










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.61 n.2 San José Jun. 2013
Comparative anatomy of leaflets of Zamia acuminata and Z. pseudomonticola (Zamiaceae) in Costa Rica
*Dirección para correspondencia:
Abstract
The genus Zamia is morphologically and ecologically the most diverse of the order Cycadales. Throughout its history this genus has been restricted to the New World and is presently almost entirely restricted to the Neotropics. Unusual anatomical traits of the leaflets, such as the sunken stomata and thick cuticle, are common in this and related genera. The objective of this research was to study and compare the leaflet anatomy of Zamia acuminata and Z. pseudomonticola and establish possible phylogenetic relationships between the anatomical traits and the near relatives of these species. The leaf material was obtained from living plants and then processed for electron microscopy study. We found that both species are very similar to each other and to Z. fairchildiana, and that they share several unusual traits with other species of the genus, such as the parenchyma morphology, the spatial distribution of tissues between the veins and the stomata morphology. The main differences between these species were seen in their fiber clusters and in the abundance of trichome basal cells on the epidermis. The anatomical similarities between the three species could be the result of their close phylogenetic relationship and the divergences between them could be the result of recent speciation during the Pleistocene, resulting from geological changes in Southern Costa Rica.
Key words: Zamia, Costa Rica, leaflet anatomy.
Resumen
Zamia es morfológica y ecológicamente el género más diverso del orden Cycadales. Este género siempre ha estado restringido a América, pero en la actualidad habita principalmente en la región neotropical. Características anatómicas inusuales en los foliolos como los estomas hundidos y las cutículas gruesas son comunes en Zamia y géneros afines. El objetivo de este trabajo consiste en comparar la anatomía de los foliolos de Zamia acuminata y Z. pseudomonticola y establecer posibles relaciones filogenéticas entre las características anatómicas y los parientes cercanos de esta especie. Las hojas de las especies seleccionadas fueron obtenidas de plantas vivas y luego procesadas para el estudio por microscopía electrónica. Ambas especies son muy similares entre sí y respecto a Z. fairchildiana y comparten varias características en común con otras especies del género como son la morfología del parénquima, de los estomas y la distribución espacial de tejidos alrededor de las haces vasculares. Las diferencias más notables entre especies se vieron a nivel de sus paquetes de fibras y en la abundancia de células basales de los tricomas en la epidermis. Las similitudes anatómicas entre estas tres especies pueden ser el resultado de su cercanía filogenética y las diferencias podrían ser el resultado de especiación durante el Pleistoceno, producto de los eventos geológicos y cambios sucedidos en el sur de Costa Rica en esa época.
Palabras clave: Zamia, Costa Rica, anatomía foliolos.
Cycadales Pers. ex Bercht. & J. Presl is the least derived group amongst the extant seed plant orders (Taylor et al. 2009), with 331 recognized species (Osborne et al. 2012). This order is morphologically conservative, with little anatomical variation within families, especially when compared to groups like the angiosperms (Norstog & Nicholls 1998). Nowadays, the order is restricted to tropical and subtropical areas of both hemispheres, extending into warm temperate areas only in Southern Africa and Australia. Hill et al. (2003) proposed a classification model that divides the order in two families supported by the divergence between Cycas L. and a clade that includes all the other genera of the order.
Zamia L. is the second most speciose genus of the order and it shows greater morphological, karyological and ecological variation than other cycad genera (Norstog 1981, Vovides 1983, Caputo et al. 1996, Stevenson 2001, Jones 2002). Also, Zamia is the only genus of Cycadales which exhibits karyotypic variation both intra-and inter-specifically (Norstog 1981, Vovides 1983, Caputo et al. 1996). According to some researchers in other plant groups such as conifers and angiosperms, this could indicate rapid evolutionary rates (Eckenwalder 2008, Takhtajan 2009).
Cycads are the only gymnosperms which have developed compound leaves. Most extant representatives show adaptations for xerophytic environments: a thick cuticle, a well defined hypodermis made up of thick-walled cells and sunken stomata, usually only on the underside of the leaflets. These traits were possibly inherited from an ancestor which inhabited dry and strongly irradiated environments (Norstog & Nicholls 1998).
All known species of Zamia have pinnate leaves (Jones 2002), but the number of leaflets per leaf is variable. The main axis of the leaf is constituted by the petiole, often with prickles, and the rachis, which holds the leaflets. The leaflets are articulated to the rachis and, unlike most other cycads, the leaflets are shed independently from the rachis. The petiole bases and cataphylls are shed completely and do not persist on the stem after senescing. The internal anatomy of the petiole-rachis axis is characterized by the dominance of parenchyma along with some mucilage ducts and relatively few and small discreet vascular strands which, in cross section, show an omega (Ω) shape arrangement. This trait is diagnostic of the order (Norstog & Nicholls 1998).
In Zamia the leaflets lack a well differentiated midvein (except in Z. restrepoi (D.W. Stev) A. Lindstr.), while the leaflet is crossed by several vascular strands (Norstog & Nicholls 1998, Jones 2002). Greguss (1968) studied in great detail the epidermal features of most cycadalean species known at the time. In more recent works, Newell (1985, 1989) studied variation in the external morphology of the leaflets of some Caribbean species of Zamia. Stevenson (1981, 1990), Stevenson et al. (1996) and Norstog & Nicholls (1998) provide general descriptions of several aspects of the leaf anatomy of diverse cycad genera. In other genera, such as Cycas, the differences in the arrangement, abundance and distribution of the tissues inside the leaflets are used to clarify the taxonomy of some confusing species groups (Hill 1996). Previous studies in Z. fairchildiana L.D. Gómez and Z. neurophyllidia D.W. Stev. have demonstrated that even though the foliar anatomy follows a general pattern, there are details in which the species differ, such as the shape and distribution of fiber bundles associated with the vascular tissue, and the size and shape of the air chambers of the mesophyll (Acuña-Castillo & Marín-Méndez 2012).
The goals of this study were to analyze and compare the leaflet anatomy of Z. acuminate Oerst. ex Dyer and Z. pseudomonticola L.D. Gómez and establish possible relationships between the leaflet anatomy and the phylogeny of some Zamia species.
Materials and methods
We worked with leaf samples of Z. acuminate and Z. pseudomonticola. The first species (as currently understood, Acuña-Castillo 2010) is endemic to Costa Rica, in San José and Puntarenas Provinces from the Central Pacific region, where it is found in wet and rain forests of tropical and premontane elevations, between 100 and 1 200m altitude. Z. pseudomonticola is known from Eastern Puntarenas Province, Costa Rica, and Western Chiriquí Province, Panama; this species grows in wet and rain forests between 1 000 and 1 600m altitude in both countries and it is one of the few Central American species of Zamia that could be considered montane (along with Z. lindleyi Warsz. ex A. Dietr.and Z. gomeziana R.H. Acuña).
The study material of Z. acuminata was obtained from wild growing plants from the Mastatal sector of La Cangreja National Park, in Puriscal, San José at 450m altitude, while material of Z. pseudomonticola was collected from cultivated plants from the José María Orozco botanical garden, University of Costa Rica, San Pedro de Montes de Oca, San José at 1 200m altitude.
All samples for microscopic observation were about 0.5cm2, from the center of the leaflet. The samples were first fixed in Karnowsky solution (glutaraldehyde 2.5% / paraformaldehyde 2% / sodium phosphate buffer 0.1M, pH. 7.4), for 24 hours at 4ºC. The material was then washed in sodium phosphate buffer, and postfixed using 1% osmium tetroxide (OsO4). The samples were further washed in distilled water and were then processed for scanning electron microscopy or light microscopy.
The samples for scanning electron microscopy were dehydrated through an ascending series of ethanol solutions and then the material was left in tert-butanol for 24 hours at 4ºC, to be sublimed thereafter in an Eiko ID-2 vacuum sublimizer. The material was mounted on aluminum bases and covered with a 30nm thick gold-palladium layer. Samples were observed using a Hitachi S-570 scanning electron microscope with an acceleration voltage of 15KV (Sánchez & Espinoza 2005). For light microscopy, the material was dehydrated through an ascending series of propanone (acetone) solutions, then embeded in Spurr’s epoxy resin. The embedded samples were sectioned at 500nm thickness with a Power Tome PC (RMC Products) Ultramicrotome, dyed with Touluidin Blue and observed with an Olympus IX-51 inverted light microscope.
Results
The leaflets of Z. acuminata are characterized by their entire margins and their long, narrow tip which represents about 25% of the total leaflet length. The veins are flat on both the upper and lower surfaces of the leaflets, which results in a smooth texture. This texture is also found in Z. pseudomonticola, but in this species the leaflets are proportionally broader and with a shorter tip. In addition, the leaflet width diminishes more abruptly towards the tip as opposed to Z. acuminata.
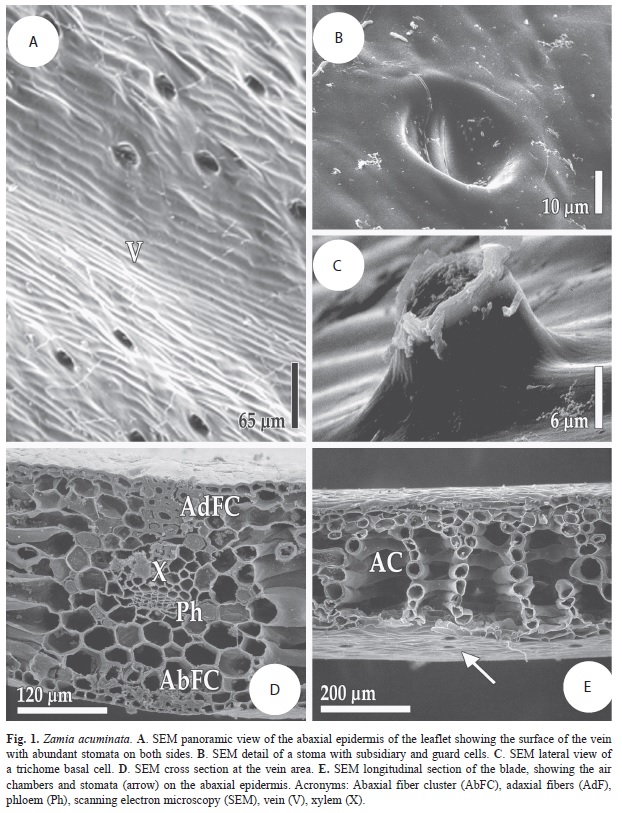
As with other species of the genus, Z. acuminata has hypostomatic leaflets. The stomata are restricted to the areas between the veins. The subsidiary cells show a concentrical pattern surrounding the guard cells (Fig. 1A), and the latter are in turn located at the bottom of a shallow depression (Fig. 1B). The other cells of the epidermis are sigmoid shaped and wider than the epidermal cells over the veins, which are narrower and more elongated. On the veins some trichome basal cells can be seen, which are ovoid or cylindrical. However, they are very scarce in this species (Fig. 1C). The adaxial surface is more uniform, and is characterized by the absence of stomata and trichomes.
In the vein area it is possible to see, immediately under the epidermis (both adaxially and abaxially), fiber bundles with different shapes. The adaxial fiber bundle is compact, narrow and extends from just under the epidermis to the periphery of the xylem. This bundle is four to five cells in width and up to seven cell layers in depth. The abaxial bundle is wider, but not as deep as the adaxial bundle since it only has two cell layers of depth. Towards the interior of the vein region the parenchyma becomes the dominant tissue and it is especially compact near the vascular tissues (Fig. 1D). In the area between the veins there are two to four cell layers of palisade parenchyma on top of the air chambers. The air chambers are large and their main axis is perpendicular to the main axis of the leaflets. The air chambers are formed by parenchymatic cells which are very enlongated and recurved, looking like branched tubes, forming walls that are one cell thick (Fig. 1E).
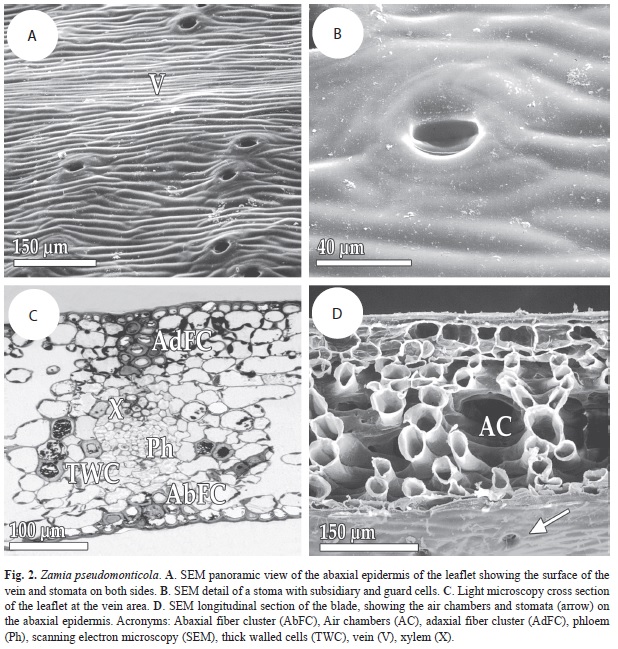
As with the previous species, Z. pseudomonticola also has hypostomatic leaflets. As occurs in other species of the genus, the epidermal cells over the veins and between the veins are dimorphic: the former are sigmoid and relatively wide in contrast to the latter, which are narrower and straighter (Fig. 2A). The stomata are sunken and are surrounded by two pairs of subsidiary cells (Fig. 2B). In the samples of this species no basal trichome cells were observed.
Under the adaxial epidermis in the vein area it is possible to distinguish a rectangular to wedge-shaped fiber bundle, relatively narrow but deep, one to three cells in width and three to five cell layers deep. Under the abaxial epidermis, there is another fiber bundle but this is less developed than in other studied species of the genus and with only one to three cell layers of depth. Between the fibers and the vascular tissues it is possible to see dense parenchyma, with polygonal cells that decrease in diameter gradually the closer they are to the xylem or the phloem. Some of these cells have developed thick walls (Fig. 2C).
In the area between the veins there are no conspicuous differences between this species and Z. fairchildiana. Palisade parenchyma formed by up to three cell layers is located above the spongy parenchyma, with very large air chambers, and whose axes are perpendicular to the main axis of the leaflet blade (Fig. 2D).
Discussion
As with other Zamia species, Z. acuminate and Z. pseudomonticola leaflets show an arrangement of traits for very different environments (Greguss 1968, Acuña-Castillo & Marín-Méndez 2012). On one hand there are adaptations for xeric environments, which are most likely evolutionary remnants from the ancestors of the genus. Such adaptations include the thick cuticle, the sunken stomata and the hypostomatic leaflets (Esau 1977). On the other hand, the leaflets also show adaptations for mesic, high humidity environments, such as the long drip tips and the large air chambers that allow the exchange of large volumes of gas.
The external macroscopic structure of the leaflets of Z. acuminata and Z. pseudomonticola is very similar to that observed in Z. fairchildiana. Most Zamia species from the Pacific slope of Southern Costa Rica and Western Panama have been given the name Z. fairchildiana, due to the similarities among the three species (Merello 2004). However, this confusion is a byproduct of their externally similar vegetative morphology and the incorrect taxonomic interpretation of Z. acuminate and Z. pseudomonticola. Some aspects of the leaflet anatomy are shared by all three species, such as the scarcity of trichome basal cells on mature leaflets, the presence of a rectangular- to wedge-shaped adaxial fiber bundle, and the relatively small abaxial fiber bundle (especially when compared to plicate-leaved Zamia species). However, the three species differ in some aspects of their foliar anatomy as well as their ecology and distribution. Even though basal trichome cells are infrequent in the studied, smooth-leaved species of Zamia, none were observed in the samples of Z. pseudomonticola, which could be because they are absent in mature leaflets of this species. The adaxial fiber bundle of Z. acuminata is more developed than in any other species of Zamia examined so far and also shows more thickwalled parenchyma cells.
Both species of this study inhabit wet and rain forests, but in their natural habitat Z. acuminate inhabits forests between 100 and 1 200m, which have a more defined seasonality, due to their more northwesterly geographic position, than the forests which are inhabited by Z. pseudomonticola. In addition, both species show significantly different morphometrics (Acuña- Castillo 2010). The apparently similar vegetative habits of Z. acuminata, Z. fairchildiana and Z. pseudomonticola, and the scarcity of specimens with reproductive structures, have caused problems for some botanists who have relied on vegetative characters to differentiate these species (e.g., Jones 2002, Merello 2004). This similarity could indicate that the three species are indeed closely related to each other and that they have diverged only recently. Events that could be responsible for such a divergence include the Pleistocene orogeny of the Fila Costeña and the Pacific slope branches of the Cordillera Talamanca (Denyer & Kussmaul 2000), as well as the fluctuation in temperature and precipitation regimes during the glacial and inter-glacial periods of the Quaternary (Haffer 1969, Colinvaux et al. 1996). These events are relatively recent in the geologic timescale; also, the spatial proximity of the geographical ranges of the three species suggests a relatively recent common ancestor for all three as occurs in other genera of Cycadales, such as Ceratozamia Brongn. (Vovides et al. 2004), Cycas (Hill 1996, 2004) and Encephalartos Lehm. (Vorster 2004), in which the species groups are distributed spatially close to each other, even though the individual species have allopatric distributions. The habitat changes and isolation between populations, resulting from orogeny and climatic changes, even in the lapse of just tens of thousands of years, could be responsible for the development of the observed anatomical differences of the studied species, as occurs in Ceratozamia in Southern Mexico, where species of the same species group, separated by just tens of kilometers, show significant and noticeable differences in their leaf architecture (Pérez-Farrera et al. 2004).
Acknowledgments
We are indebted to the staff of CIEMic for their valuable help during this research, especially to Ethel Sánchez-Chacón and Alexander Rodríguez for their advice with the electron microscopy study and the image processing. We are also thankful to Paul Hanson and three anonymous reviewers for their evaluation of the manuscript and their suggestions. Financial support for this research was provided by Vicerrectoría de Investigación, Universidad de Costa Rica through grant No 810-B0-042.
References
Acuña-Castillo, R. 2010. Revisión taxonómica, morfológica, morfometría y distribución geográfica de Zamia (Zamiaceae) en Costa Rica. Tesis de Maestría, Universidad de Costa Rica, Costa Rica. [ Links ]
Acuña-Castillo, R. & W. Marín-Méndez. 2012. Some aspects of the foliar anatomy of two Zamia species from Costa Rica: Z. fairchildiana L.D. Gómez and Z. neurophyllidia D.W. Stev. Mem. N. Y. Bot. Gard. 106: 192-203. [ Links ]
Caputo, P., S. Cozzolino, L. Gaudio, A. Moretti & D.W. Stevenson 1996. Karyology and phylogeny of some Mesoamerican species of Zamia (Zamiaceae). Amer. J. Bot. 83: 1513-1520. [ Links ]
Colinvaux, P.A., P.E. De Oliveira, J.E. Moreno, M.C. Miller & M.B. Bush. 1996. A long pollen record from lowland Amazonia: forest and cooling in glacial times. Science 274: 85-88. [ Links ]
Denyer, P. & S. Kussmaul. 2000. Geología de Costa Rica. Tecnológica, Cartago, Costa Rica. [ Links ]
Eckenwalder, J.E. 2008. Conifers of the world. Timber, Portland, Oregon, USA. [ Links ]
Esau, K. 1977. Anatomy of Seed Plants. John Wiley, New York, USA. [ Links ]
Greguss, P. 1968. Xylotomy of the living cycads, with a description of their leaves and epidermis. Akadémiai Kiadó, Budapest, Hungary. [ Links ]
Haffer, J. 1969. Speciation in Amazonian forest birds. Science 165: 131-137. [ Links ]
Hill, K.D. 1996. A taxonomic revision of the genus Cycas (Cycadaceae) in Australia. Telopea 7: 1-64. [ Links ]
Hill, K.D. 2004. Character evolution, species recognition and classification concepts in the Cycadaceae, p. 23-44. In T. Walters & R. Osborne (eds.). Cycad Classification: Concepts and Recommendations. CABI Publishing, Wallingford, United Kingdom. [ Links ]
Hill, K.D., M.W. Chase, D.W. Stevenson, H.G. Hills & B. Schutzman. 2003. The families and genera of cycads: a molecular phylogenetic analysis of cycadophyta based on nuclear and plastid DNA sequences. Int. J. Plant Sci. 164: 933-948. [ Links ]
Jones, D.L. 2002. Cycads of the World. Smithsonian Institution, Washington D.C., USA. [ Links ]
Merello, M. 2004. Zamiaceae, p. 14-16. In B.E. Hammel, M.H. Grayum, C. Herrera & N. Zamora (eds.). Manual de Plantas de Costa Rica, Vol. 2. Monogr. Syst. Bot. Missouri Bot. Gard. 92: 14-16. [ Links ]
Newell, S.J. 1985. Intrapopulational variation in leaflet morphology of Zamia pumila L. in relation to microenviroment and sex. Amer. J. Bot. 72: 217-221. [ Links ]
Newell, S.J. 1989. Variation in leaflet morphology among populations of Caribbean cycads (Zamia). Amer. J. Bot. 76: 1518-1523. [ Links ]
Norstog, K. 1981. Karyotypes of Zamia chigua (Cycadales). Caryologia 34: 255-260. [ Links ]
Norstog, K. & J.T. Nicholls. 1998. The Biology of the Cycads. Comstock, Ithaca, New York, USA. [ Links ]
Osborne, R., M. Calonje, K. Hill, L. Stanberg & D.W. Stevenson. 2012. The World list of cycads. Mem. N. Y. Bot. Gard. 106: 480-508. [ Links ]
Pérez-Farrera, M.A., A. Vovides, L. Hernández-Sandoval, D. González & M. Martínez. 2004. A morphometric analysis of the Ceratozamia norstogii complex (Zamiaceae), p 127-136. In T. Walters & R. Osborne. Cycad Classification: Concepts and Recommendations. CABI, Wallingford, United Kingdom. [ Links ]
Sánchez, E. & A. Espinoza. 2005. Ultrastructure of Oryza glumaepatula Steud, a wild rice species endemic of tropical America. Rev. Biol. Trop. 53: 15-22. [ Links ]
Stevenson, D.W. 1981. Observations on ptyxis, phenology, and trichomes in the Cycadales and their systematic implications. Amer. J. Bot. 68: 1104-1114. [ Links ]
Stevenson, D.W. 1990. Morphology and systematics of the Cycadales. Mem. N. Y. Bot. Gard. 57: 8-55. [ Links ]
Stevenson, D.W. 2001. Orden Cycadales. In R. Bernal & E. Forero (eds.). Flora de Colombia 21: 1-92. [ Links ]
Stevenson, D.W., K. Norstog & D. Molsen. 1996. Midribs of cycad pinnae. Brittonia 48: 67-74. [ Links ]
Takhtajan, A. 2009. Flowering Plants. Springer, Berlin, Germany. [ Links ]
Taylor, T.N., E.L. Taylor & M. Krings. 2009. Paleobotany: the biology and evolution of fossil plants. Academic, Burlington, Massachusetts, USA. [ Links ]
Vorster, P. 2004. Classification concepts in Encephalartos (Zamiaceae), p. 69-83. In T. Walters & R. Osborne (eds.). Cycad Classification: Concepts and Recommendations. CABI, Wallingford, United Kingdom. [ Links ]
Vovides, A.P. 1983. Systematic studies on the Mexican Zamiaceae. I. Chromosome numbers and karyotypes. Amer. J. Bot. 70: 1002-1006. [ Links ]
Vovides, A.P., M.A. Pérez-Farrera, D. González & S. Avendaño. 2004. Relationships and phytogeography in Ceratozamia (Zamiaceae), p. 109-125. In T. Walters & R. Osborne (eds.). Cycad Classification: Concepts and Recommendations. CABI, Wallingford, United Kingdom. [ Links ]
*Correspondencia:
Rafael Acuña-Castillo: Escuela de Biología, Centro de Investigación en Estructuras Microscópicas (CIEMic), Universidad de Costa Rica, San José, Costa Rica. P.O. Box 11501-2060. rafael.asurbanipal@gmail.com
Walter Marín-Méndez: Escuela de Biología, Centro de Investigación en Estructuras Microscópicas (CIEMic), Universidad de Costa Rica, San José, Costa Rica. P.O. Box 11501-2060. walter.marin@ucr.ac.cr
1. Escuela de Biología, Centro de Investigación en Estructuras Microscópicas (CIEMic), Universidad de Costa Rica, San José, Costa Rica. P.O. Box 11501-2060; rafael.asurbanipal@gmail.com, walter.marin@ucr.ac.cr
Received 19-III-2012. Corrected 20-VIII-2012. Accepted 24-IX-2012.


{kind=link}
{kind=link}