










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.61 n.3 San José Sep. 2013
Respuesta de la regeneración natural en manglares del Golfo de Urabá (Colombia) a la variabilidad ambiental y climática intra-anual
Natural regeneration response in mangroves of the Gulf of Urabá (Colombia) to the enviromental and intra-annual climate variabiblity
Natural regeneration response in mangroves of the Gulf of Urabá (Colombia) to the enviromental and intra-annual climate variabiblity
*Dirección para correspondencia:
Abstract
The natural regeneration process allows the mangrove forests remain over time. Both, biological and physical factors can affect the establishment and early stages along the development of trees. This study examined the response of natural regeneration of mangroves in the Turbo River delta and El Uno bay (Urabá Gulf, Colombia) to intra-annual environmental variability. We quantified mortality, survival and recruitment of seedlings of three mangrove species, seasonally during one year, in 72 semi-perma-nent sub-plots of 1m2. In the sub-plots, the total height and the diameter at the base of the stem of all mangrove seedlings with basal diameter less than 2.5cm were measured. Damage by herbivores was also recorded to each seedling. While Laguncularia racemosa recorded the highest rates of mortality, Rhizophora mangle showed the highest survival rate during the study period, although Avicennia germinans dominated the natural regeneration. Through a Redundancy Analysis these processes were associated to environmental variables such as: Direct Site Factor-DSF (sunlight), sediments input rate, herbivory, distances from mangroves to the river, to inner lakes, and to the coastline. These variables explained 43% of the natural regeneration variation; sedimentation rate was the most important variable, while light was very representative for the R. mangle survival. Based on historical records of precipitation, Turbo River flow rate and associated sediment loads, it was established that during the highest precipitation peak, the survival of all species decreases and during the dry season, when the conditions of flooding and sediments input were lower, it was improved. The results indicated that the sediments input rates and sunlight play an important role in the survival of natural regeneration of evaluated mangrove species.
Key words: natural regeneration, Rhizophora mangle, Avicennia germinans, Laguncularia racemosa, variables.
Resumen
En los bosques de mangle, tanto factores biológicos como físicos, afectan el establecimiento y los estados tempranos de desarrollo de los árboles y determinan la distribución espacial de las especies. Se evaluó la respuesta de la regeneración natural de los manglares del delta del río Turbo y la bahía El Uno a la variabilidad ambiental y climática intra-anual. Se cuantificaron la mortalidad, la sobrevivencia y el reclutamiento de las plántulas de tres especies, estacionalmente durante un año. Avicennia germinans dominó la regeneración natural, Rhizophora mangle presentó la mayor la supervivencia y Laguncularia racemosa la mayor mortalidad. Estos procesos se asociaron principalmente con el aporte de sedimentos aluviales, la disponibilidad de luz, la herbivoría, la distancia al río, a las lagunas internas y a la línea de costa, las cuales explicaron el 43% de la variación en la regeneración. La sedimentación fue la variable más importante para el reclutamiento de A. germinans, mientras la luz fue muy representativa para la supervivencia de R. mangle. Con base en registros históricos de precipitación, caudales y carga de sedimentos, se estableció que en la época con mayor precipitación se presenta un descenso en la supervivencia, y en la época seca se favorece el balance entre el reclutamiento y la mortalidad. Los resultados indicaron que la precipitación, el aporte de sedimentos y la luz desempeñan un papel importante en el proceso de la regeneración de las especies evaluadas.
Palabras clave: regeneración natural, Rhizophora mangle, Avicennia germinans, Laguncularia racemosa, variables ambientales, variables climáticas.
La regeneración natural permite a las especies vegetales permanecer a través del tiempo en un bosque en particular (Asquith 2002). En los bosques de mangle, tanto factores biológicos (herbivoría, competencia, depredación) como físicos (disponibilidad de luz, salinidad nivel del mar, aportes aluviales de agua y sedimentos), afectan el establecimiento y los estados tempranos de desarrollo de los árboles (Krauss et al. 2008) y determinan la distribución espacial de las especies (Delgado et al. 2001).
La permanencia y recuperación de los manglares después de una perturbación dependen principalmente de la producción y dispersión de los propágulos, así como del establecimiento exitoso de las plantas (Tomlinson 1999). Los propágulos pueden sufrir alta depredación durante su dispersión y establecimiento especialmente por cangrejos e insectos que afectan la supervivencia de estos (Ellison & Farnsworth 1993, McGuinness 1997, Farnsworth & Ellison 1997, Dahdouh-Guebas et al. 1998, Delgado et al. 2001, Lema et al. 2003, Zhang et al. 2006). Así mismo, la supervivencia y el crecimiento, posteriores al establecimiento, están fuertemente influenciados por el estrés fisicoquímico (Krauss et al. 2008), especialmente por la intensidad lumínica, debido a que los árboles de mangle están expuestos a intensidades altas de ésta y se consideran intolerantes a la sombra (Janzen 1985). Sin embargo, la tasa fotosintética de los manglares tiende a estabilizarse en niveles relativamente bajos de luz (Kathiresan & Bingham 2001) e incluso pueden presentar un mayor crecimiento con niveles de irradiación significativamente más bajos que los recibidos de la luz directa (Kathiresan & Moorthy 1993, Farnsworth & Ellison 1996, McGuiness 1997). En el caso particular de R. mangle, este presenta cambios ontogénicos en su adaptación a la luz: mientras las plántulas se adaptan al sotobosque sombreado, los árboles maduros se desempeñan mejor a plena exposición solar (Farnsworth & Ellison 1996).
La disponibilidad de luz no es el único factor que interviene en el crecimiento de los manglares. Se ha identificado que una salinidad alta del agua intersticial puede restringir el crecimiento de las plántulas de A. germinans y L. racemosa, independientemente de que cuenten con alta disponibilidad de luz (Medina et al. 1990, Benfield et al. 2005, López-Hoffman et al. 2006). A su vez, la salinidad se ve afectada por la posición de los manglares en la llanura intermareal, así como por los cambios en el nivel de las mareas, lo que puede afectar la distribución y supervivencia de las plántulas (Ellison & Farnsworth 1993, 1996, McKee 1995). Por ejemplo, en Belice las plántulas R. mangle presentaron mayor supervivencia en mareas muy bajas y medias, mientras que las de A. germinans no sobrevivieron en mareas muy bajas y si en las medias (Ellison & Farnsworth 1993). Las plántulas de L. racemosa, en Panamá, sobrevivieron en todos los intervalos de marea (Benfield et al. 2005).
La variación estacional intra-anual, en la precipitación y en los caudales de los ríos, también afecta la distribución y supervivencia de plántulas en los manglares (Ellison & Farnsworth 1996), no solo por la influencia sobre la salinidad, sino también porque aumenta la carga de sedimentos depositados (Thampanya et al. 2002a), que puede sepultar las plántulas (Terrados et al. 1997).
Los manglares del Golfo de Urabá se caracterizan por una dinámica fluvio-marina activa y marcados procesos antrópicos (Blanco et al. 2012), como la desviación del delta del Río Turbo registrada en 1954, que aceleró los procesos de sedimentación y erosión en las zonas aledañas (Correa & Vernette 2004), propiciando el avance de la barra arenosa de Punta Yarumal (Estrada & Gil 2005), y la expansión de los manglares en algunos sitios. En la bahía El Uno, además, del aprovechamiento maderero intenso de los manglares por las comunidades (CORPOURABÁ 2003), los cambios en la sedimentación aluvial y el nivel del mar, están alterando su composición específica y supervivencia. Por ello, esta investigación se propuso evaluar la respuesta de la regeneración natural de A. germinans, L. racemosa y R. mangle a la variabilidad ambiental estacional durante un año. Nuestra hipótesis de trabajo postula que la luz y la sedimentación influyen en la mortalidad, sobrevivencia y reclutamiento de estas especies. Paralelamente, previo a dicha evaluación se describió la estructura y composición de los manglares para determinar el efecto del dosel sobre la regeneración.
Materiales y Métodos
Esta investigación se realizó en los manglares de la Bahía El Uno y la punta Yarumal, del Golfo de Urabá (08º05’42” - 76º44’23” W), esquina nor-occidental de Colombia, a 1 msnm, temperatura promedio de 28ºC (Keep 2000), precipitación media anual de 2 500mm, distribución bimodal, y un periodo lluvioso entre mayo y noviembre, una leve disminución en junio y julio, y un periodo seco entre diciembre y abril (Lozano 1998). Durante la época seca es común el fenómeno de mar de leva que causa oleaje fuerte (CIOH 2010). Internamente en El Golfo se registran promedios de vientos inferiores a 4m/s, con predominio de la dirección sur. Allí, el régimen de mareas se clasifica como micromareal, con altura media mínima de 0.50m y máxima de 0.90m (SHOM 1982, IDEAM 2007). Los manglares crecen sobre depósitos aluviales no consolidados de origen Cuaternario (Molina et al. 1998). Tanto La bahía El Uno como la punta Yarumal se encuentran en un área de acreción (INVEMAR & CORPOURABÁ 2003).
Previo al análisis de la regeneración natural, se elaboró un mapa preliminar de tipos estructurales de manglares a escala 1:10 000, con base en la interpretación de imágenes satelitales de 2004 disponibles en Google Earth (versión 6.0.2.2074), Google Inc. (2009). Con base en éste, se establecieron 18 parcelas circulares de 500m2 (Fig. 1) en las cuales se midieron diámetros a la altura del pecho (DAP) y alturas totales a todos los árboles mayores de 10cm de DAP. En cada parcela, se midió conductividad eléctrica del agua de inundación, salinidad, pH y oxígeno disuelto en el agua intersticial. Así mismo, se tomaron muestras combinadas de suelos de los primeros 25cm, a las cuales se les realizó análisis granulométrico y estándar de fertilidad, en el laboratorio de suelos de la Universidad Nacional de Colombia, Sede Medellín.
Para evaluar la regeneración natural, las 18 parcelas circulares de 500m2 se dividieron en cuatro subparcelas circulares semi-permanentes de 1m2. Se realizaron tres censos semestrales en 72 sub-parcelas. En el primer censo, realizado durante la época de lluvias (agosto 2009), se identificaron y marcaron todas las plántulas de manglar con diámetro inferior a 2.5cm; se les midió altura total y diámetro en la base del tallo, y se registraron señales de agresión por herbívoros. En cada sub-parcela se caracterizó el ambiente lumínico, a través de la medición del factor de sitio directo (FSD) y el factor de sitio indirecto (FSI) por medio de fotografías hemisféricas, posteriormente analizadas con los software HemiView 2.10 (Delta-T Devices, Universidad de Kansas, U.S.A) y SideLook 1.10 (M. Nobis 2005, University of Technology, Darmstadt, Germany and Swiss Federal Institute of Technology, Zurich, Switzerland). Igualmente, se midió, por medio de la marcación de horizontes con feldespato (Van Wijnen & Bakker 2001), la cantidad de sedimentos acumulados en cuadrantes de 0.05m2 adyacentes a cada una de las 72 sub-parcelas. Pasado un año se extrajo un núcleo de sedimentos que permitió establecer la cantidad de material acumulado por encima de este horizonte. Sobre el mapa, se midieron las distancias de las sub-parcelas a las lagunas internas al río Turbo, a la línea de costa (noroccidente) y dentro de la bahía El Uno. El segundo censo se realizó a los seis meses de establecidas las sub-parcelas, durante la época seca (enero 2010), y el tercero a los 12 meses, nuevamente al inicio de la época lluviosa (julio 2010). En ambos censos se evaluaron el reclutamiento y la mortalidad de las plántulas.
Para establecer la dominancia de las especies en la regeneración, se calculó el Índice de Valor de Importancia (IVI) (Curtis & McIntosh 1951). Además, los porcentajes de mortalidad, supervivencia y reclutamiento tanto por especie como por tipo de manglar. Los datos de mortalidad y reclutamiento del primer censo se tomaron como punto de referencia (0%) para los otros dos censos, por lo cual la densidad del primer censo se asumió como el 100%, y como valor inicial de la supervivencia. La relación entre las variables ambientales y los valores de mortalidad, supervivencia y reclutamiento por especie, y por sub-parcela, después de un año, se estableció por medio de un Análisis de Redundancia-(AR) con el software CANOCO 4.5 (Ter Braak & Smilauer 1998).
Para establecer denso- dependencia por especie y la mortalidad total, se realizó un análisis de varianza (STATGRAPHICS Plus 5.10 Manugistics, Inc, 1995) con cuatro categorías de mortalidad asumidas como: Baja (M1:0-30%), Media (M2:31-50%), Alta (M3:51-80%), Muy alta (M4:≥81%), en tres de densidades: Baja (D1:0-10%), Media (D2:11-50%) y Alta (D3:≥51%). La significancia de las diferencias de medias de la mortalidad y la densidad de las especies se evaluaron por una comparación múltiple de diferencia mínima significativa (LSD) (Fisher 1935).
Para establecer relaciones de la mortalidad, la supervivencia y el reclutamiento con la precipitación, los caudales del río Turbo y la carga de sedimentos transportados por éste, se utilizaron los valores históricos promedios de precipitación (1984-2006), caudales del río Turbo (1987-2002) y de sedimentos (1990-1999), proporcionados por el Instituto de Hidrología, Meteorología y Estudios Ambientales (IDEAM), los cuales fueron relacionados con los porcentajes de supervivencia por medio de un análisis descriptivo. De la misma manera, se relacionaron cualitativamente con los registros de magnitud y dirección de vientos (1949-1984) y altura de ola media significante (Hsm) (1979-2006), tomados en la estación meteorológica Turbo y registrados en Molina et al. (2011).
Resultados
Tres tipos de manglares se identificaron como coincidentes con los descritos en el sistema de clasificación de Lugo & Snedaker (1974): i) Manglares de cuenca (Parcelas 1, 2, 3, 10, 11, 15, 16 y 17), dominados por A. germinans, situados sobre las barras arenosas (superficies de acreción), suelos bien drenados, cercanos a tierra firme, y con los mayores valores promedio de sedimentación (5.10cm/año) (Anexo 1). ii) Manglares de borde (Parcelas 4, 6, 9, 12 y 13), dominados por R. mangle, que rodean la zona interior de la bahía El Uno y las lagunas internas de punta Yarumal, sobre suelos más inestables y con los mayores porcentajes promedio de materia orgánica en el suelo (10.76%) (Anexo 1). iii) Manglares ribereños (Parcelas 5, 7, 8, 14 y 18), dominados por L. racemosa, pero con alta abundancia de R. mangle y A. germinans, ubicados detrás de los manglares de borde, en zonas irrigadas por caños interiores de agua dulce, con el dosel más abierto y por ello, con los mayores valores promedio de los índices de luz FSI y FSD (0.35 y 0.38, respectivamente) (Anexo 1).
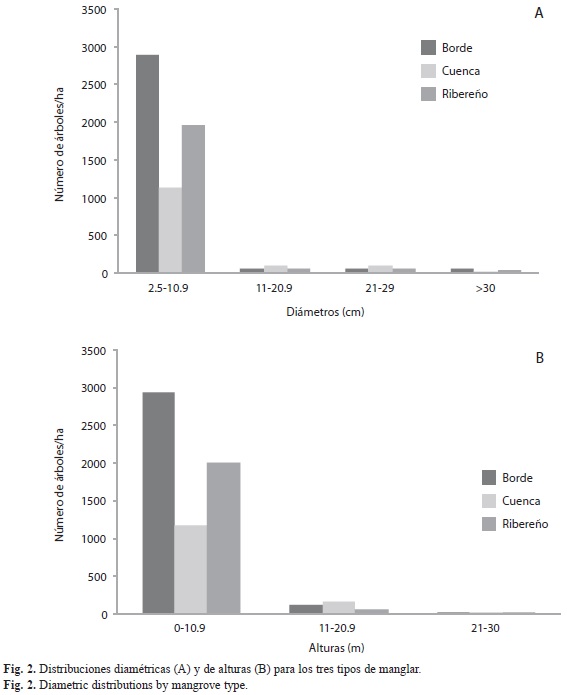
Las distribuciones diamétricas de los tres tipos de manglares (Fig. 2A) presentaron forma de “J” invertida, que expresa un alto potencial de regeneración y una muy baja representación de árboles en las categorías superiores. La mayor cantidad de individuos se registró en la categoría entre 2.5-10.9cm de DAP y dentro de ésta, los manglares de borde obtuvieron el mayor valor (2 888 árboles/ha). En las siguientes categorías (11-20.9 y 21-29cm de DAP) los manglares de cuenca obtuvieron el mayor número de individuos con 97 y 90 árboles/ ha, respectivamente. En la última categoría (mayores a 30cm de DAP) los manglares de borde presentaron el mayor número de individuos (56 árboles/ha).
Las distribuciones de alturas (Fig. 2B) también presentaron forma de “J” invertida. El mayor número de árboles (6 094 árboles/ha) se ubica en la categoría de altura de 0-10.9metros, para todos los tipos de manglares, superando ampliamente las otras dos. En esta categoría, los manglares de borde abarcaron el mayor número (2 928 árboles/ha). En la categoría de 11-20.9 metros de altura, los manglares de cuenca presentan el mayor número (165 árboles/ha). En la categoría de 21-30metros los manglares de borde registran el mayor número (28 árboles/ha).
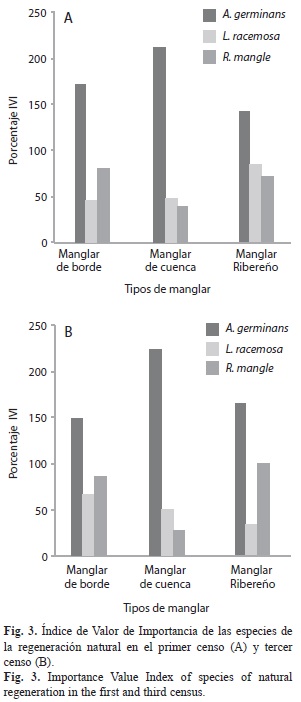
Según el IVI, la composición de la regeneración natural en el primer censo estuvo dominada por A. germinans en los tres tipos de manglares (Fig. 3A). Particularmente, en los manglares de borde R. mangle presentó el segundo lugar, mientras que en los de cuenca y ribereños el segundo lugar lo ocupó L. racemosa, aunque con escasa diferencia con respecto a R. mangle. En el segundo censo se conservaron las tendencias en el IVI, pero en el tercer censo (Fig. 3B), se presentó una variación en los manglares ribereños al ocupar R. mangle el segundo lugar.
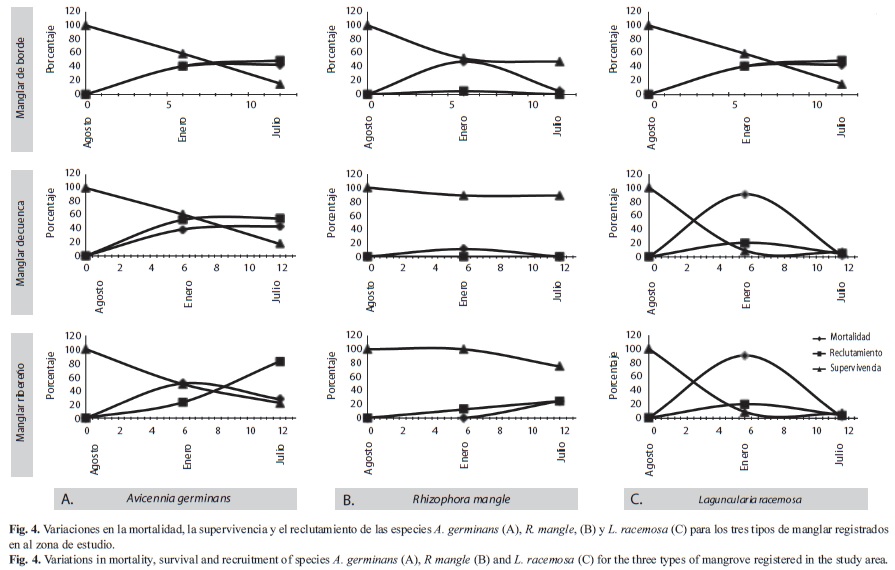
La figura 4 presenta las tasas de supervivencia, mortalidad y reclutamiento de la regeneración natural para las tres especies en los tres tipos de manglares.
En el segundo censo la mayor supervivencia en los manglares de borde la presentaron A. germinans y R. mangle (59.26% y 52.38%, respectivamente), mientras que el mayor reclutamiento lo presentó L. racemosa (80%). Entre los manglares de cuenca y ribereños, la mayor supervivencia la presentó R. mangle (88.89% y 100% respectivamente). En los tres tipos de manglares L. racemosa presentó la menor supervivencia, 47.27% (borde), 31.43% (cuenca) y 9.09% (ribereños). El mayor reclutamiento en los manglares de cuenca y ribereños los presentó A. germinans con (53.06) y (22.94), respectivamente.
En el tercer censo la supervivencia disminuyó en todos los tipos de manglar. En los de borde la mayor supervivencia la obtuvo R. mangle (47.62%), la menor L. racemosa (10.91%), mientras que el mayor reclutamiento lo presentó A. germinans (49.54%). En los manglares de cuenca la mayor supervivencia la presentó R. mangle (88.89%) y L. racemosa la menor (8.57%), mientras que A. germinans presentó el mayor reclutamiento (55.10%). De la misma forma, en los manglares ribereños R. mangle presentó el mayor porcentaje de sobrevivientes (75%) y L. racemosa el menor (6.82 %), mientras que A. germinans obtuvo el mayor porcentaje de reclutamiento (82.57%).
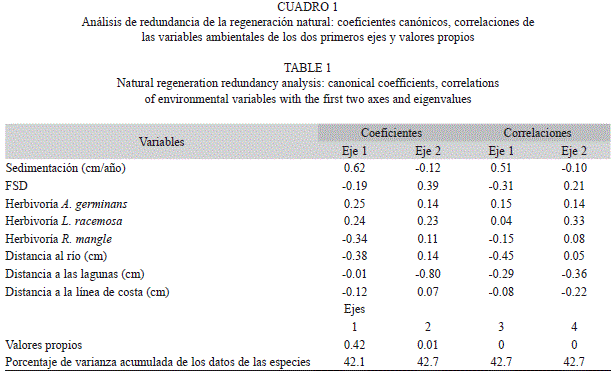
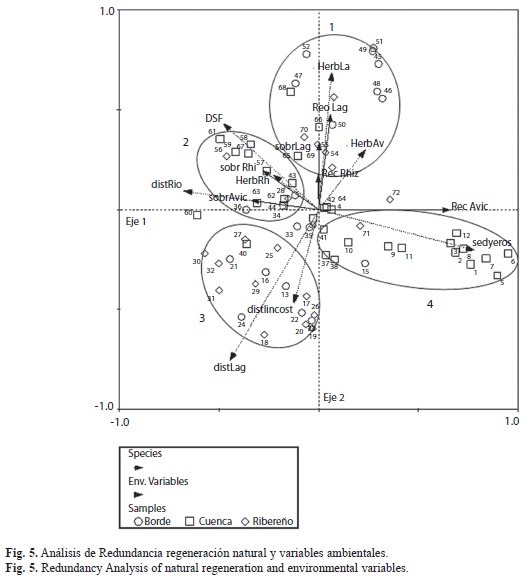
En el cuadro 1 y la figura 4, se presentan los resultados del Análisis de Redundancia (AR). Los dos primeros ejes explicaron el 43% de la variación de la regeneración. La tasa de sedimentación (R=0.51) y la distancia al río Turbo (R=-0.45) presentaron los mayores pesos en el primer eje, con coeficientes canónicos de 0.62 y -0.38, respectivamente. La distancia a las lagunas internas y la luz (FSD) presentaron los mayores pesos para el segundo eje, con coeficientes canónicos de -0.80 y 0.39, respectivamente. El segundo eje estuvo altamente correlacionado con la herbivoría sobre L. racemosa (R=0.33) y la distancia a las lagunas (R=-0.36).
Se identificaron cuatro grupos: el primero en la parte superior de la figura 5, agrupa las sub-parcelas asociadas a los mayores valores de herbivoría sobre A. germinans, reclutamiento y herbivoría de R. mangle, supervivencia y reclutamiento de L. racemosa. El segundo, incluye las sub -parcelas asociadas con los mayores valores de supervivencia de R. mangle y A. germinans, y de herbivoría de R. mangle, así como con la distancia al río Turbo y al factor de luz directa (SD). El vector que representa la supervivencia de A. germinans se presenta opuesto al vector de sedimentación y está directamente relacionado con la mayor distancia al río Turbo. El tercer grupo, está compuesto en su mayoría por las sub-parcelas pertenecientes a los manglares ribereños y de borde, asociados con la distancia al mar y a las lagunas. Por último, el cuarto grupo está conformado por las sub- parcelas asociadas al mayor reclutamiento de A. germinans y de acumulación de sedimentos (Fig. 5).
El análisis de varianza mostró denso-dependencia significativa, entre la mortalidad y la densidad de las plántulas (F3.21=14.17, p<0.01). En la relación entre mortalidad y densidad de plántulas, las pruebas de rangos múltiples mostraron diferencias significativas al 95% de confianza, entre R. mangle y A. germinans y entre R. mangle y L. racemosa, pero no entre A. germinans y L. racemosa (Cuadro 2).
Igualmente, la relación entre la mortalidad y la herbivoría, cuando se consideraron todas las especies de manglar, fue significativa (F4.211=19.66, p<0.01). Al comparar las especies se encontraron diferencias significativas entre A. germinans y L. racemosa, y entre A. germinans y R. mangle, pero no entre R. mangle y L. racemosa (Cuadro 3).
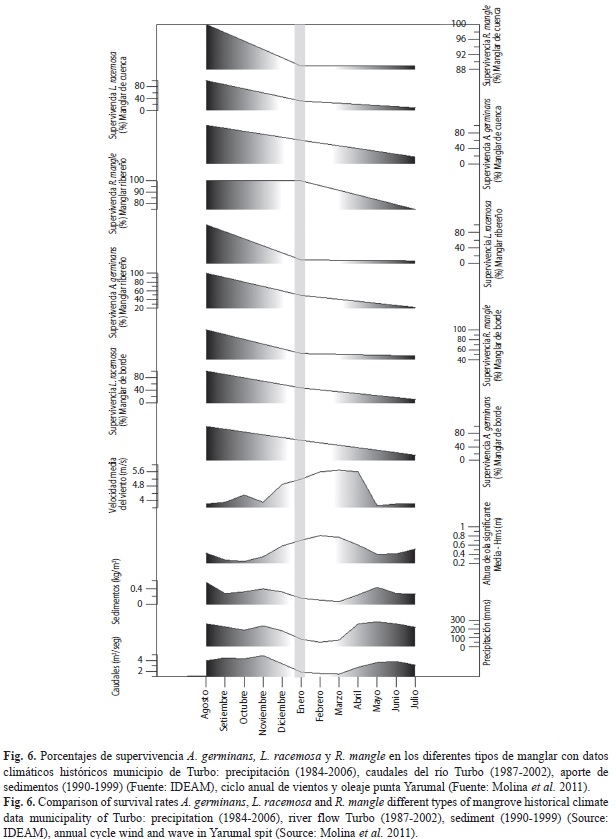
La figura 6 presenta relaciones cualitativas entre la precipitación máxima mensual, el caudal del río Turbo y la carga de sedimentos, con la supervivencia, mortalidad y reclutamiento de las especies de los manglares. De mayo a noviembre (período de lluvias) se presentó un descenso en la supervivencia de todas las especies, que coincide con la época de máximos valores promedio de precipitación, caudales de los ríos y acumulación de sedimentos. Durante estos meses el oleaje presentó las menores alturas promedio (0.25m), y las velocidades más bajas del viento de dirección sureste (3.80-4.20m/s). Igualmente, durante este periodo, el reclutamiento de A. germinans fue un poco superior a la mortalidad, mientras que en L. racemosa y R. mangle ocurrió lo contrario.
Entre diciembre y abril se presentó un descenso en los valores de precipitación, caudales y la acumulación de sedimentos. Los valores de velocidad del viento aumentaron casi a 6m/s y cambiaron de dirección, de sureste a noreste, y el oleaje presentó los valores más altos (0.58-0.81Hsm-metros). Durante estos meses la supervivencia siguió descendiendo de forma casi lineal para A. germinans, más pronunciada para L. racemosa y en menor proporción para R. mangle. Sin embargo, se registraron proporciones similares entre la mortalidad y el reclutamiento para las tres especies.
Discusión
Las distribuciones diamétricas y de alturas en forma de “J” invertida para los tres tipos de manglares, reflejan la extracción intensiva de madera proveniente de árboles de las mayores tallas, por parte de los pobladores locales (Sánchez et al. 1997, CORPOURABÁ 2003). Esta actividad ayuda a explicar también la mayor disponibilidad de luz en los manglares de borde y ribereños, que es donde se presenta la mayor extracción. Así mismo, se puede atribuir a dicha actividad, la disminución de la regeneración natural en respuesta a la reducción del número de individuos en edad reproductiva, principalmente de R. mangle y L. racemosa que son las especies más extraídas.
La dominancia de A. germinans en los tres censos, seguida por L. racemosa podría explicarse como una manifestación de la estrategia de alta producción de propágulos que tienen estas especies (Jiménez 1994) ya que ambas especies producen mayores cantidades de propágulos que R. mangle (Ellison & Farnsworth 1993). Así mismo, la dominancia de A. germinans en la regeneración podría considerarse como una expresión de su abundancia en el dosel. La tala de árboles de L. racemosa y R. mangle reduce las posibilidades de encontrar un alto número de individuos en edad reproductiva, y por tanto, de plántulas de estas especies. Además, la mayor disponibilidad de luz asociada a los claros originados por esta tala privilegia el establecimiento de unas especies (Rabinowitz 1978), como se observó en las plántulas de A. germinans.
La co-dominancia de R. mangle en los manglares ribereños y de borde, refleja la importancia de los gradientes edáficos y de marea, en la mortalidad y el reclutamiento de esta especie (Patterson & Mendelssohn 1991), que es la mejor adaptada a suelos con periodos de inundación prolongados y baja disponibilidad de oxígeno (Ellison & Farnsworth 1993). Es probablemente por esto también que R. mangle presentó la mayor supervivencia en todos los censos, en los tres tipos de manglares. El menor porcentaje de supervivencia lo presentó L. racemosa, tal como se registró en la Ciénaga Grande de Santa Marta (Elster 1998) y en el delta del Río Ranchería (Lema & Polania 2005) en Colombia, y en Belice (Ellison & Farnsworth 1993, McKee 1995). Rabinowitz (1978) determinó que las plántulas de R. mangle pueden persistir por más de un año en el sotobosque, pero las cohortes de A. germinans y L. racemosa, rara vez sobreviven más de seis meses bajo adultos congéneres o hetero-específicos. Sin embargo, las altas tasas de reclutamiento de A. germinans posibilitan su dominancia en el proceso de regeneración, tal como se ha registrado en otras zonas (Elster et al. 1999).
La disminución en los porcentajes de supervivencia podría explicarse en parte por la ocurrencia del “mar de leva”, registrado en enero de 2009 durante el segundo censo, en el cual se incrementó el nivel del mar y se inundaron áreas extensas a lo largo de toda la costa (Lacambra et al. 2003). Martínez (2010) estimó en 1.14m el valor promedio de la serie de cota de inundación en los eventos de mar de leva en la costa Atlántica Colombiana. Aunque los árboles de los manglares presentan adaptaciones morfológicas, anatómicas y fisiológicas que les permiten sobrevivir a la inundación (Tomlinson 1999), también se pueden ahogar las plántulas y juveniles de éstos, cuando quedan cubiertos por niveles altos de agua durante periodos prolongados (Reyes de la Cruz et al. 2002). Además de la inundación, durante el mar de leva se incrementa la velocidad de los vientos (hasta 35 nudos-60km/h) y por lo tanto también aumenta la energía del oleaje y la altura de las olas (2.50-4.50m) (CIOH 2010), lo que causa una remoción del suelo, y afecta el desarrollo de las plántulas que no hayan desarrollado las raíces suficientes para mantenerse en pie.
Los grupos observados en el AR relacionados con las variables ambientales no se asocian estrictamente con los tipos de manglares, lo cual evidencia que las variables ambientales medidas, operan a una escala más pequeña que las variables fisiográficas que definen los tipos de manglares.
Una de ellas es la sedimentación, relacionada a su vez con la distancia al río Turbo, principal fuente de sedimentos, y que afectó especialmente la supervivencia de A. germinans, en las sub-parcelas más cercanas a éste y donde se registraron los valores más altos de sedimentación (hasta 12cm en un año), lo cual supera el límite máximo de 10cm que podrían soportar las plántulas (Ellison 1999). A pesar del método cualitativo (Krauss et al. 2003) utilizado, se pudo establecer que en algunas sub-parcelas, alejadas de la influencia del río y cercanas a la línea de costa, la sedimentación fue muy escasa o incluso se registró erosión, al desaparecer el horizonte de feldespato, lo cual pudo causar la muerte de muchas plántulas e impedir el establecimiento de nuevas. No obstante, la alta sedimentación aunque disminuyó la supervivencia, dejó un sustrato disponible para el reclutamiento de nuevas plántulas de A. germinans, como lo indican la cercanía y dirección de los vectores de sedimentación y reclutamiento de este especie en la grafica de AR y como se ha registrado en otros manglares (Terrados et al. 1997, Ellison 1999, Thampanya et al. 2002a). La mayor supervivencia de R. mangle (Fig. 5) se asoció con una mayor disponibilidad de luz (FSD), resultados que concuerdan con la dependencia de la luz y mejor desempeño identificado en plántulas de R. mangle en grandes claros del dosel (Ellison & Farnsworth 1993). Sin embargo, R. mangle también presentó alta supervivencia en parcelas con alta sedimentación, lo cual se relaciona con la mayor longitud de sus propágulos, que le confiere una ventaja competitiva adicional con respecto a las otras dos especies de manglar (Thampanya et al. 2002b).
El tercer grupo identificado en el AR de la regeneración agrupa sub-parcelas ubicadas en manglares ribereños y de borde, y asociadas a la distancia al mar y a las lagunas costeras. Las sub-parcelas más cercanas a estos cuerpos de agua pasan mayor tiempo inundadas y pueden presentar menos oxígeno disuelto; también por la influencia de los procesos fluvio-marinos sobre las características de los suelos, principalmente el aporte de materia orgánica, nutrientes y sedimentos a los manglares.
La herbivoría presentó vectores de tamaños representativos en el AR (Coeficientes de 0.24, 0.25, 0.34 para las tres especies en el eje uno), asociada al reclutamiento y supervivencia de L. racemosa, como en otros estudios (Reyes de la Cruz et al. 2002), al igual que al reclutamiento de R. mangle. La asociación con la mortalidad denso-dependiente de las plántulas, indica su importancia como mecanismo regulador de las poblaciones. No obstante, el efecto de la herbivoría fue diferencial sobre las especies. Aunque el mayor porcentaje lo registró R. mangle (97.40%), en el segundo censo (enero 2009), en tres de las parcelas, las plántulas de A. germinans recién establecidas presentaron un ataque masivo de la oruga de Junonia evarete Cramer, tal como se registró en la Ciénaga Grande de Santa Marta (Colombia) (Elster et al. 1999). Farnsworth & Ellison (1991) atribuyen las diferencias en la herbivoría sobre las especies de manglar a la variación en la textura, palatabilidad, composición química o el contenido de nutrientes de las hojas. Es por ello, que al analizar las especies de manera independiente se encontró que la media de la herbivoría de A. germinans es diferente a la de las otras especies, lo cual puede estar afectando la competencia con L. racemosa y R. mangle por recursos y espacio. Probablemente, esta es la razón por la cual en el tercer censo no se registró denso-dependencia estadísticamente significativa entre las especies y las categorías de mortalidad, pero si un aumento en la mortalidad neta, lo que podría indicar que la disminución en la densidad de las plántulas, disminuye la competencia entre ellas.
Las tendencias de los datos históricos señalan que el aumento del caudal y la carga de sedimentos del río Turbo en la época de mayor precipitación, pueden afectar la supervivencia de la regeneración natural de todas las especies y en todos los tipos de manglar, tal como se registró en la Ciénaga Grande de Santa Marta (Elster et al. 1999).
La mayor supervivencia de R. mangle se relaciona con su adaptación a suelos más inestables, característicos de la zona intermareal baja y su mayor resistencia a la inundación (precipitación, caudales) y al aporte de sedimentos, lo cual marca la diferencia con A. germinans y L. racemosa adaptadas a las condiciones zonas intermareales más altas (McKee 1995, Reyes de la Cruz et al. 2002). En la época lluviosa el reclutamiento de R. mangle y L. racemosa fue menor que la mortalidad, probablemente asociado al exceso de sedimentos que sepulta los propágulos nuevos y no permite que estos se levanten para enraizar, disminuyendo así, la cantidad de reclutas.
Entre diciembre y abril, cuando desciende la precipitación, los caudales y la sedimentación, se favorece el balance entre el reclutamiento y la mortalidad para todas las especies en todos los tres tipos de manglar, excepto para L. racemosa en los manglares de borde, donde se presentan inundaciones con mayor nivel y mayor reducción en los suelos (McKee 1995) que también afectan la supervivencia de esta especie.
Las observaciones y los análisis realizados en esta investigación mostraron que: A. germinans dominó la regeneración natural, en la zona de estudio, favorecida por el aporte de sedimentos del río Turbo, un sustrato más firme y la formación de áreas nuevas para colonizar. Rhizophora mangle registró la mayor supervivencia en todos los tipos de manglar, por su tolerancia a altos niveles inundación, durante los periodos con mayor precipitación, caudales y aporte de sedimentos de los ríos, así como a la mayor disponibilidad de luz generada por la formación de claros creados por la intervención antrópica.
En cuanto a la mortalidad relacionada con la remoción de plántulas, L. racemosa presentó la mayor mortalidad, y se puede atribuir al mar de leva, el aumento en la inundación y la carga de sedimentos en la época más lluviosa, así como a la herbivoría. Las condiciones de menor inundación y sedimentación en la época de menor precipitación favorecieron el balance entre el reclutamiento y la mortalidad para todas las especies. La mortalidad mostró un patrón generalizado de denso-dependencia.
La luz y la sedimentación desempeñan un papel importante en el proceso de regeneración de los manglares pero se requiere una combinación de variables ambientales, biológicas y climáticas para que las plántulas de A. germinans, L. racemosa y R. mangle alcancen el estado adulto. Se recomienda para estudios posteriores tener en cuenta variables no medidas en ésta investigación, como el potencial redox, así como los niveles de inundación fluvial.
Agradecimientos
Los autores agradecen a la Dirección de investigaciones (DIME) y al Instituto de Investigaciones de la Facultad de Ciencias Agrarias, de la Universidad Nacional de Colombia, Sede Medellín, por la financiación y administración del proyecto. A COLCIENCIAS por el apoyo a la primera autora como joven investigadora. A Daniel Felipe Herrera y Luis Villegas por su colaboración en el trabajo de campo. A Luis Gabriel Molina y Andrés Osorio por compartir los datos de oleaje, caudales y vientos.
Referencias
Asquith, N.M. 2002. La dinámica del bosque y la diversidad arbórea. Parthenon, New Jersey,USA. [ Links ]
Benfield, S.L., H.M. Guzman & J.M. Mair. 2005. Temporal Mangrove Dynamics in Relation to Coastal Development in Pacific Panama. J. Environ. Manage. 76: 263-276. [ Links ]
Blanco, J.F., E.A. Estrada, L.F. Ortiz & L.E. Urrego. 2012. Ecosystem-Wide Impacts of Deforestation in Mangroves: The Urabá Gulf (Colombian Caribbean) case study. Ecology (in press). [ Links ]
CIOH (Centro de investigaciones oceanográficas e hidrográficas del Caribe). 2010. Mar de leva en el litoral caribe colombiano, Cartagena de Indias, Bolívar, Colombia. (Consultado: Junio 2011, http://www.cioh.org.co/index.php/noticias-mainmenu2677/18oceanografoperacional/1443-mar-de-leva-en-el-litoral-caribe#josc2314). [ Links ]
Correa, I.D. & I. Vernette. 2004. Introducción al problema de la erosión litoral en Urabá (sector Arboletes - Turbo), Costa Caribe colombiana. Bol. Invemar 33: 5-26. [ Links ]
CORPOURABÁ, Corporación para el Desarrollo Sostenible del Urabá. 2003. Caracterización y zonificación de los manglares del Golfo de Urabá. Apartadó, Antioquia, Colombia. [ Links ]
Curtis, J.T. & R.P. McIntosh. 1951. An Upland Forest Continuum in the Prairie-Forest Border Region of Wisconsin. Ecology 32: 476-496. [ Links ]
Dahdouh-Guebas, F., M. Verneirt, J.F. Tack, D. Van Speybroecky & N. Koedam. 1998. Propagule Predators in Kenyan Mangroves and their Possible Effect on Regeneration. Mar. Freshwater Res. 49: 345-350. [ Links ]
Delgado, P., P.F. Hensel, J.A. Jiménez & J.W. Day. 2001. The Importance of Propagule Establishment and Physical Factors in Mangrove Distributional Patterns in a Costa Rican Estuary. Aquat. Bot. 71: 157-178. [ Links ]
Ellison, A.M. & E.J. Farnsworth. 1993. Seedling Survivorship, Growth, and Response to Disturbance in Belizean Mangal. Am. J. Bot. 80: 1137-1145. [ Links ]
Ellison, A.M. & E.J. Farnsworth. 1996. Spatial and Temporal Variability in Growth of Rhizophora mangle Saplings on Coral Cays: Links with Variation in Insolation, Herbivory, and Local Sedimentation Rate. J. Ecol. 84: 717-731. [ Links ]
Ellison, J.C. 1999. Impacts of Sediment Burial on Mangroves. Mar. Pollut. Bull. 37: 420-426. [ Links ]
Elster, C. 1998. Posibilidades de regeneración del manglar en la zona de la Ciénaga Grande de Santa Marta. Deutsche Gesellshaftfür Technische Zusammenarbeit (GTZ) Gmbh, Eschborn. [ Links ]
Elster, C., L. Perdomo, J. Polania & M.L. Schnetter. 1999. Control of Avicennia germinans Recruitment and Survival by Junonia Evarete Larvae in a Disturbed Mangrove Forest in Colombia. J. Trop. Ecol. 15: 791-805. [ Links ]
Estrada, U.E. & P.S. Gil. 2005. Análisis de las condiciones morfológicas actuales como indicadoras de la evolución de la flecha litoral, en la actual desembocadura del Río Turbo, Antioquia. Tesis de pregrado, Universidad de Antioquia-Seccional Urabá, Turbo, Colombia. [ Links ]
Farnsworth, E.J. & A.M. Ellison. 1991. Patterns of Herbivory in Belizean Mangrove Swamps. Biotropica 23: 555-567. [ Links ]
Farnsworth, E.J. & A.M. Ellison. 1996. SunShade Adaptability of the Red Mangrove, Rhizophora mangle (Rhizophoraceae): Changes through Ontogeny at several Levels of Biological Organization. Am. J. Bot. 83: 1131-1143. [ Links ]
Farnsworth, E.J. & A.M. Ellison. 1997. Global Patterns of Pre-Dispersal Propagule Predation in Mangrove Forests. Biotropica 29: 318-330. [ Links ]
Fisher, R.A. 1935. The design of experiments. Macmillan, Oxford, England. [ Links ]
IDEAM, Instituto de Hidrología, Meteorología y Estudios Ambientales. 2007. Datos sistema de información nacional ambiental. Turbo, Antioquia, Colombia. [ Links ]
INVEMAR, Instituto de Investigaciones Marinas y Costeras “José Benito De Andréis” & CORPOURABÁ, Corporación para el Desarrollo Sostenible del Urabá. 2003. Diagnóstico, zonificación y definición de la estructura administrativa de manejo de la Unidad Ambiental Costera del Darién, Caribe colombiano fase I. Caracterización y diagnóstico. Informe técnico preliminar. Santa Marta, Colombia. [ Links ]
Janzen, D.H. 1985. Mangroves: Where’s the Understory? J. Trop. Ecol. 1: 89. [ Links ]
Jiménez, J.A., 1994. Los manglares del pacífico centroamericano. Editorial Fundación UNA, Heredia, Costa Rica. [ Links ]
Kathiresan, K. & P. Moorthy. 1993. Influence of Different Irradiance on Growth and Photosynthetic Characteristics in Seedlings of Rhizophora Species. Photosynthetica 29: 143-146. [ Links ]
Kathiresan, K. & B.L. Bingham. 2001. Biology of mangroves and mangrove Ecosystems. Adv. Mar. Biol. 81-251. [ Links ]
Keep, F. 2000. Monografía de Turbo. Municipio de Turbo. Turbo, Antioquia, Colombia. [ Links ]
Krauss, K.W., J.A. Allen & D.R. Cahoon. 2003. Differential rates of vertical accretion and elevation change among aerial root types in Micronesian Mangrove forest. Estuar. Coast. Shelf. S. 56: 251-259. [ Links ]
Krauss, K.W., C.E. Lovelock, K.L. McKee, L. López-Hoffman, S.M.L. Ewe & W.P. Sousa. 2008. Environmental Drivers in Mangrove Establishment and Early Development: A Review. Aquat. Bot. 89: 105-127. [ Links ]
Lacambra, C.L., C. Lozano & M. Fontalvo. 2003. Amenazas naturales y antrópicas en las zonas costeras colombianas. INVEMAR, Santa Marta, Magdalena, Colombia. [ Links ]
Lema, L.F., J. Polanía & L.E. Urrego. 2003. Dispersión y establecimiento de las especies de mangle del río Ranchería en el período de máxima fructificación. Rev. Acad. Colomb. Cienc. 27: 93-103. [ Links ]
Lema, L.F. & J. Polanía. 2005. Regeneración natural y producción del manglar del delta del río ranchería, Caribe colombiano. Actual Biol. 27: 25-33. [ Links ]
López-Hoffman, L., N.P.R. Anten, M. Martínez-Ramos & D.D. Ackerly. 2006. Salinity and light interactively affect Neotropical mangrove seedlings at the leaf and whole plant levels. Oecologia 150: 545-556. [ Links ]
Lozano, L.H. 1998. Caracterización y diagnóstico integral de la zona costera que comprende la bahía Colombia desde el río Turbo hasta Matuntugo. Escuela Naval Almirante Padilla, Cartagena de Indias, Bolívar, Colombia. [ Links ]
Lugo, A.E. & S.C. Snedaker. 1974. The Ecology of Mangroves. Annu. Rev. Ecol. Syst. 5: 39-64. [ Links ]
Martínez, J.C. 2010. Propuesta metodológica para la estimación de la cota de inundación en la zona costera del Caribe Colombiano. Tesis de Maestría, Universidad Nacional de Colombia, Medellín, Colombia. [ Links ]
McGuinness, K.A. 1997. Dispersal, Establishment and Survival of Ceriops Tagal Propagules in a North Australian Mangrove Forest. Oecologia 109: 80-87. [ Links ]
McKee, K.L. 1995. Seedling Recruitment Patterns in a Belizean Mangrove Forest: Effects of Establishment Ability and Physico-Chemical Factors. Oecologia 101: 448-460. [ Links ]
Medina, E., E. Cuevas, M. Popp & A.E. Lugo. 1990. Soil Salinity, Sun Exposure, y Growth of Acrostichum aureum, the Mangrove Fern. Bot. Gaz. 151: 41-49. [ Links ]
Molina, L.E., V.F. Pérez, J.O. Martínez, J.V. Franco, L. Marín, J.L. González & J.H. Carvajal. 1998. Geomorfología y aspectos erosivos del litoral Caribe colombiano. Publ. Geol. Esp. INGEOMINAS 21: 1-73. [ Links ]
Molina, L.G. 2011. A.F. Osorio, L.J. Otero. Capacidad de transporte longitudinal de sedimentos a escala intra-anual en la zona costera de Punta Yarumal - delta del río turbo, Golfo de Urabá, Antioquia–a partir de la simulación de un clima marítimo. XXV Congreso Latinoamericano de Hidráulica. 2012. San José, Costa Rica. [ Links ]
Patterson, C.S. & I.A. Mendelssohn. 1991. A Comparison of Physicochemical Variables Across Plant Zones in a mangal/salt Marsh Community in Louisiana. Wetlands 11: 139-161. [ Links ]
Rabinowitz, D. 1978. Mortality and Initial Propagule Size in Mangrove Seedlings in Panamá. J. Ecol. 66: 45-51. [ Links ]
Reyes de la Cruz, A., G. López Ocaña & H. Hernández Trejo. 2002. Evaluación preliminar de los efectos de la Inundación y la herbivoría sobre plántulas de mangle. Universidad y Ciencia 18: 136-139. [ Links ]
Sánchez, P.H., R. Álvarez, F. Pinto, A.S. Sánchez, J.C. Pino, I. García & M.T. Acosta. 1997. Diagnóstico y zonificación preliminar de los manglares del Caribe de Colombia. Bogotá, D.C., Colombia. [ Links ]
SHOM, Service Hydrographique et Océanographique de la Marine. 1982. Table des marées des grands ports du monde. Paris, France. [ Links ]
Ter Braak, C. & P. Smilauer. 1998. CANOCO Reference Manual and User’s Guide to Canoco for Windows: Software for Canonical Community Ordination. 4: 352. [ Links ]
Terrados, J., U. Thampanya, N. Srichai, P. Kheowvongsri, O. Geertz-Hansen, S. Boromthanarath, N. Panapitukkuly & C.M. Duarte. 1997. The Effect of Increased Sediment Accretion on the Growth of Rhizophora apiculata Seedlings. Estuar. Coast. Shelf Sci. 45: 697-701. [ Links ]
Thampanya, U., J.E. Vermaaty & J. Terrados. 2002a. The Effect of Increasing Sediment Accretion on the Seedlings of Three Common Thai Mangrove Species. Aquat. Bot. 74: 315-325. [ Links ]
Thampanya, U., J.E. Vermaaty & C.M. Duarte. 2002b. Colonization Success of Common Thai Mangrove Species as a Function of Shelter from Water Movement. Mar. Ecol. Prog. Ser. 237: 111-120. [ Links ]
Tomlinson, P.B. 1999. The Botany of Mangroves. Cambridge University, Cambridge, United Kingdom. [ Links ]
Van Wijnen, H.J. & J.P. Bakker. 2001. Long-Term Surface Elevation Change in Salt Marshes: A Prediction of Marsh Response to Future Sea-Level Rise. Estuar. Coast. Shelf. S. 52: 381-390. [ Links ]
Zhang, Y., W. Wang, Q. Wu, B. Fang & P. Lin. 2006. The growth of Kandelia candel seedlings in mangrove habitats of the Zhangjiang estuary in Fujian, China. Act. Ecol. Sinica. 26: 1648-1656. [ Links ]
Benfield, S.L., H.M. Guzman & J.M. Mair. 2005. Temporal Mangrove Dynamics in Relation to Coastal Development in Pacific Panama. J. Environ. Manage. 76: 263-276. [ Links ]
Blanco, J.F., E.A. Estrada, L.F. Ortiz & L.E. Urrego. 2012. Ecosystem-Wide Impacts of Deforestation in Mangroves: The Urabá Gulf (Colombian Caribbean) case study. Ecology (in press). [ Links ]
CIOH (Centro de investigaciones oceanográficas e hidrográficas del Caribe). 2010. Mar de leva en el litoral caribe colombiano, Cartagena de Indias, Bolívar, Colombia. (Consultado: Junio 2011, http://www.cioh.org.co/index.php/noticias-mainmenu2677/18oceanografoperacional/1443-mar-de-leva-en-el-litoral-caribe#josc2314). [ Links ]
Correa, I.D. & I. Vernette. 2004. Introducción al problema de la erosión litoral en Urabá (sector Arboletes - Turbo), Costa Caribe colombiana. Bol. Invemar 33: 5-26. [ Links ]
CORPOURABÁ, Corporación para el Desarrollo Sostenible del Urabá. 2003. Caracterización y zonificación de los manglares del Golfo de Urabá. Apartadó, Antioquia, Colombia. [ Links ]
Curtis, J.T. & R.P. McIntosh. 1951. An Upland Forest Continuum in the Prairie-Forest Border Region of Wisconsin. Ecology 32: 476-496. [ Links ]
Dahdouh-Guebas, F., M. Verneirt, J.F. Tack, D. Van Speybroecky & N. Koedam. 1998. Propagule Predators in Kenyan Mangroves and their Possible Effect on Regeneration. Mar. Freshwater Res. 49: 345-350. [ Links ]
Delgado, P., P.F. Hensel, J.A. Jiménez & J.W. Day. 2001. The Importance of Propagule Establishment and Physical Factors in Mangrove Distributional Patterns in a Costa Rican Estuary. Aquat. Bot. 71: 157-178. [ Links ]
Ellison, A.M. & E.J. Farnsworth. 1993. Seedling Survivorship, Growth, and Response to Disturbance in Belizean Mangal. Am. J. Bot. 80: 1137-1145. [ Links ]
Ellison, A.M. & E.J. Farnsworth. 1996. Spatial and Temporal Variability in Growth of Rhizophora mangle Saplings on Coral Cays: Links with Variation in Insolation, Herbivory, and Local Sedimentation Rate. J. Ecol. 84: 717-731. [ Links ]
Ellison, J.C. 1999. Impacts of Sediment Burial on Mangroves. Mar. Pollut. Bull. 37: 420-426. [ Links ]
Elster, C. 1998. Posibilidades de regeneración del manglar en la zona de la Ciénaga Grande de Santa Marta. Deutsche Gesellshaftfür Technische Zusammenarbeit (GTZ) Gmbh, Eschborn. [ Links ]
Elster, C., L. Perdomo, J. Polania & M.L. Schnetter. 1999. Control of Avicennia germinans Recruitment and Survival by Junonia Evarete Larvae in a Disturbed Mangrove Forest in Colombia. J. Trop. Ecol. 15: 791-805. [ Links ]
Estrada, U.E. & P.S. Gil. 2005. Análisis de las condiciones morfológicas actuales como indicadoras de la evolución de la flecha litoral, en la actual desembocadura del Río Turbo, Antioquia. Tesis de pregrado, Universidad de Antioquia-Seccional Urabá, Turbo, Colombia. [ Links ]
Farnsworth, E.J. & A.M. Ellison. 1991. Patterns of Herbivory in Belizean Mangrove Swamps. Biotropica 23: 555-567. [ Links ]
Farnsworth, E.J. & A.M. Ellison. 1996. SunShade Adaptability of the Red Mangrove, Rhizophora mangle (Rhizophoraceae): Changes through Ontogeny at several Levels of Biological Organization. Am. J. Bot. 83: 1131-1143. [ Links ]
Farnsworth, E.J. & A.M. Ellison. 1997. Global Patterns of Pre-Dispersal Propagule Predation in Mangrove Forests. Biotropica 29: 318-330. [ Links ]
Fisher, R.A. 1935. The design of experiments. Macmillan, Oxford, England. [ Links ]
IDEAM, Instituto de Hidrología, Meteorología y Estudios Ambientales. 2007. Datos sistema de información nacional ambiental. Turbo, Antioquia, Colombia. [ Links ]
INVEMAR, Instituto de Investigaciones Marinas y Costeras “José Benito De Andréis” & CORPOURABÁ, Corporación para el Desarrollo Sostenible del Urabá. 2003. Diagnóstico, zonificación y definición de la estructura administrativa de manejo de la Unidad Ambiental Costera del Darién, Caribe colombiano fase I. Caracterización y diagnóstico. Informe técnico preliminar. Santa Marta, Colombia. [ Links ]
Janzen, D.H. 1985. Mangroves: Where’s the Understory? J. Trop. Ecol. 1: 89. [ Links ]
Jiménez, J.A., 1994. Los manglares del pacífico centroamericano. Editorial Fundación UNA, Heredia, Costa Rica. [ Links ]
Kathiresan, K. & P. Moorthy. 1993. Influence of Different Irradiance on Growth and Photosynthetic Characteristics in Seedlings of Rhizophora Species. Photosynthetica 29: 143-146. [ Links ]
Kathiresan, K. & B.L. Bingham. 2001. Biology of mangroves and mangrove Ecosystems. Adv. Mar. Biol. 81-251. [ Links ]
Keep, F. 2000. Monografía de Turbo. Municipio de Turbo. Turbo, Antioquia, Colombia. [ Links ]
Krauss, K.W., J.A. Allen & D.R. Cahoon. 2003. Differential rates of vertical accretion and elevation change among aerial root types in Micronesian Mangrove forest. Estuar. Coast. Shelf. S. 56: 251-259. [ Links ]
Krauss, K.W., C.E. Lovelock, K.L. McKee, L. López-Hoffman, S.M.L. Ewe & W.P. Sousa. 2008. Environmental Drivers in Mangrove Establishment and Early Development: A Review. Aquat. Bot. 89: 105-127. [ Links ]
Lacambra, C.L., C. Lozano & M. Fontalvo. 2003. Amenazas naturales y antrópicas en las zonas costeras colombianas. INVEMAR, Santa Marta, Magdalena, Colombia. [ Links ]
Lema, L.F., J. Polanía & L.E. Urrego. 2003. Dispersión y establecimiento de las especies de mangle del río Ranchería en el período de máxima fructificación. Rev. Acad. Colomb. Cienc. 27: 93-103. [ Links ]
Lema, L.F. & J. Polanía. 2005. Regeneración natural y producción del manglar del delta del río ranchería, Caribe colombiano. Actual Biol. 27: 25-33. [ Links ]
López-Hoffman, L., N.P.R. Anten, M. Martínez-Ramos & D.D. Ackerly. 2006. Salinity and light interactively affect Neotropical mangrove seedlings at the leaf and whole plant levels. Oecologia 150: 545-556. [ Links ]
Lozano, L.H. 1998. Caracterización y diagnóstico integral de la zona costera que comprende la bahía Colombia desde el río Turbo hasta Matuntugo. Escuela Naval Almirante Padilla, Cartagena de Indias, Bolívar, Colombia. [ Links ]
Lugo, A.E. & S.C. Snedaker. 1974. The Ecology of Mangroves. Annu. Rev. Ecol. Syst. 5: 39-64. [ Links ]
Martínez, J.C. 2010. Propuesta metodológica para la estimación de la cota de inundación en la zona costera del Caribe Colombiano. Tesis de Maestría, Universidad Nacional de Colombia, Medellín, Colombia. [ Links ]
McGuinness, K.A. 1997. Dispersal, Establishment and Survival of Ceriops Tagal Propagules in a North Australian Mangrove Forest. Oecologia 109: 80-87. [ Links ]
McKee, K.L. 1995. Seedling Recruitment Patterns in a Belizean Mangrove Forest: Effects of Establishment Ability and Physico-Chemical Factors. Oecologia 101: 448-460. [ Links ]
Medina, E., E. Cuevas, M. Popp & A.E. Lugo. 1990. Soil Salinity, Sun Exposure, y Growth of Acrostichum aureum, the Mangrove Fern. Bot. Gaz. 151: 41-49. [ Links ]
Molina, L.E., V.F. Pérez, J.O. Martínez, J.V. Franco, L. Marín, J.L. González & J.H. Carvajal. 1998. Geomorfología y aspectos erosivos del litoral Caribe colombiano. Publ. Geol. Esp. INGEOMINAS 21: 1-73. [ Links ]
Molina, L.G. 2011. A.F. Osorio, L.J. Otero. Capacidad de transporte longitudinal de sedimentos a escala intra-anual en la zona costera de Punta Yarumal - delta del río turbo, Golfo de Urabá, Antioquia–a partir de la simulación de un clima marítimo. XXV Congreso Latinoamericano de Hidráulica. 2012. San José, Costa Rica. [ Links ]
Patterson, C.S. & I.A. Mendelssohn. 1991. A Comparison of Physicochemical Variables Across Plant Zones in a mangal/salt Marsh Community in Louisiana. Wetlands 11: 139-161. [ Links ]
Rabinowitz, D. 1978. Mortality and Initial Propagule Size in Mangrove Seedlings in Panamá. J. Ecol. 66: 45-51. [ Links ]
Reyes de la Cruz, A., G. López Ocaña & H. Hernández Trejo. 2002. Evaluación preliminar de los efectos de la Inundación y la herbivoría sobre plántulas de mangle. Universidad y Ciencia 18: 136-139. [ Links ]
Sánchez, P.H., R. Álvarez, F. Pinto, A.S. Sánchez, J.C. Pino, I. García & M.T. Acosta. 1997. Diagnóstico y zonificación preliminar de los manglares del Caribe de Colombia. Bogotá, D.C., Colombia. [ Links ]
SHOM, Service Hydrographique et Océanographique de la Marine. 1982. Table des marées des grands ports du monde. Paris, France. [ Links ]
Ter Braak, C. & P. Smilauer. 1998. CANOCO Reference Manual and User’s Guide to Canoco for Windows: Software for Canonical Community Ordination. 4: 352. [ Links ]
Terrados, J., U. Thampanya, N. Srichai, P. Kheowvongsri, O. Geertz-Hansen, S. Boromthanarath, N. Panapitukkuly & C.M. Duarte. 1997. The Effect of Increased Sediment Accretion on the Growth of Rhizophora apiculata Seedlings. Estuar. Coast. Shelf Sci. 45: 697-701. [ Links ]
Thampanya, U., J.E. Vermaaty & J. Terrados. 2002a. The Effect of Increasing Sediment Accretion on the Seedlings of Three Common Thai Mangrove Species. Aquat. Bot. 74: 315-325. [ Links ]
Thampanya, U., J.E. Vermaaty & C.M. Duarte. 2002b. Colonization Success of Common Thai Mangrove Species as a Function of Shelter from Water Movement. Mar. Ecol. Prog. Ser. 237: 111-120. [ Links ]
Tomlinson, P.B. 1999. The Botany of Mangroves. Cambridge University, Cambridge, United Kingdom. [ Links ]
Van Wijnen, H.J. & J.P. Bakker. 2001. Long-Term Surface Elevation Change in Salt Marshes: A Prediction of Marsh Response to Future Sea-Level Rise. Estuar. Coast. Shelf. S. 52: 381-390. [ Links ]
Zhang, Y., W. Wang, Q. Wu, B. Fang & P. Lin. 2006. The growth of Kandelia candel seedlings in mangrove habitats of the Zhangjiang estuary in Fujian, China. Act. Ecol. Sinica. 26: 1648-1656. [ Links ]
*Correspondencia a:
Róssalyn Hoyos G. Maestría en Bosques y Conservación Ambiental, Universidad Nacional de Colombia, Medellín, Colombia, Calle 59 A No. 63-020, Autopista Norte; rossalyn@gmail.com
Ligia Estela Urrego G. Departamento de Ciencias Forestales, Universidad Nacional de Colombia, Medellín, Colombia, Calle 59 A No. 63-020, Autopista Norte; leurrego@unal.edu.co
Álvaro Lema T.. Departamento de Ciencias Forestales, Universidad Nacional de Colombia, Medellín, Colombia, Calle 59 A No. 63-020, Autopista Norte; ajlema@unal.edu.co
1. Maestría en Bosques y Conservación Ambiental, Universidad Nacional de Colombia, Medellín, Colombia, Calle 59 A No. 63-020, Autopista Norte; rossalyn@gmail.com
2. Departamento de Ciencias Forestales, Universidad Nacional de Colombia, Medellín, Colombia, Calle 59 A No. 63-020, Autopista Norte; leurrego@unal.edu.co
3. Departamento de Ciencias Forestales, Universidad Nacional de Colombia, Medellín, Colombia, Calle 59 A No. 63-020, Autopista Norte; ajlema@unal.edu.co
Recibido 21-VI-2012. Corregido 30-X-2012. Aceptado 26-XI-2012.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}