










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.61 n.1 San José Mar. 2013
Dietary breadth of the animal protein consumed by riverine communities in the Tapajós National Forest, Brazil
*Dirección para correspondencia:
Abstract
In small-scale human settlements, the acquisition of animal protein is strictly related to subsistence activities, and yours dietary habits are determined by the availability and the selectivity permitted by the diversity of these resources. This study analyzed the consumption of animal protein sources in seven traditional riverine communities of the Tapajos National Forest, located in Eastern Brazilian Amazonia, considering fish, game meat and domestic animals. The analysis of animal protein consumption was based on the assumptions of the diet breadth model and the Optimal Foraging Theory. We compared diet breadths between communities and between rainy and dry seasons. The study focused on seven traditional riverside communities, six of them distributed along the right bank of the Tapajos River and one on the right bank of the Cupari River. Data collection was performed in four fields trips, two in the rainy season (May and July) and two in the dry season (September and November) in 2010. Data were collected through semi-structured interviews where the informant mentioned the source of animal protein consumed in the last three meals and which would be consumed at the next meal, if possible. We carried out a total of 470 interviews, where we documented 1 512 meals, and in only 12% of the meals there was no consumption of any animal protein source. The fish was consumed in 60.4% of the meals, being the most important source of animal protein consumed, differing significantly from other protein sources (χ²=23.79, df=5, p<0.001). A total of 11 species of wild animals and 46 species of fish were consumed. The choice in the consumption of game meat consisted on Tayassu pecari, Hydrochoerus hidrochaeris and Cuniculus paca, while the preference for fish consumption included Plagioscion spp., Astronotus spp., Cichla spp. and Leporinus spp.. The Simpson index did not vary significantly between the rainy and dry season (N=6, t=1.25, p=0.267) or between communities (N=6, t=5, p=0.42), although São Francisco das Chagas have significantly higher consumption of game meat (χ²=370.41, df=25, p<0.001). Fishing is an activity of paramount importance to these communities, and factors that lead to decreased availability of fish may lead to subsequent increase in hunting pressure. For the conservation of preserve of both wildlife natural resources and practices of subsistence of riverine communities of the Tapajós National Forest, it is necessary to ensure the maintenance of fish stocks and the protection of the Tapajós River areas large enough to maintain viable populations of wild animals and more tolerant to hunting and habitat loss.
Key words: dietary breadth, fish, game meat, Tapajós National Forest, traditional riverine communities.
Resumen
En pequeños asentamientos rurales, la adquisición de proteína animal está estrechamente relacionada con actividades de subsistencia, y su hábitos alimentares son determinados por la disponibilidad y diversidad de estos recursos. Este estudio examinó el consumo de pescado, caza y animales domésticos en siete comunidades tradicionales ribereñas de la Floresta Nacional do Tapajós, ubicadas en la Amazonia oriental Brasileña. La análisis se basa en los supuestos de modelos de la amplitud de la dieta y la teoría de forrajeo óptimo, se comparó la amplitud de dieta en siete comunidades ribereñas en periodo seco y lluvioso. La recolección de datos se realizó en cuatro viajes, dos en la temporada de lluvias y dos en la estación seca en el 2010, mediante entrevistas semi-estructuradas. El pescado fue consumido en el 60.4% de las comidas, siendo la fuente de proteína animal más consumida. Un total de 11 especies de animales silvestres y 46 especies de pescado fueron documentadas y la preferencia en el consumo cayó sobre T. pecari, C. paca y Hydrochoerus hidrochaeris y la preferencia por el consumo de pescado cayó sobre Plagioscion spp. Astronotus spp., Cichla spp. y Leporinus spp.
Palabras clave: amplitud de la dieta, carne de caza, comunidades tradicionales ribereñas, Floresta Nacional do Tapajós, pescado.
In small-scale human settlements, the acquisition of animal protein is strictly related to subsistence activities (Murrieta et al. 1999, Fa et al. 2002). Currently, the overhunting is considered a major threat to wildlife hunting in the humid tropics when extinguishing many species locally, which in turn threatens the dietary security of the local human populations that depend on game meat for subsistence and income (Milner-Gulland et al. 2003).
From the perspective of human ecology, the analysis of the diet based on concepts such as diversity and niche permits the systematic analysis of the exploitation of natural resources by a local population (Hanazaki & Begossi 2000). The ecological multidimensional niche approach meets all the environmental variables and resources, where a species is able to persist and maintain a stable population size (Wiens & Graham 2005). The breadth of each niche dimention can be estimated, showing the degree of spatial generalization of the use of resources (Hanazaki & Begossi 2003). If prey species composition by this population are considered to be a dimension of its niche, the niche will likely be narrower where the preferred species are available, but will become progressively wide as selectivity is reduced (MacCord & Begossi 2006).
Niche breadth encompasses a variety of factors including dietary breadth, defined by Hames & Vickers (1982) as the set of prey captured, which are selected initially by body size and secondarily by capture rates. These authors found that the age of the settlement and its distance from urban centers are the principal factors determining the efficiency of hunting, confirming predictions derived from Optimal Foraging Theory (OFT). Optimal Foraging Theory predicts that, in the Neotropics, human hunters will systematically target larger-bodied species, which provide the largest possible return in meat per unit of time or energy invested (Jerozolimski & Peres 2003). Therefore, assessing the extent of diet provides us with indirect information about the status of conservation of local wildlife and fishery used as a food source by traditional populations.
In addition to the natural factors that determine the distribution, density, and resilience of game species, patterns of human occupation mold their availability within a given area (Hill et al. 1984, Vickers 1988). The dietary habits of the human population are determined, in turn, by the availability of resources and the selectivity permitted by the diversity of these resources.
In the rivers of the Amazon Basin, the availability of fishery resources is strongly influenced by seasonal cycles, with the aquatic fauna being restricted to permanent bodies of water during the dry season, when the flood plains are exposed. During this season, the exploitation of fish and turtles increases considerably as a response to the greater concentration of this resource (Batista et al. 1998, Pezzuti et al. 2004).
In the present study, we tested the null hypothesis that the consumption of fish and game meat by the residents of traditional riverine communities in the Tapajós National Forest do not varies between the rainy and the dry seasons. We also predicted that the exploitation of these different sources of protein would vary according to population size, history of occupation, and the degree of isolation of the community, based on the dietary breadth model.
Material and Methods
Study site: The Tapajós National Forest (02°45’-04º10’ S and 054°45’-055º30’ W) is a protected area and was the second national forest established in Brazil’s Northern region, being determined by Federal Decree number 73684 of February, 1974. With a total area of 5 449.27km2, the protected area includes portions of the municipalities of Aveiro, Belterra, Placas, and Rurópolis, in the Brazilian state of Pará.
Bordered to the west by the Tapajós River, the Northern limit of the National Forest is perpendicular to km 50 of the Cuiabá-Santarém Highway, the BR-163, while it is bordered to the East by the BR-163 (IBAMA 2005). The Southern limit corresponds to the TransAmazon highway and the Cupari and Cuparaitinga Rivers. The Tapajós National Forest is a federal conservation unit of the category of sustainable use, and its main purpose is to promote the sustainable exploitation of forest resources and scientific research, with emphasis on the development of multiple procedures for the rational use of native forests (IBAMA 2005).
The predominant climate is humid tropical, consistent with Köppen’s Ami category, with annual thermal variation of 5ºC, mean annual temperature of 25.5ºC, maximum of 30.6°C and minimum of 21ºC (IBAMA 2005). Evapotranspiration is highest between October and January, while annual precipitation varies between 600-3 000mm, with a mean of 2 000mm and marked droughts during El Niño events (Nepstad et al. 2002).
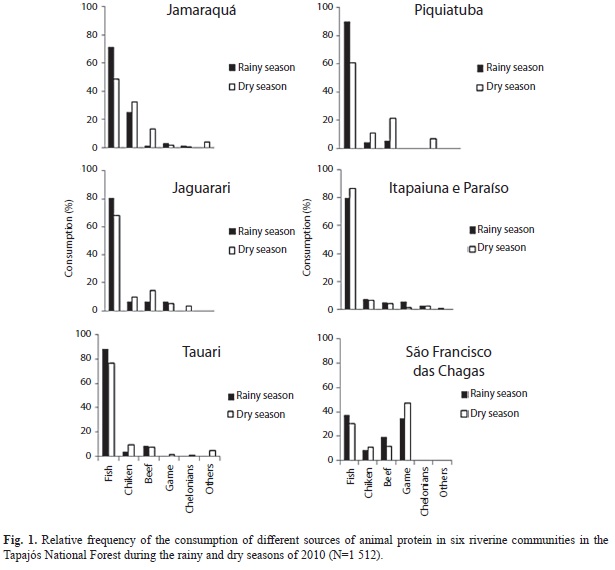
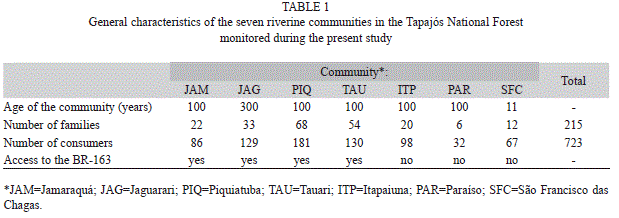
The region was originally occupied by tribes of the Tapajó people, an ancient and complex society first recorded in the 17th Century. The present day population of the Tapajós National Forest is distributed among 31 communities, 21 of which are located on the margin of the Tapajós River, with a further three on the Cupari River, and five along the BR-163 highway and in the town of Aveiro, totalizing of 10 696 residents (IBAMA 2005). The present study focused on six communities located on the right bank of the Tapajós River (Jamaraquá, Jaguarari, Piquiatuba, Tauari, Itapaiuna, and Paraíso) and one (São Francisco das Chagas) on the right bank of the Cupari River. These communities were selected during two preliminary visits to the study area in November, 2009 and January, 2010. The communities were chosen based on their location, whether or not they have access to the BR-163 highway, as well as the minimum distance of 10km between them (Fig. 1).
The demographic characteristics of the communities are presented in table 1. Paraíso, Itapaiuna, and São Francisco das Chagas are not linked directly to the BR-163 highway, and are more isolated from the nearest urban centers associated with this highway than the other communities included in the present study.
Data collection: In this study there were four field trips, two in the dry season (May and July) and two in the rainy season (September and November) in 2010. The method consisted in the application of semi-structured interviews, where the basic unit was the household. The method basically consists in interviewing with respondent following a script of questions regarding the source of animal protein consumed in the last three meals, and what is the source of animal protein for the next meal. The method does not allow analyzing changes in consumption within the home, yet provides a reasonable estimate of food consumption at the population level. Respondents were residents of the communities sampled, mostly fishermen and agriculturists. Based on the consumption data, we analyzed the dietary breadth of communities sampled during the study period. Paraíso and Itapaiuna were considered to be a single community for statistical analyses, given that they are only 500m apart, and share a common history of occupation.
Measurement of dietary breadth: To analyze the collected data, we use the dietary breadth model, which was evaluated using Simpson index as indicator, following Bodmer (1995), given the direct and proportional relationship between it and the diversity of species exploited. We used the frequency of consumption of meals to measure species diversity through the Simpson index, where the total sources of animal protein consumed by the community is the species richness. Simpson’s index (λ) varies from 0-1, and values closer to 1 indicate a higher probability that individuals belong to the same species, that is, that dominance is high and diversity is low. This index is one measure of species richness, and reflects the probability that two individuals selected randomly from the community will belong to the same species (Magurran 2004). Values are provided by the formula λ=1/Σpi2, where pi is the relative abundance of species i, that is, the number of records of this species divided by the total number of records in the sample (Magurran 2004).
Seasonal (rainy vs. dry season) differences in the dietary breadth, that is, the contribution of fish, game meat, and other sources of animal protein, were analyzed using Student’s t test for dependent samples, here season is the independent variable and the Simpson indexes were dependent variables. While possible differences in the frequency of consumption of the different sources was evaluated using Friedman’s test, where class of items consumed is the independent variable and frequency of consumption dependent variables. The Chi-square test was used to evaluate the differences in the consumption of fish and game meat among the communities, using an approach adapted from Hanazaki et al. (2009).
Results
A total of 470 interviews were applied during 2010 (Jamaraquá=52, Jaguarari=90, Piquiatuba=108, Tauari=102, Itapaiuna/Paraíso=85, São Francisco das Chagas=33). The members of the local communities consume animal protein twice a day, during their main meals. No animal protein was consumed in only 187 of the 1 512 meals recorded during this procedure. The interviews were applied to all residents in the community, and were recorded less than four meals, for there was no prediction of what would be consumed at the next meal or simply there was no meals made.
With the exception of São Francisco das Chagas, fish was by far the most important source of animal protein in all the studied communities in both seasons (Fig. 1). The consumption of fish was normally greater during the rainy season, with the exception of Itapaiuna and Paraíso, while in São Francisco das Chagas, the consumption of game meat surpassed that of fish during the dry season.
Overall, the consumption of fish was significantly greater than that of other sources of animal protein (χ²=23.79, d.f.=5, p<0.001), while the relative consumption of fish and game meat was significantly different in São Francisco das Chagas in comparison with the other communities (χ²=370.41, d.f.=25, p<0.001). The consumption of game meat in the communities that have access to the BR-163 highway (Table 1) also differed significantly from that of the communities which do not have access to the highway (χ²=81.15, d.f.=5, p<0.001), where São Francisco das Chagas had the highest consumption of game meat.
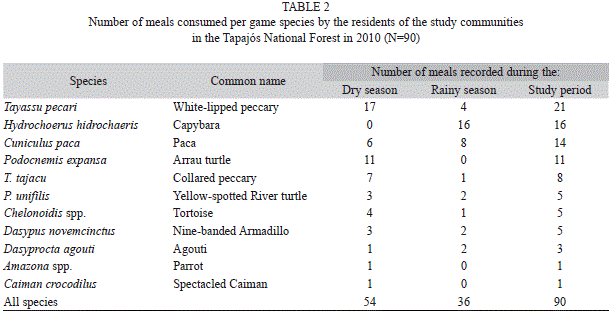
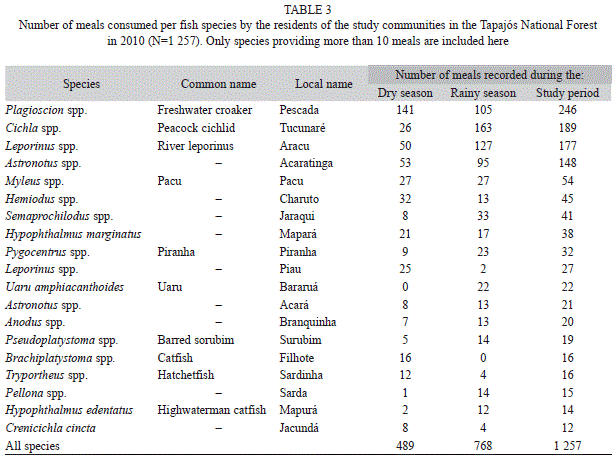
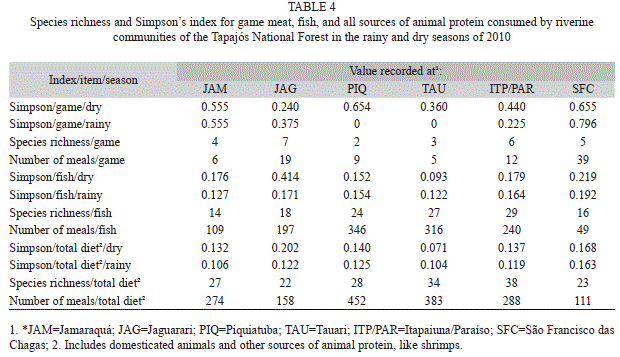
A total of 11 game species were recorded (Table 2) and 46 fish species were exploited during the period monitored (Table 3). In both cases, while some species were consumed relatively frequently, the majority were included in meals more rarely, a dominance pattern observed more explicitly in the consumption of fish. The general Simpson index (Table 4) recorded for each community did not varied significantly between the rainy and dry seasons (n=6, t=1.248, p=0.267). In addition, no seasonal differences were found when considering fish (n=6, t=1.22, p=0.28) or game meat separately (n=6, t=1.29, p=0.25). Similarly, no significant differences in dietary breadth were found among communities (n=6, t=5, p=0.42).
Discussion
According to the dietary breadth model, the number of less preferred items included in the diet will tend to increase as the preferred species become less abundant, leading to a greater generalization of the diet (MacArthur & Pianka 1966), and an increase in its breadth (Hames & Vickers 1982, MacCord & Begossi 2006). While a greater breadth may reflect the scarcity of preferred species, in the present case, it may also be a result of the increased consumption of domesticated meat, such as chicken (in Jamaraquá) and beef (in Piquiatuba), a pattern observed in North America by Gossard & York (2003). As game meat was consumed relatively rarely, the estimates of the use of this resource provided by the interviews may be somewhat unreliable. It is important to note that the diversity of game species is inversely proportional to Simpson’s index (Magurran 2004).
Consumption of fish and game meat: The consumption of fish in the study communities was significantly greater than that of any other source of animal protein. A similar pattern has been recorded on Marajó Island in Eastern Amazonia (Murrieta et al. 1999) and on the Negro (Pezzuti et al. 2004) and Juruá Rivers (Begossi et al. 1999, 2004) in the Western Amazon Basin. Fish is the main source of animal protein for all communities except São Francisco das Chagas, where fish and game were equally important during the rainy season, while the consumption of game meat exceeded that of fish during the dry season.
The significant difference observed at São Francisco das Chagas may be related to a number of specific factors, such as the reduced fish stocks on the Cupari River in comparison with the Tapajós, given that the consumption of game meat tends to be inversely related to the availability of this resource (Milner-Gulland et al. 2003). This community is also more recent and more isolated than the others, which may mean that local game populations may be less impacted than those at other sites, a pattern observed throughout the Neotropics (Lopes & Ferrari 2000, Peres 2000, Jerozolimski & Peres 2003). This also suggests that game may be preferred over fish, in general, and that the prevalence of fish in the diets of the other communities is the result of the relative scarcity of game at these sites.
Access to the BR-163 highway may be an additional factor influencing the consumption of game meat. In addition to being farthest from urban centers, Itapaiuna, Paraíso, and São Francisco das Chagas have no access to the BR-163 highway, which results in higher costs and longer delays for the acquisition of foodstuffs.
Dietary breadth: It seems likely that the analysis of dietary breadth (Simpson’s index) for the consumption of game meat is less robust than that for fish, given the greatly reduced sample available for game, with only one record available for some species (Table 2). The fish diet is characterized by a higher diversity and lower dominance of species, as well as greater species richness, in comparison with game meat. This reflects the greater variety of fish species appropriate for consumption (Pezzuti et al. 2004), as well as the relative efficiency and productivity of fishing in comparison with hunting.
The smallest diversity of fish species exploited at São Francisco das Chagas, a relatively recently-established and isolated community, corroborates the basic assumption of the dietary breadth model (Hames & Vickers 1982, Alvard 1993). Even though fish is consumed less frequently in this community, the preferred species predominate in the diet. Similarly, T. pecari, H. hydrocaeris, and C. paca are the preferred game species in this community, given their large body size, while P. expansa is the most consumed reptile. This is as predicted by OFT (Hames & Vickers 1982, Jerozolimski & Peres 2003), with the largest species being preferred due to their high returns of meat per unit of time and energy invested in harvesting. The probability of consuming smaller, less preferred species depends on the availability of only the most preferred items (Hawkes & O’Connell 1992).
The most consumed fish species in the Tapajós National Forest belonged to the genera Plagioscion spp., Cichla spp., Leporinus spp., and Astronotus spp., Plagioscion spp. and Astronotus spp. were exploited continuously throughout the study period, whereas Cichla spp. and Leporinus spp. were consumed mainly during the rainy season.
Dietary breadth did not vary significantly between seasons, which reject the hypothesis of systematic seasonal variation in the exploitation of sources of animal protein. Despite this, some fish species, such as Cichla spp. and Leporinus spp., presented marked fluctuations in consumption, with marked peaks at the beginning of the rainy season, but reduced consumption during the rest of the year. This period coincides with the highest relative abundance of fish in the region, due to the fact that river levels are at their lowest, which limits the dispersion of the animals as they become trapped in river channels and isolated bodies of water, facilitating their capture. Similar patterns have been observed on the Juruá (Begossi et al. 1999) and Negro Rivers (Pezzuti et al. 2004).
The consumption of turtle meat also occurred primarily in the dry season, which was the only source of animal protein other than Cichla spp. and Leporinus spp., but overall turtles did not represent an important resource in the study communities. Seasonal items tend to be more important where other sources of animal protein are insufficient, leading to an increase in seasonal variation. Among the Achés indigenous people of eastern Paraguay, Hill et al. (1984) observed that dietary breadth was related to the availability of foods, which varied seasonally, leading to variations in the social behavior of the group.
Implications for conservation: The monitoring of fluctuations in the behavior of riverine communities is a simple, but important ecological tool for the prediction and, if necessary, development of strategies to avoid overexploitation of faunal resources and degradation of the environment (MacCord & Begossi 2006). Hawkes & O’Connell (1992) observed that, as hunters tend to target the most worthwhile species, preferred items are rarely absent from the diet, but that increasing numbers of consumers lead to the inclusion of less preferred items in the diet, and the overexploitation of the more preferred items, making them less available to these consumers.
Based on the data collected in the present study, the diet of the riverine communities of the Tapajós National Forest appears to be in equilibrium with their traditional eating habits, suggesting no major influence of urban input or anthropogenic impacts. Despite the fact that the larger game species are not consumed frequently, it is necessary to evaluate the annual variation of the abundance of game species. We conclude that fishing satisfies the animal protein basic needs of the communities, and that seasonal climatic fluctuations have little effect their diet. However, the importance of fish to these communities means that factors that may reduce fish stocks, such as an increase in commercial fishing operations or significant growth in the population of the communities, may lead to an increase in hunting pressure, given that the excessive exploitation of game species is linked directly to the integrity of the local subsistence economy (Milner-Gulland et al. 2003).
Firstly, it is necessary to develop measures that will guarantee the maintenance of the fishery stocks at levels required for the subsistence of local riverine communities. One way of doing this would be to create restricted or prohibited fishing zones, given the need for maintenance of a diversity of environments for the commercial or subsistence exploitation of aquatic resources (McGrath et al. 1993, Ruffino et al. 1999, Da Costa et al. 1999), considering the ecology of the species involved. In addition, it will also be important to protect areas large enough to maintain viable populations of game species, which will be more resilient to harvesting and the loss of peripheral habitats (Dias 1996). A better understanding of the interrelated factors which promote hunting and the consumption of game meat is also needed (Milner-Gulland et al. 2003). Ultimately, overcoming the conceptual division between hunting and fishing is also important, given the direct interaction between the two activities observed in many cases (Pezzuti et al. 2004, Milner-Gulland et al. 2003).
Acknowledgments
I thank ICMBio for the help in field activities, Adriano Jerozolimski, Alpina Begossi, Rômulo Alves and Glenn Shepard Jr. for constructive criticism related to the original manuscript, Stephen Ferrari and Lucas Mello for translating and reviewing this manuscript into English.
References
Alvard, M.S. 1993. Testing the “ecologically noble savage” hypothesis: Interspecific prey choice by Piro hunters of Amazonian Peru. Hum. Ecol. 21: 355-387. [ Links ]
Batista, V.S., A.J.I. Silva, C.E.C. Freitas & D. Freire-Brasil. 1998. Characterization of the fishery in riverine communities in the Low-Solimões/High-Amazon region. Fish. Manag. Ecol. 5: 101-117. [ Links ]
Begossi, A., R.A.M. Silvano, B. Amaral & O.T. Oyakawa. 1999. Uses of fish and game by inhabitants of an extractive reserve (upper Juruá, Acre, Brazil). Environ. Dev. Sust. 1: 1-21. [ Links ]
Begossi, A., N. Hanazaki & R.M. Ramos. 2004. Food chains and the reasons for food fish taboos among Amazonian and Atlantic Forest fishers (Brazil). Ecol. Appl. 14: 1334-1343. [ Links ]
Bodmer, R.E. 1995. Managing Amazonian wildlife: biological correlates of game choice by detribalized hunters. Ecol. Appl. 5: 872-877. [ Links ]
Da Costa, L.R.F., R. Barthem & M.A.V. Correa. 1999. Manejo da pesca do tambaqui nos lagos de várzea da RDSM, p. 142-158. In H. Queiroz & W.G.R. Crampton (eds.). Estratégias para manejo de recursos pesqueiros em Mamirauá. Sociedade Civil Mamirauá/ CNPq, Brasília, Brasil. [ Links ]
Dias, P.C. 1996. Sources and sinks in population biology. Trend. Ecol. Evol. 11: 326-330. [ Links ]
Fa, J.E., C.A. Peres & J. Meeuwing. 2002. Bushmeat Exploitation in Tropical Forests: an International Comparison. Conservat. Biol. 16: 232-237. [ Links ]
Gossard, M.H. & R. York. 2003. Social structural influences on meat consumption. Hum. Ecol. Rev. 10: 1-9. [ Links ]
Hawkes, K. & J. O’Connell. 1992. On Optimal Foraging Models and subsistence transitions. Curr. Anthropol. 33: 63-66. [ Links ]
Hames, R.B. & W.T. Vickers. 1982. Optimal Diet Breadth Theory as a Model to Explain Variability in Amazonian Hunting. Am. Ethnol. 9: 358-378. [ Links ]
Hanazaki, N. & A. Begossi. 2000. Fishing and Niche Dimension for Food Consumption of Caiçaras from Ponta do Almada (Brazil). Hum. Ecol. Rev. 7: 52-62. [ Links ]
Hanazaki, N. & A. Begossi. 2003. Does fish still matter? Changes in the diet of two Brazilian fishing communities. Ecol. Food Nutr. 42: 279-301. [ Links ]
Hanazaki, N., R.R.N. Alves & A. Begossi. 2009. Hunting and use of terrestrial fauna used by Caiçaras from the Atlantic Forest coast (Brazil). J. Ethnobiol. Ethnomed. 5: 36. [ Links ]
Hill, K., K. Hawkes, M. Hurtado & H. Kaplan. 1984. Seasonal Variance in the Diet of Ache Hunter-Gatherers in Eastern Paraguay. Hum. Ecol. 12: 101-135. [ Links ]
IBAMA. 2005. Plano de Manejo da FLONA Tapajós, Instituto Brasileiro de Meio Ambiente e Recursos Naturais Renováveis – IBAMA, Santarém, Brasil. [ Links ]
Jerozolimski, A. & C.A. Peres. 2003. Bringing home the biggest bacon: a cross-site analysis of the structure of hunter-kill profiles in Neotropical forests. Biol. Conservat. 111: 415-425. [ Links ]
Lopes, M.A. & S.F. Ferrari. 2000. Effects of human colonization on the abundance and diversity of mammals in eastern Brazilian Amazonia. Conservat. Biol. 14: 1658-1665. [ Links ]
MacArthur, R.H. & E.R. Pianka. 1966. On optimal use of patchy environment. Am. Nat. 100: 603-609. [ Links ]
MacCord, P.L. & A. Begossi. 2006. Dietary changes over time in a caiçara community from the Brazilian Atlantic Forest. Ecol. Soc. 11: 38. [ Links ]
McGrath, D.G., F. Castro, C.R. Futemma, B.D. Amaral & J.C. Araújo. 1993. Fisheries and the evolution of resource management on the lower Amazonian floodplain. Hum. Ecol. 22: 167-195. [ Links ]
Magurran, A.E. 2004. Measuring Biological Diversity. Blackwell, Vitoria, Australia. [ Links ]
Milner-Gulland, E.J., E.L. Bennet & the SCB. 2002 Annual Meeting Wild Meat Group. 2003. Wild meat: the bigger picture. Trends. Ecol. Evol. 18: 351-357. [ Links ]
Murrieta, R.S.S., A.L. Dufour & A. Siquiera. 1999. Food consumption and subsistence in three caboclo populations on Marajó Island, Amazonia, Brazil. Hum. Ecol. 27: 455-475. [ Links ]
Nepstad, D.C., P. Moutinho, M.B. Dias Filho, E. Davidson, G. Cardinot, D. Markewitz, R. Figueiredo, N. Vianna, J. Chambers, D. Ray, J.B. Guerreiros, P. Lefebvre, L. Sternberg, M. Moreira, L. Barros, F.Y. Ishida, I. Tohlver, E. Belk, K. Kalif & K. Schwalbe. 2002. The effects of partial throughfall exclusion on canopy processes, aboveground production, and biogeochemistry of an Amazon forest. J. Geophys. Res. 107: 1-18. [ Links ]
Peres, C.A. 2000. Effects of subsistence hunting on vertebrate community structure in Amazonian forests. Conservat. Biol. 14: 240-253. [ Links ]
Pezzuti, J.C.B., J.P. Lima, D. Félix-Silva & G.H. Rêbelo. 2004. A caça e a pesca no Parque Nacional do Jaú, Amazonas, p. 213-230. In S.H. Borges, S. Iwanaga, C.C. Durigan & M. Pinheiro (eds.). Janelas para a Biodiversidade no Parque Nacional do Jaú. Fundação Vitória Amazônica, Manaus, Brasil. [ Links ]
Ruffino, M.L., B. Mitlewski, V.J. Isaac & P.R.S. Oliveira. 1999. Lago Grande de Monte Alegre: uma análise das suas comunidades pesqueiras, Recursos pesqueiros do Médio Amazonas: abordagem socioeconômica - Série Estudos Pesca, coleção Meio Ambiente, no 21. IBAMA, Brasília, Brasil. [ Links ]
Wiens, J.J. & C.H. Graham. 2005. Niche Conservatism: Integrating Evolution, Ecology, and Conservation Biology. Annu. Rev. Ecol. Evol. Systemat. 36: 519-539. [ Links ]
Vickers, W.T. 1988. Game Depletion Hypothesis of Amazonian Adaptation: Data from a Native Community. Science 239: 1521-1522. [ Links ]
*Correspondencia a:
Raphael Alves Fonseca. Museu Paraense Emílio Goeldi/MPEG/Coordenação de Zoologia - Universidade Federal do Pará, Av. Perimetral 1901, Terra Firme, 66077-530, C.P. 399, Belém/PA, Brazil; raphazed@hotmail.com
Juarez Carlos Brito Pezzuti. Núcleo de Altos Estudos Amazônicos/NAEA - Universidade Federal do Pará, Av. Augusto Correa 01, Belém/PA, 66075-110, Brazil; juca@ufpa.br
1. Museu Paraense Emílio Goeldi/MPEG/Coordenação de Zoologia - Universidade Federal do Pará, Av. Perimetral 1901, Terra Firme, 66077-530, C.P. 399, Belém/PA, Brazil; raphazed@hotmail.com
2. Núcleo de Altos Estudos Amazônicos/NAEA - Universidade Federal do Pará, Av. Augusto Correa 01, Belém/PA, 66075-110, Brazil; juca@ufpa.br
Received 07-X-2011. Corrected 19-VII-2012. Accepted 20-VIII-2012


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}