










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.61 n.3 San José Sep. 2013
Reproduction of Blackfin tuna Thunnus atlanticus (Perciformes: Scombridae) in Saint Peter and Saint Paul Archipelago, Equatorial Atlantic, Brazil
Reproducción del atún de aleta negra Thunnus atlanticus (Perciformes: Scombridae) en el Archipiélago San Pedro y San Pablo, Atlántico Ecuatorial, Brasil
Reproducción del atún de aleta negra Thunnus atlanticus (Perciformes: Scombridae) en el Archipiélago San Pedro y San Pablo, Atlántico Ecuatorial, Brasil
Natalia P.A. Bezerra1*, Cezar A.F. Fernandes1, Fernanda V. Albuquerque1, Vanessa Pedrosa1, Fábio Hazin2* & Paulo Travassos1
*Dirección para correspondencia:
Abstract
The reproduction of Blackfin tuna Thunnus atlanticus has been described for coastal regions, and for a long time, this species was considered to be a strictly continental spawner. Recently, this species was observed around a seamount habitat 500 nautical miles Northeast of Brazil, located between South America and Africa. In this study we describe the reproductive biology of Blackfin tuna at Saint Peter and Saint Paul Archipelago (SPSPA). Male and female gonads were sampled from December 2008 to July 2010, and analyzed macro and microscopically. A total of 361 fish were sampled (247 males and 114 females). Males were more common than females, with a sex ratio of 2.2 ♂:1♀. The fork length (FL) of all sampled specimens ranged from 38 to 98cm, and larger length classes were more frequent in males. It was possible to distinguish six maturity phases for females: immature, developing, spawning capable, actively spawning, regressing and recovering. Five phases were identified for males: immature, developing, spawning capable, actively spawning and recovering. The gonad index (GI) mean monthly values ranged from 6.6 (SD=4.1) to 58.4 (SD=34.7) for females, and from 2.6 (SD=1.3) to 66.2 (SD=30.4) for males. For both sexes, the largest GI values were observed at the beginning of the first semester of the year. Size at first maturity was estimated at 48cm FL and 55cm FL for females and males respectively. Approximately 80% of the specimens were adults and considered to be in reproductive conditions. Histological analysis of the ovaries and testes showed that most of the specimens were sexually mature and were reproductively active during all months of the year. However, females with mature ovaries, with large amounts of hydrated oocytes and post-ovulatory follicles, were mainly found from December to March, thus these months may constitute the main spawning season
in SPSPA. Batch fecundity varied between 272 025 and 1 140 584 oocytes for 56 and 68cm FL females respectively. Oocyte development and spawning patterns suggest a multiple spawning behavior. The results revealed that Blackfin tuna is using the SPSPA as a spawning ground, similar to other species commonly observed in the same area during the same reproductive season.
Key words: T. atlanticus, reproduction, seamounts, spawning season, gonad index (GI), reproductive cycle.
Resumen
A pesar de la importancia de las capturas del atún de aleta negra Thunnus atlanticus en el noreste de Brasil y en las cercanías del Archipiélago San Pedro y San Pablo (ASPSP), hay una escasez de información sobre su reproducción, especialmente en las islas. La reproducción del atún de aleta negra en este archipiélago se estudió de diciembre 2008 a julio 2010. Con este fin, se recolectaron 361 gónadas de hembras y machos, los machos fueron predominantemente más frecuentes en la muestra total, con una proporción sexual de 1♀:2.2 ♂. La longitud furcal todos los ejemplares muestreados varió entre 38 y 98 cm, y los machos fueron más abundantes en las clases de mayor longitud. Los valores medios mensuales del índice gonadal (IG) variaron de 6.6 (SD=4.1) a 58.4 (SD=34.7) y de 2.6 (SD=1.3) a 66.2 (SD=30.4) para hembras y machos, respectivamente. Los mayores valores de IG para ambos sexos fueron observados el inicio del primer semestre. Los análisis histológicos mostraron que la mayoría de los ovarios y testículos correspondieron a especímenes aptos para reproducirse. Sin embargo, hembras con ovarios maduros, con gran cantidad de oócitos hidratados y folículos post ovulatorios, se encontraron principalmente de diciembre a marzo, que se considera es la época de reproducción del atún de aleta negra alrededor del ASPSP.
Palabras clave: T. atlanticus, reproducción, islas oceánicas, temporada de desove, índice gonadal (IG), ciclo reproductivo.
Blackfin tuna, Thunnus atlanticus (Lesson, 1831), are present in the Western tropical Atlantic Ocean, between 40ºN and 31ºS latitude (Zavala-Camin et al. 1991), and are most common in coastal regions (Collette & Nauen 1983, Maguire et al. 2006). In Brazil, the species is caught by artisanal fishermen throughout the Northeast coast, primarily in Baía Formosa, Rio Grande do Norte State where the majority of catches are recorded, with annual landings of around 30t (Vieira et al. 2005a, Nóbrega & Lessa 2007).
Notwithstanding the species preference for coastal waters, significant catches of Blackfin tuna have been recorded in the oceanic realm in recent years off Saint Peter and Saint Paul Archipelago (SPSPA) (Bezerra et al. 2011). This species is caught along with other species traditionally fished in that area, such as Yellowfin tuna (T. albacares Bonnaterre, 1788), Wahoo (Acanthocybium solandri Cuvier, 1832), Rainbow runner (Elagatis bipinnulata Quoy e Gaimard, 1825), and Marginated flyfish (Cheilopogon cyanopterus Valenciennes, 1846) (Lubbock & Edwards 1981, Oliveira et al. 1997, Vaske Jr. et al. 2005, 2006). Excluding Yellowfin tuna, which is the most abundant species caught in that region, all other species mentioned above spawn in SPSPA surroundings (Lessa et al. 1999, Viana 2007, in prep., Duarte-Neto et al. 2009, Pinheiro et al. 2011).
These species spawn through the year and exhibit high fecundity, which is driven by high water temperatures in this equatorial oceanic zone, and may contribute to the success of multiple spawning (Hunter & Goldberg 1980, Hunter & Macewicz 1985, Travassos et al. 1999).
Despite significant catches of Blackfin tuna in Northeastern Brazil and within the vicinity of SPSPA, there is a paucity of information regarding its reproductive biology, especially in island habitats. In this context, the aim of this work was to describe the reproductive biology of the Blackfin tuna in the Saint Peter and Saint Paul Archipelago. Findings could subside the development and adoption of conservation measures needed to ensure the sustainability of exploited stocks.
Material and methods
Study site: This study was conducted from December 2008 to July 2010 in the Saint Peter and Saint Paul Archipelago located at 0º55’02” N - 29º20’42” W (Fig. 1). It is a small group of rocky islands situated about 500 nautical miles from the Brazilian coast (Campos et al. 2005). Fisheries around SPSPA are dominated by fishing boats operating trolling, handline and longline gear far from the island, mainly during the night (Vaske et al. 2006).
Sample procedures and data analyses: All gonads were obtained during scientific expeditions to SPSPA, from specimens caught by commercial fishing boats operating in the area. Fork lengths (FL) of all specimens were measured to the nearest centimeter. Differences in the length-frequency distribution of males and females were tested using a Kolmogorov-Smirnov test (p<0.05) (Zar 2010).
Gonads were fixed in situ in 10% formaldehyde and analyzed later in a laboratory. Length, width and weight of gonads from all specimens were measured, and the sex identified. Sex ratios for all months were tested using a Chi-square test (χ2, p<0.05) to identify statistically significant differences in sex proportion (Zar 2010).
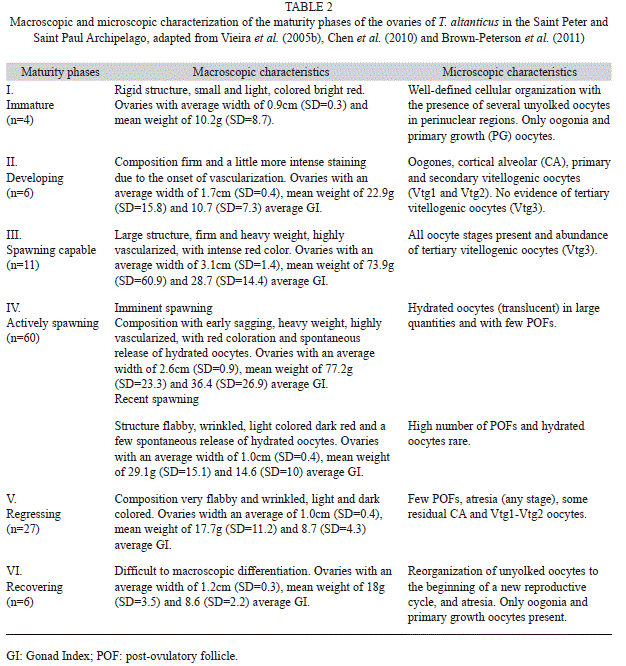
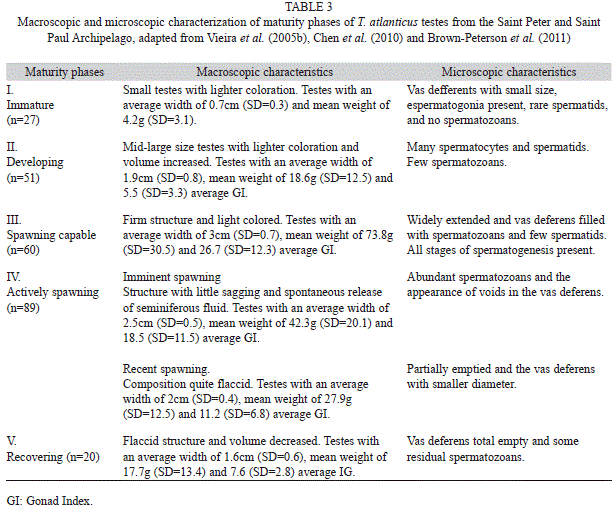
All gonads were first evaluated macroscopically for maturity stages. Then, in order to allow for histological analysis, a small section was obtained from the middle portion of each gonad, dehydrated (alcohol), cleared (xylene), embedded in paraffin, sectioned in the microtome (6μm) and finally stained (hematoxylineosin) following the method by Hunter & Macevicz (1985). Six maturity phases were established for females as follow: I-immature, II-developing, III-spawning capable, IV-actively spawning, V-regressing, VI-recovering. Five phases were identified for males: I-immature, II-developing, III-spawning capable, IV-actively spawning, V-recovering, adapted from Vieira et al. (2005b), Chen et al. (2010) and Brown- Peterson et al. (2011).
The Gonad Index (GI) of adults was calculated using the following equation from Schaefer & Orange (1956): GI=(GW*105)/FL3, where GW=gonad weight (grams) and FL=Fork length (millimeters). In order to identify the spawning season in SPSPA, the monthly mean GI was calculated for each sex separately.
Size at first maturity was estimated from a logistic curve based on relative frequency of adults in each length class, according to the formula Mf=exp [a+b*FL)/(1+exp (a+b*FL)], where Mf is the fraction of adult specimens. This method was fitted using maximum likelihood with Statistica 7 software (Zar 2010).
To estimate batch fecundity, we applied the equation: FB=nWg/w, where FB=total number of hydrated oocytes in ovaries, n=number of hydrated oocytes in the aliquot, Wg=weight of both ovaries and w=weight of ovary aliquot; this was adapted from Hunter et al. (1985). For this purpose, aliquots of 0.5g were obtained from the middle portions of 10 female gonads caught from December 2008 to March 2009, which were defined as actively spawning based on the abundance of hydrated oocytes as indicated by sampled data. However, it was not possible to exclude female gonads in the actively spawning phase that showed few post-ovulatory follicles (POF) and an elevated number of hydrated oocytes.
Results
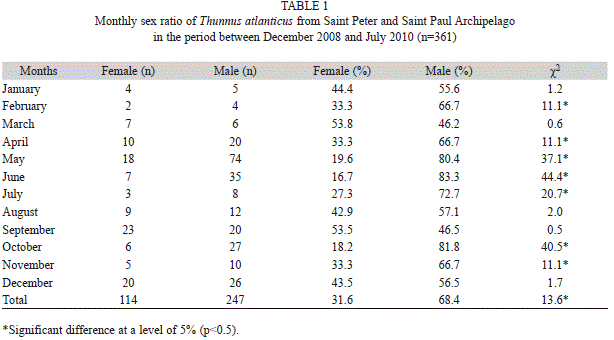
Sex ratio: A total of 361 specimens were measured and sexed (247 males and 114 females). The sexual proportion of all individuals sampled was 1♀:2.2♂, with males being predominant in the total sample (χ2=13.6, p<0.05, df=11). The Chi-square test monthly variation was also applied for grouped years. Females were more frequent than males only in March and September. Males prevailed in all remaining months, with statistically significant differences in February, April, May, June, July, October and November (Table 1).
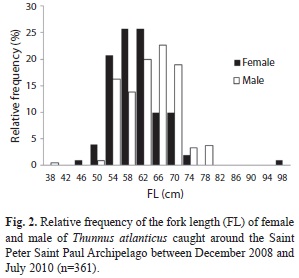
The fork length (FL) of all sampled specimens varied from 38 to 98cm. Females FL ranged from 46 to 98cm, with two modes classes: one at 58-62cm and another at 62-66cm. FL for males ranged from 38 to 78cm, with the mode at 66-70cm. There were significant differences between FL of males and females (Kolmogorov-Smirnov, p<0.05, p=0.001). Males were more frequently in larger classes (>66cm), while females predominated the smaller sizes (Fig. 2).
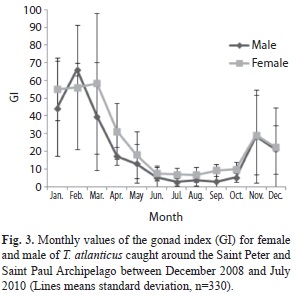
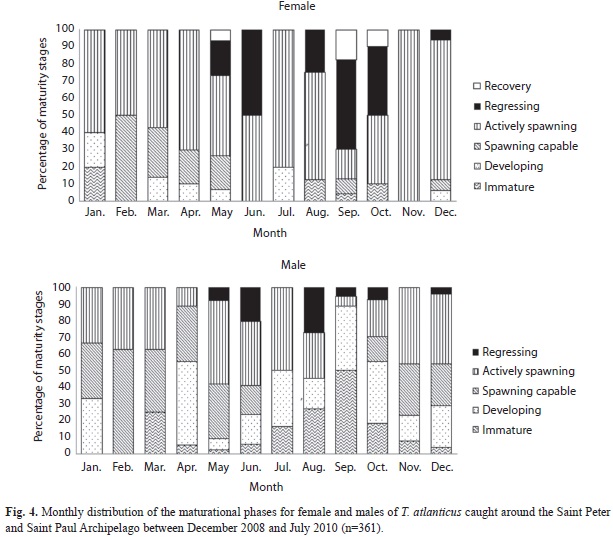
Spawning season: The gonad index (GI) average monthly values varied from 6.6 (SD=4.1) to 58.4 (SD=34.7) and from 2.6 (SD=1.3) to 66.2 (SD=30.4) for females and males, respectively. The lowest average for females was observed during August, while the highest occurred in March. High values were also found in January (mean 55.0, SD=17) and February (mean 56.2, SD=35) (Fig. 3). For males, the lowest average GI value was observed in July, while the highest values occurred in January and February. According to these results, the reproductive activity of Blackfin tuna around the SPSPA begins at the end of the second semester (November) and extends to March, with high GI values mainly in the three first months of the year.
Reproductive cycle: According to macro and microscopic analysis of the ovaries, all six maturity phases were found in females (n=114): immature (n=4; 3%), developing (n=6; 5%), spawning capable (n=11; 10%), actively spawning (n=60; 53%), regressing (n=27; 24%) and recovering (n=6; 5%). Males (n=247) were found in all five maturity phases: immature (n=27; 11%), developing (n=51; 21%), spawning capable (n=60; 24%), actively spawning (n=89; 36%) and recovering (n=20; 8%). The gonad maturation phases for both females and males are described in tables 2 and 3, respectively. According to these results, most of the ovaries and testes analyzed were from adult individuals, as 96.5% of the females and 89.0% of males showed characteristics of reproductively active specimens.
Monthly distribution of proportional female maturity phases showed few individuals with immature ovaries demonstrating spawning during all of the months, and higher reproductive activity observed in the first quarter of the year (Fig. 4).
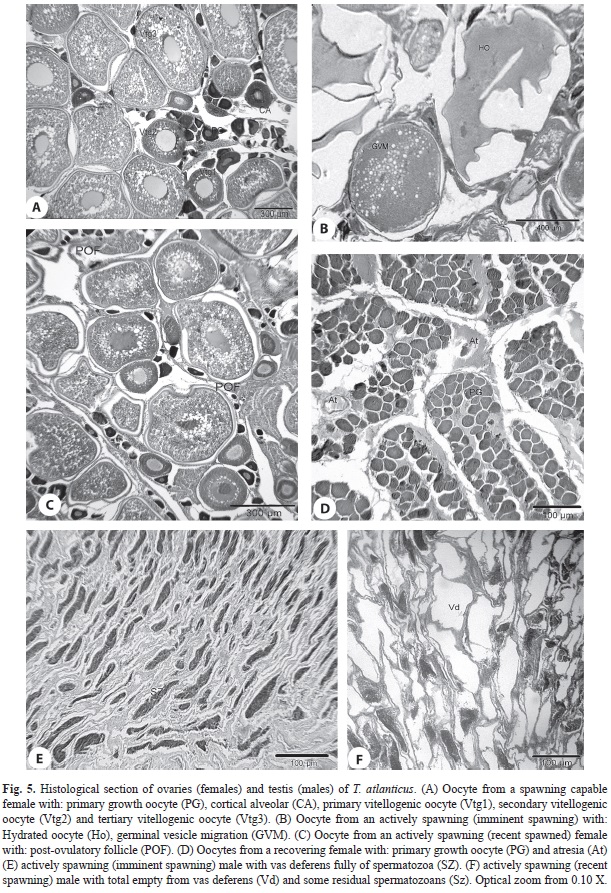
Regarding males, spawning capable specimens with their vas deferens full of sperm or actively spawning (with part of these vessels empty) were present almost all months of the year (Figs. 5 E,F). Although male individuals were more abundant throughout the period studied, testes in this maturity phase were also observed mainly during the first quarter of the year, which represented a higher reproductive activity period for Blackfin tuna in SPSPA (Fig. 4).
Higher frequency of actively spawning females, with greater amounts of hydrated oocytes and post-ovulatory follicles (Fig. 5 A,B,C), occurred between December and March, evidence of a higher reproductive activity around SPSPA during this period. Ovaries regressing and recovering, with disordered cellular organization, follicular atresia and some oogone nest, recovered rapidly for new reproductive seasons (Fig. 5D). Blackfin tuna spawn in batches around SPSPA as observed by the continuous and joint presence of hydrated oocytes and post-ovulatory follicles in the histological sections of actively spawning females, which characterizes imminent spawning (in the next 24h) and recent spawning (in the last 24h).
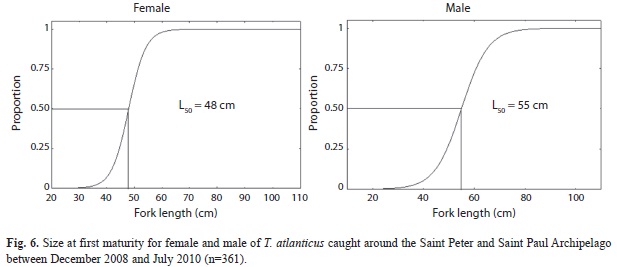
Length at first maturity (L50): The estimated length at first maturity for females and males was 48cm (CI=15) FL and 55cm (CI=15) FL, respectively (Fig. 6). Only four out of the 114 females sampled were smaller than L50 (3.5%); a total of 46 out of 247 males were smaller than L50 (18.6%).
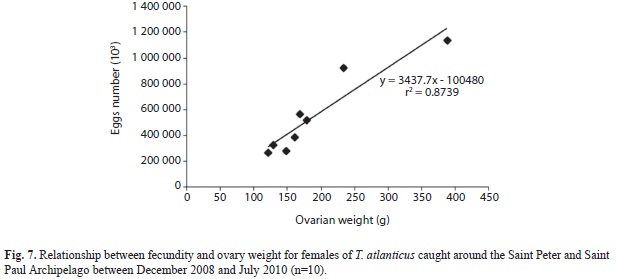
Batch fecundity: In regards to fecundity per batch, the lowest number of hydrated oocytes was 272 025, in a 56cm FL female with gonads weighing 120.90g. The highest fecundity was 1 140 584 hydrated oocytes in a 68cm FL female, with a gonads weighing 387.69g. The average fecundity per batch was 554 512 oocytes. In general, the number of hydrated oocytes was higher in samples with more gonad weight (Fig. 7). Nevertheless, the relationship between the number of hydrated oocytes and FL was not linear, due to the presence of larger individuals presenting few hydrated oocytes (98cm and 392 796 hydrated oocytes) and smaller specimens with advanced hydration (62cm and 509 030 hydrated oocytes).
Discussion
Sex ratio: The sex proportion observed was different from the male-female proportion expected for most tunas. The high natural mortality rate of females, male hostility during courtship, female predation during spawning behavior, and susceptibility to fishing gear are all factors that may have influenced male predominance in this study (Garcia Coll et al. 1984, Schaefer 1998). Vieira et al. (2005a) also observed a larger proportion of males of T. atlanticus (2.1:0.5) during a study carried out in the state of Rio Grande do Norte State. Male biased sex ratio for Blackfin tuna (1.9:1) was also reported in Northeastern Brazil (Freire et al. 2005), and similar male abundance was observed in the same region (Cruz & Paiva 1964, Cruz 1965). In Cuba, the sexual proportion of Blackfin tuna was similar to studies carried out in Brazil (Baéz-Hidalgo & Bécquer 1994). Large numbers of males were also observed in the Northern Atlantic (1.7:0.7) (Coll 1987).
In contrast, larger size tunas such as Bigeye tuna Thunnus obesus (Lowe, 1839) caught in Northeastern Brazil exhibit a greater proportion of females (1.4:1.0), which is not far from the more common relationship 1:1 (Figueiredo 2007, in prep.). Studies with Bigeye tuna and Yellowfin tuna in the Pacific Ocean reported significant differences between the sexes, and males once again predominated. This suggests that the distinction between the number of males and females may also be related to the selectivity of the several fishing gear types used to catch these individuals in different regions of the ocean (Schaeffer 2001, Zhu et al. 2010). Polygamy behavior during spawning is another hypothesis to explain the male biased sex ratio. Competition between male increases the fertility and possibility to mate with females (Pandian 2010). Higher values for GI observed for males in this study and reported by Freire et al. (2005) in Northeastern Brazil are the possible result of male competition for best reproductive performance. Sperm production increasing testes volume also indicates male domination observed by the sex ratio.
Size relationship: The size range of Blackfin tuna caught in coastal waters of Northeastern Brazil, from 36 to 89cm (Freire 2009), is very close to that found around the SPSPA, and in other studies undertaken in the North Atlantic (32 to 91cm) (Headley et al. 2009). The Rio Grande do Norte State, as in this study, showed that females were more abundant in small size classes, while males were mostly represented in the larger size classes (Vieira et al. 2005b). Females are generally larger than males among fish species, although for some tunas the inverse relationship is true (Schaefer 2001). Thus, the high energy cost of somatic and gonad growth for females and greater longevity in males may contribute to differential growth rates between sexes and possibly relates to the lower frequency of females in the upper length classes (Brill 1996, Zavala-Camin 1996).
Spawning patterns: Tunas generally migrate to waters with high temperatures (above 24ºC) to spawn. Water temperature is one of the most important environmental features for egg and larva survival, a fact observed for T. albacares, T. thynnus (Linnaeus, 1758), T. obesus and Katsuwonus pelamis (Linnaeus, 1758) (Stéquert et al. 2001, Medina et al. 2002, Mariani et al. 2010). In this study, the highest GI of Blackfin tuna was found in the first semester around the SPSPA, which coincides with the reproductive season of other pelagic fishes that also use the SPSPA as a spawning ground (Lessa et al. 1999, Viana 2007, in prep., Pinheiro et al. 2011). The variation of the gonad maturation phases distributed throughout the year also suggests successive spawning around SPSPA, possibly driven by high average annual temperatures of 27.5ºC (Travassos et al. 1999). These conditions are quite favorable to this reproductive strategy, as opposed to what occurs in temperate regions, where most fish are total spawners in the summer (Yoneda et al. 2002, Abascal et al. 2004, Ewing & Lyle 2009). Histological analyses of the ovaries of female Blackfin tuna with large amounts of hydrated oocytes and postovulatory follicles legitimizes the concept of multiple spawning and asynchronous gonad development, a characteristic common to most tropical marine fish (Wootton 1991, Hazin 1993, in prep., Goldstein et al. 2007, Chen et al. 2010). However, co-occurrence of females that are in regressing, recovering and developing phases supports the idea of rapid recovery for new spawning periods.
During the year it was also possible to note a high number of male testes with the vas deferens full of sperm and actively spawning. This supplies the high reproductive demand of active females and promotes the species’ reproductive success at the SPSPA. Testes full of sperm have also been observed in K. pelamis males in Pacific waters, due to its equally rapid reproductive cycle (Hunter et al. 1986, Ashida et al. 2010).
Length at first maturity: Eighty percent of specimens analyzed were spawning capable or actively spawning, considering the size at first maturity (L50) for females (48cm) and males (55cm). Freire et al. (2005) reported first size of capture around 58.1cm FL in Northeastern Brazil which is above the size at first maturity for the species. In studies carried out with Blackfin tuna in Baía Formosa (RN), the estimated L50 for females was 51cm total length, but showed no estimation for males (Vieira et al. 2005). Freire (2009) estimated the average size at first maturation for the same species at 49.2 and 51.3cm FL for females and males respectively; these values that are relatively close to those generated by this study. Therefore, Blackfin tuna females are able to reproduce at a smaller size than males in the Southern Atlantic. Determining the size at first maturity is crucial to managing the Blackfin tuna population in this area since it may be used to establish minimum catch sizes for management purposes.
Fecundity: In this study, the average fecundity per batch was 554 512 hydrated oocytes, a much lower value than the one found for the same species off the coast of Rio Grande do Norte (an average of 1 451 841 hydrated oocytes) (Vieira et al. 2005a). However, in this study the estimate for fecundity per batch may be biased due to inclusion of ovaries containing hydrated oocytes or with few postovulatory follicles, a result of sampling procedures that were limited by fishing time periods in the spawning grounds during spawning behavior. This difficulty was also reported by Hunter et al. (1986) and Schaefer (1996) for tunas, resulting in lower estimates of fecundity for larger individuals. The great amount of energy used by Blackfin tuna to grow and, especially, to migrate (because of its predominantly coastal geographic distribution, see Bezerra et al. 2011) might be related to the lower fecundity at the Saint Peter and Saint Paul Archipelago.
Nevertheless, further studies are necessary to elucidate the Blackfin tuna migratory movements within the tropical Atlantic Ocean, especially regarding the specimens that use the SPSPA as spawning grounds and possibly as a feeding area.
Acknowledgments
We thank Fundação de Amparo à Ciência e Tecnologia do Estado de Pernambuco (FACEPE) for granting a scholarship during the study period; Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) and the PRÓARQUIPÉLAGO/SECIRM program (Secretaria da Comissão Interministerial para os Recursos do Mar) for logistical support during the expeditions to the SPSPA; and all of the fishermen and researchers that contributedto this study.
References
Abascal, F., C. Megina & A. Medina. 2004. Testicular development in migrant and spawning bluefin tuna (Thunnus thynnus (L.)) from the eastern Atlantic and Mediterranean. Fish. Bull. 102: 407-417. [ Links ]
Ashida, H., T. Tanabe, K. Satoh, A. Fukui, S. Tanaka & N. Suzuki. 2010. Reproductive biology of male skipjack tuna Katsuwonus pelamis (Linnaeus) in the tropical Western and central Pacific Ocean. Fish. Sci. 76: 785-793. [ Links ]
Baez-Hidalgo, M. & U. Bécquer. 1994. Fecundidade del bonito Katsuwonus pelamis (Linnaeus) y la albacora Thunnus atlanticus (Lesson) en Cuba. Rev. Invest. Mar. 15: 218-222. [ Links ]
Bezerra, N.P.A., P. Travassos, F.H.V. Hazin, D.L. Viana & B.C.L. Macena. 2011. Occurrence of blackfin tuna Thunnus atlanticus, Lesson 1931 (Scombridae) in Saint Peter and Saint Paul Archipelago, Brazil. Panamjas 6: 68-70. [ Links ]
Brill, R.W. 1996. Selective advantages conferred by the high performance physiology of tunas, billfishes, and dolphin fish. Comp. Biochem. Physiol. 113: 3-15. [ Links ]
Brown-Peterson, N.J., D.M. Wyanski, F. Saborido-Rey, B.J. Macewicz & S.K. Lowerre-Barbieri. 2011. A standardized terminology for describing reproductive development in fishes. Mar. Coast. Fish. 3: 52-70. [ Links ]
Campos, T.F.C., J. Virgens-Neto, N.K. Srivastava, R.A. Petta, L.A. Artmann, J.F.S. Moraes, L. Mendes & S. Silveira. 2005. Arquipélago de São Pedro e São Paulo-Soerguimento tectônico de rochas infracrustais no Oceano Atlântico. Sítios geológicos e paleontológicos do Brasil, Brasília, Brazil. (Downloaded: March 01, 2012, www.unb.br/ig/sigep/sitio002/sitio002.pdf). [ Links ]
Chen, K.S., P.R. Crone & C. Hsu. 2010. Reproductive biology of albacore Thunnus alalunga. J. Fish. Biol. 77: 119-136. [ Links ]
Coll, I. 1987. Composiciones por largo de bonito (Katsuwonus pelamis) y albacora (Thunnus atlanticus) en dos regiones de Cuba. Rev. Invest. Mar. 8: 69-81. [ Links ]
Collette, B. & C. Nauen. 1983. FAO species catalogue Scombrids of the world. An annotated and illustrated catalogue tunas, mackerels, bonitos and related species known to date. FAO Fish Synop. 2: 85. [ Links ]
Cruz, J.F. & M.P. Paiva. 1964. Sobre a biologia pesqueira da albacora, Thunnus atlanticus (Lesson), no Nordeste do Brasil. Bol. Inst. Biol. Mar. Univ. R. G. Norte 1: 1-15. [ Links ]
Cruz, J.F. 1965. Contribuição ao estudo da biologia pesqueira da albacorinha Thunnus atlanticus (Lesson), no Nordeste do Brasil. Bol. Inst. Biol. Mar. Univ. R. G. Norte 2: 33 40. [ Links ]
Duarte-Neto, P., R. Lessa, P. Travassos & F. Hazin . 2009. Thunnus albacares, p. 263-274. In R. Lessa, M. Nóbrega & J.L. Bezerra Jr. (eds.). Dinâmica de poulações e avaliação dos estoques dos recursos pesqueiros da região Nordeste (Programa REVIZEEScore Nordeste). Martins & Cordeiro, Fortaleza, Ceará, Brazil. [ Links ]
Ewing, G. & M. Lyle. 2009. Reproductive dynamics of redbait, Emmelichthys nitidus (Emmelichthyidae), from South-Eastern Australia. Fish. Res. 97: 206-215. [ Links ]
Freire, K.M.F., R. Lessa & J.E. Lins-Oliveira. 2005. Fishery and biology of blackfin tuna Thunnus atlanticus off Northeastern Brazil. Gulf Caribb. Res. 17: 15-24. [ Links ]
Freire, K. 2009. Thunnus atlanticus, p. 275-286. In R. Lessa, M. Nóbrega & J.L. Bezerra Jr. (eds.). Dinâmica de poulações e avaliação dos estoques dos recursos pesqueiros da região Nordeste (Programa REVIZEE- Score Nordeste). Martins & Cordeiro, Fortaleza, Ceará, Brazil. [ Links ]
Garcia-Coll, I., L.S. Alvarez-Lajonchere & J.I. Noyola-Ugalde. 1984. Determinacion de la edad y el crecimiento del bonito Katsuwonus pelamis (Linnaeus) y la albacora Thunnus atlanticus (Lesson) en la región suroccidental de Cuba en el año 1979. Rev. Invest. Mar. 5: 95-126. [ Links ]
Goldstein, J., S. Heppell, A. Cooper, S. Brault & M. Lutcavage. 2007. Reproductive status and body condition of Atlantic bluefin tuna in the Gulf of Maine, 2000- 2002. Mar. Biol. 151: 2063-2075. [ Links ]
Headley, M., H. Oxenford, M. Peterson & P. Fanning. 2009. Size related variability in the summer diet of the blackfin tuna (Thunnus atlanticus Lesson, 1831) from Tobago, the Lesser Antilles. J. Appl. Ichthyol. 25: 669-675. [ Links ]
Hunter, J. & S. Goldberg. 1980. Spawning incidence and batch fecundity in northern anchovy, Engraulis mordax. Fish. Bull. 77: 641-652. [ Links ]
Hunter, J., N.C.H. Lo & R.J.H. Leong. 1985. Batch fecundity in multiple spawning fishes, p. 67-77. In R. Lasker (ed.). An egg production method for estimating spawning biomass of pelagic fish: application to the northern anchovy, Engraulis mordax. NOAA Tech. Rep. NMFS. La Jolla, California, USA. [ Links ]
Hunter, J. & B.J. Macewicz. 1985. Measurement of spawning frequency and bath fecundity in multiple spawning fishes, p. 79-94. In R. Lasker (ed.). An egg production method for estimating spawning biomass of pelagic fish: application to the Northern anchovy, Engraulis mordax. NOAA Tech. Rep. NMFS. La Jolla, California, USA. [ Links ]
Hunter, J., B. Macewicz & J. Sibert. 1986. The spawning frequency of skipjack tuna, Katsuwonus pelamis, from the South Pacific. Fish. Bull. 84: 895-903. [ Links ]
Lessa, R., P. Mafalda Jr., R. Advíncula, R. Lucchesi, J. Bezerra Jr., T. Vaske-Jr & D. Hellebrandt. 1999. Distribution and abundance of ichtryoneuston at seamounts and islands off North-Eastern Brazil. Arch. Fish. Mar. Res. 47: 239-252. [ Links ]
Lubbock, R. & A. Edwards. 1981. The fishes of Saint Paul’s Rocks. J. Fish. Biol. 18: 135-157. [ Links ]
Maguire, J., M. Sissenwine, J. Csirke, R. Grainger & S. Garcia. 2006. The state of world highly migratory, straddling and other high seas fishery resources and associated species. FAO Fish. Tech. Pap. 495: 84. [ Links ]
Mariani, P., B.R. Mackenzie, D. Iudicone & A. Bozec. 2010. Modelling retention and dispersion mechanisms of bluefin tuna eggs and larvae in the northwest Mediterranean Sea. Prog. Oceanogr. 86: 45-58. [ Links ]
Medina, A., F. Abascal, C. Megina & A. Garcia. 2002. Stereological assessment of the reproductive status of female Atlantic northern bluefin tuna during migration to Mediterranean spawning grounds through the Strait of Gibraltar. J. Fish. Biol. 60: 203-217. [ Links ]
Nóbrega, M. & R. Lessa. 2007. Descrição e composição das capturas da frota pesqueira artesanal da região nordeste do Brasil. Arq. Cienc. Mar. 40: 64-74. [ Links ]
Oliveira, G., J. Evangelista & B. Ferreira. 1997. Considerações sobre a biologia e a pesca no Arquipélago de São Pedro e São Paulo. Bol. téc. cient. CEPENE 5: 1-16. [ Links ]
Pandian, T.J. 2010. Sexuality in fishes. Science, Einfield, New Hampshire, USA. [ Links ]
Pinheiro, P., F. Hazin, P. Travassos, P. Oliveira, F. Carvalho & M. Rêgo. 2011. The reproductive biology of the rainbow runner, Elagatis bipinnulata (Quoy & Gaimard, 1825) caught in the São Pedro and São Paulo Archipelago. Braz. J. Biol. 71: 99-106. [ Links ]
Schaefer, K. 1996. Spawning time, frequency, and batch fecundity of yellowfin tuna, Thunnus albacares, near clipperton Atoll in the eastern Pacfic Ocean. Fish. Bull. 94: 98-112. [ Links ]
Schaefer, K. 1998. Reproductive biology of yellowfin tuna, Thunnus albacares, in the eastern Pacific tuna. Inter- Am. Trop. Tuna Comm. Bull. 21: 205-272. [ Links ]
Schaefer, K. 2001. Reproductive biology of tunas, p. 225- 270. In B. Block & E. Stevens (eds.). Fish Physiology, Ecology, and Evolution. Academic, San Diego, California, USA. [ Links ]
Stéquert, B., J.N. Rodriguez, B. Cuisset & F. Menn. 2001. Gonadosomatic index and seasonal variations of plasma sex steroids in skipjack tuna (Katsuwonus pelamis) and yellowfin tuna (Thunnus albacares) from the western Indian Ocean. Aquat. Living. Resour. 14: 313-318. [ Links ]
Travassos, P., F. Hazin, J. Zagaglia, R. Advíncula & J. Schober. 1999. Thermohaline structure around seamounts and island off North- Eastern Brazil. Arch. Fish. Mar. Res. 47: 211-222. [ Links ]
Vaske-Jr, T., R. Lessa, M. Nóbrega, S. Montealegre- Quijano, F. Marcante-Santana & J. Bezerra Jr. 2005. A checklist of fishes from Saint Peter and Saint Paul Archipelago. J. Appl. Ichthyol. 21: 75-79. [ Links ]
Vaske-Jr, T., R. Lessa, A. Ribeiro, M. Nóbrega, A. Pereira & C. Andrade. 2006. A pesca comercial de peixes pelágicos no arquipélago de São Pedro e São Paulo, Brasil. Trop. Ocean. 34: 31-41. [ Links ]
Vieira, K., J.L. Oliveira, M.C. Barbalho & J. Aldatz. 2005a. Aspects of the dynamic population of blackfin tuna (Thunnus atlanticus-Lesson, 1831) caught in the Northeast Brazil. Collect. Vol. Sci. Pap. ICCAT. 58: 1623-1628. [ Links ]
Vieira, K., J.L. Oliveira, M.C. Barbalho & J. Garcia Jr. 2005b. Reproductive characteristics of blackfin tuna Thunnus atlanticus (Lesson, 1831), in Northeast Brazil. Collect. Vol. Sci. Pap. ICCAT 58: 1629-1634. [ Links ]
Wootton, R.J. 1991. Ecology of teleosts fishes. Chapman and Hall, London, England. [ Links ]
Yoneda, M., K. Futagawa, M. Tokimura, H. Horikawa, S. Matsuura & M. Matsuyama. 2002. Reproductive cycle, spawning frequency and batch fecundity of the female whitefin jack Kaiwarinus equula in the East China Sea. Fish. Res. 57: 297-309. [ Links ]
Zar, J. 2010. Biostatistical analysis. Prentice-Hall, Englewood Cliffs, New Jersey, USA. [ Links ]
Zavala-Camin, L.A., R.T.B. Grassi, R.W. Seckendorff & G.G. Tiago. 1991. Ocorrência de recursos pesqueiros epipelágicos na posição 22º11’ S - 39º55’ W, Brasil. B. Inst. Pesca 18: 13-21. [ Links ]
Zavala-Camin, L.A. 1996. Introdução aos estudos sobre alimentação natural em peixes. EDUEM, Maringá, Paraná, Brazil. [ Links ]
Zhu, G., X. Dai, L. Xu & Y. Zhou. 2010. Reproductive biology of bigeye tuna, Thunnus obesus, (Scombridae) in the eastern and central tropical Pacific Ocean. Environ. Biol. Fish. 88: 253-260. [ Links ]
Ashida, H., T. Tanabe, K. Satoh, A. Fukui, S. Tanaka & N. Suzuki. 2010. Reproductive biology of male skipjack tuna Katsuwonus pelamis (Linnaeus) in the tropical Western and central Pacific Ocean. Fish. Sci. 76: 785-793. [ Links ]
Baez-Hidalgo, M. & U. Bécquer. 1994. Fecundidade del bonito Katsuwonus pelamis (Linnaeus) y la albacora Thunnus atlanticus (Lesson) en Cuba. Rev. Invest. Mar. 15: 218-222. [ Links ]
Bezerra, N.P.A., P. Travassos, F.H.V. Hazin, D.L. Viana & B.C.L. Macena. 2011. Occurrence of blackfin tuna Thunnus atlanticus, Lesson 1931 (Scombridae) in Saint Peter and Saint Paul Archipelago, Brazil. Panamjas 6: 68-70. [ Links ]
Brill, R.W. 1996. Selective advantages conferred by the high performance physiology of tunas, billfishes, and dolphin fish. Comp. Biochem. Physiol. 113: 3-15. [ Links ]
Brown-Peterson, N.J., D.M. Wyanski, F. Saborido-Rey, B.J. Macewicz & S.K. Lowerre-Barbieri. 2011. A standardized terminology for describing reproductive development in fishes. Mar. Coast. Fish. 3: 52-70. [ Links ]
Campos, T.F.C., J. Virgens-Neto, N.K. Srivastava, R.A. Petta, L.A. Artmann, J.F.S. Moraes, L. Mendes & S. Silveira. 2005. Arquipélago de São Pedro e São Paulo-Soerguimento tectônico de rochas infracrustais no Oceano Atlântico. Sítios geológicos e paleontológicos do Brasil, Brasília, Brazil. (Downloaded: March 01, 2012, www.unb.br/ig/sigep/sitio002/sitio002.pdf). [ Links ]
Chen, K.S., P.R. Crone & C. Hsu. 2010. Reproductive biology of albacore Thunnus alalunga. J. Fish. Biol. 77: 119-136. [ Links ]
Coll, I. 1987. Composiciones por largo de bonito (Katsuwonus pelamis) y albacora (Thunnus atlanticus) en dos regiones de Cuba. Rev. Invest. Mar. 8: 69-81. [ Links ]
Collette, B. & C. Nauen. 1983. FAO species catalogue Scombrids of the world. An annotated and illustrated catalogue tunas, mackerels, bonitos and related species known to date. FAO Fish Synop. 2: 85. [ Links ]
Cruz, J.F. & M.P. Paiva. 1964. Sobre a biologia pesqueira da albacora, Thunnus atlanticus (Lesson), no Nordeste do Brasil. Bol. Inst. Biol. Mar. Univ. R. G. Norte 1: 1-15. [ Links ]
Cruz, J.F. 1965. Contribuição ao estudo da biologia pesqueira da albacorinha Thunnus atlanticus (Lesson), no Nordeste do Brasil. Bol. Inst. Biol. Mar. Univ. R. G. Norte 2: 33 40. [ Links ]
Duarte-Neto, P., R. Lessa, P. Travassos & F. Hazin . 2009. Thunnus albacares, p. 263-274. In R. Lessa, M. Nóbrega & J.L. Bezerra Jr. (eds.). Dinâmica de poulações e avaliação dos estoques dos recursos pesqueiros da região Nordeste (Programa REVIZEEScore Nordeste). Martins & Cordeiro, Fortaleza, Ceará, Brazil. [ Links ]
Ewing, G. & M. Lyle. 2009. Reproductive dynamics of redbait, Emmelichthys nitidus (Emmelichthyidae), from South-Eastern Australia. Fish. Res. 97: 206-215. [ Links ]
Freire, K.M.F., R. Lessa & J.E. Lins-Oliveira. 2005. Fishery and biology of blackfin tuna Thunnus atlanticus off Northeastern Brazil. Gulf Caribb. Res. 17: 15-24. [ Links ]
Freire, K. 2009. Thunnus atlanticus, p. 275-286. In R. Lessa, M. Nóbrega & J.L. Bezerra Jr. (eds.). Dinâmica de poulações e avaliação dos estoques dos recursos pesqueiros da região Nordeste (Programa REVIZEE- Score Nordeste). Martins & Cordeiro, Fortaleza, Ceará, Brazil. [ Links ]
Garcia-Coll, I., L.S. Alvarez-Lajonchere & J.I. Noyola-Ugalde. 1984. Determinacion de la edad y el crecimiento del bonito Katsuwonus pelamis (Linnaeus) y la albacora Thunnus atlanticus (Lesson) en la región suroccidental de Cuba en el año 1979. Rev. Invest. Mar. 5: 95-126. [ Links ]
Goldstein, J., S. Heppell, A. Cooper, S. Brault & M. Lutcavage. 2007. Reproductive status and body condition of Atlantic bluefin tuna in the Gulf of Maine, 2000- 2002. Mar. Biol. 151: 2063-2075. [ Links ]
Headley, M., H. Oxenford, M. Peterson & P. Fanning. 2009. Size related variability in the summer diet of the blackfin tuna (Thunnus atlanticus Lesson, 1831) from Tobago, the Lesser Antilles. J. Appl. Ichthyol. 25: 669-675. [ Links ]
Hunter, J. & S. Goldberg. 1980. Spawning incidence and batch fecundity in northern anchovy, Engraulis mordax. Fish. Bull. 77: 641-652. [ Links ]
Hunter, J., N.C.H. Lo & R.J.H. Leong. 1985. Batch fecundity in multiple spawning fishes, p. 67-77. In R. Lasker (ed.). An egg production method for estimating spawning biomass of pelagic fish: application to the northern anchovy, Engraulis mordax. NOAA Tech. Rep. NMFS. La Jolla, California, USA. [ Links ]
Hunter, J. & B.J. Macewicz. 1985. Measurement of spawning frequency and bath fecundity in multiple spawning fishes, p. 79-94. In R. Lasker (ed.). An egg production method for estimating spawning biomass of pelagic fish: application to the Northern anchovy, Engraulis mordax. NOAA Tech. Rep. NMFS. La Jolla, California, USA. [ Links ]
Hunter, J., B. Macewicz & J. Sibert. 1986. The spawning frequency of skipjack tuna, Katsuwonus pelamis, from the South Pacific. Fish. Bull. 84: 895-903. [ Links ]
Lessa, R., P. Mafalda Jr., R. Advíncula, R. Lucchesi, J. Bezerra Jr., T. Vaske-Jr & D. Hellebrandt. 1999. Distribution and abundance of ichtryoneuston at seamounts and islands off North-Eastern Brazil. Arch. Fish. Mar. Res. 47: 239-252. [ Links ]
Lubbock, R. & A. Edwards. 1981. The fishes of Saint Paul’s Rocks. J. Fish. Biol. 18: 135-157. [ Links ]
Maguire, J., M. Sissenwine, J. Csirke, R. Grainger & S. Garcia. 2006. The state of world highly migratory, straddling and other high seas fishery resources and associated species. FAO Fish. Tech. Pap. 495: 84. [ Links ]
Mariani, P., B.R. Mackenzie, D. Iudicone & A. Bozec. 2010. Modelling retention and dispersion mechanisms of bluefin tuna eggs and larvae in the northwest Mediterranean Sea. Prog. Oceanogr. 86: 45-58. [ Links ]
Medina, A., F. Abascal, C. Megina & A. Garcia. 2002. Stereological assessment of the reproductive status of female Atlantic northern bluefin tuna during migration to Mediterranean spawning grounds through the Strait of Gibraltar. J. Fish. Biol. 60: 203-217. [ Links ]
Nóbrega, M. & R. Lessa. 2007. Descrição e composição das capturas da frota pesqueira artesanal da região nordeste do Brasil. Arq. Cienc. Mar. 40: 64-74. [ Links ]
Oliveira, G., J. Evangelista & B. Ferreira. 1997. Considerações sobre a biologia e a pesca no Arquipélago de São Pedro e São Paulo. Bol. téc. cient. CEPENE 5: 1-16. [ Links ]
Pandian, T.J. 2010. Sexuality in fishes. Science, Einfield, New Hampshire, USA. [ Links ]
Pinheiro, P., F. Hazin, P. Travassos, P. Oliveira, F. Carvalho & M. Rêgo. 2011. The reproductive biology of the rainbow runner, Elagatis bipinnulata (Quoy & Gaimard, 1825) caught in the São Pedro and São Paulo Archipelago. Braz. J. Biol. 71: 99-106. [ Links ]
Schaefer, K. 1996. Spawning time, frequency, and batch fecundity of yellowfin tuna, Thunnus albacares, near clipperton Atoll in the eastern Pacfic Ocean. Fish. Bull. 94: 98-112. [ Links ]
Schaefer, K. 1998. Reproductive biology of yellowfin tuna, Thunnus albacares, in the eastern Pacific tuna. Inter- Am. Trop. Tuna Comm. Bull. 21: 205-272. [ Links ]
Schaefer, K. 2001. Reproductive biology of tunas, p. 225- 270. In B. Block & E. Stevens (eds.). Fish Physiology, Ecology, and Evolution. Academic, San Diego, California, USA. [ Links ]
Stéquert, B., J.N. Rodriguez, B. Cuisset & F. Menn. 2001. Gonadosomatic index and seasonal variations of plasma sex steroids in skipjack tuna (Katsuwonus pelamis) and yellowfin tuna (Thunnus albacares) from the western Indian Ocean. Aquat. Living. Resour. 14: 313-318. [ Links ]
Travassos, P., F. Hazin, J. Zagaglia, R. Advíncula & J. Schober. 1999. Thermohaline structure around seamounts and island off North- Eastern Brazil. Arch. Fish. Mar. Res. 47: 211-222. [ Links ]
Vaske-Jr, T., R. Lessa, M. Nóbrega, S. Montealegre- Quijano, F. Marcante-Santana & J. Bezerra Jr. 2005. A checklist of fishes from Saint Peter and Saint Paul Archipelago. J. Appl. Ichthyol. 21: 75-79. [ Links ]
Vaske-Jr, T., R. Lessa, A. Ribeiro, M. Nóbrega, A. Pereira & C. Andrade. 2006. A pesca comercial de peixes pelágicos no arquipélago de São Pedro e São Paulo, Brasil. Trop. Ocean. 34: 31-41. [ Links ]
Vieira, K., J.L. Oliveira, M.C. Barbalho & J. Aldatz. 2005a. Aspects of the dynamic population of blackfin tuna (Thunnus atlanticus-Lesson, 1831) caught in the Northeast Brazil. Collect. Vol. Sci. Pap. ICCAT. 58: 1623-1628. [ Links ]
Vieira, K., J.L. Oliveira, M.C. Barbalho & J. Garcia Jr. 2005b. Reproductive characteristics of blackfin tuna Thunnus atlanticus (Lesson, 1831), in Northeast Brazil. Collect. Vol. Sci. Pap. ICCAT 58: 1629-1634. [ Links ]
Wootton, R.J. 1991. Ecology of teleosts fishes. Chapman and Hall, London, England. [ Links ]
Yoneda, M., K. Futagawa, M. Tokimura, H. Horikawa, S. Matsuura & M. Matsuyama. 2002. Reproductive cycle, spawning frequency and batch fecundity of the female whitefin jack Kaiwarinus equula in the East China Sea. Fish. Res. 57: 297-309. [ Links ]
Zar, J. 2010. Biostatistical analysis. Prentice-Hall, Englewood Cliffs, New Jersey, USA. [ Links ]
Zavala-Camin, L.A., R.T.B. Grassi, R.W. Seckendorff & G.G. Tiago. 1991. Ocorrência de recursos pesqueiros epipelágicos na posição 22º11’ S - 39º55’ W, Brasil. B. Inst. Pesca 18: 13-21. [ Links ]
Zavala-Camin, L.A. 1996. Introdução aos estudos sobre alimentação natural em peixes. EDUEM, Maringá, Paraná, Brazil. [ Links ]
Zhu, G., X. Dai, L. Xu & Y. Zhou. 2010. Reproductive biology of bigeye tuna, Thunnus obesus, (Scombridae) in the eastern and central tropical Pacific Ocean. Environ. Biol. Fish. 88: 253-260. [ Links ]
*Correspondencia: Natalia P.A. Bezerra: Laboratório de Ecologia Marinha, Universidade Federal Rural de Pernambuco. Rua Dom Manuel de Medeiros, s/n. Dois Irmãos, Recife, Pernambuco, Brazil, 52171-900; natalia_ufrpe@yahoo.com.br*
Cezar A.F. Fernandes: Laboratório de Ecologia Marinha, Universidade Federal Rural de Pernambuco. Rua Dom Manuel de Medeiros, s/n. Dois Irmãos, Recife, Pernambuco, Brazil, 52171-900; cezaraff@hotmail.com
Fernanda V. Albuquerque: Laboratório de Ecologia Marinha, Universidade Federal Rural de Pernambuco. Rua Dom Manuel de Medeiros, s/n. Dois Irmãos, Recife, Pernambuco, Brazil, 52171-900; nandalbuquerque@yahoo.com.br
Vanessa Pedrosa: Laboratório de Ecologia Marinha, Universidade Federal Rural de Pernambuco. Rua Dom Manuel de Medeiros, s/n. Dois Irmãos, Recife, Pernambuco, Brazil, 52171-900; vanessacont@bol.com.br
Fábio Hazin: Laboratório de Oceanografia Pesqueira, Universidade Federal Rural de Pernambuco. Rua Dom Manuel de Medeiros, s/n. Dois Irmãos, Recife, Pernambuco, Brazil, 52171-900; fhvhazin@terra.com.br
Paulo Travassos: Laboratório de Ecologia Marinha, Universidade Federal Rural de Pernambuco. Rua Dom Manuel de Medeiros, s/n. Dois Irmãos, Recife, Pernambuco, Brazil, 52171-900; p.travassos@depaq.ufrpe.br
1. Laboratório de Ecologia Marinha, Universidade Federal Rural de Pernambuco. Rua Dom Manuel de Medeiros, s/n. Dois Irmãos, Recife, Pernambuco, Brazil, 52171-900; natalia_ufrpe@yahoo.com.br*, cezaraff@hotmail.com, nandalbuquerque@yahoo.com.br, vanessacont@bol.com.br, p.travassos@depaq.ufrpe.br
2. Laboratório de Oceanografia Pesqueira, Universidade Federal Rural de Pernambuco. Rua Dom Manuel de Medeiros, s/n. Dois Irmãos, Recife, Pernambuco, Brazil, 52171-900; fhvhazin@terra.com.br
Received 16-VII-2012. Corrected 20-X-2012. Accepted 23-XI-2012.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}