










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.61 n.3 San José Sep. 2013
Mandibular and postpharyngeal gland secretions of Acromyrmex landolti (Hymenoptera: Formicidae) as chemical cues for nestmate recognition
*Dirección para correspondencia:
Abstract
Nestmate recognition is fundamental for the cohesiveness of the colony in social insects. Acromyrmex landolti is a higher Attini, a leaf-cutter ant with largest nest and morphological castes. We studied the location of the recognition signal of A. landolti on the whole ant body, the isolation effect on the capacity of nestmaterecognition, and the role of the mandibular (MG) and post-pharyngeal glands (PPG) as putative sources for chemical recognition signals in this species. We performed behavioral bioassays and chemical analyses of MG and PPG glands; MG volatiles in different castes were also characterized. In order to determine the occurrence of nestmate recognition signals, behavioral tests were performed for which the whole body, body parts or gland extracts from a nestmate and an alien worker ant, were placed on the central axis of an active field trail. Besides, the isolation effect of the nestmate chemical signal persistence was evaluated by repeating the experiments with whole ant bodies after 12h, 24h and 48h of isolation. The agonistic behavior of the ants on the trail was classified as inspection, threatening and biting. Gland volatiles were obtained by headspace solid phase microextraction, and PPG by solid sample analyses; and chemical analyses were performed using gas chromatography-mass spectrometry. Workers performed agonistic behaviors more frequently towards non-nestmate whole bodies, body parts and gland extracts, primarily those from the post-pharyngeal glands. Nestmate thoraces odorized with alien extracts were also recognized from aliens. Nestmate recognition ability persists for 12h and 24h after isolation, but decreases after 48h, suggesting that the chemical signal changes over time. Inter-colonial chemical analyses showed no differences in the PPG hydrocarbon profiles, while the relative proportions of MG compounds showed significant differences between colonies. These results showed that nestmate recognition was achieved by MG volatiles. Possibly, the PPG secretion works as a matrix that absorbs MG volatiles. We report, for the first time, some mandibular gland compounds for the genus Acromyrmex and for the tribe Attini group.
Key words: nestmate recognition, postpharyngeal gland, mandibular gland, HS-SPME, Acromyrmex landolti, castes.
Resumen
El reconocimiento de compañeras de nido es fundamental para la cohesión de la colonia en los insectos sociales. Las señales de reconocimiento de compañeras en Acromyrmex landolti fueron estudiadas mediante bioensayos de comportamiento y análisis químicos de los componentes de la glándula mandibular (GM) y postfaríngea (GPF). Los bioensayos de comportamiento fueron realizados usando cuerpos completos de hormigas, partes del cuerpo y extractos glandulares de GM y GPF de una compañera de nido y una intrusa en el centro de una trilla activa. Los comportamientos agonísticos de las obreras se clasificaron en exploración, ataque y mordida. Las obreras se mostraron más agresivas hacia los cuerpos completos, partes del cuerpo y extractos glandulares de las intrusas que hacia los de las compañeras de nido. Los volátiles glandulares se obtuvieron mediante la técnica de espacio en cabeza con microextracción en fase sólida, y los hidrocarburos de la PPG por análisis de muestra sólida. Se encontró que los volátiles de la GM conforman la señal de reconocimiento. No se encontraron diferencias intercoloniales para los volátiles de la GM. Posiblemente los hidrocarburos cuticulares actúan como una matriz que absorbe volátiles de la GM.
Palabras clave: reconocimiento de compañeras de nido, Acromyrmex landolti, glándula mandibular, glándula postfaringeal.
Nestmate recognition is paramount for the development and maintenance of colony cohesiveness in eusocial insects. Four sources of nestmate recognition signals have been proposed: environmental odors (Jutsum et al. 1979, Obin 1986), queen odors (Carlin & Hölldobler 1986, 1987, D’Ettorre & Heinze 2005), individual odors (Whitehouse & Jaffé 1995) and gestalt odors (Soroker et al. 1994, Hernández et al. 2002, Boulay et al. 2004).
Mandibular glands (MG) of Formicidae are a source of volatile compounds, comprising mainly alcohols, ketones and aldehydes (Hölldobler & Wilson 1990). Several functions have been reported for the secretion of this gland: alarm, defense, mating and nestmate recognition signals (Blum et al. 1968, Riley et al. 1974, Hernández et al. 1999, Hughes et al. 2001, Hernández et al. 2002, Hernández et al. 2006, Van Wilgenburg et al. 2010). For the leaf-cutting ants Atta laevigata Smith 1860 and Atta cephalotes Linnaeus 1758, it has been suggested that alarm pheromones from the MG are basic components in nestmate recognition signals (Jaffe 1983, Whitehouse & Jaffe 1995, Hernández et al. 2002, 2006, Sainz-Borgo et al. 2011). Further bioassays demonstrated that the MG secretion of A. laevigata is spread all over the insect cuticule (Hernández et al. 2002). In the case of Acromyrmex landolti Forel 1885, Jaffe & Navarro (1985) have reported that nestmate recognition is also due to volatile compounds present in the head.
Along with the MG, the postpharyngeal gland (PPG) has also been proposed as a source of chemical compounds responsible for nestmate recognition, providing the gestalt odor in the ants Cataglyphis niger (Andre, 1881) (Soroker et al. 1994, 1995) and Formica rufibarbis (Fabricius, 1793) (Van Zweden et al. 2010) for example. The chemical components of the PPG are mainly long chain hydrocarbons, which are also part of the profile ofcuticular hydrocarbons (Bagnéres & Morgan 1991, Do Nascimento et al. 1993, Soroker et al. 1994, Lahav et al. 1999). Individual hydrocarbon profiles are also known to change after isolation from the colony, which in turn may change the recognition value of the chemical signal (Vander Meer et al. 1989, Lenoir et al. 2001), and need the reinforced of the another members of the colony by social stimulation to maintain the nestmate recognition signals (Boulay & Lenoir 2001).
Acromyrmex landolti is considered one of most complex Attini in regards to morphology and behavior, since it is a leaf-cutter ant with large nests and many morphologically distinct castes. In order to gain insight into the nestmate recognition mechanisms in A. landolti, we studied the location of the recognition signal in the ant’s body, the effect of isolation on the capacity of nestmate recognition, and the role of the MG and PPG as putative sources for chemical recognition signals in this species.
Materials and Methods
Insects: Worker ants used for chemical analyses were obtained from 17 colonies located throughout the campus of Universidad Simón Bolívar (USB), in Caracas city, Venezuela (10°24’ N - 66°52’ W, annual temperature range: 17-25ºC, mean annual precipitation: 1 034mm, data from the Laboratorio de Sensores Remotos of USB). Ants for the behavioral bioassays were obtained from nests located in savannas near Corporación Venezolana de Guayana (CVG)-Proforca, Monagas State, Venezuela (09°32’58” N - 64°59’18” W, mean annual temperature 26±2ºC) (De Camino & Irausqui 1977) and at the CVG Field Station in Parupa, Gran Sabana, Bolívar State,
Venezuela (5º40’ N - 61º32’ W), which features an annual temperature range of 21-25ºC and a mean annual precipitation of 3 000mm (Schubert & Huber 1989).
All ants were collected while foraging on the trail, using clean soft forceps. According to the treatments to which they were assigned, the ants were either: placed in clean glass vials for immediate use; cooled down (5min) for the dissection of body parts; isolated individually in 5mL glass vials with a piece of absorbent paper moistened with distilled water; or isolated in groups (20 ants) in a plexiglass tube (20cm long) with supply of distilled water.
Behavioral bioassays: 18 different treatments were compared through behavioral bioassays (Table 1). All bioassays were performed according to Hernández et al. (2002), with some modifications. Two experimental subjects (ants or ant parts, one nestmate N and one alien A) were fixed with an entomological pin through the thorax and placed on the central axis of an active trail, between 10 and 50cm from the nest entrance. They were separated by 3cm from each other, and the behavior of passing ants was recorded either as inspection, when the subjects or objects around them were touched with the apical zone of the antennae; threatening, when the foraging ants walked towards the subjects with open mandibles; or biting, when the subjects were grasped with the mandibles.
Each experiment was recorded using a digital video camera (Sony Model DCR-TRV 20), which was placed on a tripod at a height of 1m, approximately 50cm from the nest entrance. An infrared system (night-shot) was used for night recording. For each bioassay, a total of five minutes of interactions were recorded.
MG and PPG extracts for bioassays: MG and PPG extracts for the behavioral bioassays were obtained from glands dissected from 30 medium-size nestmate or alien workers (1.5-2.0mm head size). The glands were dissected under a stereoscopic microscope (Wild Heerbrugg) in distilled water, and extracted in 100mL of n-hexane (HPLC grade, Fisher Scientific). Each experimental ant treated with either MG or PPG gland extracts odorized by brushing with 2mL of the respective extract.
In order to assess the possible variation in the chemical composition or concentration of PPG and MG secretions by isolation from nestmates, a group of ants was kept isolated from the nest during a 48h period, and compared with ants collected immediately from the nest.
For quantification of PPG secretions internal standard technique was used in the same way as described above. Individuals were collected with clean forceps, and placed in groups of 20 workers in a clean Plexiglas tube (20cm long) for 48h, and chilled to 5ºC before dissections were made. The chemical compositions of volatile compounds from the MG of 48h isolated ants were compared with those from ants collected immediately from the nest using HSSPME.
Then the samples were desorbed in the gas chromatograph injector at 250ºC for 10s (n=10). In the case of PPG hydrocarbon, the amounts in 48h isolated ants were compared with those from ants immediately collected from the nest, extracting 20 dissected PPG as described above (n=5).
MG and PPG chemical analyses: Analyses were performed by GC using a Hewlett Packard 5890A gas chromatograph equipped with a DB-5 fused silica capillary column (30m x 0.25mm ID, Quadrex, New Haven, Connecticut), and by coupled gas chromatography-mass spectrometry (GC-MS) employing a Perkin Elmer QMass-910 attached to a GC-Autosystem 2000, equipped with a DB-5 capillarycolumn (25m x 0.18mm ID, Quadrex, New Haven, Connecticut). Injector temperature was maintained at 250ºC.
For MG analyses the oven temperature program used in the MG analyses was 50ºC for 5min followed by a linear temperature increase of 6ºC/min up to 150ºC, then another ramp of 20ºC/min up to 225ºC for 20min. Coelution with available authentic synthetic standards was carried out with four compounds, 4 metyl-3-heptanol, 3 octanone, 2-nonenone (commercially available from Aldrich, 99% pure) and 3-octanol (donated by Dr. Phillip Howse). For the quantification of MG compounds derived from two different nests, peak area percentages were calculated as described in Hughes et al. (2001).
MG and PPG volatiles for headspace solid-phase microextraction (HS-SPME) analyses were obtained from dissected worker ant heads. Twenty heads were crushed individually with a clean glass rod in a silanized 2mL glass vial, and the vial was sealed with an aluminum cap and a teflon/rubber septum (Hernández et al. 2002), and placed in a water bath thermostat at 33ºC. A SPME fiber coated with 100mm polydimethylsiloxane (PDMS) was inserted into the vial for 10 min to sample headspace volatiles. The fiber was desorbed in the gas chromatograph injection port at 250ºC for 10s. Temperature and sampling time conditions were previously optimized. MG components were identified by means of GC-MS analyses. In order to improve detectability, these analyses were carried out with vials containing from six to 20 heads, and heated in a water bath at 65ºC for 90min.
Solid sampling was used for PPG analyses.PPG were dissected and placed in a glass capillary tube, then was sealed using a lighter, and finally introduced into the GC injection port at 280ºC (Morgan 1990). The oven temperature program used was 50ºC for 5min, followed by a linear temperature increase of 4ºC/min up to 280ºC for 20min.
For the behavioural bioassays, the agonistic interactions and explorations received by the experimental ants (or ant parts) were compared by the Wilcoxon’s test. For the chemical analyses of the glands a Kruskal-Wallis ANOVA test was used to compare the relative amounts of: 1) MG volatiles and PPG hydrocarbons from ants collected immediately from the nest; 2) MG and PPG compounds from ants immediately collected from the nest with 48h isolated ants and 3) volatile compounds of thorax+gaster between two different colonies.
Results
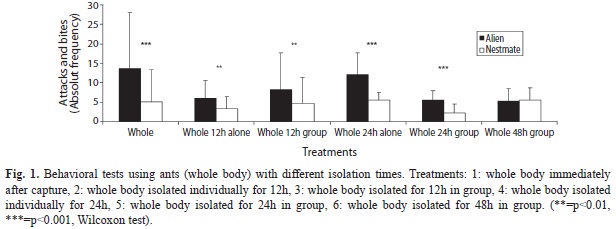
Whole body treatments: Behavioral bioassays showed that alien individuals were threatened and bitten more than nestmates, immediately upon ant collection (p<0.001) and after 12h of isolation (p<0.001). This was true also after 24h individually and 24h in group isolation (p<0.001). However, separating the ants for 48h (in a group) resulted in equal frequencies of agonistic interactions for nestmates and alien ants (p>0.9) (Fig. 1, Table 2). Although non-significant (Kruskal Wallis ANOVA p>0.05), a general decrease in the recognition ability appears to correlate with increasing isolation time (Spearman coefficient 0.85).
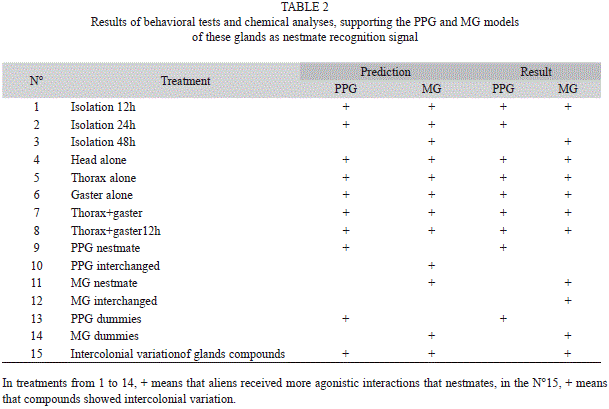
Body part treatments: Bioassays using body parts showed that alien heads (p<0.001), alien thoraces (p<0.05), alien head+thorax (p<0.05) and alien thorax+gaster (p<0.05) were threatened and bitten significantly more than the corresponding nestmate body parts (Fig. 2, Table 2). In the treatments testing only gaster and interchanged heads (p>0.05) the agonistic interactions received by alien and nestmate were not statistically significant.
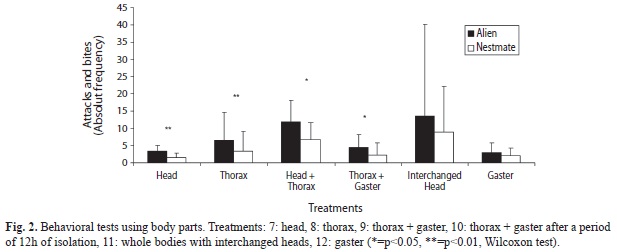
Gland extract treatments: In treatments with body parts and dummies odorized with gland extracts, both nestmate thoraces and dummies odorized with alien PPG extracts received more agonistic interactions than those odorized with nestmate PPG extracts (p=0.001, Fig. 3, Table 2). This was also the case for dummies painted with MG extracts (p=0.02, Fig. 3, Table 2), but not so for nestmate thoraces with the same treatment (p>0.05). Interestingly, in the experiments in which alien and nestmate thoraces were exchanged with regards to gland treatment, alien thoraces received more agonistic interactions than nestmate thoraces regardless of the gland extract used to odorizing them (PPG: p=0.001, MG: p=0.02, Fig. 3, Table 2).
Inspection behavior: Inspection behavior results showed statistically significant differences between aliens and nestmates in experiments using whole bodies after 24h in-group isolation (Treatment 5, Wilcoxon test, p=0.03), and in interchanged thoraces odorized with alien and nestmate MG extracts (Treatment 16, Wilcoxon test, p=0.04), that was the only treatment where nestmates were more significantly explored than aliens.
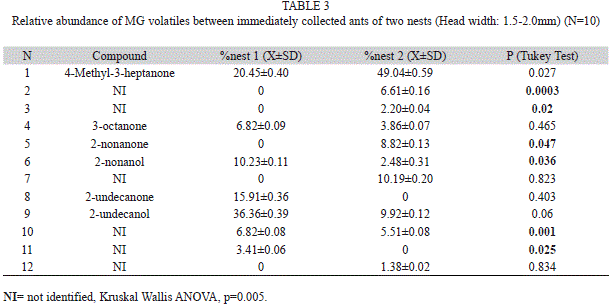
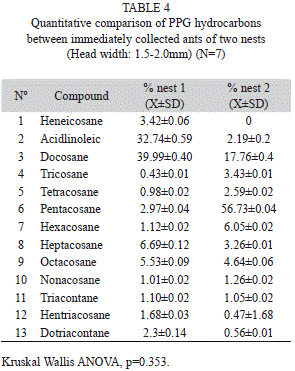
Quantification of MG and PPG compounds between two colonies: MG volatile compounds quantified by HS-SPME from ants of two different colonies were significantly different (Kruskal Wallis ANOVA, p=0.005) (Table 3). However, PPG hydrocarbons do not present intercolonial differences (Kruskal Wallis ANOVA, p=0.353). The MG volatiles and the PPG hydrocarbons do not show variation after a 48h period.
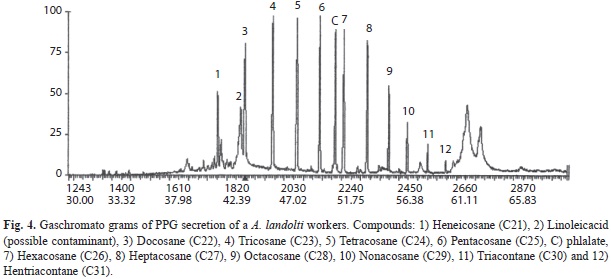
PPG hydrocarbons were identificated as (Fig. 4) (Table 4): 1) Heneicosane (C21), 2) Linoleic acid (possible contaminat), 3) Docosane (C22), 4) Tricosane (C23), 5) Tetracosane (C24), 6) Pentacosane (C25), C) phthalate 7) Hexacosane (C26), 8) Heptacosane (C27), 9) Octacosane (C28), 10) Nonacosane (C29), 11) Triacontane (C30) and 12) Hentriacontane (C31) and Dotriacontane (C32).
Quantification of MG compounds on the Thorax+Gaster: The compounds 2-nonanol and 2-undecanol derived from MG volatiles were found on the thorax+gaster. Statistically significant differences in the relative proportion of these compounds were found between two different colonies (Kruskal Wallis ANOVA, p<0.05) (Table 5).
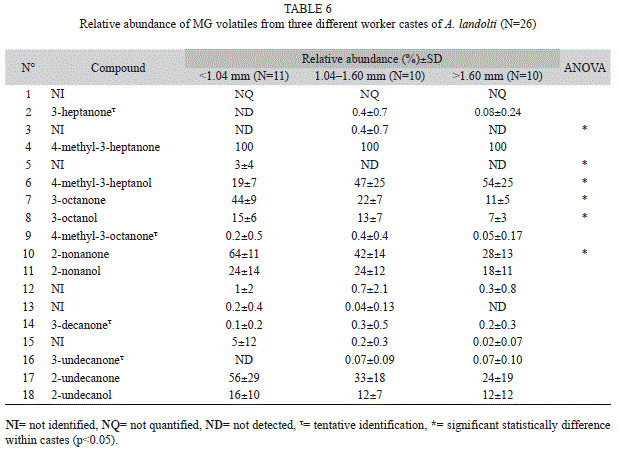
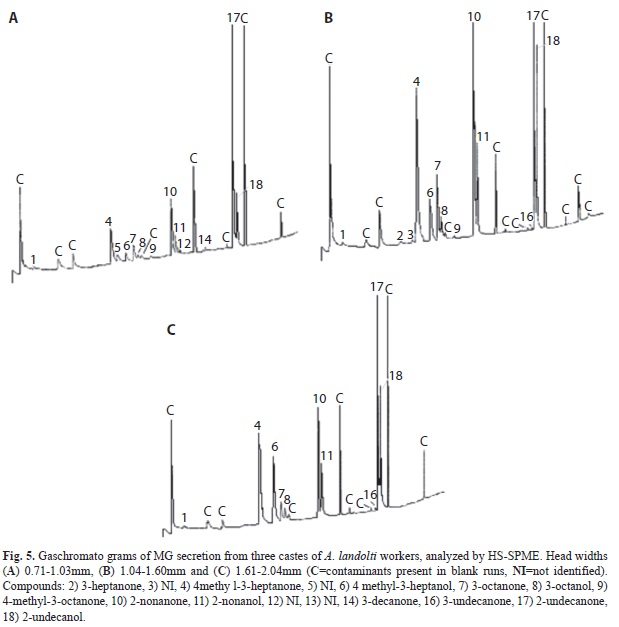
Quantification of MG compounds between castes: Six of eighteen MG volatiles
compound quantified between castes were significantly different (Kruskal Wallis ANOVA, p=0.005) (Table 6) (Fig. 5).
Discussion
Whole body treatments: Our results show that A. landolti workers are capable of recognizing their nestmates, even after individual or in-group isolation periods of 12 and 24h, indicating that recognition signals might be produced by individuals, rather than coming from the environment, fungus garden, colony or social interaction.
After being isolated from the colony for 48h, experimental subjects (foraging workers on the trail) were unable to discriminate their nestmates from aliens, showing that the signal has been modified after that isolation time from the colony. These results may suggest that the nestmate recognition signal is a volatile compound, like the mandibular gland secretions, and that then is lost during isolation time, as is the case described in Atta laevigata (Hernández et al. 2002), or perhaps by a change in the proportion of cuticular hydrocarbons in the period of isolation (Martin et al. 2012). It is also probable that the quantity of mandibular gland compounds and hydrocarbons absorbed or interchanged during the isolation period, also demonstrated by Hernandez et al. (2002) in A. laevigata, is less abundant than in the nest conditions, where hundreds of workers constantly interact with each other, while in the isolated group treatment only a few individuals are in frequent contact.
Body part treatments: Jaffé & Navarro (1985) reported that the A. landolti recognition signal is located (at least partially) in the head, as alien heads received more agonistic interactions than nestmates heads. However, we have demonstrated that they are capable of recognizing the thorax+gaster after a 12h isolation period, showing that these signals can be found all over the body for at least 12h. This result indicates that in the nestmate recognition system, at least one cue is a non-volatile one, or the cue is a volatile adsorbed by the cuticule of the thorax+gaster.
Cuticular hydrocarbons have been reported as a recognition signal, even though this is not the case with Atta laevigata (Whitehouse & Jaffe 1995), in which nestmate recognition is lost when headless ants are isolated from the colony for several hours. In our experiments, during interchanged head treatments the foraging workers did not recognize their nestmates. This suggests that these cues are mainly located in the head, even though additional signals can be also found on the thorax, because the recognition is lost in treatments where only the gaster was tested. However, in the thorax+gaster treatment, aliens were threatened more than nestmates.
The thorax+gaster received more agonistic interactions than the head alone (Treatment 7) and the thorax alone (Treatment 8).
Glands extract treatments: PPG seems to be involved in the recognition system of A. landolti, as demonstrated here by the treatment results in which thoraces were odorized with alien and nestmate PPG gland extracts. Aliens were more threatened than nestmates, contrary to the results obtained with Atta laevigata, in which only the MG extract produced alien recognition (Hernández et al. 2002). In treatments using interchanged thoraces odorized with alien and nestmate gland extracts, the alien ones received more agonistic behaviors independent of the kind of extracts, probably due to a dilution factor. Finally, in plastic dummy treatments testing PPG and MG extracts, the dummies with alien extracts were more threatened than nestmates in both cases. A different compound liberation rate from the plastic of the dummy and the cuticle is probably involved in these results.
Chemical analyses: Several of the chemical compounds identified in the MG secretions of A. landolti have been previously identified in the MG of other Attini ants from the genus Trachymyrmex and Cyphomyrmex (Crewe & Blum 1972), with the exception of the compounds 2-nonanol, 3-decanone, 3-undecanone, 2-undecanone and 2-undecanol (Table 2). Moreover, only two of these have been identified in other Acromyrmex species (A. octospinosus): 3-octanone and 3-octanol (Crewe & Blum 1972).
On the other hand, the MG secretion of A. landolti also shows differences within castes, as has been reported in other Atta species: A. sexdens rubropilosa Linnaeus 1758, A. laevigata, A. bisphaerica Forel 1908, A. capiguara Goncalves 1944, and A. cephalotes (Do Nascimento et al. 1993, Hernández et al. 1999, Hughes et al. 2001, López 2002).
Eighteen compounds were detected in MG secretion. The common presence of structurerelated compounds in these glands is known, and 3-heptanone and 4-methyl-3-heptanone have also been found in mandibular gland secretions of other Attini species: Atta sexdens, A. laevigata, A. cephalotes and Atta texana Buckley 1860 workers and in Atta sexdens virgin females (Do Nascimento et al. 1993, Hernández et al. 1999, Riley et al. 1974, Crewe & Blum 1972, Hughes et al. 2001, López 2002).
The chemical components of the PPG are mainly long chain hydrocarbons, which are also part of the profile of cuticular hydrocarbons (Bagnéres and Morgan 1991, Do Nascimento et al. 1993, Soroker et al. 1994, Lahav et al. 1999). The most of cuticular hydrocarbons are n-alkanes, monomethylalkanes, dimethylalkanes and alkenes (Martin & Drijhout 2009a), and are specie-specific, for this reason it’s important in the chemotaxonomy (Kather & Martin 2012).
Quantification of MG and PPG compounds between two colonies: The MG volatiles located in the head and in the thorax+gaster showed intercolonial variation. This fact is the main condition for a nestmate recognition cue.
The MG volatiles as nestmate recognition cues: The main function of the MG secretion is to provoke alarm behavior and to attract or repel congeners from the emission source (Morgan et al. 1978, Attygalle & Morgan 1984, Billen & Morgan 1998, Hölldobler & Wilson 1990). It is also responsible for nestmate recognition in Atta laevigata and Atta cephalotes (Hernández et al. 2002, 2006), Camponotus rufipes (Jaffé & Sánchez 1984), Camponotus atriceps (Hernández et al. 2010); and probably in Ectatomma ruidum (Jaffé & Márquez 1987) and Conomyrma sp (Jaffe et al. 1990).
In A. landolti, the paramount pieces of evidence for MG as a nestmate recognition signal were as follows: 1) Results from MG chemical analyses indicated that the A. landolti secretion shows intercolonial variation, a basic condition of a recognition signal if it is to be considered as a signal cue; 2) In the 48h isolation treatment, ants do not distinguish between nestmate and alien, indicating that recognition cues are, at least partially, a volatile compound; 3) In the interchanged head treatment, the nestmate body + alien head was more threatened and bitten than the alien body + nestmate head, indicating that the recognition signal was concentrated in the head, as was reported for A. landolti by Jaffé & Navarro (1985), 4) Nestmate thoraces odorized with MG alien extract received more agonistic interactions than the thorax odorized with MG nestmate extract; and 5) Dummies odorized with MG alien extract received more agonistic interactions than dummies odorized with nestmate MG extract.
PPG hydrocarbons as a nestmate recognition signal: The cuticular hydrocarbons have been proposed as cues for nestmate recognition in several ants, wasps, bees and termites (Blomquist et al. 1987, Singer 1998, Howard & Blomquist 2005, Martin & Drijfhout 2009a, Martin & Drijfhout 2009b). It has also been demonstrated that PPG in ants constitute a hydrocarbon reservoir for the colony (Soroker et al. 1994, Hefetz et al. 1996).
In A. landolti, we found three types of evidence for the PPG secretion functioning as recognition cues: 1) In thorax+gaster treatment (Treatment 9), aliens were more threatened and bitten than nestmates. This result indicates that the recognition signal does not volatilize in 12h; 2) The nestmate odorized with alien extract (Treatment 13) was more threatened and bitten than the thorax odorized with PPG nestmate extract; 3) Dummies odorized with PPG alien extract (Treatment 17) received more agonistic interactions than those with PPG nestmate extract. This result can be related with differences in the liberation rates of compounds of the plastic and the cuticle, being faster probably in the plastic.
These results show that the recognition system in A. landolti is colony-specific. The recognition signal is concentrated in the head, dispersed on all over the body, and involves the MG. Probably the cuticular hydrocarbons work as a matrix that absorb and release slowly MG volatiles that conformed the odor cues for nestmate recognition.
The nestmate recognition systems are different in ants. In some species the recognition cues are the cuticular hydrocarbons, as Formica japonica (Akino et al. 2004) or Cataglyphis niger (Lahav et al. 1999) but in others, like Atta laevigatta (Whitehouse & Jaffe 1995) the nestmate recognition cue is concentrated in the head; or the recognition signal is all over the body , as in O. bauri (Sainz-Borgo et al. 2011).
Acknowledgments
The authors would like to thank CVGProforca for logistical support; Beatriz Herrera-Malaver and Carmen V. Liendo-Barandiaran for the help in the chemical analyses, Solange Issa for logistical support for field trips, Andres Gonzalez for the suggestion of the manuscript, and Frances Osborn, Romano Dorta and Dan Bailey for English revision.
References
Akino, T., K. Yamamura, S. Wakamura & R. Yamaoka. 2004. Direct behavioral evidence for hydrocarbons as nestmate recognition cues in Formica japonica (Hymenoptera: Formicidae) Appl. Entomol. Zool. 39: 381-387. [ Links ]
Attygalle, A.B. & E.D. Morgan. 1984. Chemicals from the glands of ants. Chem. Soc. Rev. 13: 245-278. [ Links ]
Bagnéres, A.G. & E.D. Morgan. 1991. The postpharyngeal glands and the cuticule of Formicidae contain the same characteristic hydrocarbons. Experientia 47: 106-111. [ Links ]
Billen, J. & E.D. Morgan. 1998. Pheromone communication in social insects-sources and secretions, p. 3-33. In R.K. Vander Meer, M.D. Breed, K.E. Espelie & M.L. Winston (eds.). Pheromone Communication in Social Insects: Ants, Wasps, Bees, and Termites. Westview, Boulder, Oxford, England. [ Links ]
Blomquist, G.J., D.R. Nelson & M. De Renobales. 1987. Chemistry, biochemistry, and physiology of insect cuticular lipids. Arch. Insect Biochem. Physiol. 6: 227-265. [ Links ]
Blum, M.S., F. Padovani, H.R. Hermann & P.B. Kanowski. 1968. Chemical releasers of social behavior, XI: terpenes in the mandibular glands of Lasius umbratus. Ann. Entomol. Soc. Am. 61: 1354-1359. [ Links ]
Boulay, R. & A. Lenoir. 2001. Social isolation of mature workers affects nestmate recognition in the ant Camponotus fellah. Behav. Processes 55: 67-73. [ Links ]
Boulay, R., T. Katzav-Gozansky, A. Hefetz & A. Lenoir.2004. Odour convergence and tolerance between nestmate through trophallaxis and grooming in the ant Camponotus fellah (Dalla Torre). Insectes Soc. 51: 55-61. [ Links ]
Carlin, N.F. & B. Hölldobler. 1986. The kin recognition system of carpenter ants (Camponotus spp.). I Hierarchical cues in small colonies. Behav. Ecol. Sociobiol. 19: 123-134. [ Links ]
Carlin, N.F. & B. Hölldobler. 1987. The kin recognition systems of Carpenter ants (Camponotus spp). II. Larger colonies. Behav. Ecol. Sociobiol. 20: 209-217. [ Links ]
Crewe, R.M. & M.S. Blum. 1972. Alarm pheromones of the Attini: their phylogenetic significance. J. Insect. Physiol. 18: 31-42. [ Links ]
De Camino, R. & J. Irausqui. 1977. Proyecto para el establecimiento de una población de Pinus caribaea al sur del Edo. Monagas. Proyecto Chaguaramas, CONARE, Venezuela. [ Links ]
D’Ettorre, P. & J. Heinze. 2005. Individual Recognition in Ant Queens. Curr. Biol. 15: 2170-2174. [ Links ]
Do Nascimento, R., E. Morgan, J. Billen, E. Schoeters, T. Della Lucia & J. Bento. 1993. Variation with caste of the mandibular gland secretion in leaf-cutting ant Atta sexdens rubropilosa. J. Chem. Ecol. 19: 907-918. [ Links ]
Hefetz, A., C. Errard, A. Chambris & A. Le Negrate. 1996. Postpharyngeal gland secretion as modifier of aggressive behavior in the Myrmicinae ant Manica rubida. J. Insect. Behav. 9: 709-717. [ Links ]
Hernández, J.V., A. Cabrera & K. Jaffe. 1999. Mandibular gland secretion in different castes of the leaf-cutter ant Atta laevigata. J. Chem. Ecol. 25: 2433-2444. [ Links ]
Hernández, J.V., H. López & K. Jaffé. 2002. Nestmate recognition signal of the leaf-cutting ant Atta laevigata. J. Insect Physiol. 48: 287-295. [ Links ]
Hernández, J.V., W. Goitía, A. Osio, A. Cabrera, H. López, C. Sainz & K. Jaffe. 2006. Leaf-cutter ant species (Hymenoptera: Atta) differ in the types of cues used to differentiate between self and others. Anim. Behav. 71: 945-952. [ Links ]
Hernández, J.V., D. Rodríguez, Z. Tárano, F. Drijfhout & C. Sainz-Borgo. 2010. Are Postpharyngeal or Mandibular Gland Secretions the Source for Nestmate Recognition in Camponotus atriceps (Hymenoptera: Formicidae)? Sociobiology 56: 1-18. [ Links ]
Hölldobler, B. & E.O. Wilson. 1990. The Ants. Harvard University, Massachusetts, USA. [ Links ]
Howard, R.W. & G.J. Blomquist. 2005. Ecological, behavioral, and biochemical aspects of insect hydrocarbons. Annu. Rev. Entomol. 50: 371-393. [ Links ]
Hughes, W., P. Howse & D. Goulsom. 2001. Mandibular gland chemistry of grass cutting ants. Species, caste and colony variation. J. Chem. Ecol. 27: 109-124. [ Links ]
Jaffe, K. 1983. Chemical communication systems in the ant Atta cephalotes, p 165-180. In P. Jaisson (ed.). Social Insects in the tropics. Presses de L´Université Paris Nord, Paris, France. [ Links ]
Jaffe, K. & J. Navarro. 1985. Comunicación química en obreras de la hormiga cortadora de pasto Acromyrmex landolti Forel, 1884 (Hymenoptera, Formicidae). Rev. Bras. Entomol. 29: 350-361. [ Links ]
Jaffe, K. & M. Marquez. 1987. Chemical communication in the ponerinae ant Ectatomma ruidum. Insectes Soc. 34: 87-95. [ Links ]
Jaffe, K., M.I. Alvarez & O. Colmenares. 1990. On the communication systems of the Dolichoderinae ants Azteca foreli and Conomyrma sp. Bol. Entomol. Venez. N. S. 5: 165-182. [ Links ]
Jutsum, A.R., R.S. Saunders & J.M. Cherret. 1979. Intraespecific australian in the leaf cutting ant Acromyrmex octopinosus. Anim. Behav. 27: 839-844. [ Links ]
Kather, R & S.J. Martin. 2012. Cuticular hydrocarbon profiles as a taxonomic tool: advantages, limitations and technical aspects. Physiol. Entomol. 37: 25-32. [ Links ]
Lahav, S., V. Soroker, A. Hefetz & R.K. Vander Meer. 1999. Direct behavioral evidence for hydrocarbons as ant recognition discriminators. Naturwissenchaften 86: 246-249. [ Links ]
Lenoir, A., D. Cuisset & A. Hefetz. 2001. Effects of social isolation on hydrocarbon pattern and nestmate recognition in the ant Aphaenogaster senilis (Hymenoptera: Formicidae). Insectes Soc. 48: 101-109. [ Links ]
López, H. 2002. Estudio de los volátiles cefálicos para la hormiga A. cephalotes mediante dos métodos de extracción. Tesis de Bachillerato, Universidad Simón Bolívar, Caracas, Venezuela. [ Links ]
Martin, S., K. Trontti, S. Shemilt, F. Drijfhout, R. Butlin & D. Jackson. 2012. Weak patriline effects are present in the cuticular hydrocarbon profiles of isolated Formica exsecta ants but they disappear in the colony environment. Ecol. Evol. 2: 2333-2346. [ Links ]
Martin, S. & F. Drijfhout. 2009a. A review of ant cuticular hydrocarbons. J. Chem. Ecol. 35: 1151-1161. [ Links ]
Martin, S. & F. Drijfhout. 2009b. Nestmate and task cues are Influenced and Encoded Differently within Ant Cuticular Hydrocarbon Profiles. J. Chem. Ecol. 35: 368-374. [ Links ]
Morgan, E.D., M.R. Inwood & M.C. Cammaerts. 1978. The mandibular gland secretion of the ant Myrmica scabrinodis. Physiol. Entomol. 3: 107-114. [ Links ]
Morgan, E.D. 1990. Preparation of small-scale samples from insects for chromatography. Anal. Chim. Acta 236: 227-235. [ Links ]
Obin, M.S. 1986. Nestmate recognition cues in laboratory and field colonies of Solenopsis invicta Buren (Hymenoptera: Formicidae): effect of environmental and the role of cuticular hydrocarbons. J. Chem. Ecol. 12: 1965-1975. [ Links ]
Riley, R.G., R.M. Silverstein & J.C. Moser. 1974. Isolation, identification, synthesis and biological activity of volatile compounds from the heads of Atta ants. J. Insect Physiol. 20: 1629-1637. [ Links ]
Sainz-Borgo, C., A. Cabrera & J.V. Hernández. 2011. Nestmate recognition in the ant Odontomachus bauri (Hymenoptera: Formicidae). Sociobiology 58: 701-718. [ Links ]
Singer, T. 1998. Roles of Hydrocarbons in the Recognition Systems of Insects. Amer. Zool. 38: 394-405. [ Links ]
Schubert, C. & O. Huber. 1989. The Gran Sabana: Panorama of a region. Cuadernos Lagoven, Caracas, Venezuela. [ Links ]
Soroker, V., C. Vienne & A. Hefetz. 1995. Hydrocarbons austral within and between nestmates in Cataglyphis niger (Hymenoptera, Formicidae). J. Chem. Ecol. 21: 365-378. [ Links ]
Soroker, V., C. Vienne & A. Hefetz. 1994. The postpharyngeal gland as a “gestalt” organ for nestmate recognition in the ant Cataglyphis niger. Naturwissenschaften 81: 510-513. [ Links ]
Van Wilgenburg, E., R. Sulc, J.K. Shea & N.D. Tsutsui. 2010. Deciphering the Chemical Basis of Nestmate Recognition. J. Chem. Ecol. 36: 751-758. [ Links ]
Van Zweden, J.S., J.B. Brask, J.H. Christensen, J.J. Boomsma, T.A. Linksvayer & P.D’ettorre. 2010. Blending of heritable recognition cues among ant nestmates creates distinct colony gestalt odours but prevents within-colony nepotism. J. Evol. Biol. 23: 1498-1508. [ Links ]
Vander Meer, R.K., D. Saliwanchik & B. Lavine. 1989. Temporal changes in colony cuticular hydrocarbon patterns of Solenopsis invicta: Implications for nestmate recognition. J. Chem. Ecol. 15: 2115-2125. [ Links ]
Whitehouse, M. & K. Jaffé. 1995. Nestmate recognition in the leaf cutting ant Atta laevigata. Insectes Soc. 42: 157-166. [ Links ]
Attygalle, A.B. & E.D. Morgan. 1984. Chemicals from the glands of ants. Chem. Soc. Rev. 13: 245-278. [ Links ]
Bagnéres, A.G. & E.D. Morgan. 1991. The postpharyngeal glands and the cuticule of Formicidae contain the same characteristic hydrocarbons. Experientia 47: 106-111. [ Links ]
Billen, J. & E.D. Morgan. 1998. Pheromone communication in social insects-sources and secretions, p. 3-33. In R.K. Vander Meer, M.D. Breed, K.E. Espelie & M.L. Winston (eds.). Pheromone Communication in Social Insects: Ants, Wasps, Bees, and Termites. Westview, Boulder, Oxford, England. [ Links ]
Blomquist, G.J., D.R. Nelson & M. De Renobales. 1987. Chemistry, biochemistry, and physiology of insect cuticular lipids. Arch. Insect Biochem. Physiol. 6: 227-265. [ Links ]
Blum, M.S., F. Padovani, H.R. Hermann & P.B. Kanowski. 1968. Chemical releasers of social behavior, XI: terpenes in the mandibular glands of Lasius umbratus. Ann. Entomol. Soc. Am. 61: 1354-1359. [ Links ]
Boulay, R. & A. Lenoir. 2001. Social isolation of mature workers affects nestmate recognition in the ant Camponotus fellah. Behav. Processes 55: 67-73. [ Links ]
Boulay, R., T. Katzav-Gozansky, A. Hefetz & A. Lenoir.2004. Odour convergence and tolerance between nestmate through trophallaxis and grooming in the ant Camponotus fellah (Dalla Torre). Insectes Soc. 51: 55-61. [ Links ]
Carlin, N.F. & B. Hölldobler. 1986. The kin recognition system of carpenter ants (Camponotus spp.). I Hierarchical cues in small colonies. Behav. Ecol. Sociobiol. 19: 123-134. [ Links ]
Carlin, N.F. & B. Hölldobler. 1987. The kin recognition systems of Carpenter ants (Camponotus spp). II. Larger colonies. Behav. Ecol. Sociobiol. 20: 209-217. [ Links ]
Crewe, R.M. & M.S. Blum. 1972. Alarm pheromones of the Attini: their phylogenetic significance. J. Insect. Physiol. 18: 31-42. [ Links ]
De Camino, R. & J. Irausqui. 1977. Proyecto para el establecimiento de una población de Pinus caribaea al sur del Edo. Monagas. Proyecto Chaguaramas, CONARE, Venezuela. [ Links ]
D’Ettorre, P. & J. Heinze. 2005. Individual Recognition in Ant Queens. Curr. Biol. 15: 2170-2174. [ Links ]
Do Nascimento, R., E. Morgan, J. Billen, E. Schoeters, T. Della Lucia & J. Bento. 1993. Variation with caste of the mandibular gland secretion in leaf-cutting ant Atta sexdens rubropilosa. J. Chem. Ecol. 19: 907-918. [ Links ]
Hefetz, A., C. Errard, A. Chambris & A. Le Negrate. 1996. Postpharyngeal gland secretion as modifier of aggressive behavior in the Myrmicinae ant Manica rubida. J. Insect. Behav. 9: 709-717. [ Links ]
Hernández, J.V., A. Cabrera & K. Jaffe. 1999. Mandibular gland secretion in different castes of the leaf-cutter ant Atta laevigata. J. Chem. Ecol. 25: 2433-2444. [ Links ]
Hernández, J.V., H. López & K. Jaffé. 2002. Nestmate recognition signal of the leaf-cutting ant Atta laevigata. J. Insect Physiol. 48: 287-295. [ Links ]
Hernández, J.V., W. Goitía, A. Osio, A. Cabrera, H. López, C. Sainz & K. Jaffe. 2006. Leaf-cutter ant species (Hymenoptera: Atta) differ in the types of cues used to differentiate between self and others. Anim. Behav. 71: 945-952. [ Links ]
Hernández, J.V., D. Rodríguez, Z. Tárano, F. Drijfhout & C. Sainz-Borgo. 2010. Are Postpharyngeal or Mandibular Gland Secretions the Source for Nestmate Recognition in Camponotus atriceps (Hymenoptera: Formicidae)? Sociobiology 56: 1-18. [ Links ]
Hölldobler, B. & E.O. Wilson. 1990. The Ants. Harvard University, Massachusetts, USA. [ Links ]
Howard, R.W. & G.J. Blomquist. 2005. Ecological, behavioral, and biochemical aspects of insect hydrocarbons. Annu. Rev. Entomol. 50: 371-393. [ Links ]
Hughes, W., P. Howse & D. Goulsom. 2001. Mandibular gland chemistry of grass cutting ants. Species, caste and colony variation. J. Chem. Ecol. 27: 109-124. [ Links ]
Jaffe, K. 1983. Chemical communication systems in the ant Atta cephalotes, p 165-180. In P. Jaisson (ed.). Social Insects in the tropics. Presses de L´Université Paris Nord, Paris, France. [ Links ]
Jaffe, K. & J. Navarro. 1985. Comunicación química en obreras de la hormiga cortadora de pasto Acromyrmex landolti Forel, 1884 (Hymenoptera, Formicidae). Rev. Bras. Entomol. 29: 350-361. [ Links ]
Jaffe, K. & M. Marquez. 1987. Chemical communication in the ponerinae ant Ectatomma ruidum. Insectes Soc. 34: 87-95. [ Links ]
Jaffe, K., M.I. Alvarez & O. Colmenares. 1990. On the communication systems of the Dolichoderinae ants Azteca foreli and Conomyrma sp. Bol. Entomol. Venez. N. S. 5: 165-182. [ Links ]
Jutsum, A.R., R.S. Saunders & J.M. Cherret. 1979. Intraespecific australian in the leaf cutting ant Acromyrmex octopinosus. Anim. Behav. 27: 839-844. [ Links ]
Kather, R & S.J. Martin. 2012. Cuticular hydrocarbon profiles as a taxonomic tool: advantages, limitations and technical aspects. Physiol. Entomol. 37: 25-32. [ Links ]
Lahav, S., V. Soroker, A. Hefetz & R.K. Vander Meer. 1999. Direct behavioral evidence for hydrocarbons as ant recognition discriminators. Naturwissenchaften 86: 246-249. [ Links ]
Lenoir, A., D. Cuisset & A. Hefetz. 2001. Effects of social isolation on hydrocarbon pattern and nestmate recognition in the ant Aphaenogaster senilis (Hymenoptera: Formicidae). Insectes Soc. 48: 101-109. [ Links ]
López, H. 2002. Estudio de los volátiles cefálicos para la hormiga A. cephalotes mediante dos métodos de extracción. Tesis de Bachillerato, Universidad Simón Bolívar, Caracas, Venezuela. [ Links ]
Martin, S., K. Trontti, S. Shemilt, F. Drijfhout, R. Butlin & D. Jackson. 2012. Weak patriline effects are present in the cuticular hydrocarbon profiles of isolated Formica exsecta ants but they disappear in the colony environment. Ecol. Evol. 2: 2333-2346. [ Links ]
Martin, S. & F. Drijfhout. 2009a. A review of ant cuticular hydrocarbons. J. Chem. Ecol. 35: 1151-1161. [ Links ]
Martin, S. & F. Drijfhout. 2009b. Nestmate and task cues are Influenced and Encoded Differently within Ant Cuticular Hydrocarbon Profiles. J. Chem. Ecol. 35: 368-374. [ Links ]
Morgan, E.D., M.R. Inwood & M.C. Cammaerts. 1978. The mandibular gland secretion of the ant Myrmica scabrinodis. Physiol. Entomol. 3: 107-114. [ Links ]
Morgan, E.D. 1990. Preparation of small-scale samples from insects for chromatography. Anal. Chim. Acta 236: 227-235. [ Links ]
Obin, M.S. 1986. Nestmate recognition cues in laboratory and field colonies of Solenopsis invicta Buren (Hymenoptera: Formicidae): effect of environmental and the role of cuticular hydrocarbons. J. Chem. Ecol. 12: 1965-1975. [ Links ]
Riley, R.G., R.M. Silverstein & J.C. Moser. 1974. Isolation, identification, synthesis and biological activity of volatile compounds from the heads of Atta ants. J. Insect Physiol. 20: 1629-1637. [ Links ]
Sainz-Borgo, C., A. Cabrera & J.V. Hernández. 2011. Nestmate recognition in the ant Odontomachus bauri (Hymenoptera: Formicidae). Sociobiology 58: 701-718. [ Links ]
Singer, T. 1998. Roles of Hydrocarbons in the Recognition Systems of Insects. Amer. Zool. 38: 394-405. [ Links ]
Schubert, C. & O. Huber. 1989. The Gran Sabana: Panorama of a region. Cuadernos Lagoven, Caracas, Venezuela. [ Links ]
Soroker, V., C. Vienne & A. Hefetz. 1995. Hydrocarbons austral within and between nestmates in Cataglyphis niger (Hymenoptera, Formicidae). J. Chem. Ecol. 21: 365-378. [ Links ]
Soroker, V., C. Vienne & A. Hefetz. 1994. The postpharyngeal gland as a “gestalt” organ for nestmate recognition in the ant Cataglyphis niger. Naturwissenschaften 81: 510-513. [ Links ]
Van Wilgenburg, E., R. Sulc, J.K. Shea & N.D. Tsutsui. 2010. Deciphering the Chemical Basis of Nestmate Recognition. J. Chem. Ecol. 36: 751-758. [ Links ]
Van Zweden, J.S., J.B. Brask, J.H. Christensen, J.J. Boomsma, T.A. Linksvayer & P.D’ettorre. 2010. Blending of heritable recognition cues among ant nestmates creates distinct colony gestalt odours but prevents within-colony nepotism. J. Evol. Biol. 23: 1498-1508. [ Links ]
Vander Meer, R.K., D. Saliwanchik & B. Lavine. 1989. Temporal changes in colony cuticular hydrocarbon patterns of Solenopsis invicta: Implications for nestmate recognition. J. Chem. Ecol. 15: 2115-2125. [ Links ]
Whitehouse, M. & K. Jaffé. 1995. Nestmate recognition in the leaf cutting ant Atta laevigata. Insectes Soc. 42: 157-166. [ Links ]
*Correspondencia: Cristina Sainz-Borgo: Laboratorio de Ecología Química del Comportamiento, Departamento de Biología de Organismos, Universidad Simón Bolívar, Apartado 89000, Caracas 1080A, Venezuela; cristinasainzb@gmail.com
Bernardo Leal: Departamento de Química; Universidad Simón Bolívar, Apartado 89000, Caracas 1080A, Venezuela.
Aivlé Cabrera: Departamento de Química; Universidad Simón Bolívar, Apartado 89000, Caracas 1080A, Venezuela; acabrera@usb.ve
José V. Hernández: Laboratorio de Ecología Química del Comportamiento, Departamento de Biología de Organismos, Universidad Simón Bolívar, Apartado 89000, Caracas 1080A, Venezuela; jnandez@usb.ve
1. Laboratorio de Ecología Química del Comportamiento, Departamento de Biología de Organismos, Universidad Simón Bolívar, Apartado 89000, Caracas 1080A, Venezuela; cristinasainzb@gmail.com, jnandez@usb.ve
2. Departamento de Química; Universidad Simón Bolívar, Apartado 89000, Caracas 1080A, Venezuela; acabrera@usb.ve
Recibido 06-VIII-2012. Corregido 14-I-2013. Aceptado 08-II-2013.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}