










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.61 n.3 San José Sep. 2013
Morpho-anatomy, imbibition, viability and germination of the seed of Anadenanthera colubrina var. cebil (Fabaceae)
Morfo-anatomía, imbibición, viabilidad y germinación de las semillas de Anadenanthera colubrina var. cebil (Fabaceae)
Morfo-anatomía, imbibición, viabilidad y germinación de las semillas de Anadenanthera colubrina var. cebil (Fabaceae)
*Dirección para correspondencia:
Abstract
Seed biology is a relevant aspect of tropical forests because it is central to the understanding of processes of plant establishment, succession and natural regeneration. Anadenanthera colubrina var. cebil is a timber tree from South America that produces large seeds with thin weak teguments, which is uncommon among legumes. This study describes the morphology and anatomy of the seed coat, the viability, imbibition, and germination in this species. Seeds used during the essays came from 10 trees that grow naturally in Horco Molle, province of Tucumán, Argentina. Seed morphology was described from a sample of 20 units. The seed coat surface was examined with a scanning electron microscope. Transverse sections of hydrated and non-hydrated seeds were employed to describe the histological structure of the seed coat. Hydration, viability and germination experiments were performed under laboratory controlled conditions; and the experimental design consisted of 10 replicas of 10 seeds each. Viability and germination tests were conducted using freshly fallen seeds and seeds stored for five months. Morphologically the seeds of A. colubrina var. cebil are circular to subcircular, laterally compressed, smooth, bright brown and have a horseshoe fissure line (=pleurogram) on both sides. The seed coat comprises five tissue layers and a double (external and internal) cuticle. The outer cuticle (on the epidermis) is smooth and interrupted by microcracks and pores of variable depth. The epidermis consists of macroesclereids with non-lignified secondary walls. This layer is separated from the underlying ones during seed hydration. The other layers of internal tissues are comprised of osteosclereids, parenchyma, osteosclereids, and macrosclereids. The percentage of viable seeds was 93%, decreasing to 75% in seeds with five months old. Seed mass increased 76% after the first eight hours of hydration. Germination percentage was 75% after 76 hours. Germination of seeds stored for five months decreased to 12%. The results showed that seeds of A. colubrina var. cebil are highly permeable and germinate directly without a dormant period.
Key words: Anadenanthera, Fabaceae, seed, anatomy, imbibition, viability, germination.
Resumen
El cebil, Anadenanthera colubrina var. cebil (Griseb.) Altschul, es un árbol forestal de América del Sur con grandes semillas de tegumentos delgados y débiles. Este estudio describe la morfología y anatomía de la cubierta seminal del cebil y evalúa la viabilidad, la imbibición y germinación de las semillas. Morfológicamente las semillas son circulares a subcirculares, comprimidas lateralmente, lisas, marrón lustrosas, con una línea fisural en forma de herradura en ambas caras. Histológicamente la cubierta seminal comprende cinco capas de células y una cutícula externa e interna. La cutícula externa es lisa y presenta microfisuras y poros de profundidad variable. La epidermis consiste de un estrato de macroesclereidas con paredes secundarias no lignificadas. Los siguientes tejidos internos incluyen, osteosclereidas, parénquima, osteoscleridas y macroesceleridas. El porcentaje de semillas viables fue del 93%, disminuyendo al 75% en semillas con cinco meses de almacenamiento. Las semillas incrementaron el 76% de su masa durante las primeras 8 horas de hidratación. El porcentaje de germinación en semillas nuevas fue del 75% al término de 76 horas y del 12% en semillas almacenadas por cinco meses. Los resultados confirman que las semillas de A. colubrina var. cebil son altamente permeables y germinan sin un período de reposo.
Palabras clave: Anadenanthera, Fabaceae, semilla, anatomía, imbibición, viabilidad, germinación.
Seed biology is among the most relevant research topics for tropical forests because it is central to understand community processes, such as plant establishment, succession and natural regeneration (Janzen & Vázquez-Yanes 1990, Lamprecht 1990, Vázquez-Yanes & Orozco-Segovia 1993). Likewise, this field of study is highly useful in forestry, forest conservation, restoration, and commercial tree production (Khurana & Singh 2001). Viability is an essential requirement for seeds to germinate and to establish seedlings (Baskin & Baskin 1998). Germination involves a series of events that begins with imbibition. Diffusion of water and gases into seeds is controlled by anatomical and chemical characteristics of the seed coat (Werker et al. 1979, Baskin & Baskin 1998, De Souza & Marcos-Filho 2001, Ma et al. 2004, Qutob et al. 2008). Seed coat can be categorized as either permeable or impermeable, depending on whether or not water is absorbed. A permeable or “soft” seed absorbs water rapidly (within minutes to hours), whereas an impermeable or “hard” seed does not imbibe water even after several days or weeks of being soaked and remains dormant (Ma et al. 2004, Shao et al. 2007). Hard seed production is a survival mechanism of plants from arid or desert regions, where rainfalls are very variable or unpredictable (Baskin & Baskin 1998).
Fabaceae (Leguminosae) are abundant components of tropical flora and propagate mostly through seeds (Burkart 1952). About 80% of fabaceous species produce hard seeds (Baskin & Baskin 1998, Baskin et al. 2000). Under natural conditions, these seeds germinate after long periods (within weeks to months), when coats become permeable due to opening of a water gap in response to environmental factors, especially temperature (Baskin & Baskin 1998, De Souza & Marcos-Filho 2001, Khurana & Singh 2001). Seed impermeability has been attributed to the presence of a continuous outer cuticle and, or an epidermis of packed macrosclereids (Werker et al. 1979, Baskin & Baskin 1998, De Souza & Marcos-Filho 2001, Ma et al. 2004, Qutob et al. 2008), which are strongly attached with hydrophobic components (Tran & Cavanagh 1984, Mohamed-Yassen et al. 1994). Actually, knowledge on fabaceous seeds is mostly related to hard seeds (Baskin & Baskin 1998, Baskin et al. 2000), and studies on soft seeds are scarce, with most of such studies focusing on species of agricultural interest, mainly soybean and bean (Pérez-Herrera & Acosta-Gallegos 2002, Yaklich et al. 1986, Ma et al. 2004, Capeleti et al. 2005, Shao et al. 2007, Qutob et al. 2008).
Anadenanthera colubrina var. cebil (Griseb.) Altschul, is a tree native to South America, distributed in Brazil, Bolivia, Paraguay, Uruguay, and Argentina, locally known as “cebil”, “cebil colorado”, “cebil moro”, and “curupái”. Its timber is hard and resistant to environmental conditions and has several applications (Tortorelli 1956, Erize 1997, Justiniano & Fredericksen 1998). Cebil is also appreciated for its ornamental and medicinal properties and as source of pollen for bees. In addition, the species is used in tanneries and its gum is used as a substitute for gum arabic (Erize 1997, Justiniano & Fredericksen 1998). The seeds have a thin and weak coat. Aside from a few studies on morphology (Boelcke 1946, Digilio & Legname 1966) and field germination (Justiniano & Fredericksen 1998, Fredericksen et al. 2000) there is not information on the anatomy and viability of the seeds of this species. This study aimed to describe the morphology and anatomy of the seed coat, the viability, imbibition, and germination in A. colubrina var. cebil under laboratory conditions.
Material and Methods
Seed sampling: In October 2009, fruits and seeds were collected from 10 A. colubrina var. cebil trees (hereafter “cebil”) growing naturally in the Horco Molle Experimental Reserve (26°38’ -26°57’ S and 65°26’ - 65°20’ W; 620m.a.s.l.) of the Universidad Nacional de Tucumán, Province of Tucumán, Argentina. The area is characterized by second-growth forests belonging to the Yungas ecoregion (Cabrera 1994), and is dominated by cebil and stands of exotic forests (Eucalyptus, Ligustrum, Pinus and others). In the laboratory, seeds were separated from fruits and examined under a magnifying glass. Intact seeds of similar size and color were selected; cracked, fragmented, aborted or insect-parasitized seeds were excluded. All the experiments were made in laboratory.
Experimental design: Hydration, viability and germination assays consisted of 10 replicates of 10 seeds each; each replicate corresponded to a different tree. For viability and germination, two tests were conducted, one with freshly fallen seeds collected in October 2009 (hereinafter “new seeds”) and the other with seeds stored for five months at a constant temperature of 20°C (hereinafter “stored seeds”).
Viability assays: Viability of cebil seeds was evaluated using a solution of 0.1% 2, 3, 5-triphenyl-tetrazolium chloride (TTC), which detects seed respiration and thus viability by staining the embryo pink to red (Moore 1972). The TTC test was applied following standard protocols (Varela & Bucher 2006). Only seeds stained bright pink were considered viable.
Imbibition assays: Groups of 10 seeds were weighed on an analytical balance and then immersed in tap water (10mL) in a Petri dish for 8h. Seeds were removed at 1h intervals, blotted with filter paper, weighed and immediately returned to the dish. Weight gain of seeds is an estimate of the amount of water absorbed (Bewley & Black 1982). The assays were conducted at a constant temperature of 25°C. At the end of the assay, seeds were classified as hydrated or non-hydrated. Volume of hydrated seeds was 50-80% larger than that of non-hydrated ones.
Germination assays: Germination assays were performed in a closed chamber at 32°C. Seeds were sterilized by immersing them in 1% sodium hypochloride (NaClO) for 10min and immediately rinsed with distilled water for 1min. Seeds were germinated in Petri dishes with filter paper soaked with distilled water; water was added as needed to keep the filter paper moisture. Germination was recorded three times a week for one month. Seeds were considered to be germinated when the radicle emerged from the seed coat. At the end of the experiment, non-germinated seeds were subjected to TTC to evaluate viability.
Seed morphology: Seed morphology was characterized from a sample of 20 seeds collected from 10 trees. The nomenclature provided by Boelcke (1946) was used to describe shape, position of hilum, lens and micropyle, and characteristics of the seed coat. Seed size (length, width and thickness) was determined with a digital calliper (Davidson) to the nearest 0.01mm. Seed length was measured from hilar to chalazal end; seed width and thicknesses were taken at the mid portion and the widest part, respectively, of the seed. Seed coat color was determined with the color chart provided by Munsell (2000). The surface of the seed coat was examined with a scanning electron microscope (Servicio de Microscopia Electrónica del Noroeste Argentino, LAMENOA).
Seed anatomy: Transverse sections of the seed coat of hydrated and non-hydrated seeds were prepared by hand. Sections were stained with safranin, cresyl violet (D’Ambrogio de Argüeso 1986) or safranin-astral blue (Bruno et al. 2007), and were mounted in 50% glycerol. Observations were made under a stereoscopic microscope and a compound microscope. Images were taken with a digital camera 10 megapixel.
Percent increase in seed mass was calculated as follows: [(W1- Wd)/Wd] x 100], where W1 and Wd are wet and dry mass, respectively (Turner & Dixon 2009). Viability percentage was expressed using the same descriptive statistic (average±SD). A curve of cumulative germination frequency (%) was expressed as a function of time.
Results
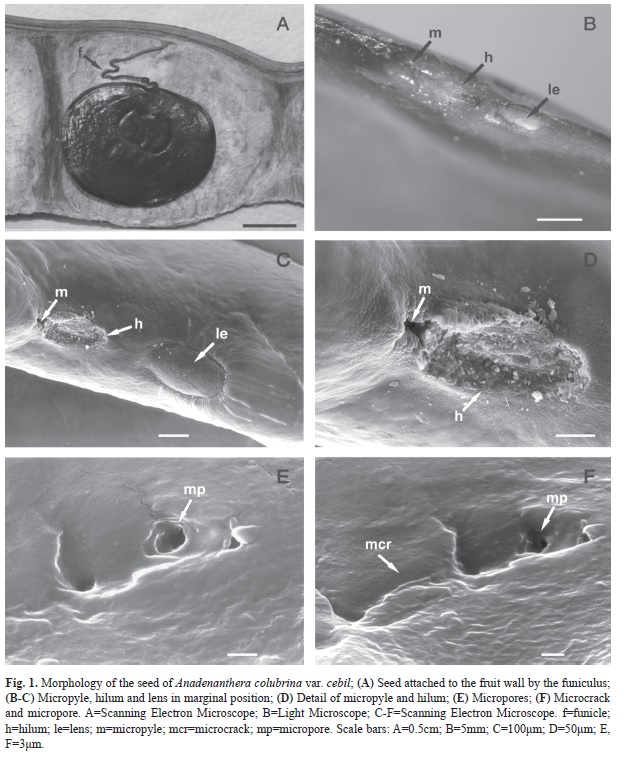
Seed morphology: Seeds 10-13 per fruit (average: 11.8; SD: 1.0), circular or subcircular, dorsoventrally compressed, with flat faces and very thin and sharp margins (Fig. 1A). Hilar and chalazar ends are rounded. Testa is light brown (dark red 5, sensu Munsell 2000) to dark brown (black 2, sensu Munsell 2000) in old seeds; surface smooth, bright; linea fissura (=pleurogram) on both faces, slightly depressed, horseshoe-shaped, covering 10% of the seed surface, arms of equal length. Funiculus is uniform thickness, dark brown (black 2 sensu Munsell 2000) (Fig. 1A). Hilum, micropyle and lens are marginally located (Fig. 1B, C). Hilum is elliptical, depressed; micropyle punctiform, closed (Fig. 1D); lens ellipsoidalshape, whitish (White 8/3 sensu Munsell 2000), near the hilum (Fig. 1B, C) and opposite the micropyle. External cuticle appears to be continuous under the magnifying glass, but scanning electron microscope reveals that it is discontinued by narrow microcracks and circular micropores of variable location, frequency and depth (Fig. 1E, F).
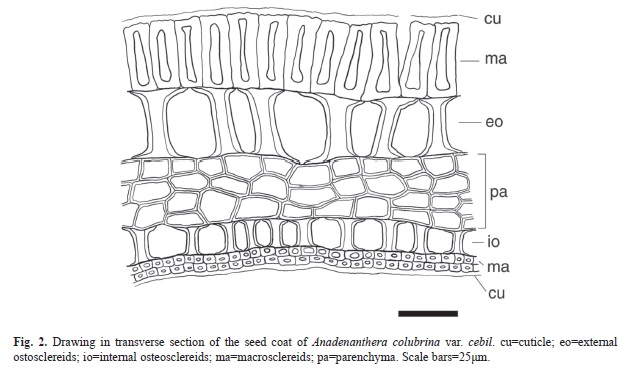
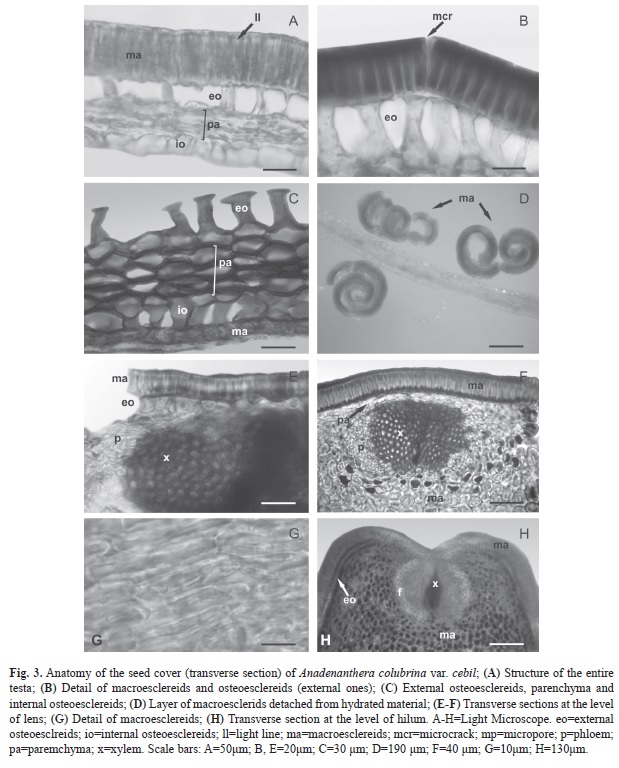
Seed coat anatomy: Transverse section of seed coat shows variable thickness, ranging between 150-205μm. The seed coat consists of five layers of tissues (Fig. 2, Fig. 3A) and a double external and internal cuticle. The outermost tissue layer is the epidermis (30-45μm), and it is composed of a stratum of columnar macrosclereids, with their main axis perpendicular to the surface. These cells are tightly bound to each other, without intercellular space, and with unevenly thickened walls that do not stain with safranin-astra blue, indicating that they are not lignified. A clear, narrow band, refractive to light, known as the light line, is observed near the apical end of these cells and parallel to the surface (Fig. 3A). The second layer is composed of a stratum of osteoscleroids. These cells have widened ends and thickened secondary walls and are separated from one another by air spaces (Fig. 3B, C). The third layer is composed of four to five strata of parenchyma cells, tangentially elongated to isodiametric and of thickened walls (Fig. 3A-C). The fourth layer consists of a stratum of osteoesclereids that are smaller than their outer counterparts (Fig. 3A-C). The fifth layer is composed of two or three strata of macroesclereids oriented perpendicularly to the epidermis layer (Fig. 3C). An inner cuticle separates the seed coat from endosperm.
Transverse sections of non-hydrated seeds were used to describe the complete structure of seed coat. In the hydrated seeds, the outer layer of macrosclereids (bound to cuticle) was separated from the underlying osteosclereids and broken off in fragments. Each fragment was curved like a curl, exposing the internal surface that was joined to the osteoesclereids. This condition partially hindered the complete view of the anatomical structure (Fig. 3D).
Transverse section of the lens showed an external stratum of macroesclereids that were smaller than those in the rest of the seed coat, followed by a stratum of osteoesclereids or of parenchyma tissue (with a vascular bundle composed of xylem of central position and lateral phloem), surrounded by multistratified macroesclereids (Fig. 3E-G). Histologically, in transverse section the hilum is similar to the lens, with a layer of osteoesclereids always below the macroesclereids (Fig. 3H).
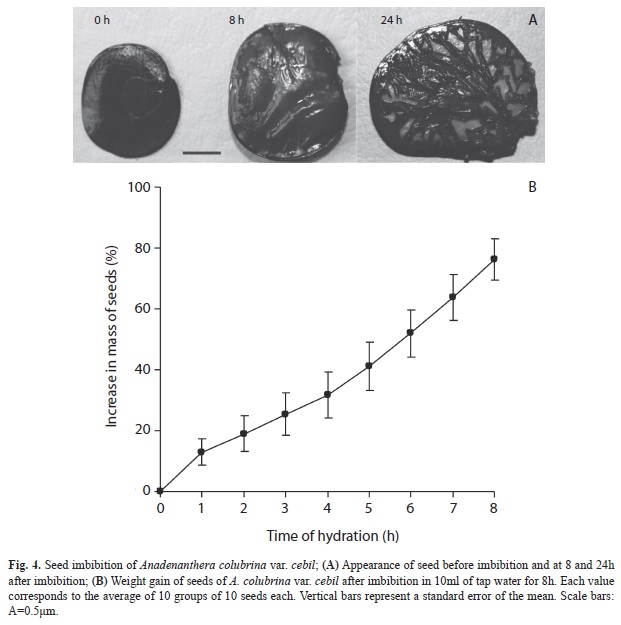
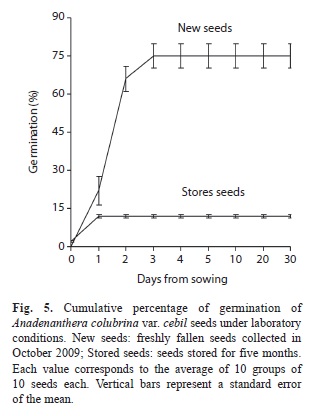
Seed viability, imbibition and germination: Mean percentage of viable seeds of cebil was 93% (SD=11%, minimum=70%, maximum=100%) in new seeds, and 75% (SD=24%, minimum=30%, maximum=100%) in seeds with five months of storage. This indicated a loss of viability of 18%. Seed mass increased 12.6% after 1h of the hydration experiment, and reached 76% after 8h (Fig. 4A), when almost doubled their initial mass (Fig. 4B). Germination in new seeds was 25% at 24h, reaching 73% at 72h (Fig. 5). In seeds stored for five months, germination percentage was 12% at 48h. At the end of the one-month assay, non-germinated seeds of both conditions (“new seeds” and “stored seeds”) were hydrated, had mucilaginous cotyledons and were not viable as shown by the TTC test.
Discussion
The seed coat of cebil shares some anatomical features with many other legumes, by exhibiting an epidermis of macroesclereids, a hypodermis of osteosclereids and an underlying parenchyma (Fahn 1985, De Souza & Marcos-Filho 2001). Cebil has a few unique features, such as the two internal layers, one consisting of osteosclereids and the other of macroesclerids.
The rapid hydration of cebil seeds indicates that the testa is highly water permeable. This trait is determined by the presence of micropores and microcracks in the epidermal cuticle, and by the weak attachment between the epidermal cells which lack lignin in their walls. The latter feature would explain the fragmentation of the epidermis plus cuticle and its separation from the underlying inner layers during hydration. The curling probably occurs because the macrosclereid layer was under great tension when the seed coat was dry. In seeds with five months of storage, where the epidermis should have been dewatered sufficiently to become waterproof, the macroscrelereids were also hidrated and curled as in the freshly fallen ones. Studies conducted in soybean cultivars showed that microcraks of the cuticle are determinant of seed permeability (Ma et al. 2004).
Discontinuity in the epidermal cuticle joined the detachment of the epidermal layer, which suggests the presence of multiple sites of water entry. This is in disagreement with findings reported for hard seeds of legumes, in which water penetrates through specific sites (hilum, micropyle, lens, strophiole, pleurogram, light line), where the seed coat is thinner than in the rest of the cuticle (Baskin & Baskin 1998).
The germination percentage of cebil found in this work was high (75%) and consistent with values reported in other studies (68-82%) for conspecifics from Brazil and Boliva (Justiniano & Fredericksen 1998). The high permeability of the cuticle makes seeds vulnerable to environmental fluctuations (e.g. temperature and humidity) and to the attack of microorganisms, shortening seed longevity (Mohamed-Yassen et al. 1994). Our data indicated that seeds stored for five months under laboratory conditions, lose 18% of their viability, and only 12% of these seeds have the potential to germinate. On the other hand, the data provided by Justiniano & Fredericksen (1998) indicated that only 32% of cebil seeds can survive after one month in Bolivia forests.
According to the characteristics of cebil seeds, such as high permeability, high germination, short longevity and anemochorous dispersal, the species is a pioneer plant, after the supporting findings reported by Justiniano & Fredericksen (1998). Pioneer plants have high light requirements and colonize clearings or open areas, generating suitable environmental conditions for the start of plant succession (Whitmore 1989, Vázquez-Yanes & Orozco-Segovia 1994). The seeds of cebil germinate under both light and dark conditions (Varela & Albornoz, pers. obs.), but juvenile and adult plants are more common in well-lit forest and disturbed sites. Because it is a heliophyte plant, it has a high lighting requirement for its regeneration (Justiniano & Fredricksen 1998). In the forests of Bolivia, cebil regeneration is highly variable among years, with maximum pulses in years of high seed production (Justiniano & Fredericksen 1998). The timely occurrence of rainfalls during the period of maximum seed rain of cebil (October-November) is likely a key factor for the regeneration of the species in Tucumán forests. The results of the present work allow us to conclude that seeds of A. colubrina var. cebil are highly permeable and germinate directly without a dormant period.
Acknowledgments
We thank Guillermo O. Lizardo for assistance in viability and germination assays, Alberto Gutiérrez and Inés Jaume for their help in the preparation of images, and Fundación Miguel Lillo that provided support for electron microscopy service. We also thank the anonymous reviewers for their timely comments and valuable suggestions. Jorgelina Brasca translated the manuscript into English.
References
Baskin, C.C. & J.M. Baskin. 1998. Seeds. Ecology, biogeography and evolution of dormancy and germination. San Diego Academic, California, USA. [ Links ]
Baskin, J.M., C.C. Baskin & X. Li. 2000. Taxonomy, anatomy and evolution of physical dormancy in seeds. Pl. Spec. Biol. 15: 139-152. [ Links ]
Bewley, J.D. & M. Black. 1982. Physiology and biochemistry of seeds in relation to germination. Viability, dormancy and environmental control. Springer-Verlag, Berlin, Germany. [ Links ]
Boelcke, O. 1946. Estudio morfológico de semillas de leguminosas Mimosoideas y Caesalpinoideas de interés agronómico en la Argentina. Darwiniana 31: 240-321. [ Links ]
Bruno, G., L. Stiefkens, M. Hadid, I. Liscovsky, M.T. Cosa & N. Dottori. 2007. Efecto de la contaminación ambiental en la anatomía de la hoja de Ligustrum lucidum (Oleaceae). Bol. Soc. Argent. Bot. 42: 231-236. [ Links ]
Burkart, A. 1952. Las leguminosas argentinas silvestres y cultivadas. Acme, Buenos Aires, Argentina. [ Links ]
Cabrera, A.L. 1994. Regiones fitogeográficas de Argentina. In W.F. Kugler (ed.). Enciclopedia Argentina de Agricultura y Jardinería. Acme, Buenos Aires, Argentina. [ Links ]
Capeleti, I., M.L. Ferrarese, F.C. Kryzanowski & O. Ferrarese-Filho. 2005. A new procedure for the quantification of lignin in soybean (Glycine max (L.) Merril) seed coat and their relationship with the resistance to mechanical damage. Seed Sci. Technol. 33: 511-515. [ Links ]
D’Ambrogio de Argüeso, A. 1986. Manual de técnicas en histología vegetal. Hemisferio Sur, Buenos Aires, Argentina. [ Links ]
De Souza, F.H.D. & J. Marcos-Filho. 2001. The seed coat as a modulator of seed-environment relationship in Fabaceae. Rev. Bras. Bot. 24: 365-375. [ Links ]
Digilio, A.P.L. & P.R. Legname. 1966. Los árboles indígenas de la Provincia de Tucumán. Ópera Lilloana XV, Tucumán, Argentina. [ Links ]
Erize, F. 1997. Nuevo libro del árbol. Tomo 1. Ateneo, Buenos Aires, Argentina. [ Links ]
Fahn, A. 1985. Anatomía vegetal. Pirámides, Buenos Aires, Argentina. [ Links ]
Fredericksen, T.S., M.J. Justiniano, B. Mostacedo, D. Kennard & L. McDonald. 2000. Comparative regeneration ecology of three leguminous timber species in a Bolivian tropical dry forest. New Forest 20: 45-64. [ Links ]
Janzen, D.H. & C. Vázquez-Yanes. 1990. Aspects of tropical seed ecology of relevance to management of tropical forested wildlands, p. 137-157. In A. GómezPompa, T.C. Whitmore & M. Hadley (eds.). Rain forest regeneration and management, UNESCO and Parthenon, Paris, France. [ Links ]
Justiniano, M.J. & T.S. Fredericksen. 1998. Ecología de especies menos conocidas: Curupaú -Anadenanthera colubrina. Proyecto BOLFOR, Santa Cruz, Bolivia. [ Links ]
Khurana, E. & J.S. Singh. 2001. Ecology of tree seed and seedlings: Implications for tropical forest conservation and restoration. Curr. Sci. 80: 748-757. [ Links ]
Lamprecht, H. 1990. Silvicultura en los trópicos. Los ecosistemas forestales en los bosques tropicales y sus especies arbóreas, posibilidades y métodos para un aprovechamiento sostenido. Deutsche Gesellschaft für Technische Zusammenarbeit (GTZ) GmbH, Eschborn, Germany. [ Links ]
Ma, F., E. Cholewa, T. Mohamed, C.A. Peterson & M. Gijzen. 2004. Craks in the palisade cuticule of soybean seed coats correlate with their permeability to water. Ann. Bot. 94: 213-228. [ Links ]
Mohamed-Yassen, Y., S.A. Barringer, W.E. Splittstoesser & S. Costanza. 1994. The role of the seeds coats in seed viability. Bot. Rev. 60: 246-260. [ Links ]
Moore, R.P. 1972. Tetrazolium staining for assessing seed quality, p. 347-366. In W. Heydecker (ed.). Seed ecology. University of Parth, London, England. [ Links ]
Munsell, A.H. 2000. Munsell soil color charts. Munsell Color Company, I.N.C. Baltimore, Maryland, USA. [ Links ]
Pérez-Herrera, P. & J.A. Acosta-Gallegos. 2002. Permeabilidad de la testa y la porción del micrópilo-hilio en semilla de frijol silvestre y cultivado. Rev. Fitotec. Mex. 25: 57-63. [ Links ]
Qutob, D., F. Ma, C.A. Peterson, M.A. Bernards & M. Gijzen. 2008. Structural and permeability properties of the soybean seed coat. Botany 86: 219-227. [ Links ]
Shao, S., C.J. Meyer, F. Ma, C.A. Peterson & M.A. Bernards. 2007. The outermost cuticle of soybean seeds: chemical composition and function during imbibition. J. Exp. Bot. 58: 1071-1082. [ Links ]
Tortorelli, L.A. 1956. Maderas y bosques argentinos. Acme, Buenos Aires, Argentina. [ Links ]
Tran, V.N. & A.K. Cavanagh. 1984. Structural aspects of dormancy, p. 1-44. In D.R. Murray (ed.). Seed physiology 2, Academic, Sydney, Australia. [ Links ]
Turner, S.R. & K.W. Dixon. 2009. Seed dormancy and germination in the Australian baobab Adansonia gregorii F.Muell. Seed Sci. Res. 19: 261-266. [ Links ]
Varela, R.O. & E.H. Bucher. 2006. Passage time, viability, and germination of seeds ingested by foxes. J. Arid Environ. 67: 566-578. [ Links ]
Vázquez-Yanes, C. & A. Orozco-Segovia. 1993. Pattern of seed longevity and germination in the tropical rain forest. Annu. Rev. Ecol. Evol. Syst. 24: 69-87. [ Links ]
Vázquez-Yanes, C. & A. Orozco-Segovia. 1994. Signals for seeds to sense and respond to gaps, p. 209-235. In A. Caldwell, M. Marlyn & R. W. Pearcy (eds.). Exploitation of environmental heterogeneity by plants. Academic, London, England. [ Links ]
Werker, E., I. Marbach & A.M. Mayer. 1979. Relation between the anatomy of the testa, water permeability and the presence of phenolics in the genus Pisum. Ann. Bot. 43: 765-771. [ Links ]
Whitmore, T.C. 1989. Canopy gaps and the two major groups of forest trees. Ecology 70: 438-536. [ Links ]
Yaklich, R.W., E.I. Vigil & W. Wergin. 1986. Pore development and seed coat permeability in soybean. Crop Sci. 26: 616-624. [ Links ]
Baskin, J.M., C.C. Baskin & X. Li. 2000. Taxonomy, anatomy and evolution of physical dormancy in seeds. Pl. Spec. Biol. 15: 139-152. [ Links ]
Bewley, J.D. & M. Black. 1982. Physiology and biochemistry of seeds in relation to germination. Viability, dormancy and environmental control. Springer-Verlag, Berlin, Germany. [ Links ]
Boelcke, O. 1946. Estudio morfológico de semillas de leguminosas Mimosoideas y Caesalpinoideas de interés agronómico en la Argentina. Darwiniana 31: 240-321. [ Links ]
Bruno, G., L. Stiefkens, M. Hadid, I. Liscovsky, M.T. Cosa & N. Dottori. 2007. Efecto de la contaminación ambiental en la anatomía de la hoja de Ligustrum lucidum (Oleaceae). Bol. Soc. Argent. Bot. 42: 231-236. [ Links ]
Burkart, A. 1952. Las leguminosas argentinas silvestres y cultivadas. Acme, Buenos Aires, Argentina. [ Links ]
Cabrera, A.L. 1994. Regiones fitogeográficas de Argentina. In W.F. Kugler (ed.). Enciclopedia Argentina de Agricultura y Jardinería. Acme, Buenos Aires, Argentina. [ Links ]
Capeleti, I., M.L. Ferrarese, F.C. Kryzanowski & O. Ferrarese-Filho. 2005. A new procedure for the quantification of lignin in soybean (Glycine max (L.) Merril) seed coat and their relationship with the resistance to mechanical damage. Seed Sci. Technol. 33: 511-515. [ Links ]
D’Ambrogio de Argüeso, A. 1986. Manual de técnicas en histología vegetal. Hemisferio Sur, Buenos Aires, Argentina. [ Links ]
De Souza, F.H.D. & J. Marcos-Filho. 2001. The seed coat as a modulator of seed-environment relationship in Fabaceae. Rev. Bras. Bot. 24: 365-375. [ Links ]
Digilio, A.P.L. & P.R. Legname. 1966. Los árboles indígenas de la Provincia de Tucumán. Ópera Lilloana XV, Tucumán, Argentina. [ Links ]
Erize, F. 1997. Nuevo libro del árbol. Tomo 1. Ateneo, Buenos Aires, Argentina. [ Links ]
Fahn, A. 1985. Anatomía vegetal. Pirámides, Buenos Aires, Argentina. [ Links ]
Fredericksen, T.S., M.J. Justiniano, B. Mostacedo, D. Kennard & L. McDonald. 2000. Comparative regeneration ecology of three leguminous timber species in a Bolivian tropical dry forest. New Forest 20: 45-64. [ Links ]
Janzen, D.H. & C. Vázquez-Yanes. 1990. Aspects of tropical seed ecology of relevance to management of tropical forested wildlands, p. 137-157. In A. GómezPompa, T.C. Whitmore & M. Hadley (eds.). Rain forest regeneration and management, UNESCO and Parthenon, Paris, France. [ Links ]
Justiniano, M.J. & T.S. Fredericksen. 1998. Ecología de especies menos conocidas: Curupaú -Anadenanthera colubrina. Proyecto BOLFOR, Santa Cruz, Bolivia. [ Links ]
Khurana, E. & J.S. Singh. 2001. Ecology of tree seed and seedlings: Implications for tropical forest conservation and restoration. Curr. Sci. 80: 748-757. [ Links ]
Lamprecht, H. 1990. Silvicultura en los trópicos. Los ecosistemas forestales en los bosques tropicales y sus especies arbóreas, posibilidades y métodos para un aprovechamiento sostenido. Deutsche Gesellschaft für Technische Zusammenarbeit (GTZ) GmbH, Eschborn, Germany. [ Links ]
Ma, F., E. Cholewa, T. Mohamed, C.A. Peterson & M. Gijzen. 2004. Craks in the palisade cuticule of soybean seed coats correlate with their permeability to water. Ann. Bot. 94: 213-228. [ Links ]
Mohamed-Yassen, Y., S.A. Barringer, W.E. Splittstoesser & S. Costanza. 1994. The role of the seeds coats in seed viability. Bot. Rev. 60: 246-260. [ Links ]
Moore, R.P. 1972. Tetrazolium staining for assessing seed quality, p. 347-366. In W. Heydecker (ed.). Seed ecology. University of Parth, London, England. [ Links ]
Munsell, A.H. 2000. Munsell soil color charts. Munsell Color Company, I.N.C. Baltimore, Maryland, USA. [ Links ]
Pérez-Herrera, P. & J.A. Acosta-Gallegos. 2002. Permeabilidad de la testa y la porción del micrópilo-hilio en semilla de frijol silvestre y cultivado. Rev. Fitotec. Mex. 25: 57-63. [ Links ]
Qutob, D., F. Ma, C.A. Peterson, M.A. Bernards & M. Gijzen. 2008. Structural and permeability properties of the soybean seed coat. Botany 86: 219-227. [ Links ]
Shao, S., C.J. Meyer, F. Ma, C.A. Peterson & M.A. Bernards. 2007. The outermost cuticle of soybean seeds: chemical composition and function during imbibition. J. Exp. Bot. 58: 1071-1082. [ Links ]
Tortorelli, L.A. 1956. Maderas y bosques argentinos. Acme, Buenos Aires, Argentina. [ Links ]
Tran, V.N. & A.K. Cavanagh. 1984. Structural aspects of dormancy, p. 1-44. In D.R. Murray (ed.). Seed physiology 2, Academic, Sydney, Australia. [ Links ]
Turner, S.R. & K.W. Dixon. 2009. Seed dormancy and germination in the Australian baobab Adansonia gregorii F.Muell. Seed Sci. Res. 19: 261-266. [ Links ]
Varela, R.O. & E.H. Bucher. 2006. Passage time, viability, and germination of seeds ingested by foxes. J. Arid Environ. 67: 566-578. [ Links ]
Vázquez-Yanes, C. & A. Orozco-Segovia. 1993. Pattern of seed longevity and germination in the tropical rain forest. Annu. Rev. Ecol. Evol. Syst. 24: 69-87. [ Links ]
Vázquez-Yanes, C. & A. Orozco-Segovia. 1994. Signals for seeds to sense and respond to gaps, p. 209-235. In A. Caldwell, M. Marlyn & R. W. Pearcy (eds.). Exploitation of environmental heterogeneity by plants. Academic, London, England. [ Links ]
Werker, E., I. Marbach & A.M. Mayer. 1979. Relation between the anatomy of the testa, water permeability and the presence of phenolics in the genus Pisum. Ann. Bot. 43: 765-771. [ Links ]
Whitmore, T.C. 1989. Canopy gaps and the two major groups of forest trees. Ecology 70: 438-536. [ Links ]
Yaklich, R.W., E.I. Vigil & W. Wergin. 1986. Pore development and seed coat permeability in soybean. Crop Sci. 26: 616-624. [ Links ]
*Correspondencia a:
Rodolfo Omar Varela. Instituto de Ecología, Fundación Miguel Lillo, Miguel Lillo 251, San Miguel de Tucumán, CP 4000, Argentina; omarvarela1@gmail.com. Corresponding author. Instituto de Ambientes de Montañas y Regiones Áridas, Universidad Nacional de Chilecito, La Rioja, Argentina.
Patricia Liliana Albornoz. Instituto de Morfología Vegetal, Fundación Miguel Lillo, Miguel Lillo 251, San Miguel de Tucumán, CP 4000, Argentina. Cátedra de Anatomía Vegetal, Facultad de Ciencias Naturales e Instituto Miguel Lillo, Universidad Nacional de Tucumán, Tucumán, Miguel Lillo 251, San Miguel de Tucumán, CP 4000, Argentina; albornoz@csnat.unt.edu.ar
1. Instituto de Ecología, Fundación Miguel Lillo, Miguel Lillo 251, San Miguel de Tucumán, CP 4000, Argentina; omarvarela1@gmail.com. Corresponding author
2. Instituto de Ambientes de Montañas y Regiones Áridas, Universidad Nacional de Chilecito, La Rioja, Argentina.
3. Instituto de Morfología Vegetal, Fundación Miguel Lillo, Miguel Lillo 251, San Miguel de Tucumán, CP 4000, Argentina.
4. Cátedra de Anatomía Vegetal, Facultad de Ciencias Naturales e Instituto Miguel Lillo, Universidad Nacional de Tucumán, Tucumán, Miguel Lillo 251, San Miguel de Tucumán, CP 4000, Argentina; albornoz@csnat.unt.edu.ar
Received 14-VIII-2012. Corrected 08-I-2013. Accepted 08-II-2013


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}