










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.61 n.3 San José Sep. 2013
Anatomía de anillos de crecimiento de 80 especies arbóreas potenciales para estudios dendrocronológicos en la Selva Central, Perú
Anatomic characterization of growth-rings in 80 potential tree species for dendrocronological studies in the Central forest, Perú
Anatomic characterization of growth-rings in 80 potential tree species for dendrocronological studies in the Central forest, Perú
*Dirección para correspondencia:
Abstract
The knowledge about the existence of annual tree rings in tropical trees, which was already found at the beginning of the last century, was ignored by many scientists for a long time. Wood samples of 80 tree species from seven different sites belonging to Satipo and Chanchamayo provinces in Central Forest, Perú. Wood slices were taken at 1.30m height, following the Peruvian Technical Norms (NTP) 251-008, COPANT norms 30:1-019 and IAWA (1989). Results showed that 24 of the 80 tree species analyzed showed a potential for dendrocronological studies, 25 had problems for growth-rings analysis, and 31 did not have potential. The problems most frequently found were: barely visible or irregular ring growth, parenchyma bands and multiseriate rays difficult to be identified in rings growth. The “T” Student test showed that the significant variation in vessel and fiber diameters between growth zones (Early-wood and late-wood) of species with potential for dendrocronology, do have a periodic cells production, so is possible to suggest the annual formation of each growth-ring. However, those species without potential to dendrocronology may be influenced by of a lot of factors, such as biotic and abiotic conditions of environment, as well as the genetic aspect of each species.
Key words: anatomic characterization, growth-rings, early wood, late wood, dendrocronology.
Resumen
El conocimiento acerca de la existencia de anillos anuales en árboles tropicales, se conoce desde principios del siglo pasado, pero fue ignorado por muchos científicos durante largo tiempo. La investigación fue realizada en el Laboratorio de Tecnología de la Madera e Industrias Forestales de la Universidad Nacional del Centro del Perú, con la finalidad de caracterizar anatómicamente los anillos de crecimiento de 80 especies arbóreas potenciales para estudios dendrocronológicos; provenientes de las provincias de Satipo y Chanchamayo en la Selva Central del Perú. Para el estudio se consideraron las normas COPANT 30:1-019, Normas Técnicas Peruanas (NTP) 251-008 y la Lista Estándar de la IAWA 1989. Entre los resultados más sobresalientes tenemos: el 30% de las ochenta especies estudiadas tienen buen potencial para estudios dendrocronológicos. La mayoría de estas especies, están delimitadas por una banda de fibras acortadas radialmente. Entre los problemas encontrados, se puede destacar la presencia de anillos con dificultad para ser visualizados, la presencia de anillos irregulares y la presencia de parénquima en bandas. Las características microscópicas, muestran que la variación significativa en dimensiones de fibras y vasos entre zonas de crecimiento (madera temprana y madera tardía), de las especies potenciales para dendrocronología, tienen una producción de células en forma periódica, lo que podría sugerir la formación anual de cada anillo.
Palabras claves: características anatómicas, anillos de crecimiento, madera temprana, madera tardía, dendroconología.
En el manejo forestal el conocimiento de la edad de los árboles es importante para determinar ciclos de corta y el tiempo durante el cual se renueva una masa forestal (Mendoza 1993). Una forma de calcular la edad en árboles tropicales es por conteo directo de los anillos de crecimiento; sin embargo, es necesario reconocer su existencia o que sean visibles en las especies de interés. La madera que se forma al inicio del anillo es denominada madera temprana y en ella las células son de paredes delgadas y lúmenes amplios. Por su parte, la madera formada al término del periodo vegetativo, tiene células de paredes anchas y lúmenes estrechos. Esta última zona recibe el nombre de madera tardía (Diaz 2003). En bosques templados dichos anillos están bien definidos por células de mayor tamaño en la madera temprana y de menor tamaño en la madera tardía. En contraste, en maderas tropicales la delimitación del anillo puede estar dada por estructuras celulares como parénquima marginal y fibras de paredes más gruesas o aplanadas en dirección radial. Por tanto, para confirmar la presencia de anillos se hacen estudios anatómicos microscópicos de los componentes celulares en la madera (elementos de vaso y fibras) (Carlquist 1988). El conocimiento acerca de la existencia de los anillos anuales en los árboles tropicales, fue encontrado a principios del siglo pasado e ignorado por muchos científicos a lo largo del tiempo (Worbes 1989). El análisis de los anillos de árboles tropicales es más difícil que el de las zonas templadas y boreales, debido a la compleja estructura anatómica de los árboles tropicales, condiciones de sitio, especies que forman más de un anillo por año (periodicidad), escasa información sobre la ecología del crecimiento para muchas especies, entre otros aspectos. Estos son factores que han dificultado el avance de estudios dendrocronológicos en los trópicos. Sin embargo, diferentes estudios demostraron la existencia de anillos anuales de crecimiento en muchas especies de árboles tropicales (Tomazello et al. 2004) y subtropicales (Navar et al. 2001). Esto debido a que muchos de los climas tropicales tienen estaciones predecibles con un exceso o escasez de disponibilidad de agua, cambios estacionales en la temperatura o una combinación de ambos, lo que origina la formación de los anillos de crecimiento (López et al. 2002).
En el Perú los estudios dendrocronológicos están fuertemente limitados por la dificultad de hallar especies potenciales para estudios dendrocronológicos, con anillos de crecimiento claramente distinguibles, por ello en pocas ocasiones se han realizado estudios dirigidos a establecer un método de recuento de anillos en estas especies.
La anatomía de la madera es importante ya que nos permite identificar las características necesarias que debe tener una especie para la dendrocronología así como también determinar la correcta identificación de los anillos de crecimiento. Esta investigación, nos ayudará a contribuir con información base para futuras investigaciones en el campo de la dendrocronología con especies tropicales que poco se ha estudiado en el Perú. El presente estudio tiene como objetivo caracterizar anatómicamente los anillos de crecimiento de 80 especies arbóreas potenciales para estudios dendrocronológicos en la Selva Central del Perú.
Materiales y Métodos
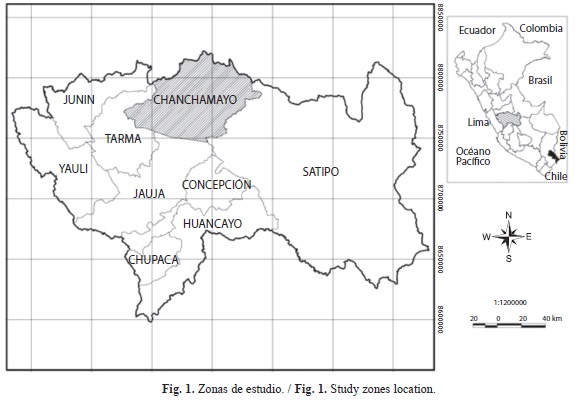
Zonas de estudio: Este estudio se llevó a cabo en las provincias de Chanchamayo y Satipo, Perú (Fig. 1). La precipitación total anual en la provincia de Satipo es de 2 324.8mm, presentándose la mínima con 67.7mm en el mes de julio y la máxima con 383.3mm durante el mes de enero (Fig. 2). La temperatura media durante el año es homogénea variando entre 24ºC-27ºC (Instituto de investigaciones de la Amazonia Peruana, 2011). La provincia de Chanchamayo tiene una precipitación total anual que varía entre 1 500-3 000mm, una temperatura que oscila entre 15-19ºC (Young & León 1999).
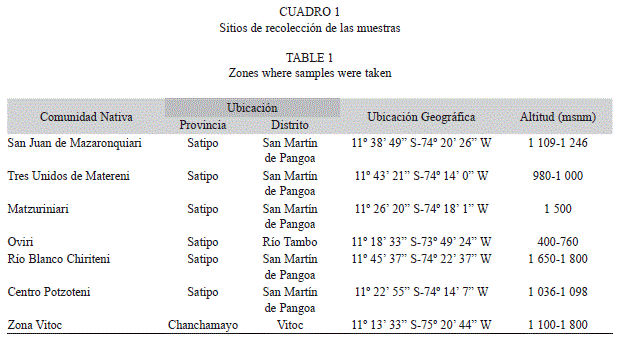
Rodajas de madera de 80 especies forestales se recolectaron de 28 familias en las zonas descritas en el cuadro 1.
Se seleccionaron de dos a tres árboles por especie de los cuales se obtuvieron rodajas de madera con una motosierra marca STIHL modelo B 175.1-2 000; a 1.30m sobre el suelo.
Se consideraron árboles de la categoría diamétrica que fluctúa entre 40-120cm y de buena sanidad física y biológica. La recolección de muestras empleó la norma NTP (Normas Técnicas Peruanas) Nº251-008. Las características macroscópicas se observaron con una lente de mano de 10X, según la norma COPANT Nº30:1-019.
Con el fin de observar la presencia de los anillos de crecimiento visibles, primero se obtuvieron tiras de madera de 2cm de lado, que fueron pulidas con lijas de diferente granulometría 80, 60, 100,120, 220, 240 y 360. Se emplearon sustancias contrastantes como: safranina al 1%, limón, kerosene, ron de quemar y thinner para mejorar la visibilidad de los anillos de crecimiento.
Para confirmar la presencia de anillos de crecimiento se hacen estudios anatómicos microscópicos de los componentes celulares en la madera (elementos de vaso y fibras). Las variables consideradas fueron las siguientes: anillos de crecimiento, diámetro de vasos y diámetro de fibras. Se empleó el analizador de imágenes Leica Aplication Suite EZ (Versión 1.6.0, 2008), adaptado a una cámara de video Modelo DC 150 7.1, y a un microscopio trinocular Leica, para efectuar las observaciones y mediciones. En el laboratorio las muestras se seccionaron en probetas de 2x2x2cm de lado, se hirvieron en agua por un lapso de 12-40h dependiendo de la dureza de la especie. Posteriormente, se procedió a ablandar las muestras con alcohol al 30% y glicerina al 70%. Con un micrótomo de deslizamiento horizontal (modelo SM 2000R), se obtuvieron cortes transversales de un espesor entre 20-30µm, dependiendo de la especie. Las láminas se deshidrataron en diferentes concentraciones de alcohol (30%, 70%, 80% y 95%) por un lapso de 20min, se aclararon con el reactivo verde brillante y se colorearon con safranina al 1%, luego se colocaron en un portaobjeto con una a dos gotas de bálsamo de Canadá, y finalmente se cubrieron con una lámina cubreobjetos.
De las especies que presentaron potencial para estudios dendrocronológicos debido a que tienen características importantes como: anillos distintos (visibles), anillos regulares; no presentando dificultades en el tipo de parénquima axial (bandas anchas de parénquima), tipo de transición gradual de zonas fibrosas y/o traqueidas, radios multiseriados; que dificulten la identificación de los anillos de crecimiento; se astilló la madera temprana y madera tardía semejante a los palitos de fósforo, de aproximadamente 1.5mm de espesor con la ayuda de una cuchilla, que se obtuvieron a partir de los cubos de 2x2x2cm. Para la maceración de las astillas estas se colocaron en un frasco con tapa hermética con una solución de ácido acético y peróxido de hidrógeno en proporciones de uno a dos, respectivamente, de manera que cubrió completamente las astillas; se secaron a 65°C por 48-72h. Además, se realizaron 50 mediciones del diámetro de vasos y de fibras de cada repetición (25 en madera temprana y 25 en madera tardía). Para determinar si existe diferencia estadística significativa entre la madera temprana y tardía, se empleó la estadística inferencial, prueba de “T” de Student al 95% de intervalo de confianza, para muestras independientes. Estos análisis estadísticos se realizaron con el software XLSTAT.
Resultados
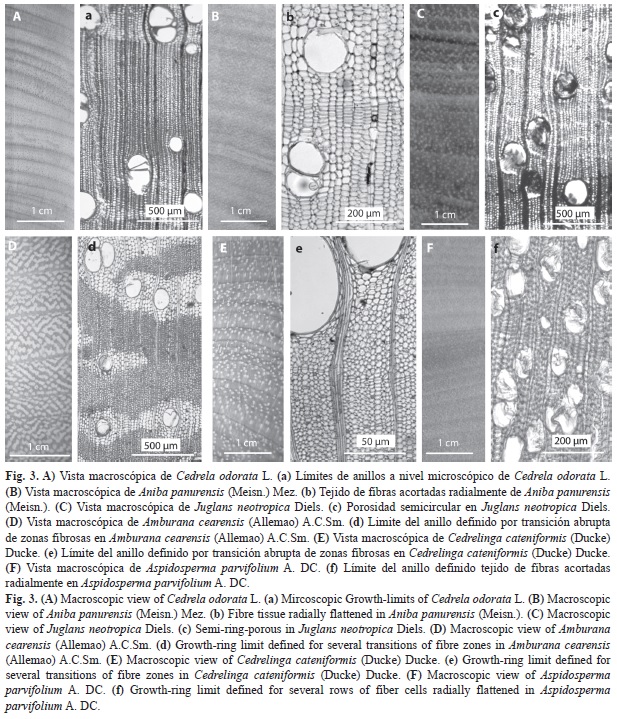
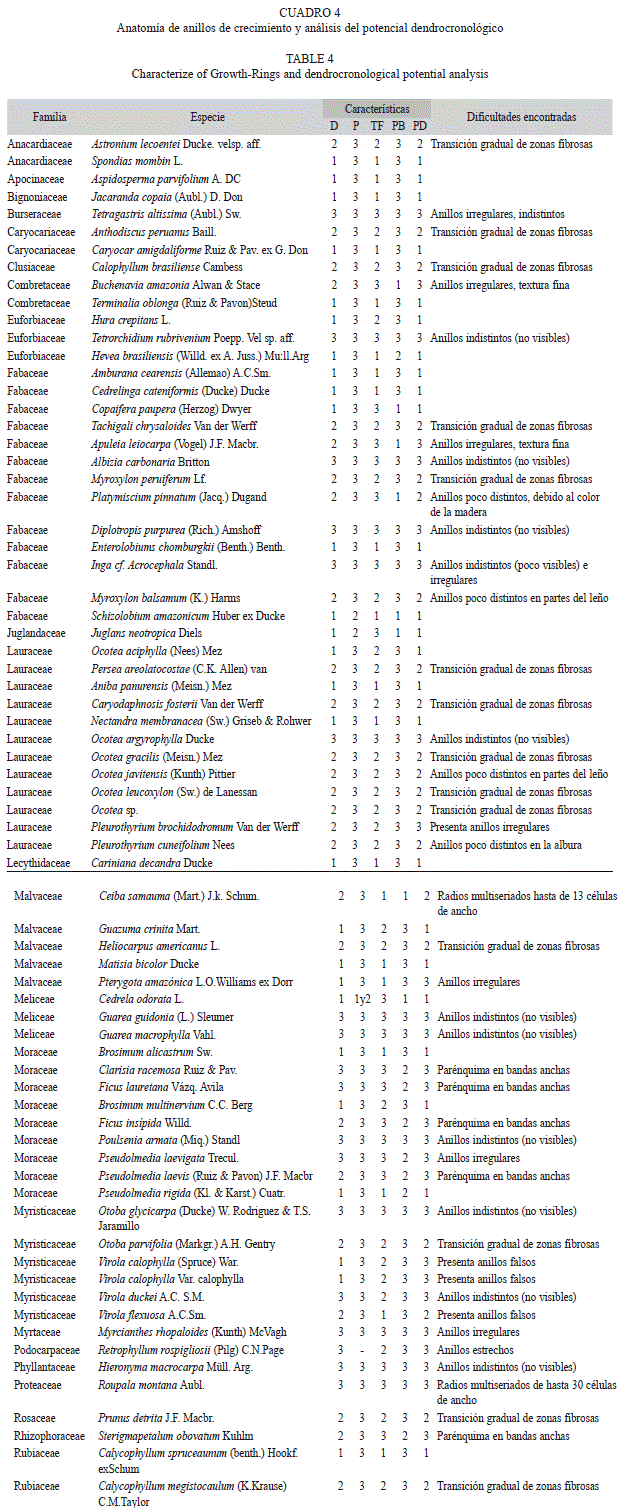
La caracterización anatómica de los anillos de crecimiento de las especies analizadas se describe a continuación en orden por familia (Fig. 3, Cuadro 4).
Anacardiaceae: Fueron investigadas dos especies de esta familia: Spondias mombin L., presenta anillos de crecimiento distintos a simple vista, regulares, delimitados por una banda de fibras acortadas radialmente, con potencial dendrocronológico; Astronium lecoentei Ducke. velsp. aff tiene anillos de crecimiento poco distintos, regulares, delimitados por una banda de fibras acortadas radialmente, con dificultad para estudios dendrocronológicos.
Apocynaceae: La especie Aspidosperma parvifolium A. DC. presentó anillos de crecimiento con potencial dendrocronológico, limitados por una banda de fibras acortadas radialmente.
Bignoniaceae: La especie Jacaranda copaia (Aubl.) D. Don. presentó anillos de crecimiento distintos a simple vista, regulares, delimitados por una banda de fibras acortadas radialmente, con potencial dendrocronológico.
Caryocaraceae: Fueron investigadas dos especies de esta familia: Caryocar amigdaliforme Ruiz & Pav. ex G. Don. presentó anillos de crecimiento distintos a simple vista, regulares, delimitados por una banda de fibras acortadas radialmente, con potencial dendrocronológico; Anthodiscus peruanus Baill, tiene anillos de crecimiento poco distintos, regulares, delimitados por una banda de fibras acortadas radialmente, con dificultad para estudios dendrocronológicos.
Clusiaceae: La especie Calophyllum brasiliense Cambess. presentó anillos de crecimiento poco distintos (visibles), delimitados por una banda de fibras acortadas radialmente, con dificultad para dendrocronología.
Combretaceae: Terminalia oblonga (Ruiz & Pavon) Steud. presentó anillos de crecimiento distintos a simple vista, regulares, delimitados por una banda de fibras acortadas radialmente, con potencial dendrocronológico. Buchenavia amazonia Alwan & Stace., presentó anillos de crecimiento poco distintos, irregulares, delimitados por una línea delgada de parénquima marginal, con dificultad para estudios dendrocronológicos.
Euphorbiaceae: De las tres especies estudiadas de esta familia dos especies tienen potencial dendrocronológico: Hevea brasiliensis (Willd. ex A. Juss.) Müll. Arg. y Hura crepitans L. presentaron anillos de crecimiento distintos a simple vista, regulares delimitados por una banda de tejido de fibras acortadas radialmente.
Fabaceae: Se estudiaron 13 especies de esta familia, de las cuales reportamos cinco especies con potencial dendrocronológico. Amburana cearensis (Allemao) A.C.Sm., Cedrelinga cateniformis (Ducke) Ducke y Enterolobium schomburgkii (Benth.) Benth, las cuales presentaron anillos de crecimiento distintos, regulares, delimitados por una banda de fibras acortadas radialmente. Copaifera paupera (Herzog) Dwyer., presenta anillos de crecimiento distintos a simple vista, regulares, delimitados por una banda de parénquima marginal y Schizolobium amazonicum Huber ex Ducke. presenta anillos de crecimientos delimitados por una banda de fibras acortadas radialmente asociado a una fina banda de parénquima marginal. Por otro lado, tres especies tienen dificultad para estudios dendrocronológicos: Tachigali chrysaloides Van der Werff y Myroxylon peruiferum Lf. tienen anillos de crecimiento poco distintos, regulares, delimitados por una banda de fibras acortadas radialmente, Platymiscium pinnatum (Jacq.) Dugand., tiene anillos de crecimiento poco distintos (debido al color del duramen) regulares, delimitados por una banda de parénquima marginal.
Juglandaceae: La única especie estudiada de esta familia es Juglans neotropica Diels, la cual tiene potencial para dendrocronología, presentó anillos de crecimiento delimitados por la diferencia en el diámetro de los poros (porosidad semicircular), asociado a una banda delgada de parénquima marginal.
Lauraceae: De las 12 especies investigadas de esta familia, tres especies mostraron potencial para dendrocronología: Ocotea aciphylla (Nees) Mez, Aniba panurensis (Meisn.) Mezy y Nectandra membranacea (Sw.) Griseb & Rohwer, las cuales presentaron anillos de crecimiento delimitados por una banda de tejido de fibras acortadas radialmente. Así mismo dos especies de esta familia presentan dificultad para dendrocronología: Persea areolatocostae (C.K. Allen) van y Ocotea gracilis (Meisn.) Mez. tienen anillos de crecimiento poco distintos, regulares, delimitados por una banda delgada de fibras acortadas radialmente.
Lecythidaceae: Cariniana decandra Ducke. presentó anillos de crecimiento distintos a simple vista, regulares, delimitados por una banda de fibras acortadas radialmente, con potencial para dendrocronología.
Malvaceae: Se estudiaron cinco especies de esta familia de las cuales Matisia bicolor Ducke y Guazuma crinita Mart. mostraron potencial para dendrocronología, presentando anillos de crecimiento distintos, regulares, delimitados por una banda de fibras acortadas radialmente. Por otro lado Ceiba samauma (Mart.) J.k. Schum. mostró anillos de crecimiento poco distintos, delimitados por una banda delgada de parénquima marginal, con dificultad para estudios dendrocronológicos.
Meliceae: Fueron investigadas tres especies de esta familia de las cuales solo Cedrela odorata L. mostró potencial dendrocronológico, esta especie presentó anillos de crecimiento distintos a simple vista, regulares, caracterizados por tener porosidad de circular a semicircular y asociados a una banda de parénquima marginal.
Moraceae: Se estudiaron nueve especies de esta familia, se reportan tres especies potenciales para dendrocronología: Brosimum alicastrum Sw. y Brosimum multinervium C.C. Berg que presentaron anillos distintos delimitados por una banda de fibras acortadas radialmente y Pseudolmedia rigida (Kl. & Karst.) con anillos delimitados por una banda de fibras acortadas radialmente, así mismo, se observó una decreciente frecuencia de bandas de parénquima desde el inicio hacia el final del anillo de crecimiento.
Myristicaceae: Las seis especies estudiadas de esta familia presentaron anillos de crecimiento con dificultad para dendrocronología, Otoba parvifolia (Markgr.) A. H. Gentry, Otoba glycicarpa (Ducke) W. Rodríguez & T.S. Jaramillo, Virola duckei A.C. S.M., Virola flexuosa A.C. Sm., al mostrar anillos de crecimiento poco distintos, regulares, delimitados por una banda de fibras acortadas radialmente.
Rosaceae: Prunus detrita J.F. Macbr. mostró anillos de crecimiento poco distintos, regulares, delimitados por una banda de fibras acortadas radialmente.
Rubiaceae: Se estudiaron tres especies de esta familia, de las cuales solo la especie Calycophyllum spruceaunum (Benth.) Hookf. Ex Schum tiene potencial para dendrocronología, con anillos delimitados por una banda de fibras acortadas radialmente. Así mismo Calycophyllum megistocaulum (K. Krause) C.M. Taylor, tienen dificultad para estudios dendrocronológicos, con anillos de crecimiento poco distintos, regulares, delimitados por una banda de fibras acortadas radialmente.
Staphyleaceae: Huertea glandulosa (Ruiz & Pavon) Steud. presentó anillos de crecimiento poco distintos, debido al color de la madera (duramen), regulares, delimitados por una banda de fibra acortadas radialmente, con dificultad para dendrocronología.
Theaceae: Gordonia fruticosa (Schrad.) H. Keng. presentó anillos de crecimiento poco distintos, regulares, delimitados por una banda de fibra de mayor espesor de pared acortados radialmente, con potencial dendrocronológico con dificultad.
Vochysiaceae: Vochysia vismifolia Spruce & Warm. mostró anillos de crecimiento poco distintos, regulares, delimitados por una banda de fibra acortadas radialmente, con dificultad para dendrocronología.
La prueba de “T” de Student realizada a las 23 especies que presentaron buen potencial para dendrocronología, concluyó que 10 especies mostraron diferencia estadística significativa en el diámetro de vasos de la madera temprana frente a la madera tardía: Pseudolmedia rigida y Copaifera paupera, presentó diferencia estadística significativa (p<0.05); así mismo Enterolobium schomburgkii Schizolobium amazonicum, Juglans neotropica, Cedrela odorata, Brosimum alicastrum, Amburana cearensis, Spondias mombin y Terminalia oblonga, presentaron diferencias estadísticas altamente significativas (p<0.01) (Cuadro 2). Con respecto al diámetro de fibras, 11 especies mostraron diferencias estadísticas significativas entre madera temprana y tardía: Juglans neotropica y Brosimum alicastrum presentaron diferencia estadística significativa (p<0.05), así mismo Schizolobium amazonicum, Aniba panurensis, Nectandra membranacea, Matisia bicolor, Hura crepitans, Amburana cearensis, Jacaranda copaia, Spondias mombin y Cedrelinga cateniformis, reportaron diferencias altamente significativas (p<0.01) (Cuadro 3).
Discusión
En relación a la porosidad, 76 especies presentaron poros difusos, Cedrela odorata L. posee porosidad de circular a semicircular, Juglans neotropica Diels y Schizolobium amazonicum Huber ex Ducke, tienen porosidad semicircular y Retrophyllum rospigliosii (Pilg) C.N. Page, no poseen poros por ser una conífera. Así también, Campos (2009), reportó veinticinco especies con porosidad difusa y Cedrela odorata con porosidad semicircular, con estudios similares realizados en Brasil y Perú. Esta característica de la porosidad difusa es corroborada por el (IAWA 1989 y Chavesta 2006), quienes reportan este tipo de porosidad en la extensa mayoría de las especies tropicales. Así mismo las condiciones ambientales que presenta la zona de estudio sería fundamental para la presencia de la porosidad difusa, donde el crecimiento está determinado por la precipitación, siendo esta de manera continua casi en todo el año, con una sola época de lluvias escasas (mayo-setiembre).
De las 23 especies que mostraron anillos de crecimiento distintos (visibles), 20 especies, están delimitadas por una banda de tejido fibroso acortado radialmente, concordando con Díaz (2003), quien menciona que el acortamiento de fibras y traqueidas está presente en la mayoría de las especies de latifoliadas. Esto se debería a que al final de la estación de crecimiento, los factores tales como sequía, reducen la concentración de auxinas y por lo tanto fomentan producción de células de diámetro pequeño; mientras tanto, la acumulación de inhibidores de crecimiento causa reducción en el ritmo de producción de nuevas células, cesando a su vez el crecimiento de nuevos brotes y desarrollo de nuevas hojas. Por lo tanto, el producto fotosintético producido por las hojas desarrolladas a plenitud, está disponible para la síntesis de la pared celular. Posteriormente, el resultado será células de paredes gruesas y diámetros pequeños (Vera et al. 2011). Esto se refleja en las dimensiones de las células de madera tardía de paredes anchas y lúmenes estrechos y madera temprana de paredes delgadas y lúmenes amplios, estas dos partes del anillo, producen diferencias de tonalidad.
Las 12 especies de la familia Lauraceae, presentaron anillos de crecimiento delimitados por una banda de tejido fibroso acortados radialmente, lo que coincide con lo reportado por Tomazello et al. (2004) y con estudios similares realizados en Brasil, quienes mencionan que es común encontrar en especies de la familia Lauraceae este tipo de delimitación. El crecimiento radial en los árboles, originado por el cambium, genera tejido xilemático secundario y floema durante el periodo de crecimiento. Este crecimiento depende en gran medida de las características genéticas de cada especie y de la influencia de factores internos (edad) y externos (clima, suelo, competencia y sitio).
De las 13 especies de la familia Fabaceae, cuatro especies presentan anillos de crecimiento delimitados por una banda de parénquima marginal, contrario a lo hallado por (Tomazello et al. 2004) quienes mencionan que las bandas de parénquima marginal son comunes en esta familia. Estas características podría deberse a muchos factores, entre ellos las condiciones bióticas y abióticas del ambiente que rodea al individuo, así como también al aspecto genético de la especie.
Retrophyllum rospigliosii (Pilg) C.N. Page, es la única conífera de las 80 especies estudiadas que muestra los anillos de crecimiento indistintos macroscópicamente, característica respaldada por León & Espinoza (2001), quienes mencionan que las células de las coníferas forman madera temprana con células de paredes delgadas y lúmenes amplios, que otorgan a esta parte del crecimiento un tono más claro que el que tiene la zona con madera tardía, y la transición entre una zona y otra puede ser gradual o abrupta. En esta especie la transición es gradual por ello no se diferencian los anillos de crecimiento macroscópicamente.
Con respecto a la variación de diámetros de los vasos de las especies estudiadas se observó que 12 especies presentan diferencias estadísticas significativas para el diámetro de los vasos, de los cuales 11 mostraron diferencias altamente significativas. Así mismo López (2006), menciona que la combinación de diámetros grandes y longitudes cortas de los vasos en madera temprana tiene una estrecha relación con la eficiencia en transporte del agua, mientras que los diámetros pequeños y longitudes grandes en madera tardía tienen como función principal evitar la ocurrencia de embolismos durante el periodo de sequía. Así también, León & Espinoza (2001), documentan que a medida que aumenta el tamaño de los poros se produce un incremento en la eficiencia de la conducción del agua. Estos resultados podrían sugerir que el crecimiento de estas especies responde a la variación de la precipitación existente en la zona de estudio.
En lo que referente a la diferencia del diámetro de fibras de madera temprana y tardía se aprecia que 10 especies presentaron diferencias estadísticas, de las cuales ocho reportaron diferencias estadísticas altamente significativas. Al respecto, Díaz (2003) menciona que las diferencias de dimensiones de las células en estas dos partes del anillo (madera temprana y madera tardía) producen diferencias de tonalidad.
Por ello, en muchas especies, la madera temprana es más clara que la madera tardía.
Agradecimientos
Al proyecto “Determinación de Incrementos Anuales y Turnos Biológicos de Rotación de Especies Forestales Tropicales para Mejorar el Aprovechamiento de Madera Comercial” del Programa de Ciencia y Tecnología, según Resolución Nº 220-PCM/FINCyT”, formulado por Jimmy Edilson Requena Rojas y Mariano Morales, por brindarnos los medios para la ejecución de esta investigación. Al coordinador del proyecto Julio Álvarez Orellana. A Isidora Gonzales Casimiro, por su apoyo y por haber sido nuestra guía durante el desarrollo de esta investigación. A Víctor Rondón Lazo, Gerente General de la Empresa Industrial Maderera “Los Ángeles” S.A.C, Mazamari; a Uldarico Gaspar, Gerente General de la Empresa Orient Trade S.A.C., Mazamari; a Fernando Travi F. Gerente General de la Empresa NEMATSA SRL., Satipo; a Alfredo Álvarez Gerente General de la Empresa San Ramón S.A.C, Satipo; a Javier Hurtado Gerente General de la Empresa Industrial Maderera Hurtado S.A.C, Satipo; a Gustavo Alcázar Serna, Gerente General de la Empresa Maderera G y G Alcázar, Satipo; A Jaime Velásquez, Gerente General de la Empresa Maderas de Exportación S.A. MADEXSA, Satipo; por su apoyo en la recolección de las muestras en campo.
Referencias
Campos, L. 2009. Dendrocronología en árboles de Tornillo, Cedrelinga cateniformis Ducke (Fabaceae), del Centro de Investigaciones Jenaro Herrera en el noreste de la Amazonia, Región Loreto-Perú. Tesis Ingeniería Forestal, Universidad Nacional Agraria La Molina, Lima, Perú [ Links ].
Carlquist, S. 1988. Comparative Wood anatomy, systematic, ecological and evolutionary aspects of dicotyledonous wood. Springer-Verlag, Berlín, Alemania. [ Links ]
Chavesta, C. 2006. Separata de Capacitación Sobre Identificación de maderas. Universidad Nacional Agraria La Molina, USAID, Lima, Perú [ Links ].
Díaz, O. 2003. Anatomía de Maderas. Universidad Austral de Chile, Valdivia, Chile. [ Links ]
IAWA. 1989. List of microscopic features for hard wood identification. International association of wood and atomists. IAWA Bulletin n.s., E.A.Wheeler, P. Baas & P.E. Gasson. Editorial Board, Leyden, Holland. [ Links ]
Instituto de investigaciones de la Amazonia Peruana (IIAP). 2011. Zonificación ecológica y económica de la provincial de Satipo. Lima, Perú [ Links ].
León, W. & N. Espinoza. 2001. Anatomía de la Madera. Consejo de Publicaciones, Mérida, Venezuela. [ Links ]
López, A., J. Valdez & R. Terrazas. 2006. Anillos de Crecimiento y su Periodicidad en Tres Especies Tropicales del Estado de Colima, México. Red de Revistas Científicas de América Latina y el Caribe, España y Portugal 4: 533-544. [ Links ]
López, L., R. Villalba & C. Peña. 2002. Periodicidad en el crecimiento de siete árboles forestales, usando como indicador cicatrices de fuego, marcados durante el incendio forestal de 1995, ocurrido en la Chonta, Guarayos Santa Cruz, Bolivia. IANIGLA/CONICET. 330: 1-27. [ Links ]
Mendoza, B. 1993. Conceptos Básicos de Manejo Forestal. Editorial UTEHA, D.F., México. [ Links ]
Návar, J., J. Nájera & E. Jurado. 2001. Preliminary Estimates of Biomass Growth in the Tamaulipan Thornscrub of Northeastern Mexico. J. Arid Environ. 47: 281-290. [ Links ]
Tomazello, M., C. Lisi, N. Hansen & G. Cury. 2004. Anatomical features of increment zones in different tree species in the State of Sao Paulo, Brazil. Sci. Forestalis 66: 46-55. [ Links ]
Vera, A., D. Ortega, P. Pérez, J. Rosero & M. Chavesta. 2011. Avances en la caracterización e identificación anatómica de los anillos de crecimiento de la especie Junglans neotropica Diels (Nogal) para estudios dendrocronológicos en el fundo La Génova, Junín. Rev. Xilema 24: 5-11. [ Links ]
Young, K. & B. León. 1999. Peru`s sumid eastern montane forest: an overview of their physical setting, biological diversity, human use and settlement, and conservation needs. Centre for Research on the Cultural and Biological Diversity of Andean Rainforests (DIVA) Technical Report 5: 1-97. [ Links ]
Worbes, M. 1989. Growth rings, increment and age of trees in inundation forest, savannas and a mountain forest in the neotropics. IAWA Bulletin new series 10: 109-122. [ Links ]
Carlquist, S. 1988. Comparative Wood anatomy, systematic, ecological and evolutionary aspects of dicotyledonous wood. Springer-Verlag, Berlín, Alemania. [ Links ]
Chavesta, C. 2006. Separata de Capacitación Sobre Identificación de maderas. Universidad Nacional Agraria La Molina, USAID, Lima, Perú [ Links ].
Díaz, O. 2003. Anatomía de Maderas. Universidad Austral de Chile, Valdivia, Chile. [ Links ]
IAWA. 1989. List of microscopic features for hard wood identification. International association of wood and atomists. IAWA Bulletin n.s., E.A.Wheeler, P. Baas & P.E. Gasson. Editorial Board, Leyden, Holland. [ Links ]
Instituto de investigaciones de la Amazonia Peruana (IIAP). 2011. Zonificación ecológica y económica de la provincial de Satipo. Lima, Perú [ Links ].
León, W. & N. Espinoza. 2001. Anatomía de la Madera. Consejo de Publicaciones, Mérida, Venezuela. [ Links ]
López, A., J. Valdez & R. Terrazas. 2006. Anillos de Crecimiento y su Periodicidad en Tres Especies Tropicales del Estado de Colima, México. Red de Revistas Científicas de América Latina y el Caribe, España y Portugal 4: 533-544. [ Links ]
López, L., R. Villalba & C. Peña. 2002. Periodicidad en el crecimiento de siete árboles forestales, usando como indicador cicatrices de fuego, marcados durante el incendio forestal de 1995, ocurrido en la Chonta, Guarayos Santa Cruz, Bolivia. IANIGLA/CONICET. 330: 1-27. [ Links ]
Mendoza, B. 1993. Conceptos Básicos de Manejo Forestal. Editorial UTEHA, D.F., México. [ Links ]
Návar, J., J. Nájera & E. Jurado. 2001. Preliminary Estimates of Biomass Growth in the Tamaulipan Thornscrub of Northeastern Mexico. J. Arid Environ. 47: 281-290. [ Links ]
Tomazello, M., C. Lisi, N. Hansen & G. Cury. 2004. Anatomical features of increment zones in different tree species in the State of Sao Paulo, Brazil. Sci. Forestalis 66: 46-55. [ Links ]
Vera, A., D. Ortega, P. Pérez, J. Rosero & M. Chavesta. 2011. Avances en la caracterización e identificación anatómica de los anillos de crecimiento de la especie Junglans neotropica Diels (Nogal) para estudios dendrocronológicos en el fundo La Génova, Junín. Rev. Xilema 24: 5-11. [ Links ]
Young, K. & B. León. 1999. Peru`s sumid eastern montane forest: an overview of their physical setting, biological diversity, human use and settlement, and conservation needs. Centre for Research on the Cultural and Biological Diversity of Andean Rainforests (DIVA) Technical Report 5: 1-97. [ Links ]
Worbes, M. 1989. Growth rings, increment and age of trees in inundation forest, savannas and a mountain forest in the neotropics. IAWA Bulletin new series 10: 109-122. [ Links ]
*Correspondencia a:
Lizandro Adal Beltrán Gutiérrez. Laboratorio de tecnología de la madera, Facultad de Ciencias Forestales y del Ambiente, Universidad Nacional del Centro del Perú, Huancayo, Perú; lizandro40bg@hotmail.com
Gina Mariela Valencia Ramos. Laboratorio de tecnología de la madera, Facultad de Ciencias Forestales y del Ambiente, Universidad Nacional del Centro del Perú, Huancayo, Perú; iraya_21@hotmail.com
1. Laboratorio de tecnología de la madera, Facultad de Ciencias Forestales y del Ambiente, Universidad Nacional del Centro del Perú, Huancayo, Perú; lizandro40bg@hotmail.com, iraya_21@hotmail.com
Recibido 09-I-2012. Corregido 20-X-2012. Aceptado 23-XI-2012.


{kind=link}
{kind=link}
{kind=link}
{kind=link}