










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.61 n.3 San José Sep. 2013
Demografía poblacional de Rhinella arenarum (Anura: Bufonidae) y Physalaemus biligonigerus (Anura: Leiuperidae) en agroecosistemas de la provincia de Córdoba, Argentina
Population demography in Rhinella arenarum (Anura: Bufonidae) and hysalaemus biligonigerus (Anura: Leiuperidae) in agroecosystems in the province of Córdoba, Argentina
Population demography in Rhinella arenarum (Anura: Bufonidae) and hysalaemus biligonigerus (Anura: Leiuperidae) in agroecosystems in the province of Córdoba, Argentina
Abstract
The advancing agricultural frontier has led to an important loss of natural habitats, with significant consequences for biodiversity. The demography for two species of anurans, Physalaemus biligonigerus and Rhinella arenarum, both associated with agricultural systems in the central region of the Córdoba Province, were analyzed and compared in this study. Four sites were sampled: three agroecosystems with different alteration degrees (C1, C2 and SM1) and a fourth site not cultivated (SM2). The sampling was conducted during two reproductive periods, from September 2008 to April 2009 and September 2009 to April 2010. Individuals were captured using live pitfall traps for the metamorphic, juveniles and adults; and visual encounter survey, for the capture of eggs and larvae. With the abundance data, the survival for each age class was estimated using the KNM method (Kiritani Nakasuki Manly). With survival rates and fertility population, Leslie matrices were elaborated to obtain a quantitative projection of the population size. Altered environments showed lower eggs and larvae survival. Population pro-jections were favorable in the site SM2 and were less favorable and a tendency to extinction, in sites dominated by crops. This study showed that the agroecosystems of this region are possibly inhospitable environments for reproduction and survival of the species studied. The aquatic stages in the life cycle of both species would be the more affected, since water bodies deterioration is present or may occur in those areas. We can recognize species-specific effects of agricultural ecosystems; P. biligonigerus was the most affected species, possibly because of their life histories and habitat requirements. We suggested that environmental degradation caused by the cropland in the central region of Argentina would impact on the demographics of the anuran populations in the area.
Key words: population demography, Rhinella arenarum, Physalaemus biligonigerus, survival, fertility, agroecosystems.
Resumen
El crecimiento de la agricultura produce pérdida del hábitat natural, con consecuencias para la biodiversidad de los anfibios. Se analizó la demografía poblacional de dos especies de anfibios en agroecosistemas de la provincia de Córdoba, Rhinella arenarum y Physalaemus biligonigerus. Los muestreos se realizaron en tres agroecosistemas y en un cuarto sitio, no afectado por cultivos. Los individuos se capturaron utilizando trampas de caída viva y relevamientos visuales. Con los datos de abundancia se estimó la supervivencia para cada clase de edad y con las tasas de supervivencia y fertilidad poblacional, se construyeron matrices de Leslie para obtener una proyección cuantitativa del tamaño poblacional. Las proyecciones poblacionales fueron favorables para el sitio SM2 y desfavorables para los sitios cultivados. Los agroecosistemas son ambientes posiblemente inhóspitos para la reproducción y supervivencia de las especies estudiadas. Particularmente se ven afectados aquellos estadios acuáticos del ciclo de vida de estas especies, debido al deterioro de los cuerpos de agua en estas áreas. Se reconocieron efectos especie-específicos de los ecosistemas agrícolas, ya que P. biligonigerus resultó la especie más afectada. Se sugiere un deterioro ambiental producido por cultivos extensivos de soja, lo que impactaría la demografía de las poblaciones de anuros.
Palabras clave: demografía poblacional, Rhinella arenarum, Physalaemus biligonigeru, supervivencia, fertilidad, agroecosistemas.
The advancing agricultural frontier has led to an important loss of natural habitats, with significant consequences for biodiversity. The demography for two species of anurans, Physalaemus biligonigerus and Rhinella arenarum, both associated with agricultural systems in the central region of the Córdoba Province, were analyzed and compared in this study. Four sites were sampled: three agroecosystems with different alteration degrees (C1, C2 and SM1) and a fourth site not cultivated (SM2). The sampling was conducted during two reproductive periods, from September 2008 to April 2009 and September 2009 to April 2010. Individuals were captured using live pitfall traps for the metamorphic, juveniles and adults; and visual encounter survey, for the capture of eggs and larvae. With the abundance data, the survival for each age class was estimated using the KNM method (Kiritani Nakasuki Manly). With survival rates and fertility population, Leslie matrices were elaborated to obtain a quantitative projection of the population size. Altered environments showed lower eggs and larvae survival. Population pro-jections were favorable in the site SM2 and were less favorable and a tendency to extinction, in sites dominated by crops. This study showed that the agroecosystems of this region are possibly inhospitable environments for reproduction and survival of the species studied. The aquatic stages in the life cycle of both species would be the more affected, since water bodies deterioration is present or may occur in those areas. We can recognize species-specific effects of agricultural ecosystems; P. biligonigerus was the most affected species, possibly because of their life histories and habitat requirements. We suggested that environmental degradation caused by the cropland in the central region of Argentina would impact on the demographics of the anuran populations in the area.
Key words: population demography, Rhinella arenarum, Physalaemus biligonigerus, survival, fertility, agroecosystems.
Resumen
El crecimiento de la agricultura produce pérdida del hábitat natural, con consecuencias para la biodiversidad de los anfibios. Se analizó la demografía poblacional de dos especies de anfibios en agroecosistemas de la provincia de Córdoba, Rhinella arenarum y Physalaemus biligonigerus. Los muestreos se realizaron en tres agroecosistemas y en un cuarto sitio, no afectado por cultivos. Los individuos se capturaron utilizando trampas de caída viva y relevamientos visuales. Con los datos de abundancia se estimó la supervivencia para cada clase de edad y con las tasas de supervivencia y fertilidad poblacional, se construyeron matrices de Leslie para obtener una proyección cuantitativa del tamaño poblacional. Las proyecciones poblacionales fueron favorables para el sitio SM2 y desfavorables para los sitios cultivados. Los agroecosistemas son ambientes posiblemente inhóspitos para la reproducción y supervivencia de las especies estudiadas. Particularmente se ven afectados aquellos estadios acuáticos del ciclo de vida de estas especies, debido al deterioro de los cuerpos de agua en estas áreas. Se reconocieron efectos especie-específicos de los ecosistemas agrícolas, ya que P. biligonigerus resultó la especie más afectada. Se sugiere un deterioro ambiental producido por cultivos extensivos de soja, lo que impactaría la demografía de las poblaciones de anuros.
Palabras clave: demografía poblacional, Rhinella arenarum, Physalaemus biligonigeru, supervivencia, fertilidad, agroecosistemas.
La proporción de bosques transformados en tierras agrícolas está creciendo rápidamente en algunas áreas Neotropicales. Particularmente, la región central de Argentina ha sido afectada por el desarrollo agrícola (Pengue 2005, Rossi 2006), con una clara tendencia de reemplazo de la agricultura tradicional por una producción más especializada, principalmente en la obtención de soja (Glycine max L.). Este progreso de la agricultura ha convertido a la región en un importante productor, procesador y exportador de granos, aceite y harina de soja (Pengue 2005, Altieri & Pengue 2006). Sin embargo, esta actividad se ha convertido en la principal causa de reducción, fragmentación y deterioro del hábitat natural y, en consecuencia, es considerada un factor relevante en la declinación registrada en las poblaciones de anfibios en las últimas décadas (di Tada et al. 1996, Wake 1998, Davidson et al. 2002, Stuart et al. 2004, Beebee & Griffiths 2005, Collins & Crump 2009, Peltzer et al. 2011).
En este contexto, resulta valioso el conocimiento sobre la demografía de las poblaciones de anfibios que habitan los ecosistemas agrícolas (Heyer 1974, Wilbur 1980, Guerry & Hunter 2002), ya que al estimar parámetros poblacionales, se puede establecer el estado de conservación de las especies. De esta manera, es posible determinar si las poblaciones de anfibios que habitan agroecosistemas son afectadas por dicha actividad, o si simplemente manifiestan fluctuaciones naturales en sus tamaños poblacionales (Beebee 1996). Las áreas cultivadas, entre otras consecuencias, producen la alteración de los ambientes acuáticos o un deterioro en la calidad de los mismos, en particular en la duración del hidroperiodo, que puede afectar la supervivencia de huevos y larvas de anfibios (Carey & Bryant 1995, Taylor et al. 2005, Altig et al. 2007, Peltzer et al. 2008). Estos efectos tienden a modificar los rasgos de historias de vida como tamaño y condición corporal, supervivencia, fertilidad, entre otros (Márquez García et al. 2009, Bionda et al. 2012), que en última instancia podrían comprometer la permanencia de las poblaciones en el tiempo. La determinación de la fecundidad y la supervivencia constituyen parámetros básicos de una población, ya que proveen información acerca de las variaciones demográficas, como las fluctuaciones numéricas o la magnitud del crecimiento poblacional (Carey 1993, Shaffer 1994, Sinsch et al. 2001), que permitiría esclarecer de qué manera ambientes como los agroecosistemas pueden afectar a las poblaciones de anfibios. En este trabajo nos proponemos analizar la demografía poblacional de dos especies de anfibios asociadas a sistemas agrícolas de la región central de la provincia de Córdoba, Rhinella arenarum Hensel 1867, y Physalaemus biligonigerus Cope 1861.
Materiales y métodos
Los sitios de muestreo se encuentran en los alrededores de la localidad de Río Cuarto (33º07’55’’18’’ S - 64º21’08’’96’’ W), provincia de Córdoba, Argentina. La región se caracteriza por tener clima semiseco, con tendencia a semihúmedo. Las lluvias máximas ocurren durante el periodo octubre-marzo, con una media anual de 800-1 000mm (Bionda 2011, Gatica et al. 2012). La fisiografía del área de estudio corresponde a llanuras suavemente onduladas. La escasa pendiente favorece el estancamiento de las aguas, sumado al aporte subterráneo que junto con las precipitaciones, ayuda a mantener cursos de agua permanentes (Bridarolli & di Tada 1996).
La selección de las especies, R. arenarum y P. biligonigerus, se basó en su representatividad en las comunidades de anfibios de la región de estudio (Bionda et al. 2011a). Ambas especies de anuros presentan una amplia distribución en el Neotrópico, siendo encontradas en Argentina, Bolivia, Brasil, Uruguay y Paraguay (IUCN 2011), y están consideradas como especies No Amenazadas (Lavilla et al. 2000) o de Preocupación Menor (IUCN 2011). La recolección de los especímenes fue realizada en cuatro sitios, tres de ellos se encuentran en zonas de cultivos de soja: Sitio Cultivo 1 (C1, 33º05’54’’38’’ S - 64º26’04’’76’’ W), Sitio Cultivo 2 (C2, 33º05’54’’79’’ S - 64º25’46’’77’’ W) y Sitio Semimodificado 1 (SM1, 33º05’39’’29’’ S - 64º25’57’’72’’ W). Los tres sitios presentan distintos grados de alteración en relación a la distancia de los cultivos a los cuerpos de agua. En C1 y C2, los cultivos están a escasos metros de los cuerpos de agua (15m aproximadamente), mientras que en SM1 se encuentran más alejados (200m aproximadamente). En los tres sitios existe la práctica ganadera, aunque el sitio C1 presenta esta actividad con una mayor intensidad (presencia de ganado a diario y en cercanía o dentro de lagunas). El cuarto sitio de muestreo, llamado Sitio Semimodificado 2 (SM2, 33º06’41’’95’’ S - 64º18’14’’71’’ W), corresponde a una zona periurbana no afectada por cultivos, cubierta por pastizales y formaciones boscosas de árboles autóctonos e introducidos (Doffo 1989).
El muestreo en cada sitio fue realizado durante dos periodos reproductivos, entre septiembre 2008 y abril 2009 y septiembre 2009 hasta abril 2010. Los muestreos fueron sistemáticos, con dos o tres visitas semanales dependiendo de las precipitaciones. Los individuos fueron capturados utilizando trampas de caída viva (Corn 1994). Estas trampas fueron dispuestas en dos líneas, estas últimas a una distancia de 6 a 8m de los cuerpos de agua. Cada línea de trampas consistió de ocho recipientes de plástico de 22cm de diámetro y 40cm de profundidad, espaciadas a 2.5m. El fondo de cada trampa fue humedecido para prevenir la desecación. Cada individuo capturado fue removido para ser marcado mediante la amputación de falanges (Waichman 1992) y así evitar la sobreestimación del tamaño de la población; además se le registró la clase de edad (metamórfico, juvenil o adulto). La clasificación de los individuos capturados en los diferentes estadios fue realizada mediante la observación de características morfológicas siguiendo a Duellman & Trueb (1994). Todos los individuos fueron liberados luego en cada sitio de captura. Asimismo, se realizaron muestreos usando la técnica de relevamiento por encuentros visuales (Crump & Scott 1994), mediante recorridos por los bordes de los cuerpos de agua, no solo para la captura y recaptura de individuos, sino también para la observación de oviposiciones y larvas. Los recorridos se realizaron durante 30min aproximadamente, registrando la mayor parte del borde correspondiente a cada laguna. Para estimar el número de huevos por puesta se utilizó la técnica descrita en Bionda et al. (2011b). Para el muestreo de larvas, se realizó una captura en cada sitio transcurridas dos semanas aproximadamente del inicio de la reproducción y la observación de oviposiciones para cada especie. Esta captura consistió de seis barridas con un copo a lo largo de una transecta desde el margen del cuerpo de agua hasta 2m dentro del mismo, con un área de muestreo de 10m2. La abundancia total de larvas para cada especie se obtuvo a partir de la suma del número de larvas capturadas entre todas las barridas. Se realizaron análisis de contingencia (Zar 1996), para corroborar si la abundancia total de cada especie (a partir de los datos de las trampas de caída y los relevamientos por encuentros visuales), es similar entre sitios.
Durante cada muestreo realizado para la captura de individuos, se registró además las siguientes variables ambientales: temperatura del aire (TA; a 100cm del suelo con un termómetro de mercurio) y del agua (TAg; a la mitad de la columna de agua y con un termómetro de mercurio), pH del agua (a 5cm de profundidad aproximadamente y con cintas indicadoras de pH) y profundidad de las lagunas (PL; con cinta métrica). Además, el registro de precipitación caída, fue provista por la estación meteorológica perteneciente a la Cátedra de Meteorología Agrícola (Universidad Nacional de Río Cuarto), distante a unos 15km aproximadamente de los sitios de muestreo. Se realizó la toma de muestras de agua en todos los sitios para la medición de niveles de nitrato (N) y fósforo (F), utilizando los métodos Colorimétrico y de Espectrofotometría visible, respectivamente (USEPA 1994).
Con los datos de abundancia fue estimada la supervivencia para cada clase de edad (huevos, larvas, metamórficos y juveniles) para la construcción de matrices de proyección poblacional o matrices de Leslie. Estas estimaciones fueron realizadas con el método KNM (Kiritani Nakasuki Manly) siguiendo a Bécart et al. (2007). Este método es el más apropiado si se trabaja con especies que exhiben un comportamiento multicohorte (Manly 1989, Rhodes 2004, Bécart et al. 2007). Por otra parte, las tasas de supervivencia por clase de edad fueron calculadas para cada evento reproductivo, y siendo que los adultos capturados para cada año no han nacido en dicho evento reproductivo sino que los mismos nacieron en años anteriores, el dato para el número de adultos fue estimado en función al número de juveniles registrados, teniendo en cuenta una mortalidad promedio de 0.45 (Perry & Goodall 1979, Wells 2007). Se realizó una validación del método KNM, comparando para ello las tasas de supervivencia obtenidas por este método y las calculadas mediante la proporción entre los valores absolutos de abundancia para las distintas clases de edad. Para corroborar si existían diferencias significativas entre ambas estimaciones, se aplico el t-test para muestras pareadas (datos previamente modificados con la transformación arcsen por tratarse de proporciones según Zar 1996) bajo la hipótesis nula de que ambas estimaciones, KNM y proporciones, son iguales. Para realizar estas comparaciones se utilizó el programa Statistica (Statsoft 2001). Además, se calculó la fuerza de la mortalidad o killing power para cada estadio siguiendo a Begon et al. (2006). Finalmente, con la tasa de supervivencia para cada clase de edad y la fertilidad poblacional (número de huevos promedio por puesta), fueron construidas las matrices de proyección poblacional o matrices de Leslie (1945) que permiten obtener una predicción cuantitativa del tamaño poblacional, realizado para 30 generaciones.
Resultados
En ambos periodos de muestreo, se registró una precipitación semejante (2008-2009=575mm y 2009-2010=444mm totales entre los meses de septiembre a abril). En el cuadro 1 se muestran los valores medios y desvíos estándar de las variables ambientales registradas en cada sitio de muestreo. En general, las temperaturas fueron más elevadas para el periodo 2009-2010, particularmente una mayor temperatura del agua fue registrada para las lagunas del C2 lo cual podría estar asociado a una menor profundidad de las mismas. Los valores de pH del agua, indican que el sitio SM1 registró los valores más elevados, al igual que el C1 para 2008-2009, en ambos casos con promedios mayores a ocho; mientras que en los sitios C2 y SM2, los valores oscilaron entre seis y siete, para ambos periodos de muestreo. De acuerdo con los valores observados en la profundidad de las lagunas, el sitio SM2 presenta las lagunas de mayor profundidad. Por su parte, el C2 indicó una disminución considerable en la profundidad de las lagunas para el último periodo de muestreo. Los niveles de nitrato presentan valores aceptables (por debajo de los valores límites de acuerdo USEPA (1994) y OMS (2006) con excepción de lo ocurrido en las lagunas del C2, con valores considerablemente mayores para las muestras tomadas en 2008-2009. En cuanto a los valores de fósforo, en todos los sitios se registraron niveles por encima de los límites establecidos de acuerdo a USEPA (1994) y OMS (2006); los valores más altos se registraron para el C2 en 2008-2009.
De acuerdo con los datos registrados para R. arenarum, la abundancia de esta especie difiere entre los sitios (X2 C1-C2=173.22, X2 SM1-C1=114.03, X2 SM1-C2=169.92, X2 SM1-SM2=108.40, X2 C1-SM2=297, X2 C2-SM2=450.45, p<0.01 y gl=14 para todos los casos). Sin embargo, para P. biligonigerus, la abundancia es similar entre los sitios alterados (X2 C1-C2=18.09, X2 SM1-C2=22.77, X2 SM1-C1=9.90, p>0.05 y gl=14 para todos los casos) y difieren de la encontrada para SM2 (X2 C1-SM2=26.25, X2 C2-SM2=28.93, X2 SM1-SM2=27.58, p<0.05 y gl=14 para todos los casos).
En el cuadro 2 se muestran las medias de supervivencia según KNM y los valores para la fuerza de la mortalidad o killing power para cada clase de edad, obtenidos a partir de los valores de supervivencia y mortalidad registrados para cada año de muestreo. En R. arenarum, puede notarse que los ambientes alterados presentan una baja supervivencia para los huevos, principalmente los sitios C1 y SM1, y que el sitio C2 además muestra la menor supervivencia para el estadio larval (Cuadro 2, A). Los estadios que más podrían comprometer a las poblaciones son los de huevos y metamórficos, teniendo en cuenta que registran los valores más altos de mortalidad (Cuadro 2, B). En P. biligonigerus, se puede destacar la baja supervivencia que presentan los huevos en los sitios C2 y SM1 (Cuadro 2, A). La mortalidad actuó con mayor fuerza en el estadio de huevos, lo que indica que este estadio es el que más podría comprometer a las poblaciones; donde C2 registró la mortalidad más alta (Cuadro 2, B). Por otra parte, el sitio SM1 registró una mortalidad alta para el estadio de los metamórficos. Cabe aclarar que para esta especie ha resultado muy difícil la captura de larvas, sólo en el sitio C2 fue capturado un número significativo de las mismas, valores que pueden ser observados en los vectores etarios de la figura 1, sin embargo, por contar con este único registro entre los cuatro sitios de muestreo, no se calculó para este estadio la tasa de supervivencia y la fuerza de la mortalidad.
En la figura 1 se muestran las proyecciones poblacionales obtenidas para 30 generaciones y los vectores etarios utilizados para el inicio de dichas simulaciones. El vector etario fue construido con el promedio de las abundancias obtenidas para cada clase de edad durante los dos años de muestreo. Las proyecciones parecen ser favorables para ambas especies en el sitio SM2. R. arenarum muestra proyecciones descendentes en sus tamaños poblacionales en los sitios de cultivos más alterados (C1 y C2) y parece mantener su tamaño poblacional a lo largo de las generaciones en el sitio SM1. Por su parte, P. biligonigerus, indicó proyecciones desfavorables para todos los sitios alterados. Particularmente, para los sitios C2 y SM1, el modelo predice la extinción de las poblaciones entre las generaciones 12 y 13, respectivamente. Esta especie muestra para el sitio de menor alteración, una proyección poblacional estable que mantiene el tamaño poblacional a lo largo de las generaciones. Al analizar los vectores etarios en cada proyección, se puede advertir que a pesar de la alta fertilidad que registran las especies en sitios como SM1 y C2, presentan proyecciones no favorables en el tiempo. Los resultados de validación para el método KNM no indicó diferencias significativas entre las tasas de supervivencia calculadas usando el método y las estimadas mediante la proporción entre los valores absolutos de abundancia por clase de edad; es decir, el método KNM provee una estimación precisa de las tasas de supervivencia para todos los estadios (t-test para muestras pareadas, p>0.05).
Discusión
La dinámica poblacional es una especialidad de la ecología que estudia los mecanismos y las consecuencias de las variaciones demográficas, usualmente fluctuaciones numéricas (Carey 1993). Las fuerzas que regulan esta dinámica ha sido un tema central y controversial en los estudios sobre la estructuración de las comunidades en general (Begon et al. 2006, Wang et al. 2009). Varias características de los agroecosistemas pueden constituir factores de mortalidad que podrían en última instancia comprometer la persistencia de las poblaciones. A escala del paisaje, la especialización de los agroecosistemas hacia ciertos cultivos como ocurre con la soja para la región de este estudio, disminuye drásticamente la biodiversidad de un ecosistema la cual proporciona la “resiliencia” o la capacidad de los mismos para recuperarse o adaptarse a un entorno cambiante (Moone & Barberi 2008). Esto plantea serios cuestionamientos sobre qué tipos de agroecosistemas permitirían conservar la biodiversidad (Perfecto & Vandermeer 2008). Específicamente a lo que se refiere a la zona neotropical, los estudios con especies de anfibios en ecosistemas agrícolas han indicado una riqueza y diversidad reducida en dichos ambientes (Peltzer et al. 2006, Bionda et al. 2011a), en este sentido, la presencia de ganado puede afectar de manera negativa a las distintas poblaciones (Knutson et al. 2004, Schmutzer et al. 2008, Burton et al. 2009, Bionda et al. 2011a), aunque estos efectos pueden ser más marcados en algunas especies que en otras. Además, se ha comprobado que la presencia de agroquímicos causan alteraciones bioquímicas y fisiológicas, incrementando de este modo la mortalidad (e.g. Lajmanovich et al. 2004, Peltzer et al. 2007, Attademo et al. 2011, Whitfield 2011). Asimismo, una mala calidad de los cuerpos de agua, un hidroperiodo reducido (Peltzer et al. 2008, Bionda et al. 2011b) o una menor disponibilidad y tipo de alimento (Bionda et al. 2012), también constituyen factores en los agroecosistemas que contribuyen a disminuir la sobrevivencia poblacional. A pesar de todas estas evidencias, cabría plantearse si estos factores podrían comprometer a las poblaciones en el tiempo.
En este estudio, la predicción en la evolución de las distintas poblaciones teniendo en cuenta los valores de fertilidad y tasas de supervivencia, ha indicado una proyección favorable en el tiempo para el sitio menos alterado y desfavorables para las poblaciones de los ambientes dominados por cultivos. Particularmente para el C2, fue observada una baja supervivencia en huevos y larvas, y proyecciones poblacionales descendentes o próximas a la extinción según la especie. La baja supervivencia en el C2 de huevos y larvas, podría deberse a una mala calidad de agua en sus lagunas, con niveles de nitrato y fósforo excesivamente altos y condiciones tales como temperaturas y profundidad de las lagunas poco propicias. Varios autores han demostrado como el nitrato en cantidades excesivas, así como una alta temperatura, alteran el desarrollo y crecimiento larval, afectando de este modo su supervivencia (Boone & Bridges 1999, Edwards et al. 2006). Esta inestabilidad en las condiciones podrían ser razones suficientes para la baja supervivencia y proyecciones poblacionales desfavorables registradas para este sitio. Los valores excesivamente altos de nitrato en el agua de las lagunas del C2 en los años 2008-2009, comparado a lo ocurrido para 2009-2010, podrían ser consecuencias de diferentes prácticas en el manejo de los cultivos, como ser, una mayor superficie sembrada o una mayor aplicación de biocidas y fertilizantes durante la temporada 2008-2009, haciendo que los niveles de nitrato en el agua de las lagunas presentaran valores por encima de los aceptados en la normativa internacional (USEPA 1994, OMS 2006). Por su parte, los sitios C1 y SM1, también registraron bajas supervivencias y proyecciones poblacionales desfavorables. Para estos sitios se podría destacar los valores alcalinos de pH, con promedios por encima de ocho. Esta última observación, sumado a lo indicado para el C2, demuestra que en general los agroecosistemas en esta región, son ambientes posiblemente inhóspitos para la reproducción y la supervivencia de las especies estudiadas, particularmente para los estadios acuáticos
Por otra parte, de acuerdo a los resultados obtenidos para R. arenarum y P. biligonigerus, es posible reconocer que los efectos de este tipo de ambientes alterados, son especie-específicos. Coincidiendo de este modo con lo mencionado por Burton et al. (2009) y Lajmanovich et al. (2010) que demostraron las respuestas diferenciales de las especies y el riesgo ecológico interespecífico en los agroecosistemas. Al respecto, es posible apreciar que las proyecciones fueron más desfavorables para P. biligonigerus. Si bien R. arenarum presentó bajas supervivencias, particularmente en los sitios más alterados, esta especie mostró un mayor reclutamiento y proyecciones más favorables que P. biligonigerus. Es probable que algunas características de la biología y los requerimientos de hábitat de P. biligonigerus la conviertan en una especie más sensible a los cambios y modificaciones del hábitat. Según Cei (1980), P. biligonigerus puede ser considerada una especie de importantes requerimientos hacia hábitats con disponibilidad de agua. En contrapartida, las especies de anfibios de hábitos más terrestres como R. arenarum, son más generalistas en sus requerimientos de hábitats, y por ello, más independientes de las variables ambientales (Keller et al. 2009). A pesar de que el modo reproductivo de R. arenarum (huevos en cordones gelatinosos depositados dentro del agua) es más dependiente del agua que el de P. biligonigerus (huevos en nidos de espuma depositados sobre la superficie del agua), sus requerimientos de hábitats más generalistas puede llevar a una mayor supervivencia de los estadios postmetamórficos resultando en un mayor reclutamiento, comparado a P. biligonigerus. En este sentido, la posible modificación de los ambientes acuáticos en los agroecosistemas, en relación a la calidad del agua y su hidroperiodo, permite presuponer que aquellas especies de hábitos más acuáticos, pueden ser más sensibles a la alteración de estos tipos de ambientes. El trabajo realizado por Peltzer (2006) para la región centro-este de Argentina es concluyente al respecto, ya que en los ambientes alterados encontraron una mayor representatividad de anuros correspondientes al gremio terrestre, seguido del gremio semiacuático, y una declinación notable del gremio acuático. Sin embargo, otros trabajos, principalmente en arrozales, no encuentran diferencias significativas entre ambientes alterados y la composición de las especies según sus hábitos (Vasconcelos & Rossa Feres 2005, Schaefer 2006). A pesar de que P. biligonigerus registra una elevada reproducción en todos los ambientes analizados, particularmente un importante número de puestas de huevos fueron observadas en el C2, lo cual podría indicar una mayor capacidad reproductiva para explotar ambientes perturbados, luego se observa un bajo o nulo reclutamiento debido a las bajas supervivencias por clase de edad. En los agroecosistemas, P. biligonigerus mostró proyecciones desfavorables, y una tendencia a la extinción de las poblaciones en la mayor parte de los sitios analizados. P. biligonigerus presenta un comportamiento explosivo, con una elevada actividad reproductiva en un corto tiempo, luego de registrarse precipitaciones importantes (Martino 1999, Bionda 2011, Bionda et al. 2011a). Para Heyer (1974) las especies que se reproducen en ambientesvariables maximizan su esfuerzo reproductivo. Otros trabajos realizados con P. albonotatus, especie ecológicamente similar a P. biligonigerus, indica a P. albonotatus como una especie invasora de habitats agrícolas debido a la abundancia registrada (Peltzer et al. 2006, Lajmanovich et al. 2010). En el mismo sentido, un estudio realizado durante cuatro años sobre las comunidades de anfibios del departamento de Río Cuarto (Córdoba), demostró que P. biligonigerus fue la especie más abundante en los agroecosistemas, pero su abundancia varió de manera considerable entre años de muestreo, indicando una dinámica poco estable en dichos sitios (Bionda 2011).
Mediante este estudio se ha advertido que los agroecosistemas por sus características representan hábitats inhóspitos para las poblaciones de R. arenarum y P. biligonigerus, las cuales constituyen especies representativas de la herpetofauna regional por su abundancia y distribución. El hecho de que los estadios acuáticos registren en general una baja supervivencia en los agroecosistemas, permite pensar en una mala calidad de agua en las lagunas de dichos ambientes. Asimismo, podemos reconocer efectos especie-específicos de los ecosistemas agrícolas, ya que P. biligonigerus resultó la especie más vulnerable, posiblemente por sus características de historias de vida y requerimientos de hábitats. Finalmente, el deterioro ambiental producido por los cultivos extensivos de soja en la región central de la Argentina, impactaría en la demografía de las poblaciones de anuros, generando una reducción en el tamaño de sus poblaciones.
Agradecimientos
Este trabajo fue financiado por el Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET), la Agencia Nacional de Promoción Científica y Tecnológica (ANCyT) y SCyT UNRC, PPI 18/C350. Nuestro estudio fue autorizado por la Agencia Córdoba Ambiente (A.C.A.S.E.).
En este contexto, resulta valioso el conocimiento sobre la demografía de las poblaciones de anfibios que habitan los ecosistemas agrícolas (Heyer 1974, Wilbur 1980, Guerry & Hunter 2002), ya que al estimar parámetros poblacionales, se puede establecer el estado de conservación de las especies. De esta manera, es posible determinar si las poblaciones de anfibios que habitan agroecosistemas son afectadas por dicha actividad, o si simplemente manifiestan fluctuaciones naturales en sus tamaños poblacionales (Beebee 1996). Las áreas cultivadas, entre otras consecuencias, producen la alteración de los ambientes acuáticos o un deterioro en la calidad de los mismos, en particular en la duración del hidroperiodo, que puede afectar la supervivencia de huevos y larvas de anfibios (Carey & Bryant 1995, Taylor et al. 2005, Altig et al. 2007, Peltzer et al. 2008). Estos efectos tienden a modificar los rasgos de historias de vida como tamaño y condición corporal, supervivencia, fertilidad, entre otros (Márquez García et al. 2009, Bionda et al. 2012), que en última instancia podrían comprometer la permanencia de las poblaciones en el tiempo. La determinación de la fecundidad y la supervivencia constituyen parámetros básicos de una población, ya que proveen información acerca de las variaciones demográficas, como las fluctuaciones numéricas o la magnitud del crecimiento poblacional (Carey 1993, Shaffer 1994, Sinsch et al. 2001), que permitiría esclarecer de qué manera ambientes como los agroecosistemas pueden afectar a las poblaciones de anfibios. En este trabajo nos proponemos analizar la demografía poblacional de dos especies de anfibios asociadas a sistemas agrícolas de la región central de la provincia de Córdoba, Rhinella arenarum Hensel 1867, y Physalaemus biligonigerus Cope 1861.
Materiales y métodos
Los sitios de muestreo se encuentran en los alrededores de la localidad de Río Cuarto (33º07’55’’18’’ S - 64º21’08’’96’’ W), provincia de Córdoba, Argentina. La región se caracteriza por tener clima semiseco, con tendencia a semihúmedo. Las lluvias máximas ocurren durante el periodo octubre-marzo, con una media anual de 800-1 000mm (Bionda 2011, Gatica et al. 2012). La fisiografía del área de estudio corresponde a llanuras suavemente onduladas. La escasa pendiente favorece el estancamiento de las aguas, sumado al aporte subterráneo que junto con las precipitaciones, ayuda a mantener cursos de agua permanentes (Bridarolli & di Tada 1996).
La selección de las especies, R. arenarum y P. biligonigerus, se basó en su representatividad en las comunidades de anfibios de la región de estudio (Bionda et al. 2011a). Ambas especies de anuros presentan una amplia distribución en el Neotrópico, siendo encontradas en Argentina, Bolivia, Brasil, Uruguay y Paraguay (IUCN 2011), y están consideradas como especies No Amenazadas (Lavilla et al. 2000) o de Preocupación Menor (IUCN 2011). La recolección de los especímenes fue realizada en cuatro sitios, tres de ellos se encuentran en zonas de cultivos de soja: Sitio Cultivo 1 (C1, 33º05’54’’38’’ S - 64º26’04’’76’’ W), Sitio Cultivo 2 (C2, 33º05’54’’79’’ S - 64º25’46’’77’’ W) y Sitio Semimodificado 1 (SM1, 33º05’39’’29’’ S - 64º25’57’’72’’ W). Los tres sitios presentan distintos grados de alteración en relación a la distancia de los cultivos a los cuerpos de agua. En C1 y C2, los cultivos están a escasos metros de los cuerpos de agua (15m aproximadamente), mientras que en SM1 se encuentran más alejados (200m aproximadamente). En los tres sitios existe la práctica ganadera, aunque el sitio C1 presenta esta actividad con una mayor intensidad (presencia de ganado a diario y en cercanía o dentro de lagunas). El cuarto sitio de muestreo, llamado Sitio Semimodificado 2 (SM2, 33º06’41’’95’’ S - 64º18’14’’71’’ W), corresponde a una zona periurbana no afectada por cultivos, cubierta por pastizales y formaciones boscosas de árboles autóctonos e introducidos (Doffo 1989).
El muestreo en cada sitio fue realizado durante dos periodos reproductivos, entre septiembre 2008 y abril 2009 y septiembre 2009 hasta abril 2010. Los muestreos fueron sistemáticos, con dos o tres visitas semanales dependiendo de las precipitaciones. Los individuos fueron capturados utilizando trampas de caída viva (Corn 1994). Estas trampas fueron dispuestas en dos líneas, estas últimas a una distancia de 6 a 8m de los cuerpos de agua. Cada línea de trampas consistió de ocho recipientes de plástico de 22cm de diámetro y 40cm de profundidad, espaciadas a 2.5m. El fondo de cada trampa fue humedecido para prevenir la desecación. Cada individuo capturado fue removido para ser marcado mediante la amputación de falanges (Waichman 1992) y así evitar la sobreestimación del tamaño de la población; además se le registró la clase de edad (metamórfico, juvenil o adulto). La clasificación de los individuos capturados en los diferentes estadios fue realizada mediante la observación de características morfológicas siguiendo a Duellman & Trueb (1994). Todos los individuos fueron liberados luego en cada sitio de captura. Asimismo, se realizaron muestreos usando la técnica de relevamiento por encuentros visuales (Crump & Scott 1994), mediante recorridos por los bordes de los cuerpos de agua, no solo para la captura y recaptura de individuos, sino también para la observación de oviposiciones y larvas. Los recorridos se realizaron durante 30min aproximadamente, registrando la mayor parte del borde correspondiente a cada laguna. Para estimar el número de huevos por puesta se utilizó la técnica descrita en Bionda et al. (2011b). Para el muestreo de larvas, se realizó una captura en cada sitio transcurridas dos semanas aproximadamente del inicio de la reproducción y la observación de oviposiciones para cada especie. Esta captura consistió de seis barridas con un copo a lo largo de una transecta desde el margen del cuerpo de agua hasta 2m dentro del mismo, con un área de muestreo de 10m2. La abundancia total de larvas para cada especie se obtuvo a partir de la suma del número de larvas capturadas entre todas las barridas. Se realizaron análisis de contingencia (Zar 1996), para corroborar si la abundancia total de cada especie (a partir de los datos de las trampas de caída y los relevamientos por encuentros visuales), es similar entre sitios.
Durante cada muestreo realizado para la captura de individuos, se registró además las siguientes variables ambientales: temperatura del aire (TA; a 100cm del suelo con un termómetro de mercurio) y del agua (TAg; a la mitad de la columna de agua y con un termómetro de mercurio), pH del agua (a 5cm de profundidad aproximadamente y con cintas indicadoras de pH) y profundidad de las lagunas (PL; con cinta métrica). Además, el registro de precipitación caída, fue provista por la estación meteorológica perteneciente a la Cátedra de Meteorología Agrícola (Universidad Nacional de Río Cuarto), distante a unos 15km aproximadamente de los sitios de muestreo. Se realizó la toma de muestras de agua en todos los sitios para la medición de niveles de nitrato (N) y fósforo (F), utilizando los métodos Colorimétrico y de Espectrofotometría visible, respectivamente (USEPA 1994).
Con los datos de abundancia fue estimada la supervivencia para cada clase de edad (huevos, larvas, metamórficos y juveniles) para la construcción de matrices de proyección poblacional o matrices de Leslie. Estas estimaciones fueron realizadas con el método KNM (Kiritani Nakasuki Manly) siguiendo a Bécart et al. (2007). Este método es el más apropiado si se trabaja con especies que exhiben un comportamiento multicohorte (Manly 1989, Rhodes 2004, Bécart et al. 2007). Por otra parte, las tasas de supervivencia por clase de edad fueron calculadas para cada evento reproductivo, y siendo que los adultos capturados para cada año no han nacido en dicho evento reproductivo sino que los mismos nacieron en años anteriores, el dato para el número de adultos fue estimado en función al número de juveniles registrados, teniendo en cuenta una mortalidad promedio de 0.45 (Perry & Goodall 1979, Wells 2007). Se realizó una validación del método KNM, comparando para ello las tasas de supervivencia obtenidas por este método y las calculadas mediante la proporción entre los valores absolutos de abundancia para las distintas clases de edad. Para corroborar si existían diferencias significativas entre ambas estimaciones, se aplico el t-test para muestras pareadas (datos previamente modificados con la transformación arcsen por tratarse de proporciones según Zar 1996) bajo la hipótesis nula de que ambas estimaciones, KNM y proporciones, son iguales. Para realizar estas comparaciones se utilizó el programa Statistica (Statsoft 2001). Además, se calculó la fuerza de la mortalidad o killing power para cada estadio siguiendo a Begon et al. (2006). Finalmente, con la tasa de supervivencia para cada clase de edad y la fertilidad poblacional (número de huevos promedio por puesta), fueron construidas las matrices de proyección poblacional o matrices de Leslie (1945) que permiten obtener una predicción cuantitativa del tamaño poblacional, realizado para 30 generaciones.
Resultados
En ambos periodos de muestreo, se registró una precipitación semejante (2008-2009=575mm y 2009-2010=444mm totales entre los meses de septiembre a abril). En el cuadro 1 se muestran los valores medios y desvíos estándar de las variables ambientales registradas en cada sitio de muestreo. En general, las temperaturas fueron más elevadas para el periodo 2009-2010, particularmente una mayor temperatura del agua fue registrada para las lagunas del C2 lo cual podría estar asociado a una menor profundidad de las mismas. Los valores de pH del agua, indican que el sitio SM1 registró los valores más elevados, al igual que el C1 para 2008-2009, en ambos casos con promedios mayores a ocho; mientras que en los sitios C2 y SM2, los valores oscilaron entre seis y siete, para ambos periodos de muestreo. De acuerdo con los valores observados en la profundidad de las lagunas, el sitio SM2 presenta las lagunas de mayor profundidad. Por su parte, el C2 indicó una disminución considerable en la profundidad de las lagunas para el último periodo de muestreo. Los niveles de nitrato presentan valores aceptables (por debajo de los valores límites de acuerdo USEPA (1994) y OMS (2006) con excepción de lo ocurrido en las lagunas del C2, con valores considerablemente mayores para las muestras tomadas en 2008-2009. En cuanto a los valores de fósforo, en todos los sitios se registraron niveles por encima de los límites establecidos de acuerdo a USEPA (1994) y OMS (2006); los valores más altos se registraron para el C2 en 2008-2009.
De acuerdo con los datos registrados para R. arenarum, la abundancia de esta especie difiere entre los sitios (X2 C1-C2=173.22, X2 SM1-C1=114.03, X2 SM1-C2=169.92, X2 SM1-SM2=108.40, X2 C1-SM2=297, X2 C2-SM2=450.45, p<0.01 y gl=14 para todos los casos). Sin embargo, para P. biligonigerus, la abundancia es similar entre los sitios alterados (X2 C1-C2=18.09, X2 SM1-C2=22.77, X2 SM1-C1=9.90, p>0.05 y gl=14 para todos los casos) y difieren de la encontrada para SM2 (X2 C1-SM2=26.25, X2 C2-SM2=28.93, X2 SM1-SM2=27.58, p<0.05 y gl=14 para todos los casos).
En el cuadro 2 se muestran las medias de supervivencia según KNM y los valores para la fuerza de la mortalidad o killing power para cada clase de edad, obtenidos a partir de los valores de supervivencia y mortalidad registrados para cada año de muestreo. En R. arenarum, puede notarse que los ambientes alterados presentan una baja supervivencia para los huevos, principalmente los sitios C1 y SM1, y que el sitio C2 además muestra la menor supervivencia para el estadio larval (Cuadro 2, A). Los estadios que más podrían comprometer a las poblaciones son los de huevos y metamórficos, teniendo en cuenta que registran los valores más altos de mortalidad (Cuadro 2, B). En P. biligonigerus, se puede destacar la baja supervivencia que presentan los huevos en los sitios C2 y SM1 (Cuadro 2, A). La mortalidad actuó con mayor fuerza en el estadio de huevos, lo que indica que este estadio es el que más podría comprometer a las poblaciones; donde C2 registró la mortalidad más alta (Cuadro 2, B). Por otra parte, el sitio SM1 registró una mortalidad alta para el estadio de los metamórficos. Cabe aclarar que para esta especie ha resultado muy difícil la captura de larvas, sólo en el sitio C2 fue capturado un número significativo de las mismas, valores que pueden ser observados en los vectores etarios de la figura 1, sin embargo, por contar con este único registro entre los cuatro sitios de muestreo, no se calculó para este estadio la tasa de supervivencia y la fuerza de la mortalidad.
En la figura 1 se muestran las proyecciones poblacionales obtenidas para 30 generaciones y los vectores etarios utilizados para el inicio de dichas simulaciones. El vector etario fue construido con el promedio de las abundancias obtenidas para cada clase de edad durante los dos años de muestreo. Las proyecciones parecen ser favorables para ambas especies en el sitio SM2. R. arenarum muestra proyecciones descendentes en sus tamaños poblacionales en los sitios de cultivos más alterados (C1 y C2) y parece mantener su tamaño poblacional a lo largo de las generaciones en el sitio SM1. Por su parte, P. biligonigerus, indicó proyecciones desfavorables para todos los sitios alterados. Particularmente, para los sitios C2 y SM1, el modelo predice la extinción de las poblaciones entre las generaciones 12 y 13, respectivamente. Esta especie muestra para el sitio de menor alteración, una proyección poblacional estable que mantiene el tamaño poblacional a lo largo de las generaciones. Al analizar los vectores etarios en cada proyección, se puede advertir que a pesar de la alta fertilidad que registran las especies en sitios como SM1 y C2, presentan proyecciones no favorables en el tiempo. Los resultados de validación para el método KNM no indicó diferencias significativas entre las tasas de supervivencia calculadas usando el método y las estimadas mediante la proporción entre los valores absolutos de abundancia por clase de edad; es decir, el método KNM provee una estimación precisa de las tasas de supervivencia para todos los estadios (t-test para muestras pareadas, p>0.05).
Discusión
La dinámica poblacional es una especialidad de la ecología que estudia los mecanismos y las consecuencias de las variaciones demográficas, usualmente fluctuaciones numéricas (Carey 1993). Las fuerzas que regulan esta dinámica ha sido un tema central y controversial en los estudios sobre la estructuración de las comunidades en general (Begon et al. 2006, Wang et al. 2009). Varias características de los agroecosistemas pueden constituir factores de mortalidad que podrían en última instancia comprometer la persistencia de las poblaciones. A escala del paisaje, la especialización de los agroecosistemas hacia ciertos cultivos como ocurre con la soja para la región de este estudio, disminuye drásticamente la biodiversidad de un ecosistema la cual proporciona la “resiliencia” o la capacidad de los mismos para recuperarse o adaptarse a un entorno cambiante (Moone & Barberi 2008). Esto plantea serios cuestionamientos sobre qué tipos de agroecosistemas permitirían conservar la biodiversidad (Perfecto & Vandermeer 2008). Específicamente a lo que se refiere a la zona neotropical, los estudios con especies de anfibios en ecosistemas agrícolas han indicado una riqueza y diversidad reducida en dichos ambientes (Peltzer et al. 2006, Bionda et al. 2011a), en este sentido, la presencia de ganado puede afectar de manera negativa a las distintas poblaciones (Knutson et al. 2004, Schmutzer et al. 2008, Burton et al. 2009, Bionda et al. 2011a), aunque estos efectos pueden ser más marcados en algunas especies que en otras. Además, se ha comprobado que la presencia de agroquímicos causan alteraciones bioquímicas y fisiológicas, incrementando de este modo la mortalidad (e.g. Lajmanovich et al. 2004, Peltzer et al. 2007, Attademo et al. 2011, Whitfield 2011). Asimismo, una mala calidad de los cuerpos de agua, un hidroperiodo reducido (Peltzer et al. 2008, Bionda et al. 2011b) o una menor disponibilidad y tipo de alimento (Bionda et al. 2012), también constituyen factores en los agroecosistemas que contribuyen a disminuir la sobrevivencia poblacional. A pesar de todas estas evidencias, cabría plantearse si estos factores podrían comprometer a las poblaciones en el tiempo.
En este estudio, la predicción en la evolución de las distintas poblaciones teniendo en cuenta los valores de fertilidad y tasas de supervivencia, ha indicado una proyección favorable en el tiempo para el sitio menos alterado y desfavorables para las poblaciones de los ambientes dominados por cultivos. Particularmente para el C2, fue observada una baja supervivencia en huevos y larvas, y proyecciones poblacionales descendentes o próximas a la extinción según la especie. La baja supervivencia en el C2 de huevos y larvas, podría deberse a una mala calidad de agua en sus lagunas, con niveles de nitrato y fósforo excesivamente altos y condiciones tales como temperaturas y profundidad de las lagunas poco propicias. Varios autores han demostrado como el nitrato en cantidades excesivas, así como una alta temperatura, alteran el desarrollo y crecimiento larval, afectando de este modo su supervivencia (Boone & Bridges 1999, Edwards et al. 2006). Esta inestabilidad en las condiciones podrían ser razones suficientes para la baja supervivencia y proyecciones poblacionales desfavorables registradas para este sitio. Los valores excesivamente altos de nitrato en el agua de las lagunas del C2 en los años 2008-2009, comparado a lo ocurrido para 2009-2010, podrían ser consecuencias de diferentes prácticas en el manejo de los cultivos, como ser, una mayor superficie sembrada o una mayor aplicación de biocidas y fertilizantes durante la temporada 2008-2009, haciendo que los niveles de nitrato en el agua de las lagunas presentaran valores por encima de los aceptados en la normativa internacional (USEPA 1994, OMS 2006). Por su parte, los sitios C1 y SM1, también registraron bajas supervivencias y proyecciones poblacionales desfavorables. Para estos sitios se podría destacar los valores alcalinos de pH, con promedios por encima de ocho. Esta última observación, sumado a lo indicado para el C2, demuestra que en general los agroecosistemas en esta región, son ambientes posiblemente inhóspitos para la reproducción y la supervivencia de las especies estudiadas, particularmente para los estadios acuáticos
Por otra parte, de acuerdo a los resultados obtenidos para R. arenarum y P. biligonigerus, es posible reconocer que los efectos de este tipo de ambientes alterados, son especie-específicos. Coincidiendo de este modo con lo mencionado por Burton et al. (2009) y Lajmanovich et al. (2010) que demostraron las respuestas diferenciales de las especies y el riesgo ecológico interespecífico en los agroecosistemas. Al respecto, es posible apreciar que las proyecciones fueron más desfavorables para P. biligonigerus. Si bien R. arenarum presentó bajas supervivencias, particularmente en los sitios más alterados, esta especie mostró un mayor reclutamiento y proyecciones más favorables que P. biligonigerus. Es probable que algunas características de la biología y los requerimientos de hábitat de P. biligonigerus la conviertan en una especie más sensible a los cambios y modificaciones del hábitat. Según Cei (1980), P. biligonigerus puede ser considerada una especie de importantes requerimientos hacia hábitats con disponibilidad de agua. En contrapartida, las especies de anfibios de hábitos más terrestres como R. arenarum, son más generalistas en sus requerimientos de hábitats, y por ello, más independientes de las variables ambientales (Keller et al. 2009). A pesar de que el modo reproductivo de R. arenarum (huevos en cordones gelatinosos depositados dentro del agua) es más dependiente del agua que el de P. biligonigerus (huevos en nidos de espuma depositados sobre la superficie del agua), sus requerimientos de hábitats más generalistas puede llevar a una mayor supervivencia de los estadios postmetamórficos resultando en un mayor reclutamiento, comparado a P. biligonigerus. En este sentido, la posible modificación de los ambientes acuáticos en los agroecosistemas, en relación a la calidad del agua y su hidroperiodo, permite presuponer que aquellas especies de hábitos más acuáticos, pueden ser más sensibles a la alteración de estos tipos de ambientes. El trabajo realizado por Peltzer (2006) para la región centro-este de Argentina es concluyente al respecto, ya que en los ambientes alterados encontraron una mayor representatividad de anuros correspondientes al gremio terrestre, seguido del gremio semiacuático, y una declinación notable del gremio acuático. Sin embargo, otros trabajos, principalmente en arrozales, no encuentran diferencias significativas entre ambientes alterados y la composición de las especies según sus hábitos (Vasconcelos & Rossa Feres 2005, Schaefer 2006). A pesar de que P. biligonigerus registra una elevada reproducción en todos los ambientes analizados, particularmente un importante número de puestas de huevos fueron observadas en el C2, lo cual podría indicar una mayor capacidad reproductiva para explotar ambientes perturbados, luego se observa un bajo o nulo reclutamiento debido a las bajas supervivencias por clase de edad. En los agroecosistemas, P. biligonigerus mostró proyecciones desfavorables, y una tendencia a la extinción de las poblaciones en la mayor parte de los sitios analizados. P. biligonigerus presenta un comportamiento explosivo, con una elevada actividad reproductiva en un corto tiempo, luego de registrarse precipitaciones importantes (Martino 1999, Bionda 2011, Bionda et al. 2011a). Para Heyer (1974) las especies que se reproducen en ambientesvariables maximizan su esfuerzo reproductivo. Otros trabajos realizados con P. albonotatus, especie ecológicamente similar a P. biligonigerus, indica a P. albonotatus como una especie invasora de habitats agrícolas debido a la abundancia registrada (Peltzer et al. 2006, Lajmanovich et al. 2010). En el mismo sentido, un estudio realizado durante cuatro años sobre las comunidades de anfibios del departamento de Río Cuarto (Córdoba), demostró que P. biligonigerus fue la especie más abundante en los agroecosistemas, pero su abundancia varió de manera considerable entre años de muestreo, indicando una dinámica poco estable en dichos sitios (Bionda 2011).
Mediante este estudio se ha advertido que los agroecosistemas por sus características representan hábitats inhóspitos para las poblaciones de R. arenarum y P. biligonigerus, las cuales constituyen especies representativas de la herpetofauna regional por su abundancia y distribución. El hecho de que los estadios acuáticos registren en general una baja supervivencia en los agroecosistemas, permite pensar en una mala calidad de agua en las lagunas de dichos ambientes. Asimismo, podemos reconocer efectos especie-específicos de los ecosistemas agrícolas, ya que P. biligonigerus resultó la especie más vulnerable, posiblemente por sus características de historias de vida y requerimientos de hábitats. Finalmente, el deterioro ambiental producido por los cultivos extensivos de soja en la región central de la Argentina, impactaría en la demografía de las poblaciones de anuros, generando una reducción en el tamaño de sus poblaciones.
Agradecimientos
Este trabajo fue financiado por el Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET), la Agencia Nacional de Promoción Científica y Tecnológica (ANCyT) y SCyT UNRC, PPI 18/C350. Nuestro estudio fue autorizado por la Agencia Córdoba Ambiente (A.C.A.S.E.).
Referencias
*Correspondencia para:
Clarisa Bionda. Ecología, Departamento de Ciencias Naturales, Facultad de Ciencias Exactas, Físico-Químicas y Naturales, UNRC, ruta 36km 601, Río Cuarto, Córdoba, Argentina; Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET), Argentina; cbionda@exa.unrc.edu.ar
Rafael Lajmanovich. Ecotoxicología, Escuela Superior de Sanidad “Dr. Ramón Carrillo”, Facultad de Bioquímica y Ciencias Biológicas, Paraje “El Pozo” s/n, Santa Fe, Argentina; Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET), Argentina; lajmanovich@hotmail.com
Nancy Salas. Ecología, Departamento de Ciencias Naturales, Facultad de Ciencias Exactas, Físico-Químicas y Naturales, UNRC, ruta 36km 601, Río Cuarto, Córdoba, Argentina; nsalas@exa.unrc.edu.ar
Adolfo Martino. Ecología, Departamento de Ciencias Naturales, Facultad de Ciencias Exactas, Físico-Químicas y Naturales, UNRC, ruta 36km 601, Río Cuarto, Córdoba, Argentina; amartino@exa.unrc.edu.ar
Ismael di Tada. Ecología, Departamento de Ciencias Naturales, Facultad de Ciencias Exactas, Físico-Químicas y Naturales, UNRC, ruta 36km 601, Río Cuarto, Córdoba, Argentina; iditada@gmail.com
1. Ecología, Departamento de Ciencias Naturales, Facultad de Ciencias Exactas, Físico-Químicas y Naturales, UNRC, ruta 36km 601, Río Cuarto, Córdoba, Argentina; cbionda@exa.unrc.edu.ar, nsalas@exa.unrc.edu.ar, amartino@exa.unrc.edu.ar, iditada@gmail.com
2. Ecotoxicología, Escuela Superior de Sanidad “Dr. Ramón Carrillo”, Facultad de Bioquímica y Ciencias Biológicas, Paraje “El Pozo” s/n, Santa Fe, Argentina; lajmanovich@hotmail.com
3. Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET), Argentina
Altieri, M. & W. Pengue. 2006. GM Soybean: Latin America’s new colonizer. GRAIN, Barcelona, España. (Consultado: Junio 6, 2011, http://www.grain.org/seedling/index. cfm?id=421). [ Links ]
Altig, R., M.R. Whiles & C.L. Taylor. 2007. What do tadpoles really eat? Assessing the trophic status of an understudied and imperiled group of consumers in freshwater habitats. Freshwat. Biol. 52: 386-395. [ Links ]
Attademo, A.M., M. Cabagna Zenklusen, R.C. Lajmanovich, P.M. Peltzer, C. Junges & A. Bassó. 2011. Besterase activities and blood cell morphology in the Frog Leptodactylus chaquensis (Amphibia: Leptodactylidae) on rice agroecosystems from Santa Fe Province (Argentina). Ecotoxicology 20: 274-282. [ Links ]
Bécart, E., A. Aubry & M. Emmerson. 2007. Monitoring the conservation status of natterjack toad (Bufo calamita) in Ireland, 2004 - 2006. Irish Wildlife Manuals, No. 31. National Parks and Wildlife Service, Department of the Environment, Heritage and Local. Government, Dublín, Irlanda. [ Links ]
Beebee, T.J.C. 1996. Ecology and conservation of amphibians. Conservation biology series. Chapman & Hall, Londres, Inglaterra. [ Links ]
Beebee, T.J.C. & R.A. Griffiths. 2005. The amphibian decline crisis: A watershed for conservation biology? Biol. Conservat. 125: 271-285. [ Links ]
Begon, M., C.R. Townsend & J.L. Harper. 2006. Ecology: from individuals to ecosystems. Blackwell, Oxford, Oxforshire, Inglaterra. [ Links ]
Bionda, C.L. 2011. Dinámica poblacional de anfibios asociados a cultivos extensivos de la provincia de Córdoba, Argentina. Tesis de Doctorado, Universidad Nacional de Río Cuarto, Río Cuarto, Córdoba, Argentina. [ Links ]
Bionda, C.L., I.E. di Tada & R.C. Lajmanovich. 2011a. Composition of amphibian assemblages in agroecosystems from the central region of Argentina. Russ. J. Herpetol. 18: 93-98. [ Links ]
Bionda, C.L., R.C. Lajmanovich, N.E. Salas, A.L. Martino & I.E. di Tada. 2011b. Reproductive Ecology of the Common South American Toad Rhinella arenarum (Anura: Bufonidae): Reproductive Effort, Clutch Size, Fecundity, and Mate Selection. J. Herpetol. 45: 261-264. [ Links ]
Bionda, C.L., N. Gari, E. Luque, N.E. Salas, R.C. Lajmanovich & A.L. Martino. 2012. Ecología trófica en larvas de Rhinella arenarum (Anura: Bufonidae) en agroecosistemas y sus posibles implicaciones para la conservación. Rev. Biol. Trop. 60: 771-779. [ Links ]
Boone, M.D. & C.M. Bridges. 1999. The effect of temperature on the potency of Carbaryl for survival of tadpoles of the green frog (Rana clamitans). Environ. Toxicol. Chem. 18: 1482-1484. [ Links ]
Bridarolli, M.E. & I.E. di Tada. 1996. Algunos aspectos de la geografía física de la provincia de Córdoba, p. 15-38. In I.E. di Tada & E.H. Bucher (eds.). Biodiversidad de la provincia de Córdoba. Fauna. Universidad Nacional de Río Cuarto, Río Cuarto, Córdoba, Argentina. [ Links ]
Burton, E.C., M.J. Gray, A.C. Schmutzer & D.L. Miller. 2009. Differential responses of postmetamorphic amphibians to cattle grazing in wetlands. J. Wildl. Manag. 73: 269-277. [ Links ]
Carey, J.R. 1993. Applied demography for biologists: with special emphasis on insects. Oxford University, Nueva York, EE.UU. [ Links ]
Carey, C. & C.J. Bryant. 1995. Possible interrelations among environmental toxicants, Amphibian development, and decline of Amphibian populations. Environ. Health Perspect. 103: 13-17. [ Links ]
Cei, J.M. 1980. Amphibians of Argentina. Monit. Zool. Ital. Nueva Serie, Florencia, Italia. [ Links ]
Collins, J.P. & M. Crump. 2009. Extinction in Our Time. Oxford University, Nueva York, EE.UU. [ Links ]
Corn, P.S. 1994. Cercas en línea recta y trampas de caída, p. 104-113. In W.R. Heyer, M.A. Donnelly, R.W. McDiarmid, L.C. Hayek & M.S. Foster (eds.). Medición y Monitoreo de la diversidad biológica. Métodos estandarizados para Anfibios. Universitaria de La Patagonia, Comodoro Rivadavia, Chubut, Argentina. [ Links ]
Crump, M.L. & N.J. Scott. 1994. Técnicas estándar para inventarios y monitoreo, p. 71-136. In W.R. Heyer, M.A. Donnelly, R.W. McDiarmid, L.C. Hayek & M.S. Foster (eds.). Medición y Monitoreo de la diversidad biológica. Métodos estandarizados para Anfibios. Universitaria de La Patagonia, Comodoro Rivadavia, Chubut, Argentina. [ Links ]
Davidson, C., H.B. Shaffer & M.R. Jennings. 2002. Spatial tests of the pesticide drift, habitat destruction, UV-B, and climate-change hypotheses for California amphibian declines. Conservat. Biol. 16: 1588-1601. [ Links ]
di Tada, I.E., M.V. Zavattieri, M.E. Bridarolli, N.E. Salas & A.L. Martino. 1996. Anfibios Anuros de la provincia de Córdoba, p. 191-215. In I.E. di Tada & E.H. Bucher (eds.). Biodiversidad de la provincia de Córdoba. Fauna. Universidad Nacional de Río Cuarto, Río Cuarto, Córdoba, Argentina. [ Links ]
Doffo, N. 1989. Geomorfología del área urbana de la ciudad de Río Cuarto y de la cuenca del arroyo El Bañado, algunas consideraciones aplicadas al manejo del medio natural. Tesis de grado, Universidad Nacional de Río Cuarto, Río Cuarto, Córdoba, Argentina. [ Links ]
Duellman, W.E. & L. Trueb. 1994. Biology of amphibians. Johns Hopkins, Londres, Inglaterra. [ Links ]
Edwards, T.M., K.A. McCoy, T. Barbeau, M.W. McCoy, J. Matthew Thro & L.J. Guillette Jr. 2006. Environmental context determines nitrate toxicity in Southern toad (Bufo terrestris) tadpoles. Aquat. Toxicol. 78: 50-58. [ Links ]
Gatica, E.A., C.A. Almeida, M.A. Mallea, M.C. Del Corigliano & P. González. 2012. Water quality assessment, by statistical analysis, on rural and urban areas of Chocancharava River (Río Cuarto), Córdoba, Argentina. Environ. Monit. Assess. DOI 10.1007/ s10661-011-2495-7. [ Links ]
Guerry, A.D. & M.L. Hunter Jr. 2002. Amphibian distribution in a landscape of forest and agriculture: an examination of landscape composition and configuration. Conservat. Biol. 16: 745-754. [ Links ]
Heyer, W.R. 1974. Niche measurements of frog larvae from a seasonal tropical location in Thailand. Ecology Tempe 55: 651-656. [ Links ]
IUCN, 2011. The IUCN Red List of Threatened Species. Version 2011.2. (Consultado: Mayo 30, 2012, http://www.iucnredlist.org). [ Links ]
Keller, A., M.O. Rödel, K.E. Linsenmair & T.U. Grafe. 2009. The importance of environmental heterogeneity for species diversity and assemblage structure in Bornean stream frogs. J. Anim. Ecol. 78: 305-314. [ Links ]
Knutson, M.G., W.B. Richardson, D.M. Reineke, B.R. Gray, J.R. Parmelee & S.E. Weick. 2004. Agricultural ponds support amphibian populations. Ecol. Appl. 14: 669-684. [ Links ]
Lajmanovich, R.C., J.C. Sánchez-Hernández, G. Stringhini & P.M. Peltzer. 2004. Levels of serum Cholinesterase activity in the Rococo toad (Bufo paracnemis) in Agrosystems of Argentina. Bull. Environ. Contam. Toxicol. 72: 586-591. [ Links ]
Lajmanovich, R.C., P.M. Peltzer, C.M. Junges, A.M. Attademo, L.C. Sanchez & A. Bassó. 2010. Activity levels of Besterases in the tadpoles of 11 species of frogs in the middle Paraná River flood plain: Implication for ecological risk assessment of soybean crops. Ecotoxicol. Environ. Saf. 73: 1517-1524. [ Links ]
Lavilla, E.O., E. Richard & G.J. Scrocchi. 2000. Categorización de los anfibios y reptiles de la República Argentina. Asociación Herpetológica Argentina, Buenos Aires, Argentina. [ Links ]
Leslie, P.H. 1945. On the use of matrices in certain population mathematics. Biometrika 35: 183-212. [ Links ]
Manly, B.F.J. 1989. A review of methods for the analysis of stage-frequency data, p. 3-69. In L.L. Mcdonald, B.F.J. Manly, J.A. Lockwood & J.A. Logan (eds.). Estimation and analysis of insect populations. Springer-Verlag, Nueva York, EE.UU. [ Links ]
Márquez García, M., M. Correa-Solis, M. Sallaberry & M.A. Méndez. 2009. Effects of pond drying on morphological and life-history traits in the anuran Rhinella spinulosa (Anura: Bufonidae). Evol. Ecol. Res. 11: 803-815. [ Links ]
Martino, A.L. 1999. Análisis estructural de una comunidad de anfibios (Barreto, Córdoba, Argentina). Tesis de Doctorado, Universidad Nacional de Río Cuarto, Río Cuarto, Córdoba, Argentina. [ Links ]
Moone, A.C. & P. Bárberi. 2008. Functional biodiversity: An agroecosystem approach. Agr. Ecosyst. Environ. 127: 7-21. [ Links ]
OMS. Organización Mundial de la Salud. 2006. Guías para la calidad del agua potable de la OMS. Vol. 1: Recomendaciones. OMS, Ginebra, Suiza. (Consultado: 15 Diciembre 2011, http://www.who.int/water_sanitation_health/dwq/es/). [ Links ]
Peltzer, P.M. 2006. La fragmentación de hábitat y su influencia en la diversidad y distribución de anfibios anuros de áreas ecotonales de los dominios fitogeográficos amazónico y chaqueño. Tesis de Doctorado, Universidad Nacional de La Plata, La Plata, Buenos Aires, Argentina. [ Links ]
Peltzer, P.M., R.C. Lajmanovich, A.M. Attademo & A.H. Beltzer. 2006. Anuran diversity across agricultural pond in Argentina. Biodivers. Conservat. 15: 3499-3513. [ Links ]
Peltzer, P.M., R.C. Lajmanovich, A.M. Attademo, M. Cabagna, G. Fiorenza, C. Junges, A. Bassó. 2007. Population and Health of Common Toads Across Agricultural Lands: Implications in Worldwide Declines. Froglog 84: 4-6. [ Links ]
Peltzer, P.M., R.C. Lajmanovich, J.C. Sánchez Hernández, M. Cabagna, A.M. Attademo & A. Bassó. 2008. Effects of agricultural pond eutrophication on survival and health status of Scinax nasicus tadpoles. Ecotoxicol. Environ. Saf. 70: 185-197. [ Links ]
Peltzer, P.M., R.C. Lajmanovich, L.C. Sanchez, A.M. Attademo, C.M. Junges, C.L. Bionda, A.L. Martino & A. Bassó. 2011. Morphological abnormalities in amphibian populations from the mid-eastern region of Argentina. Herpetol. Conserv. Biol. 6: 432-442. [ Links ]
Pengue, W.A. 2005. Transgenic crops in Argentina: the ecological and social debt. Bull. Sci. Technol. Soc. 25: 314-322. [ Links ]
Perfecto, I. & J. Vandermeer. 2008. Biodiversity Conservation in Tropical Agroecosystems: A New Conservation Paradigm. Ann. New York Acad. Sci. 1134: 173-200. [ Links ]
Perry, R.A. & D.W. Goodall. 1979. Arid land ecosystems: structure, functioning, and management. Cambridge University, Cambridge, Massacchusetts, EE.UU. [ Links ]
Rhodes, A.C.E. 2004. Marine harpacticoid copepod culture for the production of long chain highly unsaturated fatty acids and carotenoid pigments. Ph.D. Thesis, University of North Carolina, Carolina del Norte, EE.UU. [ Links ]
Rossi, R.L. 2006. Impactos recientes de la soja en la Argentina. Panorama productivo del cultivo. Agromercado 129: 4-7. [ Links ]
Schaefer, E.F. 2006. Restricciones cuantitativas asociadas con los modos reproductivos de los anfibios en áreas de impacto por la actividad arrocera en la provincia de Corrientes. Tesis de Doctorado, Universidad Nacional de La Plata, La Plata, Buenos Aires, Argentina. [ Links ]
Schmutzer, A.C., M.J. Gray, E.C. Burton & D.L. Miller. 2008. Impacts of cattle on amphibian larvae and the aquatic environment. Freshwat. Biol. 53: 2613-2625. [ Links ]
Shaffer, M.L. 1994. Demographic processes, population dynamics on heterogeneous landscapes, p. 179-205. In G.K. Meffe & C.R. Carroll (eds.). Principles of conservation biology. Sinauer, Sunderland, Massachusetts, EE.UU. [ Links ]
Sinsch, U., I.E. Di Tada & A.L. Martino. 2001. Longevity, demography and sex-specific growth of the Pampa de Achala toad, Bufo achalensis CEI, 1972. Stud. Neotrop. Fauna Environ. 36: 95-104. [ Links ]
Statsoft. 2001. Statistica for Windows, Release 6.0. Computer program manual. Statsoft, Inc. Tulsa, Oklahoma, EE.UU. [ Links ]
Stuart, S.N., J.S. Chanson, N.A. Cox, B.E. Young, A.S.L. Rodrigues, D.L. Fischman & R.W. Waller. 2004. Status and trends of amphibian declines and extinctions worldwide. Science 306: 1783-1786. [ Links ]
Taylor, B., D. Skelly, L.K. Demarchis, M.D. Slade, D. Galusha & P.M. Rabinowitz. 2005. Proximity to pollution sources and risk of amphibian limb malformation. Environ. Health Perspect. 113: 1497-1501. [ Links ]
USEPA, United States Environmental Protection Agency. 1994. National Water Quality Inventory. 1992 Report to Congress. EPA-841-R-94-001. Office of Water, Washington D.C., EE.UU. [ Links ]
Vasconcelos, T.S. & D.C. Rossa Feres. 2005. Diversidade, distribuição espacial e temporal de anfíbios anuros (Amphibia, Anura) na região noroeste do estado de São Paulo, Brasil. Biota Neotr. 5: 1-14. [ Links ]
Waichman, A.V. 1992. An alphanumeric code for toe clipping amphibians and reptiles. Herpetol. Rev. 23: 19-21. [ Links ]
Wake, D.B. 1998. Action on amphibians. Trends Ecol. Evol. 13: 379-380. [ Links ]
Wang, G., J.O. Wolff, S.H. Vessey, N.A. Slade, J.W. Witham, J.F. Merritt, M.L. Hunter Jr & S.P. Elias. 2009. Comparative population dynamics of Peromyscus leucopus in North America: influences of climate, food, and density dependence. Popul. Ecol. 51: 133-142. [ Links ]
Wells, K.D. 2007. The ecology and behavior of amphibians. Universidad de Chicago, Chicago, Illinois, EE.UU. [ Links ]
Whitfield, S.M. 2011. Enigmatic Faunal Declines at La Selva, Costa Rica: Patterns and Processes in a Collapsing Neotropical Herpetofauna. Florida International University, Miami, Florida, EE.UU. (Consultado: 14 Mayo 2012, http://digitalcommons. fiu.edu/etd/435). [ Links ]
Wilbur, H.M. 1980. Complex life cycle. Annu. Rev. Ecol. Systemat. 11: 67-93. [ Links ]
Zar, J.H. 1996. Biostatistical Analysis. Prentice Hall, Nueva Jersey, EE.UU. [ Links ]
Altig, R., M.R. Whiles & C.L. Taylor. 2007. What do tadpoles really eat? Assessing the trophic status of an understudied and imperiled group of consumers in freshwater habitats. Freshwat. Biol. 52: 386-395. [ Links ]
Attademo, A.M., M. Cabagna Zenklusen, R.C. Lajmanovich, P.M. Peltzer, C. Junges & A. Bassó. 2011. Besterase activities and blood cell morphology in the Frog Leptodactylus chaquensis (Amphibia: Leptodactylidae) on rice agroecosystems from Santa Fe Province (Argentina). Ecotoxicology 20: 274-282. [ Links ]
Bécart, E., A. Aubry & M. Emmerson. 2007. Monitoring the conservation status of natterjack toad (Bufo calamita) in Ireland, 2004 - 2006. Irish Wildlife Manuals, No. 31. National Parks and Wildlife Service, Department of the Environment, Heritage and Local. Government, Dublín, Irlanda. [ Links ]
Beebee, T.J.C. 1996. Ecology and conservation of amphibians. Conservation biology series. Chapman & Hall, Londres, Inglaterra. [ Links ]
Beebee, T.J.C. & R.A. Griffiths. 2005. The amphibian decline crisis: A watershed for conservation biology? Biol. Conservat. 125: 271-285. [ Links ]
Begon, M., C.R. Townsend & J.L. Harper. 2006. Ecology: from individuals to ecosystems. Blackwell, Oxford, Oxforshire, Inglaterra. [ Links ]
Bionda, C.L. 2011. Dinámica poblacional de anfibios asociados a cultivos extensivos de la provincia de Córdoba, Argentina. Tesis de Doctorado, Universidad Nacional de Río Cuarto, Río Cuarto, Córdoba, Argentina. [ Links ]
Bionda, C.L., I.E. di Tada & R.C. Lajmanovich. 2011a. Composition of amphibian assemblages in agroecosystems from the central region of Argentina. Russ. J. Herpetol. 18: 93-98. [ Links ]
Bionda, C.L., R.C. Lajmanovich, N.E. Salas, A.L. Martino & I.E. di Tada. 2011b. Reproductive Ecology of the Common South American Toad Rhinella arenarum (Anura: Bufonidae): Reproductive Effort, Clutch Size, Fecundity, and Mate Selection. J. Herpetol. 45: 261-264. [ Links ]
Bionda, C.L., N. Gari, E. Luque, N.E. Salas, R.C. Lajmanovich & A.L. Martino. 2012. Ecología trófica en larvas de Rhinella arenarum (Anura: Bufonidae) en agroecosistemas y sus posibles implicaciones para la conservación. Rev. Biol. Trop. 60: 771-779. [ Links ]
Boone, M.D. & C.M. Bridges. 1999. The effect of temperature on the potency of Carbaryl for survival of tadpoles of the green frog (Rana clamitans). Environ. Toxicol. Chem. 18: 1482-1484. [ Links ]
Bridarolli, M.E. & I.E. di Tada. 1996. Algunos aspectos de la geografía física de la provincia de Córdoba, p. 15-38. In I.E. di Tada & E.H. Bucher (eds.). Biodiversidad de la provincia de Córdoba. Fauna. Universidad Nacional de Río Cuarto, Río Cuarto, Córdoba, Argentina. [ Links ]
Burton, E.C., M.J. Gray, A.C. Schmutzer & D.L. Miller. 2009. Differential responses of postmetamorphic amphibians to cattle grazing in wetlands. J. Wildl. Manag. 73: 269-277. [ Links ]
Carey, J.R. 1993. Applied demography for biologists: with special emphasis on insects. Oxford University, Nueva York, EE.UU. [ Links ]
Carey, C. & C.J. Bryant. 1995. Possible interrelations among environmental toxicants, Amphibian development, and decline of Amphibian populations. Environ. Health Perspect. 103: 13-17. [ Links ]
Cei, J.M. 1980. Amphibians of Argentina. Monit. Zool. Ital. Nueva Serie, Florencia, Italia. [ Links ]
Collins, J.P. & M. Crump. 2009. Extinction in Our Time. Oxford University, Nueva York, EE.UU. [ Links ]
Corn, P.S. 1994. Cercas en línea recta y trampas de caída, p. 104-113. In W.R. Heyer, M.A. Donnelly, R.W. McDiarmid, L.C. Hayek & M.S. Foster (eds.). Medición y Monitoreo de la diversidad biológica. Métodos estandarizados para Anfibios. Universitaria de La Patagonia, Comodoro Rivadavia, Chubut, Argentina. [ Links ]
Crump, M.L. & N.J. Scott. 1994. Técnicas estándar para inventarios y monitoreo, p. 71-136. In W.R. Heyer, M.A. Donnelly, R.W. McDiarmid, L.C. Hayek & M.S. Foster (eds.). Medición y Monitoreo de la diversidad biológica. Métodos estandarizados para Anfibios. Universitaria de La Patagonia, Comodoro Rivadavia, Chubut, Argentina. [ Links ]
Davidson, C., H.B. Shaffer & M.R. Jennings. 2002. Spatial tests of the pesticide drift, habitat destruction, UV-B, and climate-change hypotheses for California amphibian declines. Conservat. Biol. 16: 1588-1601. [ Links ]
di Tada, I.E., M.V. Zavattieri, M.E. Bridarolli, N.E. Salas & A.L. Martino. 1996. Anfibios Anuros de la provincia de Córdoba, p. 191-215. In I.E. di Tada & E.H. Bucher (eds.). Biodiversidad de la provincia de Córdoba. Fauna. Universidad Nacional de Río Cuarto, Río Cuarto, Córdoba, Argentina. [ Links ]
Doffo, N. 1989. Geomorfología del área urbana de la ciudad de Río Cuarto y de la cuenca del arroyo El Bañado, algunas consideraciones aplicadas al manejo del medio natural. Tesis de grado, Universidad Nacional de Río Cuarto, Río Cuarto, Córdoba, Argentina. [ Links ]
Duellman, W.E. & L. Trueb. 1994. Biology of amphibians. Johns Hopkins, Londres, Inglaterra. [ Links ]
Edwards, T.M., K.A. McCoy, T. Barbeau, M.W. McCoy, J. Matthew Thro & L.J. Guillette Jr. 2006. Environmental context determines nitrate toxicity in Southern toad (Bufo terrestris) tadpoles. Aquat. Toxicol. 78: 50-58. [ Links ]
Gatica, E.A., C.A. Almeida, M.A. Mallea, M.C. Del Corigliano & P. González. 2012. Water quality assessment, by statistical analysis, on rural and urban areas of Chocancharava River (Río Cuarto), Córdoba, Argentina. Environ. Monit. Assess. DOI 10.1007/ s10661-011-2495-7. [ Links ]
Guerry, A.D. & M.L. Hunter Jr. 2002. Amphibian distribution in a landscape of forest and agriculture: an examination of landscape composition and configuration. Conservat. Biol. 16: 745-754. [ Links ]
Heyer, W.R. 1974. Niche measurements of frog larvae from a seasonal tropical location in Thailand. Ecology Tempe 55: 651-656. [ Links ]
IUCN, 2011. The IUCN Red List of Threatened Species. Version 2011.2. (Consultado: Mayo 30, 2012, http://www.iucnredlist.org). [ Links ]
Keller, A., M.O. Rödel, K.E. Linsenmair & T.U. Grafe. 2009. The importance of environmental heterogeneity for species diversity and assemblage structure in Bornean stream frogs. J. Anim. Ecol. 78: 305-314. [ Links ]
Knutson, M.G., W.B. Richardson, D.M. Reineke, B.R. Gray, J.R. Parmelee & S.E. Weick. 2004. Agricultural ponds support amphibian populations. Ecol. Appl. 14: 669-684. [ Links ]
Lajmanovich, R.C., J.C. Sánchez-Hernández, G. Stringhini & P.M. Peltzer. 2004. Levels of serum Cholinesterase activity in the Rococo toad (Bufo paracnemis) in Agrosystems of Argentina. Bull. Environ. Contam. Toxicol. 72: 586-591. [ Links ]
Lajmanovich, R.C., P.M. Peltzer, C.M. Junges, A.M. Attademo, L.C. Sanchez & A. Bassó. 2010. Activity levels of Besterases in the tadpoles of 11 species of frogs in the middle Paraná River flood plain: Implication for ecological risk assessment of soybean crops. Ecotoxicol. Environ. Saf. 73: 1517-1524. [ Links ]
Lavilla, E.O., E. Richard & G.J. Scrocchi. 2000. Categorización de los anfibios y reptiles de la República Argentina. Asociación Herpetológica Argentina, Buenos Aires, Argentina. [ Links ]
Leslie, P.H. 1945. On the use of matrices in certain population mathematics. Biometrika 35: 183-212. [ Links ]
Manly, B.F.J. 1989. A review of methods for the analysis of stage-frequency data, p. 3-69. In L.L. Mcdonald, B.F.J. Manly, J.A. Lockwood & J.A. Logan (eds.). Estimation and analysis of insect populations. Springer-Verlag, Nueva York, EE.UU. [ Links ]
Márquez García, M., M. Correa-Solis, M. Sallaberry & M.A. Méndez. 2009. Effects of pond drying on morphological and life-history traits in the anuran Rhinella spinulosa (Anura: Bufonidae). Evol. Ecol. Res. 11: 803-815. [ Links ]
Martino, A.L. 1999. Análisis estructural de una comunidad de anfibios (Barreto, Córdoba, Argentina). Tesis de Doctorado, Universidad Nacional de Río Cuarto, Río Cuarto, Córdoba, Argentina. [ Links ]
Moone, A.C. & P. Bárberi. 2008. Functional biodiversity: An agroecosystem approach. Agr. Ecosyst. Environ. 127: 7-21. [ Links ]
OMS. Organización Mundial de la Salud. 2006. Guías para la calidad del agua potable de la OMS. Vol. 1: Recomendaciones. OMS, Ginebra, Suiza. (Consultado: 15 Diciembre 2011, http://www.who.int/water_sanitation_health/dwq/es/). [ Links ]
Peltzer, P.M. 2006. La fragmentación de hábitat y su influencia en la diversidad y distribución de anfibios anuros de áreas ecotonales de los dominios fitogeográficos amazónico y chaqueño. Tesis de Doctorado, Universidad Nacional de La Plata, La Plata, Buenos Aires, Argentina. [ Links ]
Peltzer, P.M., R.C. Lajmanovich, A.M. Attademo & A.H. Beltzer. 2006. Anuran diversity across agricultural pond in Argentina. Biodivers. Conservat. 15: 3499-3513. [ Links ]
Peltzer, P.M., R.C. Lajmanovich, A.M. Attademo, M. Cabagna, G. Fiorenza, C. Junges, A. Bassó. 2007. Population and Health of Common Toads Across Agricultural Lands: Implications in Worldwide Declines. Froglog 84: 4-6. [ Links ]
Peltzer, P.M., R.C. Lajmanovich, J.C. Sánchez Hernández, M. Cabagna, A.M. Attademo & A. Bassó. 2008. Effects of agricultural pond eutrophication on survival and health status of Scinax nasicus tadpoles. Ecotoxicol. Environ. Saf. 70: 185-197. [ Links ]
Peltzer, P.M., R.C. Lajmanovich, L.C. Sanchez, A.M. Attademo, C.M. Junges, C.L. Bionda, A.L. Martino & A. Bassó. 2011. Morphological abnormalities in amphibian populations from the mid-eastern region of Argentina. Herpetol. Conserv. Biol. 6: 432-442. [ Links ]
Pengue, W.A. 2005. Transgenic crops in Argentina: the ecological and social debt. Bull. Sci. Technol. Soc. 25: 314-322. [ Links ]
Perfecto, I. & J. Vandermeer. 2008. Biodiversity Conservation in Tropical Agroecosystems: A New Conservation Paradigm. Ann. New York Acad. Sci. 1134: 173-200. [ Links ]
Perry, R.A. & D.W. Goodall. 1979. Arid land ecosystems: structure, functioning, and management. Cambridge University, Cambridge, Massacchusetts, EE.UU. [ Links ]
Rhodes, A.C.E. 2004. Marine harpacticoid copepod culture for the production of long chain highly unsaturated fatty acids and carotenoid pigments. Ph.D. Thesis, University of North Carolina, Carolina del Norte, EE.UU. [ Links ]
Rossi, R.L. 2006. Impactos recientes de la soja en la Argentina. Panorama productivo del cultivo. Agromercado 129: 4-7. [ Links ]
Schaefer, E.F. 2006. Restricciones cuantitativas asociadas con los modos reproductivos de los anfibios en áreas de impacto por la actividad arrocera en la provincia de Corrientes. Tesis de Doctorado, Universidad Nacional de La Plata, La Plata, Buenos Aires, Argentina. [ Links ]
Schmutzer, A.C., M.J. Gray, E.C. Burton & D.L. Miller. 2008. Impacts of cattle on amphibian larvae and the aquatic environment. Freshwat. Biol. 53: 2613-2625. [ Links ]
Shaffer, M.L. 1994. Demographic processes, population dynamics on heterogeneous landscapes, p. 179-205. In G.K. Meffe & C.R. Carroll (eds.). Principles of conservation biology. Sinauer, Sunderland, Massachusetts, EE.UU. [ Links ]
Sinsch, U., I.E. Di Tada & A.L. Martino. 2001. Longevity, demography and sex-specific growth of the Pampa de Achala toad, Bufo achalensis CEI, 1972. Stud. Neotrop. Fauna Environ. 36: 95-104. [ Links ]
Statsoft. 2001. Statistica for Windows, Release 6.0. Computer program manual. Statsoft, Inc. Tulsa, Oklahoma, EE.UU. [ Links ]
Stuart, S.N., J.S. Chanson, N.A. Cox, B.E. Young, A.S.L. Rodrigues, D.L. Fischman & R.W. Waller. 2004. Status and trends of amphibian declines and extinctions worldwide. Science 306: 1783-1786. [ Links ]
Taylor, B., D. Skelly, L.K. Demarchis, M.D. Slade, D. Galusha & P.M. Rabinowitz. 2005. Proximity to pollution sources and risk of amphibian limb malformation. Environ. Health Perspect. 113: 1497-1501. [ Links ]
USEPA, United States Environmental Protection Agency. 1994. National Water Quality Inventory. 1992 Report to Congress. EPA-841-R-94-001. Office of Water, Washington D.C., EE.UU. [ Links ]
Vasconcelos, T.S. & D.C. Rossa Feres. 2005. Diversidade, distribuição espacial e temporal de anfíbios anuros (Amphibia, Anura) na região noroeste do estado de São Paulo, Brasil. Biota Neotr. 5: 1-14. [ Links ]
Waichman, A.V. 1992. An alphanumeric code for toe clipping amphibians and reptiles. Herpetol. Rev. 23: 19-21. [ Links ]
Wake, D.B. 1998. Action on amphibians. Trends Ecol. Evol. 13: 379-380. [ Links ]
Wang, G., J.O. Wolff, S.H. Vessey, N.A. Slade, J.W. Witham, J.F. Merritt, M.L. Hunter Jr & S.P. Elias. 2009. Comparative population dynamics of Peromyscus leucopus in North America: influences of climate, food, and density dependence. Popul. Ecol. 51: 133-142. [ Links ]
Wells, K.D. 2007. The ecology and behavior of amphibians. Universidad de Chicago, Chicago, Illinois, EE.UU. [ Links ]
Whitfield, S.M. 2011. Enigmatic Faunal Declines at La Selva, Costa Rica: Patterns and Processes in a Collapsing Neotropical Herpetofauna. Florida International University, Miami, Florida, EE.UU. (Consultado: 14 Mayo 2012, http://digitalcommons. fiu.edu/etd/435). [ Links ]
Wilbur, H.M. 1980. Complex life cycle. Annu. Rev. Ecol. Systemat. 11: 67-93. [ Links ]
Zar, J.H. 1996. Biostatistical Analysis. Prentice Hall, Nueva Jersey, EE.UU. [ Links ]
*Correspondencia para:
Clarisa Bionda. Ecología, Departamento de Ciencias Naturales, Facultad de Ciencias Exactas, Físico-Químicas y Naturales, UNRC, ruta 36km 601, Río Cuarto, Córdoba, Argentina; Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET), Argentina; cbionda@exa.unrc.edu.ar
Rafael Lajmanovich. Ecotoxicología, Escuela Superior de Sanidad “Dr. Ramón Carrillo”, Facultad de Bioquímica y Ciencias Biológicas, Paraje “El Pozo” s/n, Santa Fe, Argentina; Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET), Argentina; lajmanovich@hotmail.com
Nancy Salas. Ecología, Departamento de Ciencias Naturales, Facultad de Ciencias Exactas, Físico-Químicas y Naturales, UNRC, ruta 36km 601, Río Cuarto, Córdoba, Argentina; nsalas@exa.unrc.edu.ar
Adolfo Martino. Ecología, Departamento de Ciencias Naturales, Facultad de Ciencias Exactas, Físico-Químicas y Naturales, UNRC, ruta 36km 601, Río Cuarto, Córdoba, Argentina; amartino@exa.unrc.edu.ar
Ismael di Tada. Ecología, Departamento de Ciencias Naturales, Facultad de Ciencias Exactas, Físico-Químicas y Naturales, UNRC, ruta 36km 601, Río Cuarto, Córdoba, Argentina; iditada@gmail.com
1. Ecología, Departamento de Ciencias Naturales, Facultad de Ciencias Exactas, Físico-Químicas y Naturales, UNRC, ruta 36km 601, Río Cuarto, Córdoba, Argentina; cbionda@exa.unrc.edu.ar, nsalas@exa.unrc.edu.ar, amartino@exa.unrc.edu.ar, iditada@gmail.com
2. Ecotoxicología, Escuela Superior de Sanidad “Dr. Ramón Carrillo”, Facultad de Bioquímica y Ciencias Biológicas, Paraje “El Pozo” s/n, Santa Fe, Argentina; lajmanovich@hotmail.com
3. Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET), Argentina
Recibido 22-III-2012. Corregido 10-X-2012. Aceptado 12-XI-2012.


{kind=link}
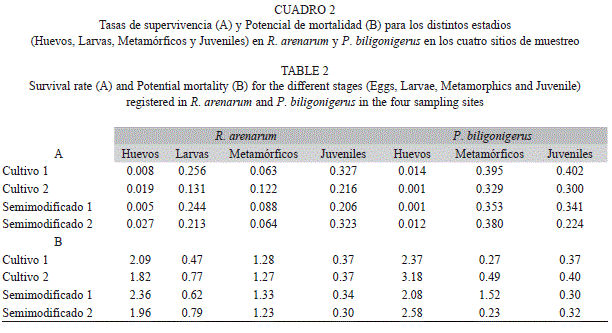{kind=link}
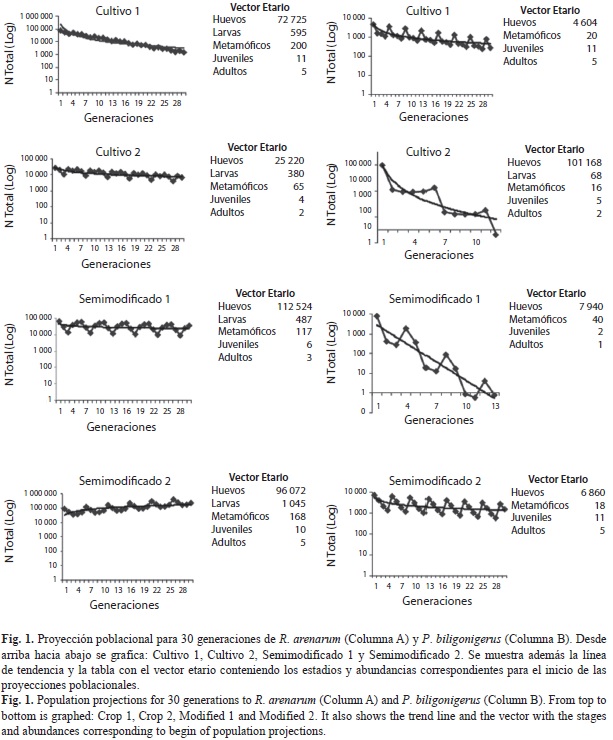{kind=link}