










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.61 n.3 San José Sep. 2013
A ten-month diseases survey on wild Litopenaeus setiferus (Decapoda: Penaeidae) from Southern Gulf of Mexico
Diez meses de análisis de las enfermedades en especímenes silvestres de Litopenaeus setiferus en el sur del Golfo de México
Diez meses de análisis de las enfermedades en especímenes silvestres de Litopenaeus setiferus en el sur del Golfo de México
Rodolfo Enrique del Río-Rodríguez1*, Daniel Pech1,3*, Sonia Araceli Soto-Rodriguez2*, Monica Isela Gomez-Solano1 & Atahualpa Sosa-Lopez1
*Dirección para correspondencia:
Abstract
The development of shrimp aquaculture in Mexican coasts of the Gulf of Mexico began to be explored using the Pacific white shrimp Litopenaeus vannamei in the mid 90´s. Many concerns over the risk of disease transmission to the economically important native penaeids, have been the main deterrent for the aquaculture of L. vannamei in the region. Concurrently, more than 10 years of research experience on the aquaculture suitability of the native Litopenaeus setiferus from the Terminos Lagoon, in the Yucatán Peninsula, have been accumulated. The aim of this study was then to determine the seasonal variations of the naturally acquired diseases and the possible detection of exotic pathogens. For this, random subsamples (n~60) of juveniles L. setiferus were collected from monthly captures. In order to detect the widest range of pathogens, including infectious hypodermal and hematopoietic necrosis (IHHNv) and white spot syndrome (WSSv) viruses, both histopathological and molecular methods were employed. Monthly prevalence (%) was calculated for every finding. We were able to detect a total of 16 distinct histological anomalies, most of which the presump- tive aetiological agent was readily identified. PCR results for viruses were negative. For some pathogens and symbionts, the prevalence was significantly different between the adult and juvenile populations. Prevalence of diseases tended to be higher in juvenile shrimp than in adults. The results of this study indicated that L. setiferus carry a wide variety of pathogens and symbionts that seem to be endemic to penaeids of the Gulf of Mexico, and those juveniles were more conspicuous to acquire pathogens and symbionts than adults.
Key words: Litopenaeus setiferus, wild shrimp, diseases, Southern Mexico.
Resumen
Durante la década de los 90´s se introdujo el camarón blanco del Pacífico Litopenaeus vannamei a los Estados costeros mexicanos del Golfo de México con fines acuícolas, por lo que desde entonces existe preocupación por la posible introducción de enfermedades que puedan afectar a las poblaciones de camarones nativos. La investigación sobre la domesticación de especies nativas para una acuacultura sustentable se ha realizado por más de 10 años, sin embargo, aún existe escasa información sobre las enfermedades que se presentan de manera natural en estas poblaciones y posible trasfaunación. El presente estudio aborda el problema de las enfermedades encontradas en subpoblaciones de jóvenes y adultos de Litopenaeus setiferus del Área natural protegida Laguna de Términos, estado de Campeche, México. Técnicas de histología y biología molecular fueron utilizadas como herramientas de diagnóstico. Se encontró que L. setiferus es portador de patógenos y simbiontes endémicos del Golfo de México, y comparativamente, los jóvenes son más susceptibles en adquirir estas infecciones que los adultos, como probable respuesta al ambiente lacustre que ocupan. No se encontró evidencia de los virus IHHNv y WSSv, aunque en trabajos más recientes en algunos Estados del Norte ya se han detectado en poblaciones silvestres.
Palabras clave: Litopenaeus setiferus, peneidos silvestres, enfermedades, Sur del Golfo de México
The decline of shrimp fisheries in the Gulf of Mexico in the last 10 years, have prompted the state governments of the region to promote the aquaculture of penaeid shrimp at rural and commercial levels. During mid 90´s, the shrimp aquaculture potential of the Mexican coasts of the Gulf of Mexico began to be explored with the Pacific white shrimp Litopenaeus vannamei. However, due to outbreaks of Taura syndrome and White Spot viruses in shrimp farms of the Pacific coast (Jiménez et al. 1999, Lyle-Fritch et al. 2006), the Federal Government expressed concerns over the risk of disease transmission to economically important native penaeids of the Gulf of México, since shrimp larvae supply originated from the Pacific production. This concern has been one of the main deterrents for the L. vannamei culture expansion in Southeast Mexico.
Between 1997 and 2002, three independent studies were carried out to determine the frequency and prevalence of diseases and/or symbionts of wild and cultured penaeids of the Gulf of Mexico. Vidal-Martinez et al. (2002) informed the presence of one flagellate, five ciliates, one Microsporideans, one gregarine and six metazoan parasites on four native shrimp species along the coast of Yucatan state; the ectosymbionts shown the highest prevalence in all hosts. In May and September 1999, a team lead by Chavez-Sanchez et al. (2002) sampled wild and cultured penaeid species in 10 stations along three coastal states of the Gulf of Mexico; Tamaulipas (6), veracruz (1) and Campeche (3). They informed a wide range of diseases and infestations (from bacteria to metazoan parasites) occurring in wild native penaeids, but stressed that no serious disease (i. e. viral diseases) was detected. The higher prevalence and severity grades were observed for parasites (Cestoda) and for invasive and non-invasive protozoans. The sampling scheme was repeated in 2000, but included only five out of 10 of the former stations; they confirmed that parasites and symbionts are the most conspicuous biological associations of the wild penaeids from the Mexican coast of the Gulf. Furthermore, Lopez-Tellez et al. (2009) determined the seasonal variation of ectosymbiotic ciliates on farmed and wild shrimps during a 12-month period (December 2001-November 2002) from the Yucatan state coasts. They found that patterns of ciliate invasions differed between cultured (L. vannamei) and wild stocks (Farfantepenaeus duorarum and F. brasiliensis); heavier intensities of colonization by Zoothamniun sp. and Epistylis sp. (in order of importance) occurred in cultured shrimp during the rainy season, (September-November) while in wild shrimp, higher intensities of infection occurred later in the year (winter frontal storms, December to the following February). For the cultured L. vannamei, temperature, turbidity-as indirect measure of pond fertilization-and survival variations, were accounted as significant explanatory variables of ciliates seasonality patterns. Explanatory variables for the ciliate mean intensity of infestations of the wild counterparts were not informed.
Litopenaeus setiferus, the white shrimp of the Gulf of Mexico has one of its major breeding grounds within the influence area of the Terminos Lagoon. In the last 10 years there has been an important effort to evaluate the suitability of L. setiferus domestication for its use in aquaculture. This Atlantic species has been considered as alternative species for shrimp culture in the American Atlantic area, due to some promising aquaculture traits and that represents a low ecological or disease risk as a consequence of unintended release associated with floods or hurricanes (Arena et al. 2003). However a detailed knowledge on the natural disease fluctuation affecting this species is still needed. It is yet to be discovered if there are significant pathogens affecting these natural populations and if exotic viruses-already common in the Mexican Pacific-have managed to establish. Also, it is important to generate back- ground information for candidate aquaculture species as in this case and elucidate if pathogens and simbionts affect juvenile and adult populations in the same manner and occurrence during a continued and extended period in an area where the establishment of shrimp culture, depends on the domestication of native shrimp.
Since 2002 a collaborative series of inshore research experiments using wild stocks of L. setiferus has been carried out in order to establish the baseline knowledge of its aqua- culture traits and the diseases affecting these particular stocks, also testing the probable presence of exotic pathogens. Here by using histology as the main presumptive diagnostic technique, the microbial diseases and parasites affecting juveniles and adults of L. setiferus are presented. Molecular methods were also used to screen the possible presence of IHHNv and WSSv, currently the two most common virus agents in the Mexican Pacific aquaculture affecting L. vannamei.
Materials and Methods
Sample collection: Monthly captures of shrimp (L. setiferus) from July 2002 to June 2003 were carried out at two selected sites from the Terminos lagoon (Fig. 1), Southeast Gulf of Mexico for aquaculture experiments. The sample schedule was intended to cover a 12-month period, but samplings of January and February 2003 were not carried out due to an unusual persistence of stormy conditions that made navigation difficult. The cumulative number of juvenile and adult shrimp for the histopathological study was over the minimum necessary to comply with a 95% of confidence level, assuming a 10% diseases prevalence in a population conformed by more than 100 000 specimens (OIE 2009).
Adult (ADU) shrimp (avg wt 36.26g, SD±3.47; avg tot length 17.35g SD±1.08; 139 males and 163 females) were collected at “Punta de las Disciplinas” (site 1, 91°55´14´´ N - 18°43´34´´ W) sea bound North-West mouth of the lagoon. Juvenile (JUv) shrimp (avg wt 10.68g, SD±4.16; avg total length 11.58g SD±1.82) were captured using gill nets at “Desembocadura de Boca Chica” (site 2, 92°15´46´´ N - 18°43´21´´ W). Shrimp were fixed, processed for histology and stained with Mayer-Bennett’s Hematoxylin and Eosin-Phloxine (H&E) according to Bell & Lightner (1988) and Lightner (1996).
Molecular methods for viral detection: A small portion of muscle, hepatopancreas and pleopods from each shrimp were excised and fixed in ethanol 96° (prior to histological fixation). A subsample (n=90) was selected subsequently to histopathological observations that could indicate possible virus involvement (i. e. spheroids in lymphoid organ, apparent inclusion bodies, cuticular epithelium and antennal gland necrosis) and was analyzed for white spot syndrome virus (WSSv) and infectious hypodermal and hematopoietic necrosis virus (IHHNV) by PCR methods following the protocols of Centro de Investigación en Alimentación y Desarrollo (CIAD Mazatlan Unit). DNA extraction kit (IQ2000) was used for extraction of viral DNA according to the manufacturer’s instructions. Briefly, approximately 20mg of tissue was homogenized in 500mL lysis buffer, incubated for 10min at 95ºC followed by centrifugation for 10min at 1 200g and the supernatant was transferred to a new tube. DNA was precipitated by the addition of 400mL of 95% cold ethanol. Pel- leted DNA was washed with 95% ethanol by centrifugation and airdried. The dried DNA pellets were suspended in 100mL of double dis- tilled water (ddH2O) or Diethylpyrocarbonate treated Water (DEPC - H2O).
The DNA samples were submitted to one-step for IHHNv and nested PCR tests for WSSv using the IQ2000TM detection system. PCR reactions were carried out in a 10.0 and 17.0μL (WSSV) reaction mixture for the first and nested step, and 13μL for IHHNV. Amplification was performed in a programmable thermal cycler (Eppendorf Mastercycler). The amplified products were fragments of 550 and 296bp for WSSv and fragments of 438 or 644bp for IHHNv. Products were separated in 2.0% agarose gels, stained with ethidium bromide and visualized using a Gel Documenta- tion System (UvP BioImaging Systems).
Prevalence was taken in this study as the number of instances of disease expressed in percentage in a known population, at a desig- nated time, without distinction between old and new cases (Thrusfield 2007). The percentage of infected host (PIH) is defined as the number of hosts of one species infected with one or more infectious agent in a sample divided by the total number of hosts of that species within a discrete time (Pech et al. 2010). A Kruskal-Wallis non-parametric ANOVA was used to test differences in histological anomalies between juvenile and adult shrimps and among months. The significance of statistical analysis was established at α=0.05, unless otherwise stated.
Results
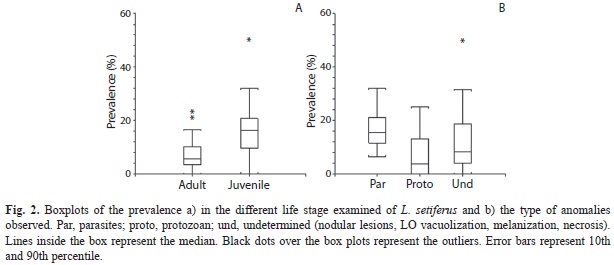
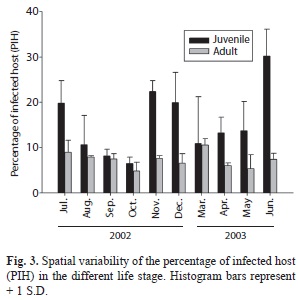
Prevalence of diseases between juvenile and adult shrimp: A total of 16 distinctly histological anomalies were detected, and in most of them the aetiological agent was generically identified. The anomalies found and their prevalence is depicted in Table 1. Significant differences in the prevalence as a function of the life stage of L. setiferus and the type of infection were observed. No effect of sampling month was observed (Table 2). Juvenile (JUV) organisms showed the highest prevalence (Fig. 2A) of metazoan parasites and undetermined anomalies seem to be the most conspicuous in L. setiferus (Fig. 2B). Using PIH as a variable summarizing all the anomalies detected in L. setiferus, significant differences were observed showing that JUv life stage showed the most propensity at acquiring infections (Table 2) independently of the sampling month (Fig. 3).
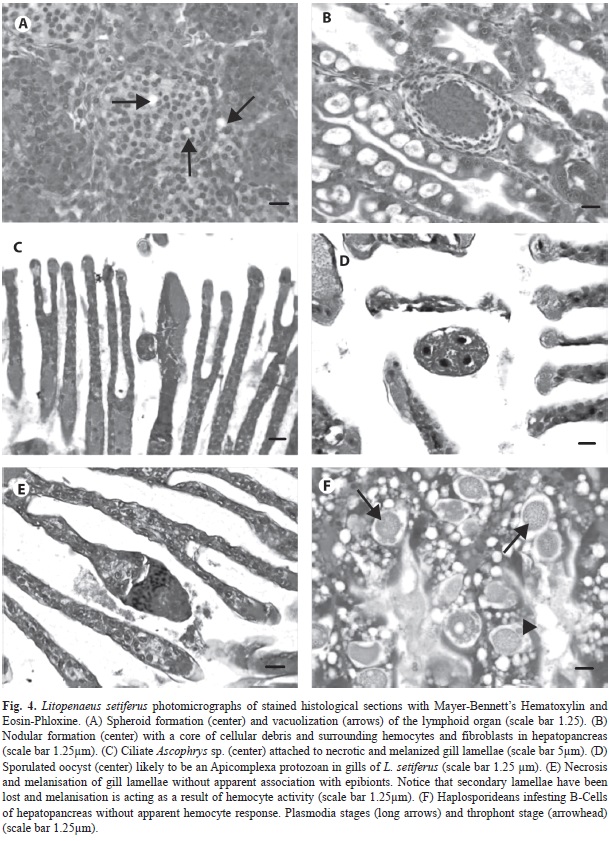
Microbial diseases and simbionts: In adult (ADU) and JUv specimens, a low prevalence of spheroid formations was detected in the lymphoid organ (LO) (Fig. 4A). During fresh examination the specimens with this pathology did not displayed any of the known gross signs of disease caused by major known viruses in the Americas (WSSV, TSV, IHHNV) and other tissues did not provide pathogno- monic characteristics of viral infection. PCR results were negative for IHHNV and WSSV viruses (Table 3).
Nodular lesions were observed in mid and distal hepatopancreas intratubular epithelium in both age groups (Fig. 4B). The presence of these nodules peaked in March and prevailed in JUv in April (Table 2), which suggest that this type of infection is more prevalent during the dry season. These lesions, similar to advanced stages of necrotizing hepatopancreatitis (NHP), involve hemocytic response, which develop into a core of melanized cells and debris.
Gill tissue contained the most diverse range of microorganisms from all the histopathological findings. Colonial peritrichous protozooans treated here as Epistylis-Zoothamnium complex were observed in JUV, with the highest prevalence in November (34%). It was not always possible to discern the presence of the myonem in Zoothamnium sp. from the histological sections, that would allow us differentiating between the common characteristics shared with Epistylis sp. (i. e. stalked, “U” shaped macronucleus). For the sake of argument, we treated this finding as a binomial-genus complex. The frequency of this epibiont complex was always higher in JUV than in ADU; they were not observed in either JUv or ADU shrimp during the dry months (March, April, June). In the present study, other ciliates were also observed generally in low prevalence, with the exception of Ascophrys spp. (Fig. 4C) that infected half of the JUv shrimp during June 2003 (Table 1). An unidentified protozoan was observed in the gill tissue; this protozoan shows a tetrad of cells seemingly enclosed in a kind of cyst (Fig. 4D) and we believe that represents an sporulated oocyst of the Apicomplexa.
Parasites: Intramuscular invasive protozoan Thelohania (Agmasoma) penai was observed infecting ADU all over the sampling period except in April, May and June, indicating that this infection prevails in this segment of the population during the main rainy period of the year. Infection prevalence, however, seems to remain low (Table 1). This microsporidean was observed in JUVonly in November, April and May, which suggests that pluvial runoff into the lagoon may play a role in its infection dynamics.
Intracellular haplosporideans were readily identified in tissue sections of HP (Fig. 4F). It was possible to observe several plasmodia stages occurring in one single organism, from uninucleated immature to ruptured mature- releasing trophonts. The presence of this parasite was almost confined to JUV specimens with a peak of infection in November (86%). In March, only one ADU specimen was positive, indicating that this parasite is well adapted to JUv shrimp as host, in which no apparent hemocytic response is elicited. Apparently, most “B” cells of the HP tubules appeared heavily infected.
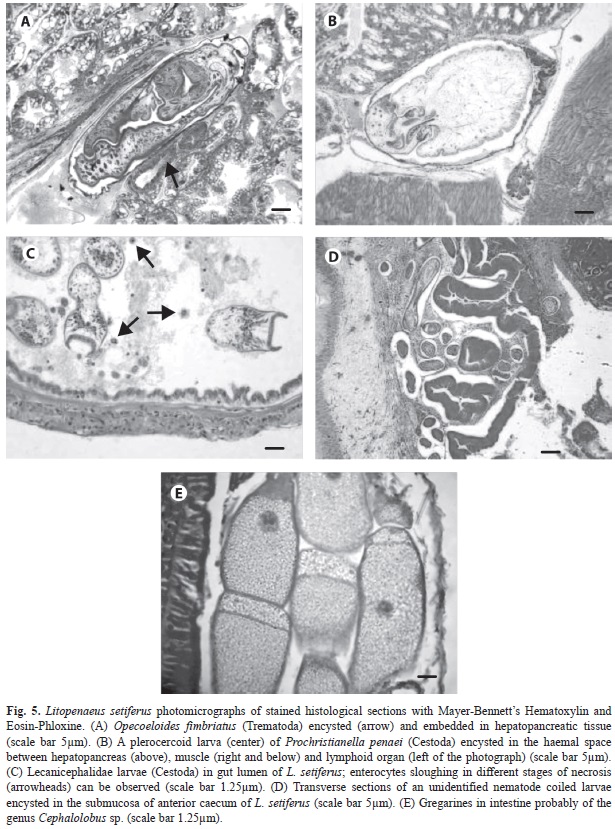
We found a trematode embedded in the hepatopancreatic intratubular lumina, forming a very thin capsule wall, that judging on the size of the acetabulum with respect to the oral sucker, it is likely to represent a larval stage of the digenean O. fimbriatus (Fig. 5A). The affected tissue showed a mild inflammatory response. Its prevalence suggests that this par- asite infests preferently JUV shrimp rather than ADU.
Prochristrianella penaei was invariably observed immersed in the hepatopancreas or related tissues (Fig. 5B). There was a peak in July (2002) and March (2003) in JUv and ADU, respectively. Also cestode larvae bearing a prominent apical sucker were observed in the intestine of L. setiferus, where apparently causes sloughing of epithelial cells of the mucosal layer (Fig. 5C). Enterocytes going through necrotic stages could be observed detached into the lumen (hypertrophy, karyor- rhexys, pyknosis).
An unidentified nematode was commonly found coiled and circumscribed to the sub-mucosa of the anterior ceca of L. setiferus where it prompts a relatively mild inflammaory response involving mainly fibroblasts and collagen fibers, rather than hemocytes (Fig. 5D). This response does not appear to have an effect on the parasite, which remains practi- cally intact in the host’s tissues, and it does not seem to burrow in the posterior ceca. This nematode was present in JUv all over the year except in June. Coincidentally, with respect to the cestode, the peak of JUV infection occurred in July, but for ADU appear to be bimodal with a peak in September and another in November (2002), that corresponds close to the end of the rainy season and the beginning of the North- wind season, respectively.
Gregarines were observed in only one JUV specimen during November (Fig. 5E). It is likely that the intensity of the infection by this parasite might have been underestimated since more than one stage can inhabit between the stomach and intestine; gut tract was not always possible to section all thorough for histology.
Histological anomalies of uncertain aetiology: A number of anomalies, for which it was not possible to associate with a causative agent, were observed in some tissues. Some of them might be related to microbial infections that could not be readily identified in tissues stained with H & E, or early immunological responses to subclinical infections or a challenging environment. Spermatophore melanization, a degenerative process was observed in adult males in low prevalence (Table 1). Melanosis and necrosis of gill tissues, in apparent absence of aetiological agents, were recurrent pathologies (Fig. 4E). These damages were observed in varying severity grades (data not shown) and occurred all over the sample period, with peaks of prevalence (JUV/ADUL) in December (23/40) and June (56/63). Necrosis without melanization was also a common observation.
Discussion
The etiology of spheroid formation in LO could not be determined in this study, since PCR results were negative. LO spheroids have been observed in positive IHHN and TSv in wild (Morales-Covarrubias & Chavez-Sanchez 1999) and experimentally induced infections in L. vannamei (Hasson et al. 1999); they have also been related to other systemic viral infections (Pantoja & Ligthner 2003). However, shrimp of this study may be developing in a challenging environment since LO spheroid formation is considered a major defense mechanism (Anggraeni & Owens 2000).
The nodular lesions observed in this study could be caused by bacteria from the natural environment, and may be associated to animals whose health was compromised and for which this environment is more challenging during the dry season. According to Gomez-Gil et al. (1998) bacteria can be isolated from several tissues including hepatopancreas of normal shrimp reared under experimental conditions; they isolated a wide range of species of the genus Vibrio spp. in those conditions. It has also been proven that wild and cultured penaeids harbor similar gut bacterial floral composition where members of the genus Vibrio were quantitatively dominant (Oxley et al. 2002). A high density of vibrios has been recently informed from the hepatopancreas of farmed L. vannamei jóvenes from Northwestern Mexico (Soto-Rodriguez et al. 2010). It is possible that a diverse range of vibrios are likely to be present in hepatopancreas since this organ runs through the digestive tract of shrimp and it is contaminated with bacterial flora. This differs from diseased animals where one or two species of bacteria predominate. High prevalence of Vibrio spp. (100%) has been informed for natural populations of L. vannamei broodstock from the Pacific coast of Mexico (Morales-Covarrubias et al. 1999); authors suggested that sampled population might have been severely stressed, although no high mortality of this species has been recorded or was observed in the area during that study.
Since early 70´s, microsporideans are known to infect L. setiferus in the coast of United States where their occurrence is common (Couch 1978). Furthermore, T. penaei was first informed in wild juveniles of the pink shrimp – Farfantepenaeus duoraum- and the red shrimp –F. brasiliensis- in three coastal locations of the State of Yucatan (vidal-Martinez et al. 2002), as well as in farmed L. vannamei in a nearby location; this may suggest a possible disease transference between wild and cultured shrimp. Microsporidians are common in wild shrimp inhabiting waters near to aquaculture devel- opments (Toubiana et al. 2004). It is likely that this parasite has a wider geographic and species range distribution in Mexican waters of the gulf.
Dyková et al. (1988) first described patho- genic haplosporideans infesting shrimp in 1998 from introduced L. vannamei into Cuba from Nicaragua, and recently, they seem to have reemerged –in the same host- in Belize (Nunan et al. 2007). To our knowledge, haplosporideans have been scarcely studied in wild penaeids. For the Mexican coasts, haplosporidiosis has been informed in the Pacific blue shrimp L. stylirostris (Lightner 1996) and from the Gulf of Mexico, only one specimen of F. duorarum from a locality known as Champoton, was informed infected with haplosporideans for the first time (Chavez-Sanchez et al. 2002); therefore, L. setiferus must also be registered as a new host. The prevalence of this parasite observed in the present study, suggest that this haplosporidean might be endemic to juvenile L. setiferus from the Terminos lagoon and since pathogenic, it may influence the recruiting rate of juveniles into the adult population during the rainy season.
With respect to the gregarines, two unspecified genera (Nematopsis spp. and Cephalolo- bus spp.) were informed by Chavez-Sanchez and co-workers (2002) with a prevalence that reached up to 53% in fresh samples of several species of native penaeids, distributed from Tamaulipas to Campeche, Mexico. A high prevalence of this parasite has been observed in the Bob shrimp Xiphophorus kroyeri, captured by our team in the same area (unpublished data). It is likely that the gregarines observed in L. setiferus belong to the genus Cephalolobus, as it shows a widening in the anterior portion of the trophozoite, characteristic of the genus according to Cuellar-Anjel (2008). Despite its relatively large geographic distribution, gregarines have not been informed in introduced Pacific white shrimp L. vannamei for aquaculture in the Gulf of Mexico so far.
During the rainy months, epibionts were a common finding in the study subject. Recently, Lopez-Tellez et al. (2009) informed that epibionts –especially Epistylis sp. and Zoothamnium sp.– are very common as mixed infections in wild shrimps from the coast of Yucatán during a year cycle, peaking during the Northern winter storm season. Interestingly, those researchers did also notice that the prevalence of Epistylis sp. was higher than Zoothamnium sp. in wild specimens, contrary to what they found for farmed L. vannamei in the study area. Zoothamnium sp. seems to have a higher infesting capacity in pond environments, a fact noticed by Overstreet in 1973. In this work, our results suggests that rainy conditions (from June to December) seem to favor the colonization of juveniles over adults, tending to drop below detection levels –established for this survey– during the dry season; this confirmed the former tendency. However, more dedicated research on Zoothamnium sp. and Epistylis sp. colonization on free living crustaceans, has revealed that their infestation prevalence, higher during the spring and summer months, is not significantly associated to environmental variables but to the host abundance (Pinto-Utz 2003), emphasizing the opportunistic nature of these obligate epibionts.
To the best of our knowledge, Gregarine oocysts have not been previously informed in penaeids gill tissue (free or embedded) but in oysters (Azevedo & Cachola 1992). The oocysts observed might be the free aquatic intermediate phase of Cephalolobus spp. or any other Apicomplexa that produces oocysts with at least four infective sporozoites. Nematopsis spp. can be discarded as they produce oocysts with a single sporozoite. According to Clopton (2002), no oocysts are known for Cephalolobus spp., although the family Cephalolobidae contains five species divided into two genera and all are intestinal parasites of crustacea. Oocysts are also shed along the faeces of infected vertebrate animals (Modry et al. 2001), and L. setiferus may be playing a role in keeping the infection in the lagoon system, as this protozoan was present during the rainy months (July, September and November) and mostly in juveniles.
Metazoan parasites, such as trematodes, cestodes and nematodes affecting the HP and the anterior ceca appear to have an important involvement in the health status of both age groups of L. setiferus. Metazoan parasites have been identified in fresh samples of penaeid shrimps from the Mexican part of the Gulf of Mexico as Helicometrina nimia, Opecoeloides fimbriatus (Trematoda), Prochristianella penaei (Cestoda) and Hysterothylacyum sp. (Nematoda) by Chavez-Sánchez et al. (2002) and Vidal-Martinez et al. (2002). Opecoeloides fimbriatus has been long informed in wild penaeid shrimp, being more common during the summer months; L. setiferus captured from Ossabaw Sound, Georgia (USA) harbors this parasite that matures in teleost fish mainly in the Sciaenidae family (Overstreet 1973). It has been also informed by vidal et al. (2002) in F. aztecus from one locality in the Yucatán Peninsula. The presence and pathology of P. penaei in penaeid shrimp have been studied at some extent in the Gulf of Mexico (Overstreet 1973, Couch 1978, Johnson 1989). It is widely distributed among several species of shrimps, and sometimes is capable of eliciting a hostdefense response of varying intensity, in which the thickness of the cellular cyst-made of migrating hemocytes fibroblast and collagen deposits-, gives an idea of the parasites length of residence in the host and/or host’s immunological vigor. These cestodes use L. setiferus as intermediate hosts and we found the infection to be persistent in all sampling months. Johnson (1989) suggested that the life cycle is probably completed in stingrays such as Dasyatis sabina and D. say. The former species is informed as a common inhabitant of the Terminos lagoon, where it penetrates to feed on juveniles or migrating sea-bound adult shrimps (Ramos-Miranda et al. 2005).
Hutton in 1959 (cited by Overstreet 1973), found Lecanicephalidae cestode larvae parasitizing wild shrimp in high numbers. Reports on these larvae in shrimp are scarce. More recently high prevalence of Lecanicephalidae larvae were described from wild L. schmitti from Maracaibo Lake in venezuela (Boada et al. 1999), and is probably the same unidentified larvae found in low prevalence in L. setiferus (3%) and F. aztecus (17%) from two locations in the Gulf of Mexico by Chavez-Sánchez et al. (2002). We observed that the prevalence and frequency of these larvae was also low.
At least four genera of nematodes occur in shrimp species of the Gulf of Mexico (Hutton 1962, Overstreet 1973, Feigenbaum 1975). It is not possible to suggest an identification of the nematode found here from histological sections. However, the genus Leptolaimus has been informed to inhabit the anterior caecum of F. aztecus and L. setiferus as a comensal (Hutton 1962, Overstreet 1973), and Thynnascaris sp. as larvae in F. brasiliensis (Feigenbaum 1975). Hysterothylacium sp. was informed in juveniles of F. brasiliensis captured in a coastal environment from Yucatán by Vidal-Martínez et al. (2002), although the organ in which this nematode was collected from was not specified. The study by Chavez-Sánchez et al. (2002), did not report nematodes in juvenile shrimps from the Gulf of Mexico, therefore examination of fresh material would be necessary to confirm the identification of our finding.
Gills are exposed organs susceptible to many aquatic diseases; therefore they are considered aquatic environmental indicators. Severe black-pigmented lesions have been informed to occur in gills of shrimp after 15 days exposure to cadmium and nickel under experimental conditions (Denton & Camp- bell 1990). Couch (1977) described extensive lesions in gills after exposure to certain concentrations of cadmium chloride and suggested that contamination can act as a cofactor in gill respiratory dysfunction in subjects submitted to environmental stressors, such as salinity fluctuation. The two stations, from which the shrimp of the present study were obtained, are close to the largest receiving areas of oil extraction platforms in the country, which is also located in the influence area of the Terminos lagoon, considered the largest coastal fluvial wetland system in the Gulf of Mexico. Besides heavy metals, at least other 30 pollutants from natural spillovers and continental runoffs have been indentified in shrimp grounds in the Campeche sound (Vidal-Martinez et al. 2006); pollution, in combination with the salinity fluctuation, may be contributing to the aetiology and prevalence of gill necrosis found here, although the contributing factors or their synergism as the origin of this specific pathology, might be worthy of further research.
The role of disease in species with aquaculture potential: Litopenaeus setiferus specimens have been collected for aquaculture research purposes from the Terminos lagoon for a number of years. Terminos is the largest coastal lagoon ecosystem of México and its complex environmental evolution has been continuously studied. Similarly to the results of other surveys in open environments, L. setiferus carry a wide variety of pathogens and opportunists, which mostly seem to be endemic to penaeids of the Gulf of Mexico. Their varying prevalence may be the consequence of the hydrological regime (Yañez-Arancibia & Day 1988, Cu-Escamilla 2003) that provides the micro-environments in which these pathogens and its host interact. A fact noticed by Fein- genbaun since 1975, is that shrimp species of the Gulf of Mexico harbor a heavier load of parasites when compared to species of the Mexican Pacific. Results of a pathological survey by Morales-Covarrubias & Chavez- Sanchez (1999) and our results, confirm the former statement.
At the end of the 90´s, some reports on viruses in wild penaeid populations from the Mexican Pacific coast were issued (Morales- Covarrubias et al. 1999, Pantoja et al. 1999); no definitive evidence indicating the presence of major viral pathogens was found in shrimp from this survey sites. Unfortunately, occurrence of IHHNv and TSV has been informed for wild L. setiferus and F. aztecus from Laguna Madre in Tamaulipas, the bordering state of the Gulf of México. Guzmán-Sáenz et al. (2009) found low prevalence of infected individuals of both species, suggesting that the origin of these infections in 2005 are due to local shrimp farming with L. vannamei or its closeness to Texas where TSV impacted shrimp aquaculture in 2004. Moreover, between 2008 and 2010, farmed L. vannamei infected with IHHNV was informed for the first time in Tamaulipas and Tabasco (Lopez-Tellez et al.2010), the latter a neighboring state close to the
Terminos Lagoon.
The Terminos lagoon is part of a natural protected area with human settlements dedicated to diverse production activities. Attempts were made in the past to introduce the Pacific white shrimp L. vannamei for aquaculture purposes that did not progress, and federal law currently prohibits it. Native shrimp species are still under study aiming to rural aquaculture development. The establishment of disease status of a native species previous to being developed in enclosed environments may help to solve future controversies with respect to the assessment of disease outbreaks sources.
Acknowledgments
This study was partially financed by the National Council for Science and Technology (CONACYT Mexico, Project – 36780B): “Risk factors and prevalence associated with the presence of IHHNv, TSV and BP in wild and cultured shrimp (Farfantepenaeus duorarum, Litopenaeus setiferus and L. vannamei) from the coast of Campeche and Yucatán” (in Spanish). The authors are indebted to the researchers and personnel of the Unidad Multidisciplinaria de Investigación UNAM currently located at Sisal Yucatán, formerly known as Laboratorio de Ecofisiología, Facultad de Ciencias UNAM, and also to Edgar Mendoza- Franco for critically reviewing this manuscript. Daniel Pech was funded by FOMIX Yucatán award “Sensibilidad y vulnerabilidad de los ecosistemas costeros del sureste de México ante el Cambio Climático Global”.
References
Anggraeni, M.S. & L. Owens. 2000. The haemocytic origin of lymphoid organ spheroid cells in the penaeid prawn Penaeus monodon. Dis. Aquat. Organ. 40: 85-92. [ Links ]
Arena, L., M. Montalvan, G. Espinoza, G. Gaxiola, A. Sánchez, A. van Wormhoudt, D. Hernández, R. Díaz & C. Rosas. 2003. Genetic relationship between Litopenaeus setiferus (L.) and L. schmitti (Burkenroad) determined by using 16S mitochondrial sequences and enzymatic analysis. Aquac. Res. 34: 981-990. [ Links ]
Azevedo, C. & R. Cachola. 1992. Fine structure of the apicomplexa of Nematopsis sp. of two marine bivalve mollusks. Dis. Aquat. Organ. 14: 69-73. [ Links ]
Bell, T.A. & D.v. Lightner. 1988. A handbook of nor- mal shrimp histology. Special publication No. 4. World Aquaculture Society, Baton Rouge, Louisiana, EE.UU. [ Links ]
Boada, M., D. Rueda & R. Parra. 1999. Parásitos intestinales de juveniles de Litopenaeus schmitti (Burkenroad, 1936) de lago de Maracaibo, Estado Zulia, venezue- la. Memorias de la Fundación La Salle de Ciencias Naturales 59: 3-15. [ Links ]
Chavez-Sanchez, M.C., M. Hernandez-Martinez, S. Abad- Rosales, E. Fajer-Avila, L. Montoya-Rodriguez & P. Alvarez-Torres. 2002. A survey of infectious diseases and parasites of penaeid shrimp from the Gulf of Mexico. J. World. Aquac. Soc. 33: 316-329. [ Links ]
Clopton, R.E. 2002. Gregarines: A generic level review. Phylum Apicomplexa Levine, 1970: Order Eugrega- rinorida Léger, 1900, p. 205-288. In J.J. Lee, G. Lee- dale, D. Patterson & P.C. Bradbury (eds.). Illustrated Guide to the Protozoa, Society of Protozoologists, Lawrence, Kansas, EE.UU. [ Links ]
Couch, J.A. 1977. Ultrastructural study of lesions in gills of a marine shrimp exposed to cadmiun11. J. Invertebr. Pathol. 29: 267-288. [ Links ]
Couch, J.A. 1978. Diseases, parasites, and toxic responses of commercial penaeid shrimps of the Gulf of Mexico and South Atlantic coast of North America. Fish. Bull. 76: 1-44. [ Links ]
Cuellar-Anjel, J. 2008. Enfermedades por parásitos, p. 137-157. In v. Morales & J. Cuellar-Angel (eds.). Guía Técnica-Patología e Inmunología de camarones peneidos. Programa CYTED Red II-D vannamei, Panamá, República de Panamá [ Links ].
Cu-Escamilla, A. D. 2003. Análisis de la calidad del agua de la Laguna de Términos, ciclo 2001.2002. Tesis de Licenciatura, Universidad Autónoma de Campeche, México. [ Links ]
Denton, G.R.W. & R.S.F. Campbell. 1990. The pathology of Cadmium and Nickel toxicity in the banana shrimp (Penaeus merguiensis de Man). Asian Fish. Sci. 3: 287-297. [ Links ]
Dyková, I., J. Lom & E. Fajer. 1988. A new haplosporean infecting the hepatopancreas in the penaeid shrimp, Penaeus vannamei. J. Fish Dis. 11: 15-22. [ Links ]
Feigenbaum, D. 1975. Parasites of the commercial shrimp Penaeus vannamei Boone and Penaeus brasiliensis Latreille. Bull. Mar. Sci. 25: 491-514. [ Links ]
Gomez-Gil, B., L. Tron-Mayen, A. Roque, J.F. Turnbull, v. Inglis & A.L. Guerra-Flores. 1998. Species of Vibrio isolates from hepatopancreas haemolymph and digestive tract population for healthy juvenile Penaeus vannamei. Aquaculture 163: 1-9. [ Links ]
Guzman-Saenz, F.M., Z.J. Molina-Garza, R. Perez-Casta- ñeda, J.C. Ibarra-Gomez & L. Galaviz-Silva. 2009. Infectious Hypodermal and Hematopoietic Necrosis virus (IHHNv) and Taura Syndrome virus (TSv) in wild shrimp (Farfantepenaeus aztecus Ives, 1891 and Litopenaeus setiferus Linnaeus, 1767) of Laguna Madre, Gulf of Mexico. Rev. Biol. Mar. Oceanogr. 44: 663-672. [ Links ]
Hasson, K.W., D.v. Lightner, L.L. Mohney, R.M. Red- man & B.M. White. 1999. Role of lymphoid organ spheroids in chronic Taura syndrome virus (TSv) infections in Penaeus vannamei. Dis. Aquat. Organ. 38: 93-105. [ Links ]
Hutton, R.F. 1962. Studies on parasites of Penaeus duorarum, the pink shrimp: final report, Florida State Board of Conservation Marine Laboratory, Florida, EE.UU. [ Links ]
Jiménez, F., F. Galaviz, N. Segovia, M. Salinas, Z. Ramos & G. Molina. 1999. Presencia del virus de la Mancha Blanca (WSSv) en México. Boletín del Programa Nacional de Sanidad Acuícola y la Red de Diagnóstico 2: 1-3. [ Links ]
Johnson, S.K. 1989. Handbook of shrimp diseases. Department of Wildlife and Fisheries, Aquaculture Branch. Texas University, Texas, EE.UU. [ Links ]
Lightner, D.v. 1996. A handbook of shrimp pathology and diagnostic procedures for diseased cultured penaeid shrimp. World Aquaculture Society, Baton Rouge, Louisiana, EE.UU. [ Links ]
Lopez-Tellez, N.A., V.M. Vidal-Martínez & R.M. Overstreet. 2009. Seasonal variation of ectosymbiotic ciliates on farmed and wild shrimps from coastal Yucatan, Mexico. Aquaculture 287: 271-277. [ Links ]
López-Téllez, N.A., R. Rodríguez, J.V. Itzá & M.L. Unzueta. 2010. Condición sanitaria de las granjas de camarón blanco Litopenaeus Vannamei cultivado en el Golfo de México. I Reunión Nacional de Innovación Acuícola y Pesquera, Libro de Resúmenes, Campeche, México. [ Links ]
Lyle-Fritch, L.P., E. Romero-Beltran & F. Paez-Osuna. 2006. A survey on use of the chemical and biological products for shrimp farming in Sinaloa (NW Mexico). Aquac. Eng. 35: 135-146. [ Links ]
Modry, D., P. Daszak, J. Volf, M. vesely, S.J. Ball & B. Koudela. 2001. Five new species of coccidian (Apicomplexa: Eimeriidae) from Madagascan chameleons (Sauria: Chamaleonidae). Syst. Parasitol. 48: 117-123. [ Links ]
Morales-Covarrubias, M.S. & C. Chavez-Sanchez. 1999. Histopathological studies on wild broodstock of white shrimp Penaeus vannamei in the Platanitos area adjacent to San Blas, Nayarit, Mexico. J. World Aquac. Soc. 30: 192-200. [ Links ]
Morales-Covarrubias, M., L.M. Nunan, D.v. Lightner, J.C. Mota-Urbina, M.A. Aguirre-Garza & C. Chávez- Sánchez. 1999. Prevalence of infectious hypodermal and hematopoietic necrosis virus (IHHNv) in wild adult blue shrimp Penaeus stylirostris from the Northern Gulf of California, Mexico. J. Aquat. Anim. Health 11: 296-301. [ Links ]
Nunan, L.M., D.v. Lightner, C.R. Pantoja, N.A. Stokes & K.S. Reece. 2007. Characterization of a rediscovered haplosporidean parasite from cultured Penaeus vannamei. Dis. Aquat. Organ. 74: 67-75. [ Links ]
OIE. 2009. Manual of diagnostic test for aquatic animals, Office International des Epizooties. Paris, France. [ Links ]
Overstreet, R.M. 1973. Parasites of some penaeid shrimps with emphasis on reared hosts. Aquaculture 2: 105-140. [ Links ]
Oxley, A.P.A., W. Shipton, L. Owens & D. Mckay. 2002. Bacterial flora from the gut of the wild and cultured banana prawn, Penaeus merguiensis. J. Appl. Micro- biol. 93: 214-223. [ Links ]
Pantoja, C.R., D.V. Lightner & K.H. Holtschmit. 1999. Prevalence and Geographic distribution of Infectious Hypodermal and Hematopoietic Necrosis virus (IHHNV) in wild blue shrimp Penaeus stylirostris from the Gulf of California, Mexico. J. Aquat. Anim. Health. 11: 23-34. [ Links ]
Pantoja, C.R. & D.V. Lightner. 2003. Similarity between the histopathology of White Spot Syndrome virus and Yellow Head Syndrome virus and its relevance to diagnosis of YHV disease in the Americas. Aquaculture 218: 47-54. [ Links ]
Pech, D., M.L. Aguirre-Macedo, J.W. Lewis & V.M. Vidal-Martínez. 2010. Rainfall induces time-lagged changes in the proportion of the tropical aquatic hosts infected with metazoan parasites. Int. J. Parasitol. 40: 937-944. [ Links ]
Pinto-Utz, L.R. 2003. Identification, life history, and ecology of peritrich ciliates as epibionts on Calanoid copepods in the Chesapeake Bay, Ph. D. Thesis, Smithsonian Environmental Research Center, EE.UU. [ Links ]
Ramos-Miranda, J., L. Quiniou, D. Flores-Hernandez, T. Do-Chi, L. Ayala Perez & A. Sosa-Lopez. 2005. Spatial and temporal changes in the nekton of the Terminos lagoon, Campeche, Mexico. J. Fish Biol. 66: 513-530. [ Links ]
Soto-Rodriguez, S., B. Gomez-Gil, R. Lozano & A. Roque. 2010. Density of vibrios in hemolymph and hepatopancreas of diseased Pacific white shrimp Litopenaeus vannamei from northwestern Mexico. J. World Aquacult. Soc. 41: 76-83. [ Links ]
Thrusfield, M. 2007. Describing disease occurrence, p. 46-74. In: M. Thrusfield (ed). veterinary Epidemiology. Blackwell Publishing, Oxford, Inglaterra. [ Links ]
Toubiana, M., O. Guelorget, J.L. Bouchereau, H. Lucien-Brun & A. Marques. 2004. Microsporideans in penaeid shrimp along the west coast of Madagascar. Dis. Aquat. Organ. 58: 79-82. [ Links ]
Vidal-Martinez, V.M., A.M. Jimenez-Cueto & R. Sima-Alvarez. 2002. Parasites and simbionts of native and cultured penaeids from Yucatán, Mexico. J. Aquat. Anim. Health 14: 57-64. [ Links ]
Vidal-Martínez, V.M., M.L. Aguirre-Macedo, R.E. del Rio-Rodriguez, G. Gold-Bouchot, J. Rendon-von Osten & G.A. Miranda-Rosas. 2006. The pink shrimp Farfantepenaeus duorarum, its symbionts and helminthes as bioindicators of chemical pollution in Campeche Sound, Mexico. J. Helminthol. 80: 159-174. [ Links ]
Yañez-Arancibia, A. & J.W. Day. 1988. Ecological cha- racterization of Terminos Lagoon, a tropical lagoonestuarine system in the southern Gulf of Mexico, p. 1-26, In A. Yañez-Arancibia & J.W. Jr Day (eds.) Ecology of a coastal ecosystem in the southern part of the Gulf of Mexico: Terminos Lagoon region. Ins- tituto de Ciencias del Mar y Limnología, Universidad Autónoma de México, México. [ Links ]
Arena, L., M. Montalvan, G. Espinoza, G. Gaxiola, A. Sánchez, A. van Wormhoudt, D. Hernández, R. Díaz & C. Rosas. 2003. Genetic relationship between Litopenaeus setiferus (L.) and L. schmitti (Burkenroad) determined by using 16S mitochondrial sequences and enzymatic analysis. Aquac. Res. 34: 981-990. [ Links ]
Azevedo, C. & R. Cachola. 1992. Fine structure of the apicomplexa of Nematopsis sp. of two marine bivalve mollusks. Dis. Aquat. Organ. 14: 69-73. [ Links ]
Bell, T.A. & D.v. Lightner. 1988. A handbook of nor- mal shrimp histology. Special publication No. 4. World Aquaculture Society, Baton Rouge, Louisiana, EE.UU. [ Links ]
Boada, M., D. Rueda & R. Parra. 1999. Parásitos intestinales de juveniles de Litopenaeus schmitti (Burkenroad, 1936) de lago de Maracaibo, Estado Zulia, venezue- la. Memorias de la Fundación La Salle de Ciencias Naturales 59: 3-15. [ Links ]
Chavez-Sanchez, M.C., M. Hernandez-Martinez, S. Abad- Rosales, E. Fajer-Avila, L. Montoya-Rodriguez & P. Alvarez-Torres. 2002. A survey of infectious diseases and parasites of penaeid shrimp from the Gulf of Mexico. J. World. Aquac. Soc. 33: 316-329. [ Links ]
Clopton, R.E. 2002. Gregarines: A generic level review. Phylum Apicomplexa Levine, 1970: Order Eugrega- rinorida Léger, 1900, p. 205-288. In J.J. Lee, G. Lee- dale, D. Patterson & P.C. Bradbury (eds.). Illustrated Guide to the Protozoa, Society of Protozoologists, Lawrence, Kansas, EE.UU. [ Links ]
Couch, J.A. 1977. Ultrastructural study of lesions in gills of a marine shrimp exposed to cadmiun11. J. Invertebr. Pathol. 29: 267-288. [ Links ]
Couch, J.A. 1978. Diseases, parasites, and toxic responses of commercial penaeid shrimps of the Gulf of Mexico and South Atlantic coast of North America. Fish. Bull. 76: 1-44. [ Links ]
Cuellar-Anjel, J. 2008. Enfermedades por parásitos, p. 137-157. In v. Morales & J. Cuellar-Angel (eds.). Guía Técnica-Patología e Inmunología de camarones peneidos. Programa CYTED Red II-D vannamei, Panamá, República de Panamá [ Links ].
Cu-Escamilla, A. D. 2003. Análisis de la calidad del agua de la Laguna de Términos, ciclo 2001.2002. Tesis de Licenciatura, Universidad Autónoma de Campeche, México. [ Links ]
Denton, G.R.W. & R.S.F. Campbell. 1990. The pathology of Cadmium and Nickel toxicity in the banana shrimp (Penaeus merguiensis de Man). Asian Fish. Sci. 3: 287-297. [ Links ]
Dyková, I., J. Lom & E. Fajer. 1988. A new haplosporean infecting the hepatopancreas in the penaeid shrimp, Penaeus vannamei. J. Fish Dis. 11: 15-22. [ Links ]
Feigenbaum, D. 1975. Parasites of the commercial shrimp Penaeus vannamei Boone and Penaeus brasiliensis Latreille. Bull. Mar. Sci. 25: 491-514. [ Links ]
Gomez-Gil, B., L. Tron-Mayen, A. Roque, J.F. Turnbull, v. Inglis & A.L. Guerra-Flores. 1998. Species of Vibrio isolates from hepatopancreas haemolymph and digestive tract population for healthy juvenile Penaeus vannamei. Aquaculture 163: 1-9. [ Links ]
Guzman-Saenz, F.M., Z.J. Molina-Garza, R. Perez-Casta- ñeda, J.C. Ibarra-Gomez & L. Galaviz-Silva. 2009. Infectious Hypodermal and Hematopoietic Necrosis virus (IHHNv) and Taura Syndrome virus (TSv) in wild shrimp (Farfantepenaeus aztecus Ives, 1891 and Litopenaeus setiferus Linnaeus, 1767) of Laguna Madre, Gulf of Mexico. Rev. Biol. Mar. Oceanogr. 44: 663-672. [ Links ]
Hasson, K.W., D.v. Lightner, L.L. Mohney, R.M. Red- man & B.M. White. 1999. Role of lymphoid organ spheroids in chronic Taura syndrome virus (TSv) infections in Penaeus vannamei. Dis. Aquat. Organ. 38: 93-105. [ Links ]
Hutton, R.F. 1962. Studies on parasites of Penaeus duorarum, the pink shrimp: final report, Florida State Board of Conservation Marine Laboratory, Florida, EE.UU. [ Links ]
Jiménez, F., F. Galaviz, N. Segovia, M. Salinas, Z. Ramos & G. Molina. 1999. Presencia del virus de la Mancha Blanca (WSSv) en México. Boletín del Programa Nacional de Sanidad Acuícola y la Red de Diagnóstico 2: 1-3. [ Links ]
Johnson, S.K. 1989. Handbook of shrimp diseases. Department of Wildlife and Fisheries, Aquaculture Branch. Texas University, Texas, EE.UU. [ Links ]
Lightner, D.v. 1996. A handbook of shrimp pathology and diagnostic procedures for diseased cultured penaeid shrimp. World Aquaculture Society, Baton Rouge, Louisiana, EE.UU. [ Links ]
Lopez-Tellez, N.A., V.M. Vidal-Martínez & R.M. Overstreet. 2009. Seasonal variation of ectosymbiotic ciliates on farmed and wild shrimps from coastal Yucatan, Mexico. Aquaculture 287: 271-277. [ Links ]
López-Téllez, N.A., R. Rodríguez, J.V. Itzá & M.L. Unzueta. 2010. Condición sanitaria de las granjas de camarón blanco Litopenaeus Vannamei cultivado en el Golfo de México. I Reunión Nacional de Innovación Acuícola y Pesquera, Libro de Resúmenes, Campeche, México. [ Links ]
Lyle-Fritch, L.P., E. Romero-Beltran & F. Paez-Osuna. 2006. A survey on use of the chemical and biological products for shrimp farming in Sinaloa (NW Mexico). Aquac. Eng. 35: 135-146. [ Links ]
Modry, D., P. Daszak, J. Volf, M. vesely, S.J. Ball & B. Koudela. 2001. Five new species of coccidian (Apicomplexa: Eimeriidae) from Madagascan chameleons (Sauria: Chamaleonidae). Syst. Parasitol. 48: 117-123. [ Links ]
Morales-Covarrubias, M.S. & C. Chavez-Sanchez. 1999. Histopathological studies on wild broodstock of white shrimp Penaeus vannamei in the Platanitos area adjacent to San Blas, Nayarit, Mexico. J. World Aquac. Soc. 30: 192-200. [ Links ]
Morales-Covarrubias, M., L.M. Nunan, D.v. Lightner, J.C. Mota-Urbina, M.A. Aguirre-Garza & C. Chávez- Sánchez. 1999. Prevalence of infectious hypodermal and hematopoietic necrosis virus (IHHNv) in wild adult blue shrimp Penaeus stylirostris from the Northern Gulf of California, Mexico. J. Aquat. Anim. Health 11: 296-301. [ Links ]
Nunan, L.M., D.v. Lightner, C.R. Pantoja, N.A. Stokes & K.S. Reece. 2007. Characterization of a rediscovered haplosporidean parasite from cultured Penaeus vannamei. Dis. Aquat. Organ. 74: 67-75. [ Links ]
OIE. 2009. Manual of diagnostic test for aquatic animals, Office International des Epizooties. Paris, France. [ Links ]
Overstreet, R.M. 1973. Parasites of some penaeid shrimps with emphasis on reared hosts. Aquaculture 2: 105-140. [ Links ]
Oxley, A.P.A., W. Shipton, L. Owens & D. Mckay. 2002. Bacterial flora from the gut of the wild and cultured banana prawn, Penaeus merguiensis. J. Appl. Micro- biol. 93: 214-223. [ Links ]
Pantoja, C.R., D.V. Lightner & K.H. Holtschmit. 1999. Prevalence and Geographic distribution of Infectious Hypodermal and Hematopoietic Necrosis virus (IHHNV) in wild blue shrimp Penaeus stylirostris from the Gulf of California, Mexico. J. Aquat. Anim. Health. 11: 23-34. [ Links ]
Pantoja, C.R. & D.V. Lightner. 2003. Similarity between the histopathology of White Spot Syndrome virus and Yellow Head Syndrome virus and its relevance to diagnosis of YHV disease in the Americas. Aquaculture 218: 47-54. [ Links ]
Pech, D., M.L. Aguirre-Macedo, J.W. Lewis & V.M. Vidal-Martínez. 2010. Rainfall induces time-lagged changes in the proportion of the tropical aquatic hosts infected with metazoan parasites. Int. J. Parasitol. 40: 937-944. [ Links ]
Pinto-Utz, L.R. 2003. Identification, life history, and ecology of peritrich ciliates as epibionts on Calanoid copepods in the Chesapeake Bay, Ph. D. Thesis, Smithsonian Environmental Research Center, EE.UU. [ Links ]
Ramos-Miranda, J., L. Quiniou, D. Flores-Hernandez, T. Do-Chi, L. Ayala Perez & A. Sosa-Lopez. 2005. Spatial and temporal changes in the nekton of the Terminos lagoon, Campeche, Mexico. J. Fish Biol. 66: 513-530. [ Links ]
Soto-Rodriguez, S., B. Gomez-Gil, R. Lozano & A. Roque. 2010. Density of vibrios in hemolymph and hepatopancreas of diseased Pacific white shrimp Litopenaeus vannamei from northwestern Mexico. J. World Aquacult. Soc. 41: 76-83. [ Links ]
Thrusfield, M. 2007. Describing disease occurrence, p. 46-74. In: M. Thrusfield (ed). veterinary Epidemiology. Blackwell Publishing, Oxford, Inglaterra. [ Links ]
Toubiana, M., O. Guelorget, J.L. Bouchereau, H. Lucien-Brun & A. Marques. 2004. Microsporideans in penaeid shrimp along the west coast of Madagascar. Dis. Aquat. Organ. 58: 79-82. [ Links ]
Vidal-Martinez, V.M., A.M. Jimenez-Cueto & R. Sima-Alvarez. 2002. Parasites and simbionts of native and cultured penaeids from Yucatán, Mexico. J. Aquat. Anim. Health 14: 57-64. [ Links ]
Vidal-Martínez, V.M., M.L. Aguirre-Macedo, R.E. del Rio-Rodriguez, G. Gold-Bouchot, J. Rendon-von Osten & G.A. Miranda-Rosas. 2006. The pink shrimp Farfantepenaeus duorarum, its symbionts and helminthes as bioindicators of chemical pollution in Campeche Sound, Mexico. J. Helminthol. 80: 159-174. [ Links ]
Yañez-Arancibia, A. & J.W. Day. 1988. Ecological cha- racterization of Terminos Lagoon, a tropical lagoonestuarine system in the southern Gulf of Mexico, p. 1-26, In A. Yañez-Arancibia & J.W. Jr Day (eds.) Ecology of a coastal ecosystem in the southern part of the Gulf of Mexico: Terminos Lagoon region. Ins- tituto de Ciencias del Mar y Limnología, Universidad Autónoma de México, México. [ Links ]
*Correspondencia: Rodolfo Enrique del Río-Rodríguez: Instituto EPOMEX and Facultad de Ciencias Químico-Biológicas, Universidad Autónoma de Campeche, Av. Agustín Melgar s/n, entre Juan de la Barrera y calle 20, Colonia Buenavista, CP 24039, Campeche, Campeche México; redelrio@uacam.mx
Daniel Pech: Instituto EPOMEX and Facultad de Ciencias Químico-Biológicas, Universidad Autónoma de Campeche, Av. Agustín Melgar s/n, entre Juan de la Barrera y calle 20, Colonia Buenavista, CP 24039, Campeche, Campeche México. Departamento de Ciencias de la Sustentabilidad, ECOSUR, Unidad Campeche, Av. Rancho Polígono 2-A, Col. Ciudad Industrial, 24500 Lerma, Campeche, México; danielpech@uacam.mx
Sonia Araceli Soto-Rodriguez: Unidad Mazatlán en Acuicultura y Manejo Ambiental del CIAD, Av. Sabalo Cerritos s/n, Estero del Yugo, A. P. 711, CP 82010, Mazatlán Sinaloa, México; ssoto@ciad.mx
Monica Isela Gomez-Solano: Instituto EPOMEX and Facultad de Ciencias Químico-Biológicas, Universidad Autónoma de Campeche, Av. Agustín Melgar s/n, entre Juan de la Barrera y calle 20, Colonia Buenavista, CP 24039, Campeche, Campeche México; moigomez@uacam.mx
Atahualpa Sosa-Lopez: Instituto EPOMEX and Facultad de Ciencias Químico-Biológicas, Universidad Autónoma de Campeche, Av. Agustín Melgar s/n, entre Juan de la Barrera y calle 20, Colonia Buenavista, CP 24039, Campeche, Campeche México; moigomez@uacam.mx, atahsosa@uacam.mx
1. Instituto EPOMEX and Facultad de Ciencias Químico-Biológicas, Universidad Autónoma de Campeche, Av. Agustín Melgar s/n, entre Juan de la Barrera y calle 20, Colonia Buenavista, CP 24039, Campeche, Campeche México; redelrio@uacam.mx, danielpech@uacam.mx, moigomez@uacam.mx, atahsosa@uacam.mx
2. Unidad Mazatlán en Acuicultura y Manejo Ambiental del CIAD, Av. Sabalo Cerritos s/n, Estero del Yugo, A. P. 711, CP 82010, Mazatlán Sinaloa, México; ssoto@ciad.mx
3. Departamento de Ciencias de la Sustentabilidad, ECOSUR, Unidad Campeche, Av. Rancho Polígono 2-A, Col. Ciudad Industrial, 24500 Lerma, Campeche, México.
Received 20-vIII-2012. Corrected 10-XII-2012. Accepted 22-I-2013.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}