 375-382. [ Links ]
Pammenter, N.W., F. Loreto & T.D. Sharkey. 1993. End-product feedback effects on photosynthetic electron-transport. Photosynth. Res. 35: 5-14. [ Links ]
Peña-Claros, M., R.G.A. Boot, J. Dorado-Lora & A. Zonta. 2002. Enrichment planting of Bertholletia excelsa in secondary forest in the Bolivian Amazon: effect of cutting line width on survival, growth and crown traits. For. Ecol. Manage. 161: 159-168. [ Links ]
Piotto, D. 2007. Growth of native tree species planted in open pasture, young secondary forest and mature forest in humid tropical Costa Rica. J. Trop. For. Sci. 19: 92-102. [ Links ]
Pons, T.L. & R.A.M. Welschen. 2002. Overestimation of respiration rates in commercially available clampon leaf chambers. Complications with measurement of net photosynthesis. Plant Cell Environ. 25: 1367-1372. [ Links ]
Posada, J.M., M.J. Lechowicz & K. Kitajima. 2009. Optimal photosynthetic use of light by tropical tree crowns achieved by adjustment of individual leaf angles and nitrogen content. Ann. Bot-London 103: 795-805. [ Links ]
Raines, C.A. 2003. The Calvin cycle revisited. Photosynth. Res. 75: 1-10. [ Links ]
Reich, P.B., J. Oleksyn & I.J. Wright. 2009. Leaf phosphorus influences the photosynthesis-nitrogen relation: a cross-biome analysis of 314 species. Oecologia 160: 207-212. [ Links ]
Riddoch, I., T. Lehto & J. Grace. 1991. Photosynthesis of tropical tree seedlings in relation to light and nutrient supply. New Phytol. 119: 137-147. [ Links ]
Romell, E., G. Hallsby, A. Karlsson & C. García. 2008. Artificial canopy gaps in a Macaranga spp. dominated secondary tropical rain forest - Effects on survival and above ground increment of four under-planted dipterocarp species. For. Ecol. Manage. 255: 1452-1460. [ Links ]
Schatral, A. & J.E.D. Fox. 1994. Quality and viability of seeds in the genus Hibbertia. Seed Sci. Technol. 22: 273-284. [ Links ]
Siddique, I., V.L. Engel, J.A. Parrotta, D. Lamb, G.B. Nardoto, J. Ometto, L.A. Martinelli & S. Schmidt. 2008. Dominance of legume trees alters nutrient relations in mixed species forest restoration plantings within seven years. Biogeochemistry 88: 89-101. [ Links ]
Sims, D.A. & R.W. Pearcy. 1991. Photosynthesis and respiration in Alocasia-macrorrhiza following transfers to high and low light. Oecologia 86: 447-453. [ Links ]
Vankooten, O. & J.F.H. Snel. 1990. The use of chlorophyll fluorescence nomenclature in plant stress physiology. Photosynth. Res. 25: 147-150. [ Links ]
Verhoeven, A.S., B. DemmigAdams & W.W. Adams. 1997. Enhanced employment of the xanthophyll cycle and thermal energy dissipation in spinach exposed to high light and N stress. Plant Physiol. 113: 817-824. [ Links ]
Zimmerman, J.K., T.M. Aide & A.E. Lugo. 2007. Implications of land use history for natural forest regeneration and restoration strategies in Puerto Rico, p. 51-74. In R.J. Hobbs & V.A. Cramer (eds.). Old fields, Island, Washington, DC, USA. [ Links ]
Anderson, J.M., W.S. Chow & J. De Las Rivas. 2008. Dynamic flexibility in the structure and function of photosystem II in higher plant thylakoid membranes: the grana enigma. Photosynth. Res. 98: 575-587. [ Links ]
Aragao, D.V., L.B. Fortini, S. Mulkey, D.J. Zarin, M.M. Araujo & C.J.R. De Carvalho. 2005. Correlation but no causation between leaf nitrogen and maximum assimilation: The role of drought and reproduction in gas exchange in an understory tropical plant Miconia ciliata (Melastomataceae). Am. J. Bot. 92: 456-461. [ Links ]
Bonilla-Moheno, M. & K.D. Holl. 2010. Direct seeding to restore tropical mature-forest species in areas of slash-and-burn agriculture. Restor. Ecol. 18: 438-445. [ Links ]
Bungard, R.A., S.A. Zipperlen, M.C. Press & J.D. Scholes. 2002. The influence of nutrients on growth and photosynthesis of seedlings of two rainforest dipterocarp species. Funct. Plant Biol. 29: 505-515. [ Links ]
Camargo, J.L.C., I.D.K. Ferraz & A.M. Imakawa. 2002. Rehabilitation of degraded areas of central Amazonia using direct sowing of forest tree seeds. Restor. Ecol. 10: 636-644. [ Links ]
Celentano, D., R.A. Zahawi, B. Finegan, R. Ostertag, R.J. Cole & K.D. Holl. 2011. Litterfall dynamics under different tropical forest restoration strategies. Biotropica 43: 279-287. [ Links ]
Cernusak, L.A., K. Winter, J. Aranda & B.L. Turner. 2008. Conifers, angiosperm trees, and lianas: Growth, whole-plant water and nitrogen use efficiency, and stable isotope composition (delta C-13 and delta O-18) of seedlings grown in a tropical environment. Plant Physiol. 148: 642-659. [ Links ]
Chazdon, R.L. 2008. Beyond deforestation: Restoring forests and ecosystem services on degraded lands. Science 320: 1458-1460. [ Links ]
Cole, R.J., K.D. Holl & R.A. Zahawi. 2010. Seed rain under tree islands planted to restore degraded lands in a tropical agricultural landscape. Ecol. App. 20: 1255-1269. [ Links ]
Cole, R.J., C. Keene, R.A. Zahawi & K.D. Holl. 2011. Direct seeding of late successional trees to restore tropical montane forest. For. Ecol. Manage. 261: 1590-1597. [ Links ]
Coomes, D.A. & P.J. Grubb. 1998. Responses of juvenile trees to above- and belowground competition in nutrient-starved Amazonian rain forest. Ecology 79: 768-782. [ Links ]
Coste, S., J.C. Roggy, P. Imbert, C. Born, D. Bonal & E. Dreyer. 2005. Leaf photosynthetic traits of 14 tropical rain forest species in relation to leaf nitrogen concentration and shade tolerance. Tree Physiol. 25: 1127-1137. [ Links ]
Cusack, D. & F. Montagnini. 2004. The role of native species plantations in recovery of understory woody diversity in degraded pasturelands of Costa Rica. For. Ecol. Manage. 188: 1-15. [ Links ]
Demmig-Adams, B. & W.W. Adams. 2006. Photoprotection in an ecological context: the remarkable complexity of thermal energy dissipation. New Phytol. 172: 11-21. [ Links ]
Doucet, J.L., Y.L. Kouadio, D. Monticelli & P. Lejeune. 2009. Enrichment of logging gaps with moabi (Baillonella toxisperma Pierre) in a Central African rain forest. For. Ecol. Manage. 258: 2407-2415. [ Links ]
Ellis, A.R., S.P. Hubbell & C. Potvin. 2000. In situ field measurements of photosynthetic rates of tropical tree species: a test of the functional group hypothesis. Can. J. Bot. 78: 1336-1347. [ Links ]
Engel, V.L. & J.A. Parrotta. 2001. An evaluation of direct seeding for reforestation of degraded lands in central São Paulo state, Brazil. For. Ecol. Manage. 152: 169-181. [ Links ]
Holdridge, L.R., W.C. Grenke, W.H. Hatheway, T. Liang & J.A. Tosi Jr. 1971. Forest environments in tropical life zones. Pergamon, Oxford. [ Links ]
Holl, K.D. 1999. Factors limiting tropical rain forest regeneration in abandoned pasture: seed rain, seed germination, microclimate, and soil. Biotropica 31: 229-241. [ Links ]
Holl, K.D. 2002. Tropical moist forest, p. 539-558. In M.R. Perrow & A.J. Davy (eds.). Handbook of ecological restoration. Cambridge University, Cambridge, United Kingdom. [ Links ]
Holl, K.D. & T.M. Aide. 2011. When and where to actively restore ecosystems? For. Ecol. Manage. 261: 1588-1563. [ Links ]
Holl, K.D., R.A. Zahawi, R.J. Cole, R. Ostertag & S. Cordell. 2011. Planting seedlings in plantations versus tree islands as a large-scale tropical forest restoration strategy. Restor. Ecol. 19: 470-479. [ Links ]
Johnson, J.D., R. Tognetti, M. Michelozzi, S. Pinzauti, G. Minotta & M. Borghetti. 1997. Ecophysiological responses of Fagus sylvatica seedlings to changing light conditions. The interaction of light environment and soil fertility on seedling physiology. Physiol. Plantarum 101: 124-134. [ Links ]
Kitao, M., R. Yoneda, H. Tobita, Y. Matsumoto, Y. Maruyama, A. Arifin, A.M. Azani & M.N. Muhamad. 2006. Susceptibility to photoinhibition in seedlings of six tropical fruit tree species native to Malaysia following transplantation to a degraded land. Trees-Struct. Funct. 20: 601-610. [ Links ]
Lamb, D., P.D. Erskine & J.D. Parrotta. 2005. Restoration of degraded tropical forest landscapes. Science 310: 1628-1632. [ Links ]
Li, X.P., P. Muller-Moule, A.M. Gilmore & K.K. Niyogi. 2002. PsbS-dependent enhancement of feedback de-excitation protects Photosystem II from photoinhibition. Proc. Nat. Acad. Sci. USA 99: 15222-15227. [ Links ]
Logan, B.A., S.G. Terry & K.K. Niyogi. 2008. Arabidopsis genotypes with differing levels of psbS expression differ in Photosystem II quantum yield, xanthophyll cycle pool size, and aboveground growth. Int. J. Plant Sci. 169: 597-604. [ Links ]
Loik, M.E. & K.D. Holl. 1999. Photosynthetic responses to light for rainforest seedlings planted in abandoned pasture, Costa Rica. Restor. Ecol. 7: 382-391. [ Links ]
Long, S.P. & C.J. Bernacchi. 2003. Gas exchange measurements, what can they tell us about the underlying limitations to photosynthesis? Procedures and sources of error. J. Exp. Bot. 54: 2393-2401. [ Links ]
Martínez-Garza, C. & H.F. Howe. 2003. Restoring tropical diversity: beating the time tax on species loss. J. App. Ecol. 40: 423-429. [ Links ]
Meir, P., J. Grace & A.C. Miranda. 2001. Leaf respiration in two tropical rainforests: constraints on physiology by phosphorus, nitrogen and temperature. Funct. Ecol. 15: 378-387. [ Links ]
Minchin, P.E.H. & A. Lacointe. 2005. New understanding on phloem physiology and possible consequences for modelling long-distance carbon transport. New Phytol. 166: 771-779. [ Links ]
Niyogi, K.K., X.P. Li, V. Rosenberg & H.S. Jung. 2005. Is PsbS the site of non-photochemical quenching in photosynthesis? J. Exp. Bot. 56: 375-382. [ Links ]
Pammenter, N.W., F. Loreto & T.D. Sharkey. 1993. End-product feedback effects on photosynthetic electron-transport. Photosynth. Res. 35: 5-14. [ Links ]
Peña-Claros, M., R.G.A. Boot, J. Dorado-Lora & A. Zonta. 2002. Enrichment planting of Bertholletia excelsa in secondary forest in the Bolivian Amazon: effect of cutting line width on survival, growth and crown traits. For. Ecol. Manage. 161: 159-168. [ Links ]
Piotto, D. 2007. Growth of native tree species planted in open pasture, young secondary forest and mature forest in humid tropical Costa Rica. J. Trop. For. Sci. 19: 92-102. [ Links ]
Pons, T.L. & R.A.M. Welschen. 2002. Overestimation of respiration rates in commercially available clampon leaf chambers. Complications with measurement of net photosynthesis. Plant Cell Environ. 25: 1367-1372. [ Links ]
Posada, J.M., M.J. Lechowicz & K. Kitajima. 2009. Optimal photosynthetic use of light by tropical tree crowns achieved by adjustment of individual leaf angles and nitrogen content. Ann. Bot-London 103: 795-805. [ Links ]
Raines, C.A. 2003. The Calvin cycle revisited. Photosynth. Res. 75: 1-10. [ Links ]
Reich, P.B., J. Oleksyn & I.J. Wright. 2009. Leaf phosphorus influences the photosynthesis-nitrogen relation: a cross-biome analysis of 314 species. Oecologia 160: 207-212. [ Links ]
Riddoch, I., T. Lehto & J. Grace. 1991. Photosynthesis of tropical tree seedlings in relation to light and nutrient supply. New Phytol. 119: 137-147. [ Links ]
Romell, E., G. Hallsby, A. Karlsson & C. García. 2008. Artificial canopy gaps in a Macaranga spp. dominated secondary tropical rain forest - Effects on survival and above ground increment of four under-planted dipterocarp species. For. Ecol. Manage. 255: 1452-1460. [ Links ]
Schatral, A. & J.E.D. Fox. 1994. Quality and viability of seeds in the genus Hibbertia. Seed Sci. Technol. 22: 273-284. [ Links ]
Siddique, I., V.L. Engel, J.A. Parrotta, D. Lamb, G.B. Nardoto, J. Ometto, L.A. Martinelli & S. Schmidt. 2008. Dominance of legume trees alters nutrient relations in mixed species forest restoration plantings within seven years. Biogeochemistry 88: 89-101. [ Links ]
Sims, D.A. & R.W. Pearcy. 1991. Photosynthesis and respiration in Alocasia-macrorrhiza following transfers to high and low light. Oecologia 86: 447-453. [ Links ]
Vankooten, O. & J.F.H. Snel. 1990. The use of chlorophyll fluorescence nomenclature in plant stress physiology. Photosynth. Res. 25: 147-150. [ Links ]
Verhoeven, A.S., B. DemmigAdams & W.W. Adams. 1997. Enhanced employment of the xanthophyll cycle and thermal energy dissipation in spinach exposed to high light and N stress. Plant Physiol. 113: 817-824. [ Links ]
Zimmerman, J.K., T.M. Aide & A.E. Lugo. 2007. Implications of land use history for natural forest regeneration and restoration strategies in Puerto Rico, p. 51-74. In R.J. Hobbs & V.A. Cramer (eds.). Old fields, Island, Washington, DC, USA. [ Links ]
*Correspondencia a:
Michael E. Loik. Department of Environmental Studies, University of California, Santa Cruz, CA 95064, U.S.A.; mloik@ucsc.edu
Rebecca J. Cole. Department of Environmental Studies, University of California, Santa Cruz, CA 95064, U.S.A.; cole.rebeccaj@gmail.com
Karen D. Holl. Department of Environmental Studies, University of California, Santa Cruz, CA 95064, U.S.A.; kholl@ucsc.edu
Gabriel C. Sady. Department of Environmental Studies, University of California, Santa Cruz, CA 95064, U.S.A.; gabrielsady@gmail.com
1. Department of Environmental Studies, University of California, Santa Cruz, CA 95064, U.S.A.; mloik@ucsc.edu, cole.rebeccaj@gmail.com, kholl@ucsc.edu, gabrielsady@gmail.com
Received 19-VII-2012. Corrected 12-XII-2012. Accepted 24-I-2013.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}