










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.61 n.3 San José Sep. 2013
Species composition and bathymetric distribution of gorgonians (Anthozoa: Octocorallia) on the Southern Mexican Pacific coast
Composición de especies y distribución batimétrica de gorgonios (Anthozoa: Octocorallia) en la costa del Pacífico mexicano
Composición de especies y distribución batimétrica de gorgonios (Anthozoa: Octocorallia) en la costa del Pacífico mexicano
*Dirección para correspondencia:
Abstract
Gorgonians are important components of coastal ecosystems, as they provide niches, natural compounds with medical applications and are used as bioindicators. Species composition and assemblage structure of gorgonians (Anthozoa: Octocorallia) were studied along a bathymetric profile in the Southern Mexican Pacific coast. Species composition was based on specimens collected within a depth range of 0-70m in 15 sites. The relative abundance of species was determined in six sites at four depths (5, 10, 20 and 25m) using three 10m2 transects at each depth level. Twenty-seven species of gorgonians belonging to six genera and three families were registered. The species composition varied with depth: 11 species were distributed between 0-25m depth, while 17 species were found between 40-70m depth interval. The shallow zone is characterized by a relatively large abundance of gorgonians, dominated by colonies of Leptogorgia cuspidata and L. ena. In contrast, the deepest zone was characterized by relatively low abundance of gorgonians, dominated by L. alba, the only species observed in both depth intervals. The similarity analysis showed differences in the composition and abundance of species by depth and site, suggesting that the main factor in determining the assemblage structure is depth. Results of this study suggest that the highest richness of gorgonian species in the study area may be located at depths of 40-70m, whereas the highest abundances are found between 5 and 10m depth. This study represents a contribution to the poorly known eastern Pacific gorgonian biota.
Key words: abundance, bathymetrical distribution, diversity, gorgonians assemblages, Mexican Pacific.
Resumen
La composición de especies y estructura de la comunidad de gorgonáceos se determinó a lo largo de un perfil batimétrico en la costa suroeste del Pacífico Mexicano. La composición de especies se realizó a través de una revisión y recolecta de especímenes entre 0 y 70m de profundidad en 15 sitios de muestreo. La caracterización de la estructura de la comunidad se realizó en seis sitios, en los cuales se colocaron tres transectos de 10m2 cada uno a 5, 10, 20 y 25m de profundidad. Se encontraron 27 especies de gorgonáceos pertenecientes a seis géneros y tres familias; aunque seis especies requieren ser confirmadas. La mayor riqueza de especies se registró entre 40 y 70m. La estructura de la comunidad entre 5 y 25m de profundidad mostró la existencia de dos zonas con la misma composición de especies pero con diferente abundancia relativa: una somera entre 5 y 10m de profundidad, caracterizada por una mayor abundancia de gorgonáceos y dominado por colonias de Leptogorgia cuspidata y Leptogorgia ena y una zona profunda entre 20 y 25m de profundidad, caracterizada por una baja abundancia de gorgonáceos y dominada por colonias de Leptogorgia alba.
Palabras clave: abundancia, distribución batimétrica, gorgonáceos, diversidad, Pacífico mexicano.
Gorgonians are characteristic and abundant components of coastal benthic communities of the eastern Pacific waters (Breedy & Guzmán 2002). Due to their arborescent form, gorgonians increase spatial and ecological heterogeneity, providing niches for many associated species (Lasker 1985, vreeland & Lasker 1989, Gerhardt 1990, Zea 1993). Some gorgonian species are significant sources of natural compounds used in biomedical research and pharmacology (McEnroe & Fenical 1978, Rodríguez & Ramírez 1994, Gutiérrez et al. 2004, 2005, 2006), and the presence and abundance of certain species has been shown to be beneficial to commercial fishing (Ruiz & Rada 2006). Gorgonians are also useful in environmental monitoring programs because various species are environmentally sensitive and can serve as bioindicators (Hernández-Muñoz et al. 2008).
Currently, efforts are underway to increase the body of knowledge of the richness of gorgonians in the Tropical eastern Pacific. Seven genera and four families have been reported in the shallow waters (Breedy & Guzmán 2003). Complete taxonomic reviews have been undertaken for the genera Pacifigorgia, Leptogorgia and Eugorgia (Breedy & Guzmán 2002, 2007, 2009), and new species are being described (Breedy et al. 2012). Nevertheless, our understanding of the species composition and distribution of gorgonian assemblages remains limited.
Knowledge of the species diversity and distribution patterns of gorgonians in the Tropical eastern Pacific is so far limited to surveys from Costa Rica (Breedy 2009) and Panamá (Guzmán et al. 2004, 2008). For the Mexican Pacific coast, which is approximately 8 000km long with diverse oceanographic settings, there is a single work by Reyes-Bonilla et al. (1997), reporting 10 gorgonian species at Cabo Pulmo.
The purpose of this study is to identify the gorgonian species that inhabit the bathymetric range of 0-70m and characterize the relative abundance of gorgonian species along a 5-25m bathymetric profile on the Southern Pacific coast of Mexico.
Materials and Methods
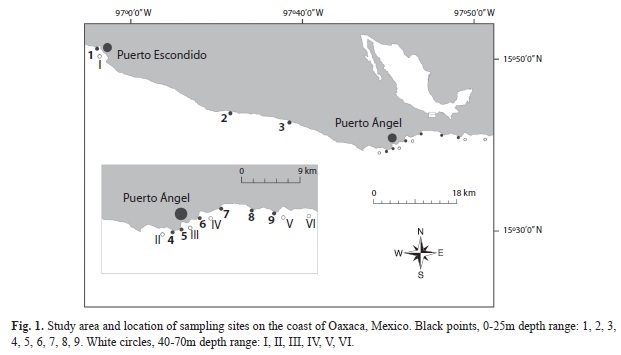
Study area: The study area is located in the Southern Mexican Pacific, along the coast of Oaxaca, between Puerto Escondido (15°52’10.48” N - 97°06’44.13” W) and Puerto Ángel (15°41’29.52” N - 96°14’13.38” W).
Fifteen sites were examined along 70km of coastline using SCUBA (Fig. 1). Bathymetric features, such as terraces and slope breaks, naturally divide the area into two types of sites: (1) adjacent to the coast, with a depth range from 0 to 25m, and (2) sites >100m from the coast, with a depth range from 40 to 70m. This division is likely related to the rate of sea level rise during the last 15 000 years and its effects on coastal erosion (Barrie & Conway 2002). All of the sites had rocky substratum, which is characteristic of the study area.
Taxonomy and species composition: To study the species composition, gorgonian colonies were identified in situ and collected for later validation. The census was conducted at 15 sites, nine sites were in the 0-25m depth range and six in the 40-70m depth range. Since the sampling effort may affect the final number of observed species, each site was visited the same number of times until no more species were recorded, resulting in a total of 10 visits per site.
The specimens were preserved by airdrying or fixed in 70% ethanol and identified through an analysis of the external morphology of the colony and the characteristics and composition of the sclerites following Bayer (1961) and Breedy & Guzmán (2002). A small fragment of each colony was placed in a hypochlorite sodium solution to separate the calcium carbonate sclerites from the organic material. Afterwards they were washed with water and examined under a light microscope (Olympus CX31). The specimens found were deposited in the Biological Collection of the Universidad del Mar, Oaxaca, Mexico, no. MHNUMAR-002B-01 - MHNUMAR-002B-27.
Gorgonian assemblages in relation to depth (5-25m) and sites: The species composition and relative abundance of gorgonians was determined for six of the nine sites in the 5-25m depth range (4, 5, 6, 7, 8 and 9). Stratified random sampling was conducted in each of the six study sites. Sampling was carried out along three 10m line transects at four depths (5, 10, 20 and 25m). These transects were marked with a 10m plastic tape to indicate ten consecutive 1m2 quadrats, which formed a total area of 10m2 for each transect. All gorgonians within each quadrat were identified and counted.
For the assemblage structure analysis, the original data were log(x+1) transformed to down-weight the contributions of dominant species relative to less common species (Clarke & Green 1988) and a Bray-Curtis dissimilarity matrix was constructed (Clarke et al. 2006). A two-way crossed similarity analysis (ANOSIM) with replicates was used to discover differences in the assemblage structures of gorgonians at different depths (5, 10, 20 and 25m) and among sites.
An analysis of similarity percentage (SIMPER) was used to identify the species responsible for major differences between depth groups. Analyses were performed with the program PRIMER 6 (Clarke 1993).
Results
Taxonomy and species composition: In total, 27 gorgonian species belonging to six genera and three families were found, although six species require confirmation (Table 1).
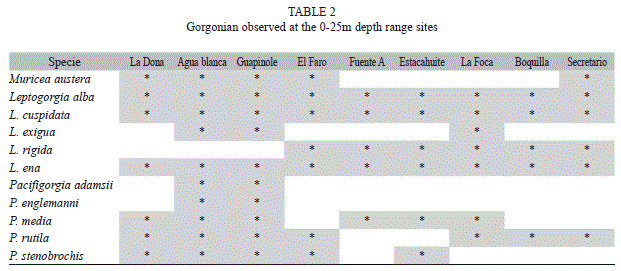
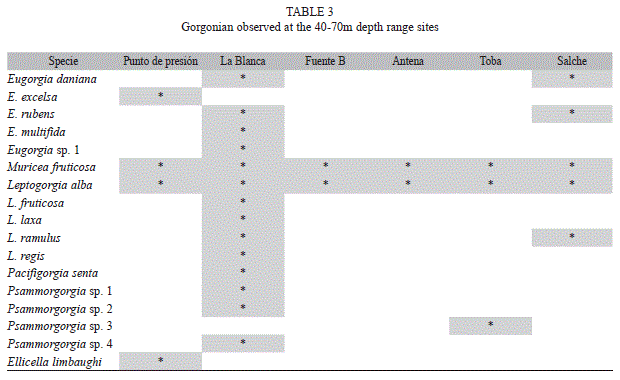
The composition of gorgonians varied with depth. Of the 27 species observed, 11 were found within the 0-25m depth interval (Table 2) and 17 were found in the 40-70m depth interval (Table 3). Leptogorgia alba was the only species observed in both depth intervals.
At the 0-25m depth, all species belonged to the genera Leptogorgia (n=5) and Pacifigorgia (n=5), with the exception of Muricea austera. Four species (L. alba, L. cuspidata, L. ena and L. rigida) were present in all sites at 0-25m. Pacifigorgia adamsii and P. englemanni were only found in Guapinole and Agua Blanca, the two sites with the greatest species richness (n=10) (Table 1).
The species in the 40-70m depth interval included seven genera. Leptogorgia had the highest number of species (n=6), whereas only one species was found for Pacifigorgia and Muricea. All species belonging to the genera Eugorgia (n=5) or Psammogorgia (n=4) were found in deeper waters. Only L. alba and M. fruticosa were present in all sites. Fifty percent of the species found in the 40-70m depth interval (n=9) were observed only in La Blanca, which was the site with the greatest species richness (n=15; Table 3). E. excelsa and Psammogorgia sp3 were each only found in one site (Punto de Presión and Toba, respectively; Table 3).
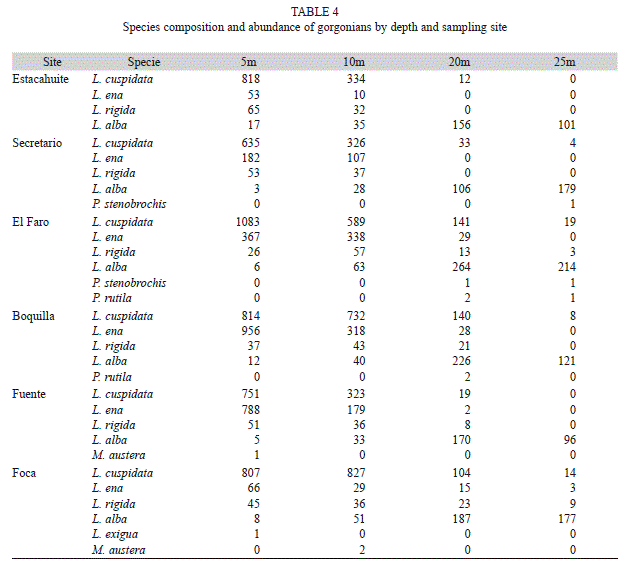
Assemblage structure in relation to depth (5-25m) and sites: Eight species were registered in the six shallow sites selected for this analysis. L. cuspidata, L. ena and L. alba were observed in all six sites, but only L. alba was found at all depths (Table 4).
The similarity analysis (ANOSIM) showed differences in the composition and abundance of species by depth (R global=0.68, p<0.1%) and site (R global=0.32, p<0.1%), suggesting that the main factor in determining the assemblage structure is depth.
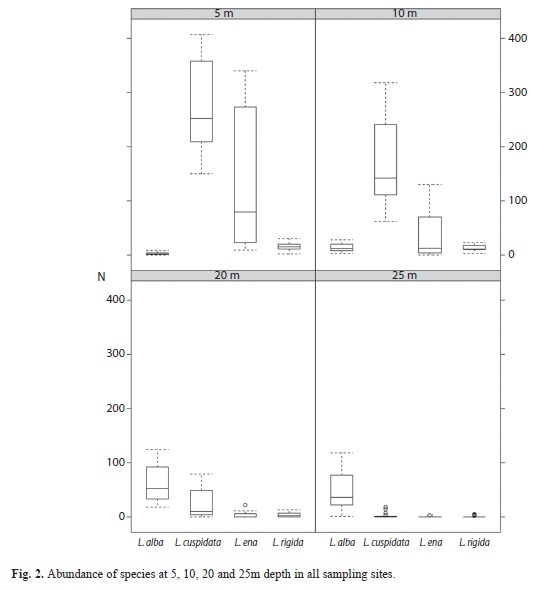
The ANOSIM pair wise comparisons with depth as a factor showed that the least similarity was found between the shallowest and the deepest groups (5 and 25m; R=0.97, p<0.1%). The most similarity was found between the closest depths, both at the deeper (20 and 25m; R=0.40, p<0.2%) and shallower depth ranges (5 and 10m; R=0.64 p<0.1%). These findings suggest the existence of two main zones with the same composition but different relative abundance of gorgonian species: a shallow zone at 5-10m depth, and a deep zone at 20-25m depth. The shallow zone is characterized by a relatively large abundance of gorgonians, dominated by colonies of L. cuspidata and L. ena. In contrast, the deepest zone was characterized by relatively low abundance of gorgonians, dominated by L. alba (Fig. 2).
The species that contributed the most to the dissimilarity (SIMPER analysis) between the two levels of shallow strata (5 and 10m) were L. cuspidata (49.0%), L. ena (43.3%), L. alba (4.5%) and L. rigida (3.4%); and the species that contributed the most to the dissimilarity between 20 and 25m assemblage were L. alba (60%), L. cuspidata (30.0%), L. rigida (5.4%) and L. ena (4.3%).
Discussion
Taxonomy and species composition: The gorgonian fauna along the study area is species-rich, with 27 species found compared to the 10 species recorded in the Gulf of California, Mexico (Reyes-Bonilla et al. 1997). However, further studies across the Mexican Pacific coast, including a wider bathymetric range, are required to achieve a complete inventory of species richness.
The gorgonian species richness in the present study area is similar to that of gorgonian communities in other areas of the Tropical eastern Pacific. Thirty species have been reported in Costa Rica (Breedy & Guzmán 2003, Breedy & Cortés 2008, Breedy 2009), 34 and 38 species respectively have been reported in the Gulfs of Chiriquí and Panamá (Las Perlas Archipelago) in Panamá (Guzmán et al. 2004, 2008) and 25 species have been reported in the Colombian Pacific (Prahl et al. 1987). Gorgonian species richness in the study area was higher than the one reported for some other regions of the Tropical eastern Pacific: 11 species in El Salvador, 7 in Nicaragua and 13 in Perú (Breedy & Guzmán 2003, 2007, 2009).
Gorgonian species reported in this study represent new records for the Pacific coast of México. Twenty five out of 27 species were documented in Oaxaca for the first time (the only exceptions are Eugorgia rubens and Eugorgia daniana). Leptogorgia ramulus, Leptogorgia regis and Ellisella limbaughi constitute new records for México. All other species reported here were previously documented as Mexican Pacific gorgonian fauna (Reyes-Bonilla et al. 1997, Breedy & Guzmán 2003, 2007, 2009).
Leptogorgia alba was the only species that was found at all sampling sites and depths. This species is distributed all along the Tropical eastern Pacific coast and reportedly occurs around oceanic islands (e.g. Isla del Coco, Costa Rica and the Galapagos Islands, Ecuador; Breedy et al. 2009). However, in this study we documented a more extensive bathymetric range distribution than previously found for this species (Breedy & Cortés 2008), reaching 70m, giving L. alba the widest depth range of any gorgonian in the Tropical eastern Pacific.
At the 40-70m depth interval, six genera were identified and 66% of the species were observed, suggesting that deeper areas harbor more gorgonians species than shallow ones. This finding confirms other reports of a higher richness and abundance of gorgonian species at depths greater than 40m (Sánchez 1999, Ruiz & Rada 2006, Matsumoto et al. 2007, Rossi et al. 2008, Watanabe et al. 2009, Gori et al. 2010). This may be because shallow habitats are more severe due to water movement and competition with algae (Gori et al. 2011) compared with the relatively stable deeper environments.
In the 0-25m depth interval, the species composition was distributed more homogeneously among sites than in the deep strata (40-70m). This suggests a high level of ecological connectivity through superficial water circulation between sites, as has been reported for the Campeche Bank (Jordán-Dahlgren 2002), or by a long dispersal range of larvae. In contrast, at the deeper strata (40-70m), 61% of the species seem to be site specific, indicating limited connectivity between sites, which may be due to short dispersal range of the larvae and settlement near the parental colony, as has been observed in Paramuricea clavata (Coma et al. 1995, Linares et al. 2007). Studies of tropical reefs have documented the effect of variability in the dispersal capability of larvae on the resulting spatial structure of gorgonian assemblage (Jordán-Dahlgren 2002). Quantifying the effect of larval dispersal on gorgonian distribution on the Pacific coast of Mexico requires further study of larval biology, population biology, as well as the oceanographic processes in the study area.
Assemblage structure in relation to depth (5-25m) and sites: The main species that characterize the gorgonian assemblages at the 5-25m depth interval were L. cuspidata, L. ena, L. alba and L. rigida, because they were the most frequent and abundant species in all sampling sites. These assemblages with few species may be due to high environmental severity conditions of the study area related to high wave regime (Young 1999, Chen et al. 2002), high turbidity and sedimentation by river discharge (Glynn & Leyte-Morales 1997, Granja-Fernández & López-Pérez 2008) that may limit the presence of many species.
The main four gorgonian species exhibit different spatial distribution patterns along the 5-25m bathymetric profile. L. cuspidata and L. ena define the shallower (5-10m) sites and L. alba the deeper (20-25m) sites. These bathymetrical distribution pattern is not directly related to light because their tissues do not contain symbiotic algae (pers. observ.) as has been observed in all species reported for the Tropical eastern Pacific (Breedy & Guzmán 2003, 2007, 2009). According to Fabricius & De’ath (2008), gorgonians that do not have symbionts do not require light, but instead depend on the flow of water to bring phytoplankton and other suspended food particles to their tentacles. The study area is a region of upwelling so there is enough food available to maintain great abundance of gorgonians.
A number of previous studies have demonstrated that wave action is an important factor in determining the zoning patterns of gorgonians in shallow habitats (Kinzie 1973, Opresko 1973, Birkeland 1974, Dinesen 1983, Botero 1987, Yoshioka & Yoshioka 1989, Sánchez et al. 1998). The greatest abundance of L. cuspidata and L. ena were found at depths of 5 and 10m, suggesting that they are well suited to this specific environment, perhaps due to their ability to grip the substrate. Their strong grip helps the colonies avoid detachment by wave action, which is one of the most significant factors in gorgonian mortality (Grigg 1975, Yoshioka & Yoshioka 1991).
Leptogorgia alba was the only species present at all depths, but it was more conspicuous at the deeper areas. Its extensive distribution might be related to its tolerance to a wide range of environmental conditions, as has been observed for Eunicella singularis in the Mediterranean Sea (Gori et al. 2010). Its low abundance at 5 and 10m suggest that in this environment this species is less tolerant or efficient than L. cuspidata and L. ena. Several studies note that larvae may reach different depths and then not survive due to specific forms of competition (La Barre et al. 1986, Dai 1990).
Further studies of environmental or biological factors that affect the distribution and relative abundance of gorgonian species, like type of substrate, coverage of substrate, and sedimentation in the Tropical eastern Pacific, are required to explain the abundance and distribution patterns of gorgonians species at different sites and depths.
Acknowledgments
We thank valeria Hernández Urraca, Imelda Pérez Maldonado, Fernando Díaz and Gonzáles for assistance in the field. This manuscript was greatly improved by comments from Eric Jordán Dahlgren, Rosa Rodríguez Martínez, Lorenzo Álvarez Filip and Andrés Ramón López Pérez. This work was supported by the Smithsonian Tropical Research Institute, Panama and Universidad del Mar, Puerto Angel, México.
References
Barrie, J.v. & K.W. Conway. 2002. Rapid sea-level change and coastal evolution on the Pacific margin of Canada. Sediment. Geol. 150: 171-183. [ Links ]
Bayer, F.M. 1961. The shallow water Octocorallia of the West Indian region. Martinus Nijhoff, The Hague, Holland. [ Links ]
Birkeland, C. 1974. The effect of wave action on the population dynamics of Gorgonia ventalina Linnaeus. Stud. Trop. Oceanogr. 12: 115-126. [ Links ]
Botero, L. 1987. Zonación de octocorales gorgonáceos en el área de Santa Marta y Parque Nacional Tayrona, Costa Caribe Colombiana. An. Inst. Inv. Mar. Punta de Betin 17: 61-80. [ Links ]
Breedy, O. 2009. Octocorals, p. 161-167. In I.S. Wehrtmann & J. Cortés (eds.). Marine biodiversity of Costa Rica, Central America. Monographiae Biologicae 86. Springer verlag, Berlin, Germany. [ Links ]
Breedy, O., R. Abeytia & H.M. Guzman. 2012. A new species of Leptogorgia (Cnidaria: Anthozoa: Octocorallia) from the Mexican Pacific coast. Bull. Mar. Sci. 88: 319-325. [ Links ]
Breedy, O. & J. Cortés. 2008. Octocorals (Coelenterata: Anthozoa: Octocorallia) of Isla del Coco, Costa Rica. Rev. Biol. Trop. 56: 71-77. [ Links ]
Breedy, O. & H.M. Guzmán. 2002. A revision of the genus Pacifigorgia (Coelenterata: Octocorallia: Gorgoniidae). Proc. Bio. Soc. Wash. 115: 782-839. [ Links ]
Breedy, O. & H.M. Guzmán. 2003. Octocorals from Costa Rica: the genus Pacifigorgia (Coelenterata: Octocorallia: Gorgoniidae). Zootaxa 281: 1-60. [ Links ]
Breedy, O. & H.M. Guzmán. 2007. A revision of the genus Leptogorgia Milne Edwards & Haime, 1857 (Coelenterata: Octocorallia: Gorgoniidae) in the eastern Pacific. Zootaxa 1419: 1-90. [ Links ]
Breedy, O. & H.M. Guzmán. 2009. A revision of the genus Eugorgia verril, 1868 (Coelenterata: Octocorallia: Gorgoniidae). Zootaxa 2151: 1-46. [ Links ]
Breedy, O., H.M. Guzman & S. vargas. 2009. A revision of the genus Eugorgia verrill, 1868 (Coelenterata: Octocorallia: Gorgoniidae. Zootaxa 2151: 1-46. [ Links ]
Chen, G., B. Chapron, R. Ezraty & D. vandemark. A global view of swell and wind sea climate in the ocean by satellite altimeter and scatterometer. 2002. J. Atmos. Oceanic Technol. 19: 1849-1859. [ Links ]
Clarke, K.R. 1993. Non-parametric multivariate analysis of changes in community structure. Aust. J. Ecol. 18: 117-143. [ Links ]
Clarke, K.R. & R.H. Green. 1988. Statistical design and analysis for a ‘biological effects’ study. Mar. Ecol. Prog. Ser. 46: 213-226. [ Links ]
Clarke, K.R., P.J. Somerfield & M.G. Chapman. 2006. On resemblance measures for ecological studies, including taxonomic dissimilarities and a Zero-adjusted Bray-Curtis coefficient for denuded assemblages. J. Exp. Mar. Biol. Ecol. 330: 55-80. [ Links ]
Coma, R., M. Ribes, M. Zabala & J. Gili. 1995. Reproduction and cycle of gonadal development in the Mediterranean gorgonian Paramuricea clavata. Mar. Ecol. Prog. Ser. 117: 173-183. [ Links ]
Dai, C.F. 1991. Distribution and adaptive strategies of Alcyonacean corals in Nanwan Bay, Taiwan. Hydrobiologia 216/217: 241-246. [ Links ]
Dinesen, Z. 1983. Patterns in the distribution of soft corals across the central Great Barrier Reef. Coral Reefs 1: 229-236. [ Links ]
Fabricius, K.E. & G. De’ath. 2008. Photosynthetic symbionts and energy supply determine octocoral biodiversity in coral reefs. Ecology 89: 3163-3173. [ Links ]
Gerhardt, D.J. 1990. Fouling and gastropod predation: consequences of grazing from the octocoral. Mar. Ecol. Prog. Ser. 62: 103-108. [ Links ]
Glynn, P.W. & G.E. Leyte-Morales. 1997. Coral reefs of Huatulco, West México: Reef development in upwelling Gulf of Tehuantepec. Rev. Biol. Trop. 45: 1033-1047. [ Links ]
Gori, A., S. Rossi, E. Berganzo, J.L. Pretus, M.R.T. Dale & J.M. Gili. 2010. Spatial distributions patterns of the gorgonians Eunicella singularis, Paramuricea clavata and Leptogorgia sarmentosa (Cap de Creus, northwestern Mediterranean Sea). Mar. Biol. 158: 143-158. [ Links ]
Gori, A., S. Rossi, C. Linares, E. Berganzo, C. Orejas, M.R.T. Dale & J.M. Gili. 2011. Size and spatial structure in deep versus shallow populations of the Mediterranean gorgonian Eunicella singularis (Cap de Creus, northwestern Mediterranean Sea). Mar. Biol. 158: 1721-1732. [ Links ]
Granja-Fernández, M.R. & R.A. López-Pérez. 2008. Sedimentación en comunidades arrecifales de Bahías de Huatulco, Oaxaca, México. Rev. Biol. Trop. 56: 1179-1187. [ Links ]
Grigg, R.W. 1975. Age structure of a longevous coral: A relative index of habitat suitability and stability. Am. Nat. 109: 647-657. [ Links ]
Gutiérrez, M., T.L. Capson, H.M. Guzmán, E. Quiñoa & R. Riguera. 2004. L-galactosa as a natural product: isolation from a marine octocoral of the first a-Lgalactosyl saponin. Tetrahedron Lett. 45: 7833-7836. [ Links ]
Gutiérrez, M., T.L. Capson, H.M. Guzmán, J. González, E. Ortega-Barria, E. Quiñoa & R. Riguera. 2005. Leptolide, a new furanocembranolide diterpene from Leptogorgia alba. J. Nat. Prod. 68: 614-616. [ Links ]
Gutiérrez, M., T.L. Capson, H.M. Guzmán, J. González, E. Ortega-Barria, E. Quiñoa & R. Riguera. 2006. Antiplasmodial metabolites isolated from the marine octocoral Muricea austera. J. Nat. Prod. 69: 1379-1383. [ Links ]
Guzmán, H.M., C.A. Guevara & O. Breedy. 2004. Distribution, diversity and conservation of coral reefs and coral communities in the largest marine protected area of Pacific Panama (Coiba Island). Envir. Conserv. 31: 111-121. [ Links ]
Guzmán, H.M., S. Benfield, O. Breedy & J. Mair. 2008. Broadening reef protection across the marine conservation corridor of the Eastern Tropical Pacific: distribution and diversity of reefs in Las Perlas Archipelago, Panama. Envir. Conser. 35: 46-54. [ Links ]
Hernández-Muñoz, D., P.M. Alcolado & M. Hernández-González. 2008. Efectos de un emisario submarino de aguas “residuales” urbanas sobre comunidades de octocorales (Octocorallia: Alcyonacea) en Cuba. Rev. Biol. Trop. 56: 65-75. [ Links ]
Jordán-Dahlgren, E. 2002. Gorgonian distribution patterns in coral reef environments of the Gulf of Mexico: evidence of sporadic ecological connectivity? Coral Reefs 21: 205-215. [ Links ]
Kinzie, R.A. 1973. The zonation of the West Indian gorgonians. Bull. Mar. Sci. 23: 95-155. [ Links ]
La Barre, S., J.C. Coll & P.W. Sammarco. 1986. Defensive strategies of soft corals (Coelenterata: Octocorallia) of the Great Barrier Reef. II. The relationship between toxicity and feeding deterrence. Biol. Bull. 171: 565-576. [ Links ]
Lasker, H. 1985. Prey preferences and browsing pressure of the butterflyfish Chaetodon capistratus on Caribbean gorgonians. Mar. Ecol. Prog. Ser. 21: 213-220. [ Links ]
Linares, C., D. Doak, R. Coma, D. Díaz & M. Zabala. 2007. Life history and population viability of a longlived marine invertebrate: the octocoral Paramuricea clavata. Ecology 88: 918-928. [ Links ]
Matsumoto, A.K., F. Iwase, Y. Imahara & H. Namikawa. 2007. Bathymetric distribution and biodiversity of deep-water octocorals (Coelenterata: Octocorallia) in Sagami Bay and adjacent waters of Japan, p. 231-251. In G. Robert & S. Cairns (eds.). Conservation and adaptive management of seamount and deep-sea coral ecosystems, Rosenstiel School of Marine and Atmospheric Science, University of Miami, USA. [ Links ]
McEnroe, F.J. & W. Fenical. 1978. Structures and synthesis of some new antibacterial sesquiterpenoids from the gorgonian coral Pseudopterogorgia rigida. Tetrahedron 34: 1661-1664. [ Links ]
Opresko, D.M. 1973. Abundance and distribution of shallow-water gorgonians in the area of Miami, Florida. Bull. Marine Sci. 23: 535-558. [ Links ]
Prahl, H.v., D. Escobar & G. Molina. 1987. Octocorales (Octocorallia: Gorgoniidae y Plexauridae) de aguas someras del Pacífico Colombiano. Rev. Biol. Trop. 34: 13-33. [ Links ]
Reyes-Bonilla, H., F. Sinsel-Duarte & O. Arizpe. 1997. Gorgonias y corales pétreos (Anthozoa: Gorgonacea y Scleractinia) de Cabo Pulmo, México. Rev. Biol. Trop. 45: 1439-1443. [ Links ]
Rodríguez, A.D. & C. Ramírez. 1994. Further butenolides from the Caribbean octocoral Pterogorgia citrina. J. Nat. Prod. 7: 339-347. [ Links ]
Rossi, S., G. Tsounis, C. Orejas, T. Padrón, J.M. Gili, L. Bramanti, N. Teixidó & J. Gutt. 2008. Survey of deep-dwelling red coral (Corallium rubrum) populations at Cap de Creus (NW Mediterranean). Mar. Biol. 154: 533-545. [ Links ]
Ruiz, L. & M. Rada. 2006. Octocorales de las aguas profundas del Oriente de venezuela. Invest Mar. valparaiso 34: 71-79. [ Links ]
Sánchez, J.A. 1999. Black coral-octocoral distribution patterns on Imelda Bank, a deep-water reef, Colombia, Caribbean Sea. Bull. Mar. Sci. 65: 215-225. [ Links ]
Sánchez, J.A., S. Zea & J.M. Díaz. 1998. Patterns of octocoral and black coral distribution in the oceanic barrier reef-complex of Providencia Island, Southwestern Caribbean. Caribb. J. Sci. 34: 250-264. [ Links ]
vreeland, H.v. & H.R. Lasker. 1989. Selective feeding of the polychaete Hermodice carunculata Pallas on Caribbean gorgonians. J. Exp. Mar. Biol. Ecol. 129: 265-267. [ Links ]
Watanabe, S., A. Metaxas, J. Sameoto & P. Lawton. 2009. Patterns in abundance and size in two deep-water gorgonian octocorals, in relation to depth and substrate features off Nova Scotia. Deep-Sea Res. 56: 2235-2248. [ Links ]
Yoshioka, P.M. & B.B. Yoshioka. 1989. Effects of wave energy, topographic relief and sediment transport on the distribution of shallow-water gorgonians of Puerto Rico. Coral Reefs 8: 145-152. [ Links ]
Yoshioka, P.M. & B.B. Yoshioka. 1991. A comparison of the survivorship and growth of shallow-water gorgonian species of Puerto Rico. Mar. Ecol. Prog. Ser. 69: 253-260. [ Links ]
Young, I.R. 1999. Seasonal variability of the global ocean wind and wave climate. Int. J. Climatol. 19: 931-950. [ Links ]
Zea, S. 1993. Cover of sponges and other sessile organisms in rocky and coral reefs habitats, of Santa Mata, Colombian Caribbean Sea. Carib. J. Sci. 29: 75-88. [ Links ]
Bayer, F.M. 1961. The shallow water Octocorallia of the West Indian region. Martinus Nijhoff, The Hague, Holland. [ Links ]
Birkeland, C. 1974. The effect of wave action on the population dynamics of Gorgonia ventalina Linnaeus. Stud. Trop. Oceanogr. 12: 115-126. [ Links ]
Botero, L. 1987. Zonación de octocorales gorgonáceos en el área de Santa Marta y Parque Nacional Tayrona, Costa Caribe Colombiana. An. Inst. Inv. Mar. Punta de Betin 17: 61-80. [ Links ]
Breedy, O. 2009. Octocorals, p. 161-167. In I.S. Wehrtmann & J. Cortés (eds.). Marine biodiversity of Costa Rica, Central America. Monographiae Biologicae 86. Springer verlag, Berlin, Germany. [ Links ]
Breedy, O., R. Abeytia & H.M. Guzman. 2012. A new species of Leptogorgia (Cnidaria: Anthozoa: Octocorallia) from the Mexican Pacific coast. Bull. Mar. Sci. 88: 319-325. [ Links ]
Breedy, O. & J. Cortés. 2008. Octocorals (Coelenterata: Anthozoa: Octocorallia) of Isla del Coco, Costa Rica. Rev. Biol. Trop. 56: 71-77. [ Links ]
Breedy, O. & H.M. Guzmán. 2002. A revision of the genus Pacifigorgia (Coelenterata: Octocorallia: Gorgoniidae). Proc. Bio. Soc. Wash. 115: 782-839. [ Links ]
Breedy, O. & H.M. Guzmán. 2003. Octocorals from Costa Rica: the genus Pacifigorgia (Coelenterata: Octocorallia: Gorgoniidae). Zootaxa 281: 1-60. [ Links ]
Breedy, O. & H.M. Guzmán. 2007. A revision of the genus Leptogorgia Milne Edwards & Haime, 1857 (Coelenterata: Octocorallia: Gorgoniidae) in the eastern Pacific. Zootaxa 1419: 1-90. [ Links ]
Breedy, O. & H.M. Guzmán. 2009. A revision of the genus Eugorgia verril, 1868 (Coelenterata: Octocorallia: Gorgoniidae). Zootaxa 2151: 1-46. [ Links ]
Breedy, O., H.M. Guzman & S. vargas. 2009. A revision of the genus Eugorgia verrill, 1868 (Coelenterata: Octocorallia: Gorgoniidae. Zootaxa 2151: 1-46. [ Links ]
Chen, G., B. Chapron, R. Ezraty & D. vandemark. A global view of swell and wind sea climate in the ocean by satellite altimeter and scatterometer. 2002. J. Atmos. Oceanic Technol. 19: 1849-1859. [ Links ]
Clarke, K.R. 1993. Non-parametric multivariate analysis of changes in community structure. Aust. J. Ecol. 18: 117-143. [ Links ]
Clarke, K.R. & R.H. Green. 1988. Statistical design and analysis for a ‘biological effects’ study. Mar. Ecol. Prog. Ser. 46: 213-226. [ Links ]
Clarke, K.R., P.J. Somerfield & M.G. Chapman. 2006. On resemblance measures for ecological studies, including taxonomic dissimilarities and a Zero-adjusted Bray-Curtis coefficient for denuded assemblages. J. Exp. Mar. Biol. Ecol. 330: 55-80. [ Links ]
Coma, R., M. Ribes, M. Zabala & J. Gili. 1995. Reproduction and cycle of gonadal development in the Mediterranean gorgonian Paramuricea clavata. Mar. Ecol. Prog. Ser. 117: 173-183. [ Links ]
Dai, C.F. 1991. Distribution and adaptive strategies of Alcyonacean corals in Nanwan Bay, Taiwan. Hydrobiologia 216/217: 241-246. [ Links ]
Dinesen, Z. 1983. Patterns in the distribution of soft corals across the central Great Barrier Reef. Coral Reefs 1: 229-236. [ Links ]
Fabricius, K.E. & G. De’ath. 2008. Photosynthetic symbionts and energy supply determine octocoral biodiversity in coral reefs. Ecology 89: 3163-3173. [ Links ]
Gerhardt, D.J. 1990. Fouling and gastropod predation: consequences of grazing from the octocoral. Mar. Ecol. Prog. Ser. 62: 103-108. [ Links ]
Glynn, P.W. & G.E. Leyte-Morales. 1997. Coral reefs of Huatulco, West México: Reef development in upwelling Gulf of Tehuantepec. Rev. Biol. Trop. 45: 1033-1047. [ Links ]
Gori, A., S. Rossi, E. Berganzo, J.L. Pretus, M.R.T. Dale & J.M. Gili. 2010. Spatial distributions patterns of the gorgonians Eunicella singularis, Paramuricea clavata and Leptogorgia sarmentosa (Cap de Creus, northwestern Mediterranean Sea). Mar. Biol. 158: 143-158. [ Links ]
Gori, A., S. Rossi, C. Linares, E. Berganzo, C. Orejas, M.R.T. Dale & J.M. Gili. 2011. Size and spatial structure in deep versus shallow populations of the Mediterranean gorgonian Eunicella singularis (Cap de Creus, northwestern Mediterranean Sea). Mar. Biol. 158: 1721-1732. [ Links ]
Granja-Fernández, M.R. & R.A. López-Pérez. 2008. Sedimentación en comunidades arrecifales de Bahías de Huatulco, Oaxaca, México. Rev. Biol. Trop. 56: 1179-1187. [ Links ]
Grigg, R.W. 1975. Age structure of a longevous coral: A relative index of habitat suitability and stability. Am. Nat. 109: 647-657. [ Links ]
Gutiérrez, M., T.L. Capson, H.M. Guzmán, E. Quiñoa & R. Riguera. 2004. L-galactosa as a natural product: isolation from a marine octocoral of the first a-Lgalactosyl saponin. Tetrahedron Lett. 45: 7833-7836. [ Links ]
Gutiérrez, M., T.L. Capson, H.M. Guzmán, J. González, E. Ortega-Barria, E. Quiñoa & R. Riguera. 2005. Leptolide, a new furanocembranolide diterpene from Leptogorgia alba. J. Nat. Prod. 68: 614-616. [ Links ]
Gutiérrez, M., T.L. Capson, H.M. Guzmán, J. González, E. Ortega-Barria, E. Quiñoa & R. Riguera. 2006. Antiplasmodial metabolites isolated from the marine octocoral Muricea austera. J. Nat. Prod. 69: 1379-1383. [ Links ]
Guzmán, H.M., C.A. Guevara & O. Breedy. 2004. Distribution, diversity and conservation of coral reefs and coral communities in the largest marine protected area of Pacific Panama (Coiba Island). Envir. Conserv. 31: 111-121. [ Links ]
Guzmán, H.M., S. Benfield, O. Breedy & J. Mair. 2008. Broadening reef protection across the marine conservation corridor of the Eastern Tropical Pacific: distribution and diversity of reefs in Las Perlas Archipelago, Panama. Envir. Conser. 35: 46-54. [ Links ]
Hernández-Muñoz, D., P.M. Alcolado & M. Hernández-González. 2008. Efectos de un emisario submarino de aguas “residuales” urbanas sobre comunidades de octocorales (Octocorallia: Alcyonacea) en Cuba. Rev. Biol. Trop. 56: 65-75. [ Links ]
Jordán-Dahlgren, E. 2002. Gorgonian distribution patterns in coral reef environments of the Gulf of Mexico: evidence of sporadic ecological connectivity? Coral Reefs 21: 205-215. [ Links ]
Kinzie, R.A. 1973. The zonation of the West Indian gorgonians. Bull. Mar. Sci. 23: 95-155. [ Links ]
La Barre, S., J.C. Coll & P.W. Sammarco. 1986. Defensive strategies of soft corals (Coelenterata: Octocorallia) of the Great Barrier Reef. II. The relationship between toxicity and feeding deterrence. Biol. Bull. 171: 565-576. [ Links ]
Lasker, H. 1985. Prey preferences and browsing pressure of the butterflyfish Chaetodon capistratus on Caribbean gorgonians. Mar. Ecol. Prog. Ser. 21: 213-220. [ Links ]
Linares, C., D. Doak, R. Coma, D. Díaz & M. Zabala. 2007. Life history and population viability of a longlived marine invertebrate: the octocoral Paramuricea clavata. Ecology 88: 918-928. [ Links ]
Matsumoto, A.K., F. Iwase, Y. Imahara & H. Namikawa. 2007. Bathymetric distribution and biodiversity of deep-water octocorals (Coelenterata: Octocorallia) in Sagami Bay and adjacent waters of Japan, p. 231-251. In G. Robert & S. Cairns (eds.). Conservation and adaptive management of seamount and deep-sea coral ecosystems, Rosenstiel School of Marine and Atmospheric Science, University of Miami, USA. [ Links ]
McEnroe, F.J. & W. Fenical. 1978. Structures and synthesis of some new antibacterial sesquiterpenoids from the gorgonian coral Pseudopterogorgia rigida. Tetrahedron 34: 1661-1664. [ Links ]
Opresko, D.M. 1973. Abundance and distribution of shallow-water gorgonians in the area of Miami, Florida. Bull. Marine Sci. 23: 535-558. [ Links ]
Prahl, H.v., D. Escobar & G. Molina. 1987. Octocorales (Octocorallia: Gorgoniidae y Plexauridae) de aguas someras del Pacífico Colombiano. Rev. Biol. Trop. 34: 13-33. [ Links ]
Reyes-Bonilla, H., F. Sinsel-Duarte & O. Arizpe. 1997. Gorgonias y corales pétreos (Anthozoa: Gorgonacea y Scleractinia) de Cabo Pulmo, México. Rev. Biol. Trop. 45: 1439-1443. [ Links ]
Rodríguez, A.D. & C. Ramírez. 1994. Further butenolides from the Caribbean octocoral Pterogorgia citrina. J. Nat. Prod. 7: 339-347. [ Links ]
Rossi, S., G. Tsounis, C. Orejas, T. Padrón, J.M. Gili, L. Bramanti, N. Teixidó & J. Gutt. 2008. Survey of deep-dwelling red coral (Corallium rubrum) populations at Cap de Creus (NW Mediterranean). Mar. Biol. 154: 533-545. [ Links ]
Ruiz, L. & M. Rada. 2006. Octocorales de las aguas profundas del Oriente de venezuela. Invest Mar. valparaiso 34: 71-79. [ Links ]
Sánchez, J.A. 1999. Black coral-octocoral distribution patterns on Imelda Bank, a deep-water reef, Colombia, Caribbean Sea. Bull. Mar. Sci. 65: 215-225. [ Links ]
Sánchez, J.A., S. Zea & J.M. Díaz. 1998. Patterns of octocoral and black coral distribution in the oceanic barrier reef-complex of Providencia Island, Southwestern Caribbean. Caribb. J. Sci. 34: 250-264. [ Links ]
vreeland, H.v. & H.R. Lasker. 1989. Selective feeding of the polychaete Hermodice carunculata Pallas on Caribbean gorgonians. J. Exp. Mar. Biol. Ecol. 129: 265-267. [ Links ]
Watanabe, S., A. Metaxas, J. Sameoto & P. Lawton. 2009. Patterns in abundance and size in two deep-water gorgonian octocorals, in relation to depth and substrate features off Nova Scotia. Deep-Sea Res. 56: 2235-2248. [ Links ]
Yoshioka, P.M. & B.B. Yoshioka. 1989. Effects of wave energy, topographic relief and sediment transport on the distribution of shallow-water gorgonians of Puerto Rico. Coral Reefs 8: 145-152. [ Links ]
Yoshioka, P.M. & B.B. Yoshioka. 1991. A comparison of the survivorship and growth of shallow-water gorgonian species of Puerto Rico. Mar. Ecol. Prog. Ser. 69: 253-260. [ Links ]
Young, I.R. 1999. Seasonal variability of the global ocean wind and wave climate. Int. J. Climatol. 19: 931-950. [ Links ]
Zea, S. 1993. Cover of sponges and other sessile organisms in rocky and coral reefs habitats, of Santa Mata, Colombian Caribbean Sea. Carib. J. Sci. 29: 75-88. [ Links ]
*Correspondencia a:
Rosalinda Abeytia. Unidad Académica Puerto Morelos. Instituto de Ciencias del Mar y Limnología, Universidad Nacional Autónoma de México, Ap. Postal 1152, 77500 Cancún, QR 77500, México; r.abeytia@gmail.com
Héctor M. Guzmán. Smithsonian Tropical Research Institute, Apartado 0843-03092, Balboa, República de Panamá; GuzmanH@si.edu
Odalisca Breedy. Centro de Investigación en Ciencias del Mar y Limnología, Centro de Investigación en Estructuras Microscópicas, Universidad de Costa Rica, San José, Costa Rica; odalisca.breedy@ucr.ac.cr
1. Unidad Académica Puerto Morelos. Instituto de Ciencias del Mar y Limnología, Universidad Nacional Autónoma de México, Ap. Postal 1152, 77500 Cancún, QR 77500, México; r.abeytia@gmail.com
2. Smithsonian Tropical Research Institute, Apartado 0843-03092, Balboa, República de Panamá; GuzmanH@si.edu
3. Centro de Investigación en Ciencias del Mar y Limnología, Centro de Investigación en Estructuras Microscópicas, Universidad de Costa Rica, San José, Costa Rica; odalisca.breedy@ucr.ac.cr
Received 29-V-2012. Corrected 01-XI-2012. Accepted 28-XI-2012


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}