










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.60 suppl.3 San José Nov. 2012
Characterization of deepwater invertebrates at Isla del Coco National Park and Las Gemelas Seamount, Costa Rica
*Dirección para correspondencia:
Abstract
The deepwater faunas of oceanic islands and seamounts of the Eastern Tropical Pacific are poorly known. From 11-22 September 2009, we conducted an exploration of the deepwater areas around Isla del Coco National Park and Las Gemelas Seamount, located about 50km southwest of Isla del Coco, Costa Rica using a manned submersible to survey the seafloor habitats. The goal of the exploration was to characterize the habitats and biota, and conduct quantitative surveys of the deepwater portions of Isla del Coco National Park and Las Gemelas. We completed a total of 22 successful submersible dives, spanning more than 80hr underwater, and collected a total of 36hr of video. With respect to invertebrates, our objectives were to gather quantitative information on species composition, density, distribution and habitat associations as well as to compare the invertebrate communities between the two sites. A total of 7 172 invertebrates were counted from analysis of the video collected on this project. Larger organisms were counted and placed into 27 taxonomic groups to characterize the deepwater invertebrate fauna of Las Gemelas Seamount and Isla del Coco National Park. The Shannon-Weiner Index for biodiversity (H’) was calculated to be 0.14 ± 0.02 for Isla del Coco and 0.07 ± 0.03 for Las Gemelas surveys. Although richness was fairly equal between the two sites, evenness was greater at Isla del Coco (J = 0.04 ± 0.006) when compared to Las Gemelas (J = 0.02 ± 0.01). This lower level of evenness in the community at Las Gemelas was a result of high densities of a few dominant species groups, specifically sea urchins and black corals. We also evaluated invertebrate percent cover at both Isla del Coco and Las Gemelas Seamount with respect to habitat type, slope and rugosity. Results indicated that highly rugose habitats contained the highest frequencies of all invertebrates at both sites, with the exception of glass sponges and polychaetes at Isla del Coco, which were found in greater quantities at intermediate levels of rugosity. Information obtained from these submersible surveys indicate that seamounts in the tropical eastern Pacific Ocean may be an important source of biodiversity and that more quantitative surveys are needed to characterize the fauna of the region.
Key words: Diversity surveys, seamounts, deepwater invertebrates, submersible observations, biodiversity.
Resumen
La fauna de aguas profundas de islas oceánicas y de montes submarinos del Pacífico Tropical Oriental son muy poco conocidas. Para caracterizar las faunas de aguas profundas del Parque Nacional Isla del Coco y el Monte Submarino Las Gemelas, Costa Rica, llevamos a cabo un estudio cuantitativo de los hábitats y su fauna. Se tomaron videos de transectos desde un sumergible entre 50 y 402m de profundidad del 11 al 22 de setiembre 2009. Se recolectó información cuantitativa de la composición de especies de invertebrados, densidad, distribución y hábitats asociados en ambas localidades y se comparó. Se contaron 7,172 invertebrados en los videos analizados, y se ubicaron en 27 categorías taxonómicas. El Índice de Shannon-Weiner (H’) fue de 0.14 ± 0.02 para la Isla del Coco y 0.07 ± 0.03 para Las Gemelas. La riqueza fue parecida en ambos sitios pero la equitabilidad fue mayor en la Isla del Coco (J = 0.04 ± 0.006) comparado con Las Gemelas (J = 0.02 ± 0.01). Este menor nivel de equitabilidad en Las gemelas de debió a la alta densidad de unos pocos grupos dominantes, específicamente erizos de mar y coral negro. También evaluamos el porcentaje de cobertura de los invertebrados con respecto a tipo de hábitat, pendiente y rugosidad del sustrato. Los resultados indican que hábitats con alta rugosidad contiene más invertebrados en ambos sitios, excepto la esponjas silíceas y los poliquetos en la Isla del Coco, que se encontraron en mayor cantidad en niveles intermedios de rugosidad. La información obtenida con este estudio indican que los montes submarinos en el Pacífico Tropical Oriental pueden ser una fuente importante de biodiversidad y que se necesitan más estudios cuantitativos para caracterizar la fauna de la región.
Palabras clave: Encuestas sobre diversidad, montes marinos, invertebrados en aguas profundas, observaciones en sumergible, biodiversidad.
Isla del Coco, also known as Cocos Island, is located 550km southwest of the Pacific coast of Costa Rica and sits atop the Coco Volcanic Cordillera, a submarine mountain range that exists offshore of the southern part of Costa Rica (Cortés 2008, Rojas & Alvarado 2012). The island is near the intersection of the Panama current and the northern equatorial counter-current. This convergence of large current systems creates localized currents that flow up the sides of the undersea ridge from the deep ocean, bringing cooler, nutrient rich water upward where it mixes with warmer surface waters to support an extremely productive ecosystem (Lizano 2008). The deep waters of Isla del Coco were explored in 1986 with the Johnson-Sea-Link I as part of a prospecting survey for the generation of pharmacetical compounds (Cortés 2008). Three papers were published, one on hard corals (Cairns 1991a), one on calcified hydroids of the family Stylasteridae (Cairns 1991b), and recently, Reiswig (2010), described a new species of hexactinellid sponge. In recognition of the large diversity and uniqueness of flora and fauna associated with Isla del Coco, the government of Costa Rica designated it a national park in 1978. No deepwater exploration of the island had taken place until 2006, when the DeepSee submersible started operations (Cortés & Blum 2008). Since then more than 1,500 dives have been logged at Isla del Coco.
In 2007, a conservation gap analysis was conducted, and Las Gemelas Seamount was identified as a possible location for inclusion into the system of marine reserves in Costa Rica (SINAC, MINAET 2008). Reports from Costa Rican fishers, however, indicated that this seamount had been fished only occasion- ally in the last 15 years. In order to determine if the habitats and species of Las Gemelas Seamount were suitable for inclusion in a reserve system, we conducted submersible surveys to compare it with the habitats and fauna of the deeper portions of waters around Isla del Coco National Park. The goal of this project was to characterize the habitats and biota, and conduct quantitative surveys of the deepwater portions of Isla del Coco National Park and Las Gemelas Seamount, located about 50km southwest of Isla del Coco. The objectives of the cruise were to gather information about demersal fishes, evaluate zonation of fishes (Starr et al. 2012) and invertebrates, conduct observational surveys of macroplankton, collect images of habitats and species, and collect octocorals (Octocorallia), black corals (Antipatharia), echinoderms (Echinodermata), crustaceans (Crustacea), other invertebrates, and rocks. Additionally, we gathered quantitative information about species composition, density, distribution and habitat associations and compared the invertebrate communities between the two sites.
Methods
From 11-22 September 2009, scientists from the US National Geographic Society, University of Costa Rica, Moss Landing Marine Laboratories, Monterey Bay Aquarium Research Institute, Ocean Research & Conservation Association, and the University of California conducted an exploration of the deepwater areas around Isla del Coco National Park (5°33’N, 87°02’W) and Las Gemelas Seamount (4°59’N, 87°38’W, Lizano 2012). We used the Undersea Hunter Group’s DeepSee submersible to explore the water column and seafloor habitats to a depth of 400m, described by Cortés & Blum (2008).
At Las Gemelas Seamount we surveyed a general area that was suggested to us by commercial fishermen. When we arrived in the general vicinity of the seamount, we conducted bathymetric surveys of the region using the support vessel’s echosounder to locate the shallowest parts of the seamount, and then used the DeepSee submersible to survey two of the shallow peaks. At Isla del Coco National Park most submersible dives occurred at dive locations along the drop-off at the northern edge of the island in areas normally visited by the DeepSee during its regular trips with commercial passengers. At all dive sites observers usually spent 30-60min exploring the area. After getting a sense of the habitat types associated with the dive site, observers haphazardly chose starting depths and directions for quantitative surveys. Observers most frequently chose to start a transect at a habitat and depth representative of the site (often a rock outcrop). Upon starting a transect, the pilot would slowly (~0.15 m/sec) drive the submersible along a preselected course for 10min. The course was almost always either parallel to an isobath or angled about 45 degrees above or below the isobath. On one transect, the submersible transect was nearly vertical. During a typical 3hr-long dive, we conducted two to four quantitative, 10min transects. The number and starting location of transects were selected by the scientific observer in the submersible.
Submersible transects were patterned after strip transect surveys that have been commonly used to evaluate invertebrates in temperate environments (Yoklavich & O’Connell 2008, Tissot et al. 2007, Starr & Yoklavich 2008). During these quantitative transects, observers looked forward and downward through the submersible dome, for a set time period (usually 10 minutes), and identified and observed invertebrates in a 1m wide swath. Lasers that were mounted 33cm apart, on either side of the cam- era housing, shined parallel beams of light and allowed us to establish transect width. Pilots maneuvered the submersible and/or adjusted the camera so that the camera’s field of view was as close to 1 m wide as possible. Transect lengths were determined by distance traveled as measured by a Doppler velocity log attached to the sub. In addition to direct observations, a video record of the transect swath was recorded by the submersible’s high-definition digital camera on mini-DV tape. All video was subsequently reviewed to verify the identification of taxa, to record invertebrate density and percent cover, and to describe and classify habitat-associations.
In order to compare invertebrate communities at Las Gemelas Seamount and Isla del Coco National Park, we used only data from submersible dives that occurred at similar depths and covered similar habitats at each site. The dives and associated transects at Las Gemelas Seamount covered primarily rocky habitats at depths greater than 150m. Thus, for comparison purposes, we used only the seven submersible dives at Isla del Coco that contained transects covering similar depths and habitats and that could be used to contrast with the four submersible surveys at Las Gemelas Seamount. The comparisons included taxa composition and density of corresponding taxonomic groups.
To evaluate taxa composition, we estimated taxa richness (defined as the number of discrete taxa identified) and diversity (Shannon-Wiener Index, Zar (1999)) at each site – using those macroinvertebrates that we could identify and count individually (generally invertebrates >5cm in size). We then calculated density of each taxonomic group for each dive by summing the number of macroinvertebrates observed on transects and dividing that number by the total transect area of that dive (i.e., transect lengths x 1m width). Invertebrate densities for each site were calculated by dividing the total number of macroinvertebrates by the total area of each site. Although these values are not true estimates of richness or diversity because many of the taxa identified are comprised of several species, we present the results for comparison purposes. Additionally, we conducted multidimensional scaling (MDS) analyses of invertebrate densities at the two study sites using the software program Primer.
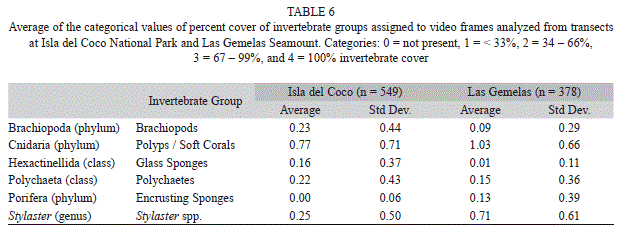
In addition to analyzing densities of individual taxa, the percent cover of six smaller invetebrate groups was estimated. These abundant, smaller taxa generally covered large areas of the seafloor and were comprised of species in the following taxonomic groups: Brachiopoda (phylum), Cnidaria (phylum), Hexactinellida (class), Polychaeta (class), Porifera (phylum), and Stylaster spp. To document percent cover we selected a random starting point within the first minute of the transect and then analyzed individual video frames every 20 seconds of the transect. For each of the still frames analyzed from transects we assigned a percent cover value for each of the six invertebrate taxa evaluated. The percent cover value was a categorical value, i.e., 0 = not present, 1 = < 33%, 2 = 34–66%, 3 = 67–99%, and 4 = 100% invertebrate cover. A percent cover index for each dive site was obtained by averaging the categorical numbers assigned to each video frame analyzed.
Habitats were defined by a combination of substrate type, slope, and rugosity of the seafloor, as described in Greene et al. (1999) and Tissot et al. (2007). We used seven primary substrate codes: boulder (B), cobble (C), gravel (G), pebble (P), bedrock or rock outcrop (R), sand (S), and a code for a vertical pinnacle (T). We defined bottom type as a two-letter code representing the approximate percent cover of the two most prevalent substrata in a particular habitat patch. The first character of the code represents the substratum that accounted for at least 50% of the habitat, and the second represents the second most prevalent habitat, accounting for at least 20% of the patch (e.g., the code RB represents a habitat in which at least 50% is bedrock and at least 20% of the bottom is covered by boulders). If the bottom was a single type of substrate, we used a single code twice (e.g., “BB” for > 70% cover by boulders). We defined three types of slope (<30°, 30 – 60°, >60°) and three rugosity levels (Low, Medium, High) that were arbitrarily defined, based on the capability of the crevices in the substrate to hide fishes (e.g., no room, can hide small fishes, crevices large enough to hide large fishes).
Results
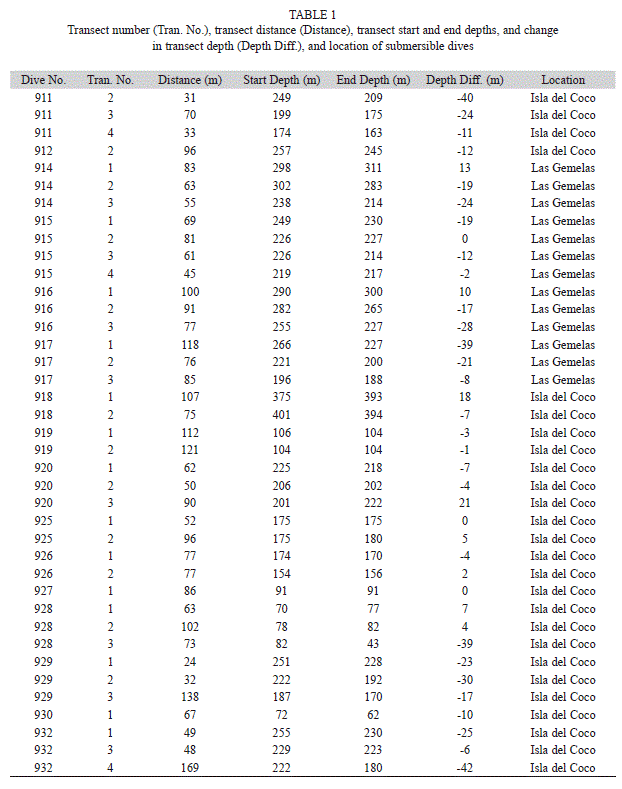
Submersible dives and transects: We completed a total of 22 submersible dives. Rough weather limited our opportunity to launch the submersible at Las Gemelas Seamount, and we were only able to conduct surveys there on two days. We were able to launch the submersible on 12 days at Isla del Coco National Park. We conducted a total of four dives at the Las Gemelas Seamount and 18 dives around Isla del Coco. Maximum depths of dives ranged from 50-402m, and dive duration averaged 3.7hr. Total duration of visual observations during dive explorations was more than 80hr. Quantitative data were available from 16 submersible dives (Table 1). Four of these dives occurred at Las Gemelas Seamount, and 12 dives occurred around Isla del Coco National Park. A total of 38 quantitative transects were completed in this study. Transect lengths varied from 23-169m. The total area surveyed equaled 3003m2. At Isla del Coco National Park, we conducted 25 quantitative transects on 12 dives, and surveyed 1999m2. At Las Gemelas Seamount, we completed 13 video transects on four dives, and surveyed 1004m2. We collected 36hr of video documentation from the 38 transects.
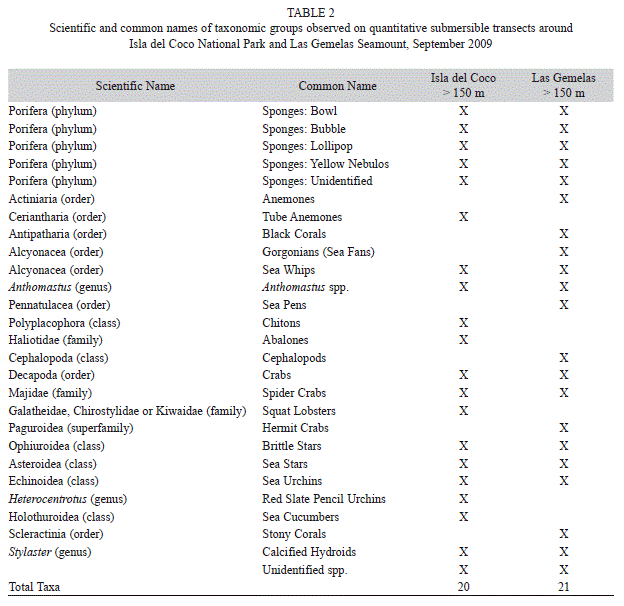
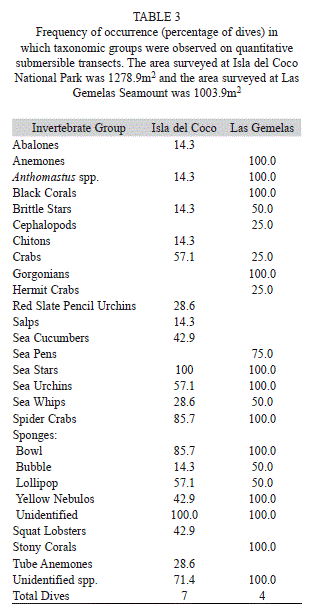
Quantification of Individual Organisms: A total of 7 172 individual invertebrates were counted from analysis of the video collected on this project. These macroinvertebrate species were counted and placed into 27 taxonomic groups to characterize the deepwater invertebrate fauna of Las Gemelas Seamount and Isla del Coco National Park (Table 2). Invertebrate species richness was slightly greater at Las Gemelas Seamount than at Isla del Coco. We encountered 21 taxonomic groups on quantitative transects around Las Gemelas, whereas only 20 taxonomic groups were observed at Isla del Coco. Because we encountered several invertebrates that have not yet been reported in the scientific literature, we grouped species into higher taxonomic levels for our analyses (Table 2). Octocorals (Genus Anthomastus), brittle stars (Class Ophiuroidea), sea urchins (Class Echinoidea), sponges (Phylum Porifera) and unidentified species were common at both locations, although encountered more frequently during Las Gemelas dives. Breedy and Cortés (2008) described some of the octocorals, but more voucher specimens are needed to identify them to species. Anemones (Order Actiniaria), black corals (Order Alcyonacea), gorgonians (Order Alcyonacea) and stony corals (Order Scleractinia) were found on each transect at Las Gemelas, but were absent from Isla del Coco transects (Table 3).
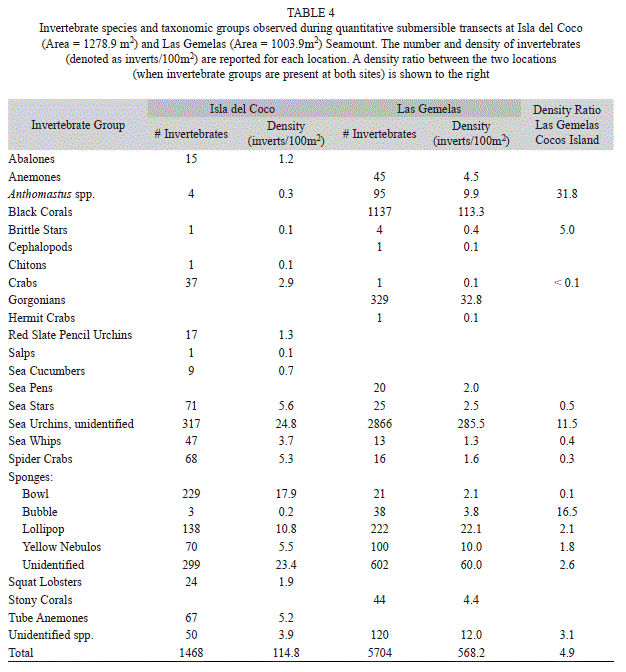
The Shannon-Weiner Index for biodiversity (H’) was calculated to be 0.14 ± 0.02 for Isla del Coco and 0.07 ± 0.03 for Las Gemelas Seamount surveys. Although richness was fairly equal between the two sites, evenness was greater at Isla del Coco (J = 0.04 ± 0.006) when compared to Las Gemelas (J = 0.02 ± 0.01). This lower level of evenness in the community at Las Gemelas was a result of high densities of a few dominant species groups, specifically sea urchins and black corals (Table 4).
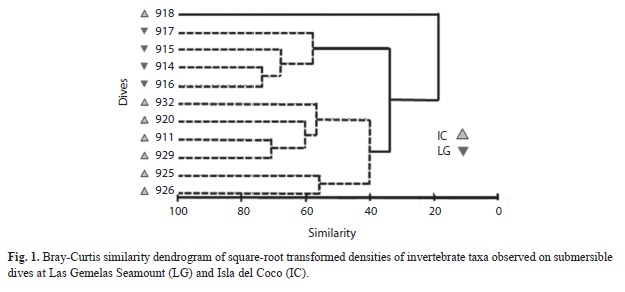
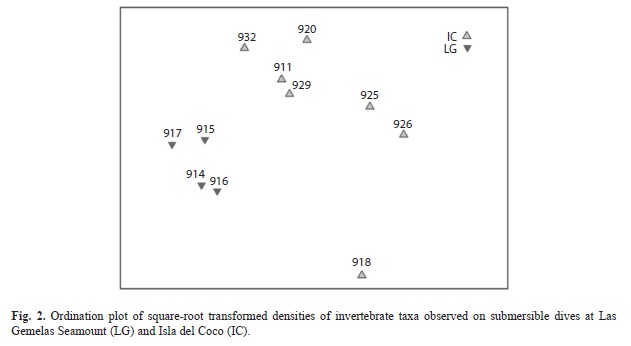
Invertebrate Density: Bray-Curtis similarity indices demonstrated that the invertebrate taxa and densities at Las Gemelas Seamount are different than at Isla del Coco. A cluster analysis of square-root transformed density data (Fig. 1) indicated that occurrences of invertebrate taxa in dives at Las Gemelas Seamount were much more similar than invertebrates at Isla del Coco. Similarly, an ordination plot of the data using multidimensional scaling on the similarity matrix indicated a clear separation between the two sites (Fig. 2). The combined density of all invertebrate groups was 4.9 times higher at Las Gemelas (568 invertebrates/100m2) than at Isla del Coco (115 invertebrates/100m2). This large difference in overall invertebrate density between the two sites can be attributed to the extremely high densities of sea urchins and black corals (and to a smaller extent that of gorgonians) observed on submersible dives at Las Gemelas (Table 4). These invertebrates were distributed throughout all transects, as evidenced by the 100% occurrence on all dives. Also, Anthomastus spp. were present at both sites, but exhibited substantially higher densities around Las Gemelas Seamount than at Isla del Coco.
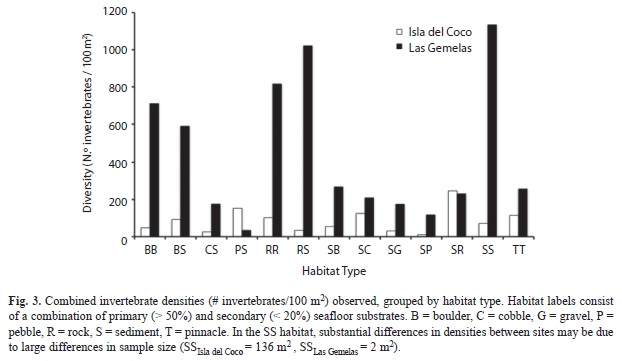
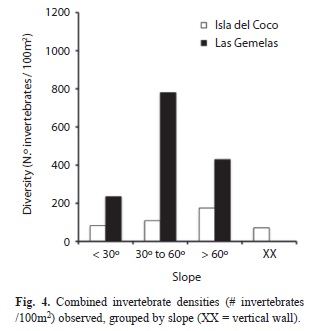
We examined invertebrate densities at both Isla del Coco and Las Gemelas Seamount with respect to habitat type, slope, and rugosity. Whereas invertebrate densities were highest in sand, rock, and boulder habitats at Las Gemelas, invertebrates at Isla del Coco were denser in pinnacle, rock, mixed areas of sand and rock, pebble, and boulder habitats (Fig. 3). When comparing density with slope, we found that invertebrate densities at Isla del Coco increased with increasing slope, whereas invertebrate densities at Las Gemelas were highest in moderate slopes of 30° to 60° (Fig. 4). A comparison of invertebrate densities by rugosity category indicated that medium to high rugosity habitats contained the highest invertebrate densities at both sites (Fig. 5).
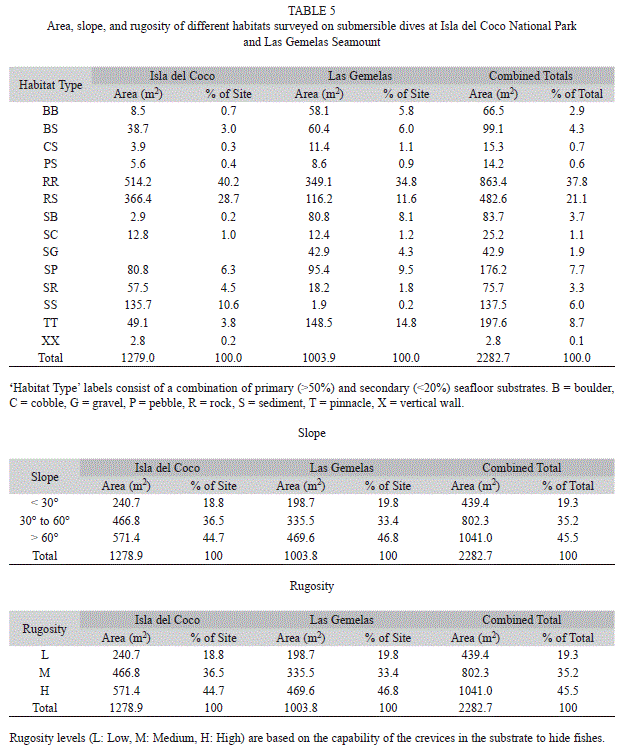
Habitat types: We encountered a total of 14 combinations of the seven substratum codes. At Isla del Coco, 58% of the habitats surveyed were comprised primarily of rock and 34% were primarily sand, whereas at Las Gemelas, 73% of the transects covered rocky habitats and 25% covered sandy habitats (Table 5). Transects at Las Gemelas Seamount occurred over somewhat more rugose habitats. At Isla del Coco, 39.9%, 28.5%, and 31.6% of the habitat area surveyed was high, medium, and low rugosity, respectively. At Las Gemelas Seamount, 46.8%, 34.3%, and 18.9% of the habitat area surveyed was high, medium, and low rugosity, respectively. Similarly, transects at Las Gemelas more typically occurred on higher slopes. At Las Gemelas Seamount, 30%, 56%, and 14% of slops in the hábitat area surveyed were <30°, 30–60°, and >60°, respectively. At Isla del Coco, 48%, 35%, and 17% of the slopes in the habitat area surveyed were <30°, 30–60°, and >60°, respectively.
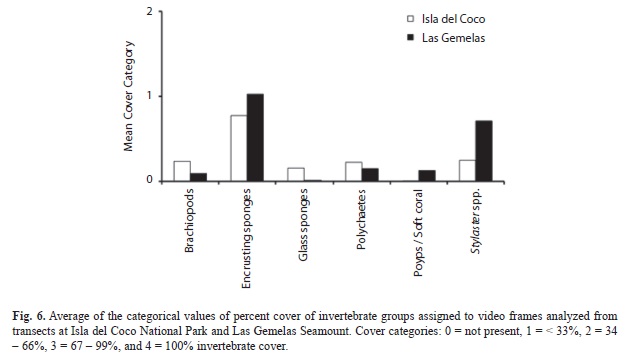
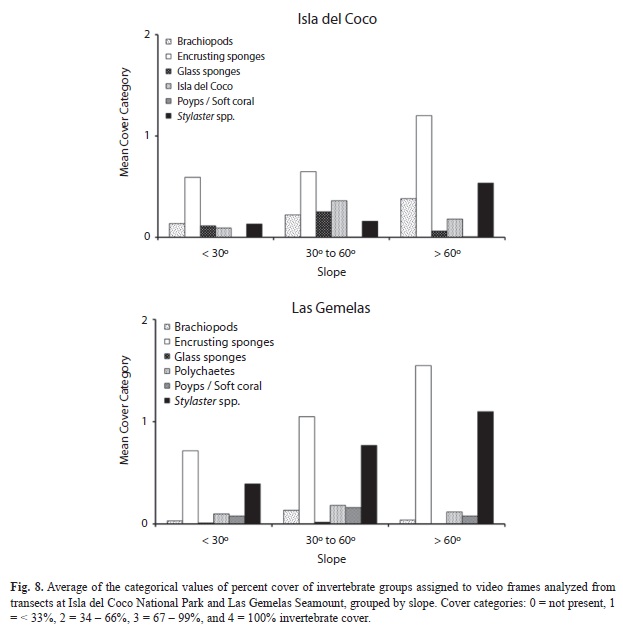
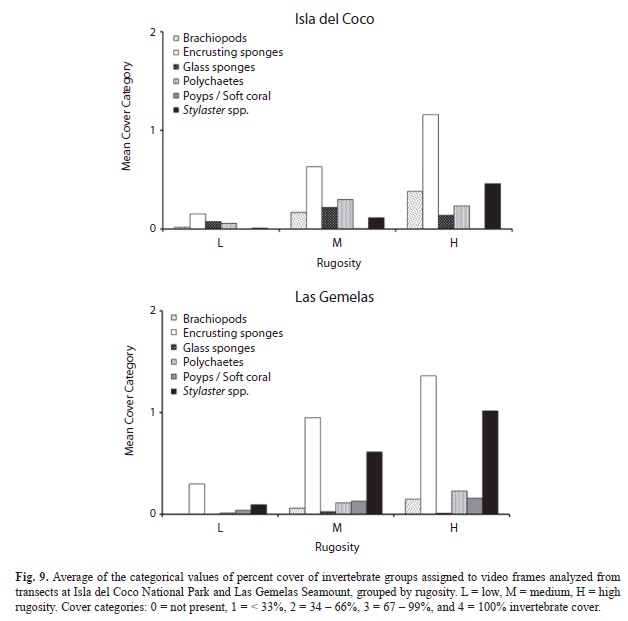
Quantification of Percent Cover: All six taxonomic groups used in the quantification of percent invertebrate cover were present at both Isla del Coco and Las Gemelas Seamount (Table 6). Brachiopods, glass sponges and polychaetes were more prevalent at Isla del Coco, whereas encrusting sponges, polyps/soft corals and Stylaster spp. were found in higher frequencies at Las Gemelas (Fig. 6). Encrusting sponges were the most commonly encountered taxonomic group at both sites. We evaluated invertebrate percent cover at both Isla del Coco and Las Gemelas Seamount with respect to habitat type, slope and rugosity. Whereas encrusting sponges and Stylaster spp. dominated every habitat type at Las Gemelas, percent cover of encrusting sponges and Stylaster spp. at Isla del Coco were greater in rock, pinnacle, and mixed areas of sand and rock or pebble habitats only (Fig. 7). Glass sponges dominated habitats consisting primarily of boulders at Isla del Coco. When evaluating percent cover with slope, we found that all invertebrate groups were more prevalent in areas with intermediate (30° to 60°) or steep (> 60°) slopes (Fig. 8). A comparison of percent cover and rugosity indicated that high rugosity habitats contained the highest frequencies of all invertebrates at both sites, with the exception of glass sponges and polychaetes at Isla del Coco, which were found in greater quantities at intermediate levels of rugosity (g. 9).
Discussion
In September 2009, we surveyed the habitats and invertebrate communities in the deep- water portions of Las Gemelas Seamount and Isla del Coco. We used a combination of quantitative strip transects and general observations to characterize invertebrate communities at each study site. We surveyed habitats in depths ranging from 50-402m, but reported only the data obtained from dives in rocky habitats that were deeper than 150m to facilitate a comparison of deepwater communities at Las Gemelas Seamount and Isla del Coco. Habitats surveyed at Isla del Coco included vertical rock walls and steep slopes consisting of highly fragmented areas of volcanic rock outcrops and sand. Sea urchins and sponges were among the most commonly observed taxonomic groups. Higher densities of these organisms were found at slopes greater than 60° and in areas displaying high levels of rugosity. Pinnacles and rocky habitats housed the highest overall densities of invertebrates, although the highest density of an invertebrate taxon at Isla del Coco was still much lower than from Las Gemelas Seamount.
Seamounts are important features in the world’s oceans, and an increasing amount of research has been devoted to studying the biological communities and patterns of benthic biodiversity of seamounts. Not surprisingly, with increased information more questions are generated about the ecological role of seamounts and their vulnerability to disturbance. Many researchers have suggested that seamounts are unique environments, containing hotspots of biodiversity and endemism (Clark et al. 2010, Rowden et al. 2010b), and are very fragile (Koslow et al. 2001). Although Samadi et al. (2006) and Thoma et al. (2009) indicated that genetic studies might refute the concept of increased endemism on seamounts, they did agree with the hypothesis that seamounts are diversity hotspots, possessing benthic assemblages with particularly high species richness.
Habitats surveyed at Las Gemelas contained rich, dense assemblages of invertebrates. Some of the prominent benthic invertebrates included sea urchins, black corals, sponges, anemones, corals, gorgonians, echinoderms, and crustaceans. Two taxonomic groups (sea urchins and black corals) accounted for more than 70% of the total combined invertebrate density at Las Gemelas. These very prevalent taxa were found at higher frequencies in areas of rock and/or sand, intermediate slopes (30° to 60°), and medium levels of rugosity. These differences in assemblages of deepwater invertebrates is similar to those shown by Ricciardi and Bourget (1999) and Adami et al. (2004) for intertidal invertebrates. They showed that significant differences in community structure were found between vertical and horizontal samples in different locations. Also, differences in structure were much greater between localities than among replicates within each locality. Rowden et al. (2010a) analyzed more than 5000 sea-floor images on 20 southwest Pacific seamounts and reported that the mean biomass of epibenthic megafauna was nearly four times greater than on the adjacent continental slope at comparable depths.
The difference in invertebrate densities between the two sites may also be attributed to habitat heterogeneity. At Isla del Coco, 74% of habitats were either all rock or some mixture of rock and sand. Habitats at Las Gemelas, however, were more diverse, and consisted pri- marily of rock, sand, and/or pinnacles. Another potential explanation for the difference in invertebrate densities is that the habitats surveyed at Las Gemelas covered a smaller range of slope categories (most slopes were 30º to 60º) than present at Isla del Coco, which was comprised primarily of more extreme slopes (typically <30º or >60º). Mendoza et al. (2010) showed that deep, steep underwater slopes as prime habitats for many decapod crustaceans and suggested that these habitats are under- studied, thus may contain many new species. Until more surveys are conducted to enable an analysis of species-area curves, we cannot fully explain the observed differences. Importantly, we encountered different habitats in each of our dives at Las Gemelas, indicating that the sea- mounts are likely to include a wider variety of habitats than those surveyed. This suggests that the diversity of taxa at Las Gemelas Seamount is potentially much greater than we were able to determine with the available submersible dives.
One important qualitative observation is that we saw a larger number of encrusting and structure-forming invertebrates at Las Gemelas Seamount than at Isla del Coco. This habitat complexity further indicates that habitats at Las Gemelas may be able to harbor a greater diversity and biomass of both invertebrate and fish species than at Isla del Coco. O’Hara et al. (2008) reported that the large biogenic structures formed by colonial cold-water scleractinian corals provide valuable habitat for marine invertebrates on seamounts. They suggested that cold-water scleractinian corals are a key habitat for a many ophiuroid species that occur in the Western Pacific Ocean.
Given the diverse and rich assemblages of invertebrates carpeting the bottom habitats at Las Gemelas Seamount, as well as the fact that the area contains numerous species which have not yet been described in the scientific literature (and thus, are important for the maintenance of biodiversity), Las Gemelas Seamount may be a richer environment for invertebrates than the deep water surrounding Isla del Coco. Koslow et al. (2001) discussed concerns about the vulnerability of seamount communities to human impacts, especially with the development of large-scale bottom trawl fisheries in the deep sea. They reported that 24 to 43% of the invertebrate species they recovered from experimental trawling were new to science, and between 16 and 33% appeared to be restricted to the seamount environment. The concern expressed by Koslow et al. (2001) is that seamount invertebrate communities are fragile. O’Hara et al. (2008) believe that the slow growth rate of seamount corals that recovery of damaged cold-water coral assemblages will be a long-term process. The high densities of invertebrates in the area around Isla del Coco National Park and Las Gemelas seamount signifies the need for increased protection and study of these areas, because of the paucity of information related to the distribution and relative abundance of the important resources that could easily be overexploited.
Acknowledgments
We thank Shmulik Blum, Sylvia Earle, Avi Klapfer, Bruce Robison, Edith Widder, and the crew of the Argo for safe submersible operations and help with observations of fishes and macroinvertebrates. Katie Schmidt helped excerpt information from video and Dan Malone provided the MDS plots. Fund- ing was provided by the National Geographic Society, California Sea Grant Program, Moss Landing Marine Labs, and the Universidad de Costa Rica.
References
Adami M., A. Tablado & J. Lopez-Gappa. 2004. Spatial and temporal variability in intertidal assemblages dominated by the mussel Brachidontes rodriguezii (d’orbigny, 1846). Hydrobiologia 520: 49-59. [ Links ]
Bray J.R. & J.T. Curtis. 1957. An ordination of the upland forest communities of Southern Wisconsin. Ecol. Monogr. 27: 325-349. [ Links ]
Breedy, O. & J. Cortés. 2008. Octocorals (Coelenterata: Anthozoa: Octocorallia) of Isla del Coco, Costa Rica. Rev. Biol. Trop. 56 (Suppl. 2): 71-77. [ Links ]
Cairns, S.D. 1991a. A revision of the ahermatypic Scleractinia of the Galápagos and Cocos Islands. Smithsonian Contr. Zool. 504: 1-33. [ Links ]
Cairns, S.D. 1991b. New records of Stylasteridae (Hydrozoa: Hydroida) from the Galápagos and Cocos Islands. Proc. Biol. Soc. Washington 104: 209-228. [ Links ]
Clark, M.R., A. A. Rowden, T. Schlacher, A. Williams, M. Consalvey, K.I. Stocks, A.D. Rogers, T.D. O’Hara, M. White, T.M. Shank & J.M. Hall-Spencer. 2010. The ecology of seamounts: Structure, function, and Human impacts. Ann. Rev. Mar. Sci. 2: 253-278. [ Links ]
Cortés, J. 2008. Historia de la investigación marina de la Isla del Coco, Costa Rica. Rev. Biol. Trop. 56 (Suppl. 2): 1-18. [ Links ]
Cortés, J. & S. Blum. 2008. Life to 450 m depth at Isla del Coco, Costa Rica. Rev. Biol. Trop. 56 (Suppl. 2): 189-206. [ Links ]
Greene, H.G., M.M. Yoklavich, R.M. Starr, V.M. O’Connell, W.W.Wakefield, D.E. Sullivan, J.E. McRea & G.M. Cailliet. 1999. A classification scheme for deep sea- floor habitats. Oceanol. Acta 22: 663-678. [ Links ]
Koslow J.A., G.W. Boehlert, J.D.M. Gordon, R.L. Haedrich, P. Lorance & N. Parin. 2000. Continental slope and deep-sea fisheries: Implications for a fragile ecosystem. ICES J. Mar. Sci. 57: 548-557. [ Links ]
Koslow J.A., K. Gowlett-Holmes, J.K. Lowry, T. O’Hara, G. Poore & A. Williams. 2001. Seamount benthic macrofauna off southern Tasmania: Community structure and impacts of trawling. Mar. Ecol. Prog. Ser. 213: 111-125. [ Links ]
Lizano, O.G. 2008. Dinámica de aguas alrededor de la Isla del Coco, Costa Rica. Rev. Biol. Trop. 56 (Suppl. 2): 31-48. [ Links ]
Lizano, O.G. 2012. Rasgos morfológicos alrededor de la Isla del Coco y de sus montes submarinos vecinos, Pacífico de Costa Rica. Rev. Biol. Trop. 60 (Suppl.
3): 43-51. [ Links ]
O’Hara T.D., A.A. Rowden & A. Williams. 2008. Cold- water coral habitats on seamounts: Do they have a specialist fauna? Biodivers. Lett. 14: 925-934. [ Links ]
Mendoza, J.C.E., T. Naruse, S. Tan, T. Chan, D.F. Richer & P.K.L. Ng. 2010. Case studies on decapod crustaceans from the Philippines reveal deep, steep underwater slopes as prime habitats for ‘rare’ species. Biodivers. Conserv. 19: 575-586. [ Links ]
Reiswig, H.M. 2010. A new species of Tretodictyum (Porifera: Hexactinellida: Tretodictyidae) from off Cocos Island, tropical eastern Pacific Ocean. Proc. Biol. Soc. Washington 123: 242-250. [ Links ]
Ricciardi, A. & E. Bourget. 1999. Global patterns of macroinvertebrate biomass in marine intertidal communities. Mar. Ecol. Prog. Ser. 185: 21-35. [ Links ]
Richer de Forges, B., J.A. Koslow & G.C.B. Poore. 2000. Diversity and endemism of the benthic seamount fauna in the southwest Pacific. Nature 405: 944-947. [ Links ]
Rojas, W. & G.E. Alvarado. 2012. Geología y contexto geotectónico de la Isla del Coco y la zona marítima frente al Pacífico Central de Costa Rica. Rev. Biol. Trop. 60 (Suppl. 3): 15-32. [ Links ]
Rowden, A.A., T.A. Schlacher, A. Williams, M.R. Clark, R. Stewart, F. Althaus, D.A. Bowden, M. Consalvey, W. Robinson & J. Dowdney. 2010a. A test of the seamount oasis hypothesis: Seamounts support higher epibenthic megafaunal biomass than adjacent slopes. Mar. Ecol. 31: 95-106. [ Links ]
Rowden, A.A., J.F. Dower, T. A. Schlacher, M. Consalvey & M.R. Clark. 2010b. Paradigms in seamount ecology: fact, fiction and future. Mar. Ecol. (Suppl.) 131: 226-241. [ Links ]
Samadi, S., L. Bottan, E. Macpherson, B. Richer De Forges & M. Boisselier. 2006. Seamount endemism questioned by the geographic distribution and population genetic structure of marine invertebrates. Mar. Biol. 149: 1463-1475. [ Links ]
SINAC, MINAET. 2008. GRUAS II: Propuesta de Ordenamiento Territorial para la conservación de la biodiversidadde Costa Rica. Volumen 3: Análisis de Vacíos en la Representatividad e Integridad de la biodiversidad marina y costera. Sistema Nacional de Áreas de Conservación (SINAC), Ministerio de Ambiente, Energía y Telecomunicaciones (MINAET), San José, Costa Rica. 60 p. [ Links ]
Starr, R.M. & M.M. Yoklavich. 2008. Monitoring MPAs in Deep Water off Central California: 2007 IMPACT Submersible Baseline Survey. California Sea Grant College Program Publication T-067. 24 p. [ Links ]
Starr, R.M., K. Green & E. Sala. 2012. Deepwater fish assemblages at Isla del Coco National Park and Las Gemelas Seamounts, Costa Rica. Rev. Biol. Trop. 60 (Suppl. 3): 347-362. [ Links ]
Thoma, J.N., E. Pante, M.R. Brugler & S.C. France. 2009. Deep-sea octocorals and antipatharians show no evidence of seamount-scale endemism in the NW Atlantic. Mar. Ecol. Prog. Ser. 397: 25-35. [ Links ]
Tissot, B.N., M.A. Hixon & D.L. Stein. 2007. Habitat- based submersible assessment of macro-invertebrate and groundfish assemblages at Heceta Bank, Oregon, from 1988 to 1990. J. Exp. Mar. Biol. Ecol. 352: 50-64. [ Links ]
Yoklavich, M.M. & V. O’Connell. 2008. Twenty Years of Research on Demersal Communities Using the Delta Submersible in the Northeast Pacific. In J.R. Reynolds & H.G. Greene (eds.). Marine Habitat Mapping Technology for Alaska. Alaska Sea Grant College Program, University of Alaska Fairbanks. doi:10.4027/mhmta.2008.10. [ Links ]
Zar, H. 1999. Biostatistical Analysis. 4th Ed. Prentice Hall, Upper Saddle River, New Jersey. [ Links ]
*Correspondencia a:
Richard M. Starr. University of California Sea Grant Program and Moss Landing Marine Laboratories, 8272 Moss Landing Road, Moss Landing, California 95039, USA; starr@mlml.calstate.edu
Jorge Cortés. Centro de Investigación en Ciencias del Mar y Limnología (CIMAR), Ciudad de la Investigación, Universidad de Costa Rica, San Pedro, 11501-2060 San José, Costa Rica; jorge.cortes@ucr.ac.cr
Cheryl L. Barnes. University of California Sea Grant Program and Moss Landing Marine Laboratories, 8272 Moss Landing Road, Moss Landing, California 95039, USA.
Kristen Green. Alaska Department of Fish and Game, 304 Lake Street, Room 103, Sitka, Alaska, 99835; kristen.green@alaska.gov
Odalisca Breedy. Centro de Investigación en Ciencias del Mar y Limnología (CIMAR), Ciudad de la Investigación, Universidad de Costa Rica, San Pedro, 11501-2060 San José, Costa Rica
1. University of California Sea Grant Program and Moss Landing Marine Laboratories, 8272 Moss Landing Road, Moss Landing, California 95039, USA; starr@mlml.calstate.edu
2. Centro de Investigación en Ciencias del Mar y Limnología (CIMAR), Ciudad de la Investigación, Universidad de Costa Rica, San Pedro, 11501-2060 San José, Costa Rica; jorge.cortes@ucr.ac.cr
3. Alaska Department of Fish and Game, 304 Lake Street, Room 103, Sitka, Alaska, 99835; kristen.green@alaska.gov
*Dirección para correspondencia:
Abstract
The deepwater faunas of oceanic islands and seamounts of the Eastern Tropical Pacific are poorly known. From 11-22 September 2009, we conducted an exploration of the deepwater areas around Isla del Coco National Park and Las Gemelas Seamount, located about 50km southwest of Isla del Coco, Costa Rica using a manned submersible to survey the seafloor habitats. The goal of the exploration was to characterize the habitats and biota, and conduct quantitative surveys of the deepwater portions of Isla del Coco National Park and Las Gemelas. We completed a total of 22 successful submersible dives, spanning more than 80hr underwater, and collected a total of 36hr of video. With respect to invertebrates, our objectives were to gather quantitative information on species composition, density, distribution and habitat associations as well as to compare the invertebrate communities between the two sites. A total of 7 172 invertebrates were counted from analysis of the video collected on this project. Larger organisms were counted and placed into 27 taxonomic groups to characterize the deepwater invertebrate fauna of Las Gemelas Seamount and Isla del Coco National Park. The Shannon-Weiner Index for biodiversity (H’) was calculated to be 0.14 ± 0.02 for Isla del Coco and 0.07 ± 0.03 for Las Gemelas surveys. Although richness was fairly equal between the two sites, evenness was greater at Isla del Coco (J = 0.04 ± 0.006) when compared to Las Gemelas (J = 0.02 ± 0.01). This lower level of evenness in the community at Las Gemelas was a result of high densities of a few dominant species groups, specifically sea urchins and black corals. We also evaluated invertebrate percent cover at both Isla del Coco and Las Gemelas Seamount with respect to habitat type, slope and rugosity. Results indicated that highly rugose habitats contained the highest frequencies of all invertebrates at both sites, with the exception of glass sponges and polychaetes at Isla del Coco, which were found in greater quantities at intermediate levels of rugosity. Information obtained from these submersible surveys indicate that seamounts in the tropical eastern Pacific Ocean may be an important source of biodiversity and that more quantitative surveys are needed to characterize the fauna of the region.
Key words: Diversity surveys, seamounts, deepwater invertebrates, submersible observations, biodiversity.
Resumen
La fauna de aguas profundas de islas oceánicas y de montes submarinos del Pacífico Tropical Oriental son muy poco conocidas. Para caracterizar las faunas de aguas profundas del Parque Nacional Isla del Coco y el Monte Submarino Las Gemelas, Costa Rica, llevamos a cabo un estudio cuantitativo de los hábitats y su fauna. Se tomaron videos de transectos desde un sumergible entre 50 y 402m de profundidad del 11 al 22 de setiembre 2009. Se recolectó información cuantitativa de la composición de especies de invertebrados, densidad, distribución y hábitats asociados en ambas localidades y se comparó. Se contaron 7,172 invertebrados en los videos analizados, y se ubicaron en 27 categorías taxonómicas. El Índice de Shannon-Weiner (H’) fue de 0.14 ± 0.02 para la Isla del Coco y 0.07 ± 0.03 para Las Gemelas. La riqueza fue parecida en ambos sitios pero la equitabilidad fue mayor en la Isla del Coco (J = 0.04 ± 0.006) comparado con Las Gemelas (J = 0.02 ± 0.01). Este menor nivel de equitabilidad en Las gemelas de debió a la alta densidad de unos pocos grupos dominantes, específicamente erizos de mar y coral negro. También evaluamos el porcentaje de cobertura de los invertebrados con respecto a tipo de hábitat, pendiente y rugosidad del sustrato. Los resultados indican que hábitats con alta rugosidad contiene más invertebrados en ambos sitios, excepto la esponjas silíceas y los poliquetos en la Isla del Coco, que se encontraron en mayor cantidad en niveles intermedios de rugosidad. La información obtenida con este estudio indican que los montes submarinos en el Pacífico Tropical Oriental pueden ser una fuente importante de biodiversidad y que se necesitan más estudios cuantitativos para caracterizar la fauna de la región.
Palabras clave: Encuestas sobre diversidad, montes marinos, invertebrados en aguas profundas, observaciones en sumergible, biodiversidad.
Isla del Coco, also known as Cocos Island, is located 550km southwest of the Pacific coast of Costa Rica and sits atop the Coco Volcanic Cordillera, a submarine mountain range that exists offshore of the southern part of Costa Rica (Cortés 2008, Rojas & Alvarado 2012). The island is near the intersection of the Panama current and the northern equatorial counter-current. This convergence of large current systems creates localized currents that flow up the sides of the undersea ridge from the deep ocean, bringing cooler, nutrient rich water upward where it mixes with warmer surface waters to support an extremely productive ecosystem (Lizano 2008). The deep waters of Isla del Coco were explored in 1986 with the Johnson-Sea-Link I as part of a prospecting survey for the generation of pharmacetical compounds (Cortés 2008). Three papers were published, one on hard corals (Cairns 1991a), one on calcified hydroids of the family Stylasteridae (Cairns 1991b), and recently, Reiswig (2010), described a new species of hexactinellid sponge. In recognition of the large diversity and uniqueness of flora and fauna associated with Isla del Coco, the government of Costa Rica designated it a national park in 1978. No deepwater exploration of the island had taken place until 2006, when the DeepSee submersible started operations (Cortés & Blum 2008). Since then more than 1,500 dives have been logged at Isla del Coco.
In 2007, a conservation gap analysis was conducted, and Las Gemelas Seamount was identified as a possible location for inclusion into the system of marine reserves in Costa Rica (SINAC, MINAET 2008). Reports from Costa Rican fishers, however, indicated that this seamount had been fished only occasion- ally in the last 15 years. In order to determine if the habitats and species of Las Gemelas Seamount were suitable for inclusion in a reserve system, we conducted submersible surveys to compare it with the habitats and fauna of the deeper portions of waters around Isla del Coco National Park. The goal of this project was to characterize the habitats and biota, and conduct quantitative surveys of the deepwater portions of Isla del Coco National Park and Las Gemelas Seamount, located about 50km southwest of Isla del Coco. The objectives of the cruise were to gather information about demersal fishes, evaluate zonation of fishes (Starr et al. 2012) and invertebrates, conduct observational surveys of macroplankton, collect images of habitats and species, and collect octocorals (Octocorallia), black corals (Antipatharia), echinoderms (Echinodermata), crustaceans (Crustacea), other invertebrates, and rocks. Additionally, we gathered quantitative information about species composition, density, distribution and habitat associations and compared the invertebrate communities between the two sites.
Methods
From 11-22 September 2009, scientists from the US National Geographic Society, University of Costa Rica, Moss Landing Marine Laboratories, Monterey Bay Aquarium Research Institute, Ocean Research & Conservation Association, and the University of California conducted an exploration of the deepwater areas around Isla del Coco National Park (5°33’N, 87°02’W) and Las Gemelas Seamount (4°59’N, 87°38’W, Lizano 2012). We used the Undersea Hunter Group’s DeepSee submersible to explore the water column and seafloor habitats to a depth of 400m, described by Cortés & Blum (2008).
At Las Gemelas Seamount we surveyed a general area that was suggested to us by commercial fishermen. When we arrived in the general vicinity of the seamount, we conducted bathymetric surveys of the region using the support vessel’s echosounder to locate the shallowest parts of the seamount, and then used the DeepSee submersible to survey two of the shallow peaks. At Isla del Coco National Park most submersible dives occurred at dive locations along the drop-off at the northern edge of the island in areas normally visited by the DeepSee during its regular trips with commercial passengers. At all dive sites observers usually spent 30-60min exploring the area. After getting a sense of the habitat types associated with the dive site, observers haphazardly chose starting depths and directions for quantitative surveys. Observers most frequently chose to start a transect at a habitat and depth representative of the site (often a rock outcrop). Upon starting a transect, the pilot would slowly (~0.15 m/sec) drive the submersible along a preselected course for 10min. The course was almost always either parallel to an isobath or angled about 45 degrees above or below the isobath. On one transect, the submersible transect was nearly vertical. During a typical 3hr-long dive, we conducted two to four quantitative, 10min transects. The number and starting location of transects were selected by the scientific observer in the submersible.
Submersible transects were patterned after strip transect surveys that have been commonly used to evaluate invertebrates in temperate environments (Yoklavich & O’Connell 2008, Tissot et al. 2007, Starr & Yoklavich 2008). During these quantitative transects, observers looked forward and downward through the submersible dome, for a set time period (usually 10 minutes), and identified and observed invertebrates in a 1m wide swath. Lasers that were mounted 33cm apart, on either side of the cam- era housing, shined parallel beams of light and allowed us to establish transect width. Pilots maneuvered the submersible and/or adjusted the camera so that the camera’s field of view was as close to 1 m wide as possible. Transect lengths were determined by distance traveled as measured by a Doppler velocity log attached to the sub. In addition to direct observations, a video record of the transect swath was recorded by the submersible’s high-definition digital camera on mini-DV tape. All video was subsequently reviewed to verify the identification of taxa, to record invertebrate density and percent cover, and to describe and classify habitat-associations.
In order to compare invertebrate communities at Las Gemelas Seamount and Isla del Coco National Park, we used only data from submersible dives that occurred at similar depths and covered similar habitats at each site. The dives and associated transects at Las Gemelas Seamount covered primarily rocky habitats at depths greater than 150m. Thus, for comparison purposes, we used only the seven submersible dives at Isla del Coco that contained transects covering similar depths and habitats and that could be used to contrast with the four submersible surveys at Las Gemelas Seamount. The comparisons included taxa composition and density of corresponding taxonomic groups.
To evaluate taxa composition, we estimated taxa richness (defined as the number of discrete taxa identified) and diversity (Shannon-Wiener Index, Zar (1999)) at each site – using those macroinvertebrates that we could identify and count individually (generally invertebrates >5cm in size). We then calculated density of each taxonomic group for each dive by summing the number of macroinvertebrates observed on transects and dividing that number by the total transect area of that dive (i.e., transect lengths x 1m width). Invertebrate densities for each site were calculated by dividing the total number of macroinvertebrates by the total area of each site. Although these values are not true estimates of richness or diversity because many of the taxa identified are comprised of several species, we present the results for comparison purposes. Additionally, we conducted multidimensional scaling (MDS) analyses of invertebrate densities at the two study sites using the software program Primer.
In addition to analyzing densities of individual taxa, the percent cover of six smaller invetebrate groups was estimated. These abundant, smaller taxa generally covered large areas of the seafloor and were comprised of species in the following taxonomic groups: Brachiopoda (phylum), Cnidaria (phylum), Hexactinellida (class), Polychaeta (class), Porifera (phylum), and Stylaster spp. To document percent cover we selected a random starting point within the first minute of the transect and then analyzed individual video frames every 20 seconds of the transect. For each of the still frames analyzed from transects we assigned a percent cover value for each of the six invertebrate taxa evaluated. The percent cover value was a categorical value, i.e., 0 = not present, 1 = < 33%, 2 = 34–66%, 3 = 67–99%, and 4 = 100% invertebrate cover. A percent cover index for each dive site was obtained by averaging the categorical numbers assigned to each video frame analyzed.
Habitats were defined by a combination of substrate type, slope, and rugosity of the seafloor, as described in Greene et al. (1999) and Tissot et al. (2007). We used seven primary substrate codes: boulder (B), cobble (C), gravel (G), pebble (P), bedrock or rock outcrop (R), sand (S), and a code for a vertical pinnacle (T). We defined bottom type as a two-letter code representing the approximate percent cover of the two most prevalent substrata in a particular habitat patch. The first character of the code represents the substratum that accounted for at least 50% of the habitat, and the second represents the second most prevalent habitat, accounting for at least 20% of the patch (e.g., the code RB represents a habitat in which at least 50% is bedrock and at least 20% of the bottom is covered by boulders). If the bottom was a single type of substrate, we used a single code twice (e.g., “BB” for > 70% cover by boulders). We defined three types of slope (<30°, 30 – 60°, >60°) and three rugosity levels (Low, Medium, High) that were arbitrarily defined, based on the capability of the crevices in the substrate to hide fishes (e.g., no room, can hide small fishes, crevices large enough to hide large fishes).
Results
Submersible dives and transects: We completed a total of 22 submersible dives. Rough weather limited our opportunity to launch the submersible at Las Gemelas Seamount, and we were only able to conduct surveys there on two days. We were able to launch the submersible on 12 days at Isla del Coco National Park. We conducted a total of four dives at the Las Gemelas Seamount and 18 dives around Isla del Coco. Maximum depths of dives ranged from 50-402m, and dive duration averaged 3.7hr. Total duration of visual observations during dive explorations was more than 80hr. Quantitative data were available from 16 submersible dives (Table 1). Four of these dives occurred at Las Gemelas Seamount, and 12 dives occurred around Isla del Coco National Park. A total of 38 quantitative transects were completed in this study. Transect lengths varied from 23-169m. The total area surveyed equaled 3003m2. At Isla del Coco National Park, we conducted 25 quantitative transects on 12 dives, and surveyed 1999m2. At Las Gemelas Seamount, we completed 13 video transects on four dives, and surveyed 1004m2. We collected 36hr of video documentation from the 38 transects.
Quantification of Individual Organisms: A total of 7 172 individual invertebrates were counted from analysis of the video collected on this project. These macroinvertebrate species were counted and placed into 27 taxonomic groups to characterize the deepwater invertebrate fauna of Las Gemelas Seamount and Isla del Coco National Park (Table 2). Invertebrate species richness was slightly greater at Las Gemelas Seamount than at Isla del Coco. We encountered 21 taxonomic groups on quantitative transects around Las Gemelas, whereas only 20 taxonomic groups were observed at Isla del Coco. Because we encountered several invertebrates that have not yet been reported in the scientific literature, we grouped species into higher taxonomic levels for our analyses (Table 2). Octocorals (Genus Anthomastus), brittle stars (Class Ophiuroidea), sea urchins (Class Echinoidea), sponges (Phylum Porifera) and unidentified species were common at both locations, although encountered more frequently during Las Gemelas dives. Breedy and Cortés (2008) described some of the octocorals, but more voucher specimens are needed to identify them to species. Anemones (Order Actiniaria), black corals (Order Alcyonacea), gorgonians (Order Alcyonacea) and stony corals (Order Scleractinia) were found on each transect at Las Gemelas, but were absent from Isla del Coco transects (Table 3).
The Shannon-Weiner Index for biodiversity (H’) was calculated to be 0.14 ± 0.02 for Isla del Coco and 0.07 ± 0.03 for Las Gemelas Seamount surveys. Although richness was fairly equal between the two sites, evenness was greater at Isla del Coco (J = 0.04 ± 0.006) when compared to Las Gemelas (J = 0.02 ± 0.01). This lower level of evenness in the community at Las Gemelas was a result of high densities of a few dominant species groups, specifically sea urchins and black corals (Table 4).
Invertebrate Density: Bray-Curtis similarity indices demonstrated that the invertebrate taxa and densities at Las Gemelas Seamount are different than at Isla del Coco. A cluster analysis of square-root transformed density data (Fig. 1) indicated that occurrences of invertebrate taxa in dives at Las Gemelas Seamount were much more similar than invertebrates at Isla del Coco. Similarly, an ordination plot of the data using multidimensional scaling on the similarity matrix indicated a clear separation between the two sites (Fig. 2). The combined density of all invertebrate groups was 4.9 times higher at Las Gemelas (568 invertebrates/100m2) than at Isla del Coco (115 invertebrates/100m2). This large difference in overall invertebrate density between the two sites can be attributed to the extremely high densities of sea urchins and black corals (and to a smaller extent that of gorgonians) observed on submersible dives at Las Gemelas (Table 4). These invertebrates were distributed throughout all transects, as evidenced by the 100% occurrence on all dives. Also, Anthomastus spp. were present at both sites, but exhibited substantially higher densities around Las Gemelas Seamount than at Isla del Coco.
We examined invertebrate densities at both Isla del Coco and Las Gemelas Seamount with respect to habitat type, slope, and rugosity. Whereas invertebrate densities were highest in sand, rock, and boulder habitats at Las Gemelas, invertebrates at Isla del Coco were denser in pinnacle, rock, mixed areas of sand and rock, pebble, and boulder habitats (Fig. 3). When comparing density with slope, we found that invertebrate densities at Isla del Coco increased with increasing slope, whereas invertebrate densities at Las Gemelas were highest in moderate slopes of 30° to 60° (Fig. 4). A comparison of invertebrate densities by rugosity category indicated that medium to high rugosity habitats contained the highest invertebrate densities at both sites (Fig. 5).
Habitat types: We encountered a total of 14 combinations of the seven substratum codes. At Isla del Coco, 58% of the habitats surveyed were comprised primarily of rock and 34% were primarily sand, whereas at Las Gemelas, 73% of the transects covered rocky habitats and 25% covered sandy habitats (Table 5). Transects at Las Gemelas Seamount occurred over somewhat more rugose habitats. At Isla del Coco, 39.9%, 28.5%, and 31.6% of the habitat area surveyed was high, medium, and low rugosity, respectively. At Las Gemelas Seamount, 46.8%, 34.3%, and 18.9% of the habitat area surveyed was high, medium, and low rugosity, respectively. Similarly, transects at Las Gemelas more typically occurred on higher slopes. At Las Gemelas Seamount, 30%, 56%, and 14% of slops in the hábitat area surveyed were <30°, 30–60°, and >60°, respectively. At Isla del Coco, 48%, 35%, and 17% of the slopes in the habitat area surveyed were <30°, 30–60°, and >60°, respectively.
Quantification of Percent Cover: All six taxonomic groups used in the quantification of percent invertebrate cover were present at both Isla del Coco and Las Gemelas Seamount (Table 6). Brachiopods, glass sponges and polychaetes were more prevalent at Isla del Coco, whereas encrusting sponges, polyps/soft corals and Stylaster spp. were found in higher frequencies at Las Gemelas (Fig. 6). Encrusting sponges were the most commonly encountered taxonomic group at both sites. We evaluated invertebrate percent cover at both Isla del Coco and Las Gemelas Seamount with respect to habitat type, slope and rugosity. Whereas encrusting sponges and Stylaster spp. dominated every habitat type at Las Gemelas, percent cover of encrusting sponges and Stylaster spp. at Isla del Coco were greater in rock, pinnacle, and mixed areas of sand and rock or pebble habitats only (Fig. 7). Glass sponges dominated habitats consisting primarily of boulders at Isla del Coco. When evaluating percent cover with slope, we found that all invertebrate groups were more prevalent in areas with intermediate (30° to 60°) or steep (> 60°) slopes (Fig. 8). A comparison of percent cover and rugosity indicated that high rugosity habitats contained the highest frequencies of all invertebrates at both sites, with the exception of glass sponges and polychaetes at Isla del Coco, which were found in greater quantities at intermediate levels of rugosity (g. 9).
Discussion
In September 2009, we surveyed the habitats and invertebrate communities in the deep- water portions of Las Gemelas Seamount and Isla del Coco. We used a combination of quantitative strip transects and general observations to characterize invertebrate communities at each study site. We surveyed habitats in depths ranging from 50-402m, but reported only the data obtained from dives in rocky habitats that were deeper than 150m to facilitate a comparison of deepwater communities at Las Gemelas Seamount and Isla del Coco. Habitats surveyed at Isla del Coco included vertical rock walls and steep slopes consisting of highly fragmented areas of volcanic rock outcrops and sand. Sea urchins and sponges were among the most commonly observed taxonomic groups. Higher densities of these organisms were found at slopes greater than 60° and in areas displaying high levels of rugosity. Pinnacles and rocky habitats housed the highest overall densities of invertebrates, although the highest density of an invertebrate taxon at Isla del Coco was still much lower than from Las Gemelas Seamount.
Seamounts are important features in the world’s oceans, and an increasing amount of research has been devoted to studying the biological communities and patterns of benthic biodiversity of seamounts. Not surprisingly, with increased information more questions are generated about the ecological role of seamounts and their vulnerability to disturbance. Many researchers have suggested that seamounts are unique environments, containing hotspots of biodiversity and endemism (Clark et al. 2010, Rowden et al. 2010b), and are very fragile (Koslow et al. 2001). Although Samadi et al. (2006) and Thoma et al. (2009) indicated that genetic studies might refute the concept of increased endemism on seamounts, they did agree with the hypothesis that seamounts are diversity hotspots, possessing benthic assemblages with particularly high species richness.
Habitats surveyed at Las Gemelas contained rich, dense assemblages of invertebrates. Some of the prominent benthic invertebrates included sea urchins, black corals, sponges, anemones, corals, gorgonians, echinoderms, and crustaceans. Two taxonomic groups (sea urchins and black corals) accounted for more than 70% of the total combined invertebrate density at Las Gemelas. These very prevalent taxa were found at higher frequencies in areas of rock and/or sand, intermediate slopes (30° to 60°), and medium levels of rugosity. These differences in assemblages of deepwater invertebrates is similar to those shown by Ricciardi and Bourget (1999) and Adami et al. (2004) for intertidal invertebrates. They showed that significant differences in community structure were found between vertical and horizontal samples in different locations. Also, differences in structure were much greater between localities than among replicates within each locality. Rowden et al. (2010a) analyzed more than 5000 sea-floor images on 20 southwest Pacific seamounts and reported that the mean biomass of epibenthic megafauna was nearly four times greater than on the adjacent continental slope at comparable depths.
The difference in invertebrate densities between the two sites may also be attributed to habitat heterogeneity. At Isla del Coco, 74% of habitats were either all rock or some mixture of rock and sand. Habitats at Las Gemelas, however, were more diverse, and consisted pri- marily of rock, sand, and/or pinnacles. Another potential explanation for the difference in invertebrate densities is that the habitats surveyed at Las Gemelas covered a smaller range of slope categories (most slopes were 30º to 60º) than present at Isla del Coco, which was comprised primarily of more extreme slopes (typically <30º or >60º). Mendoza et al. (2010) showed that deep, steep underwater slopes as prime habitats for many decapod crustaceans and suggested that these habitats are under- studied, thus may contain many new species. Until more surveys are conducted to enable an analysis of species-area curves, we cannot fully explain the observed differences. Importantly, we encountered different habitats in each of our dives at Las Gemelas, indicating that the sea- mounts are likely to include a wider variety of habitats than those surveyed. This suggests that the diversity of taxa at Las Gemelas Seamount is potentially much greater than we were able to determine with the available submersible dives.
One important qualitative observation is that we saw a larger number of encrusting and structure-forming invertebrates at Las Gemelas Seamount than at Isla del Coco. This habitat complexity further indicates that habitats at Las Gemelas may be able to harbor a greater diversity and biomass of both invertebrate and fish species than at Isla del Coco. O’Hara et al. (2008) reported that the large biogenic structures formed by colonial cold-water scleractinian corals provide valuable habitat for marine invertebrates on seamounts. They suggested that cold-water scleractinian corals are a key habitat for a many ophiuroid species that occur in the Western Pacific Ocean.
Given the diverse and rich assemblages of invertebrates carpeting the bottom habitats at Las Gemelas Seamount, as well as the fact that the area contains numerous species which have not yet been described in the scientific literature (and thus, are important for the maintenance of biodiversity), Las Gemelas Seamount may be a richer environment for invertebrates than the deep water surrounding Isla del Coco. Koslow et al. (2001) discussed concerns about the vulnerability of seamount communities to human impacts, especially with the development of large-scale bottom trawl fisheries in the deep sea. They reported that 24 to 43% of the invertebrate species they recovered from experimental trawling were new to science, and between 16 and 33% appeared to be restricted to the seamount environment. The concern expressed by Koslow et al. (2001) is that seamount invertebrate communities are fragile. O’Hara et al. (2008) believe that the slow growth rate of seamount corals that recovery of damaged cold-water coral assemblages will be a long-term process. The high densities of invertebrates in the area around Isla del Coco National Park and Las Gemelas seamount signifies the need for increased protection and study of these areas, because of the paucity of information related to the distribution and relative abundance of the important resources that could easily be overexploited.
Acknowledgments
We thank Shmulik Blum, Sylvia Earle, Avi Klapfer, Bruce Robison, Edith Widder, and the crew of the Argo for safe submersible operations and help with observations of fishes and macroinvertebrates. Katie Schmidt helped excerpt information from video and Dan Malone provided the MDS plots. Fund- ing was provided by the National Geographic Society, California Sea Grant Program, Moss Landing Marine Labs, and the Universidad de Costa Rica.
References
Adami M., A. Tablado & J. Lopez-Gappa. 2004. Spatial and temporal variability in intertidal assemblages dominated by the mussel Brachidontes rodriguezii (d’orbigny, 1846). Hydrobiologia 520: 49-59. [ Links ]
Bray J.R. & J.T. Curtis. 1957. An ordination of the upland forest communities of Southern Wisconsin. Ecol. Monogr. 27: 325-349. [ Links ]
Breedy, O. & J. Cortés. 2008. Octocorals (Coelenterata: Anthozoa: Octocorallia) of Isla del Coco, Costa Rica. Rev. Biol. Trop. 56 (Suppl. 2): 71-77. [ Links ]
Cairns, S.D. 1991a. A revision of the ahermatypic Scleractinia of the Galápagos and Cocos Islands. Smithsonian Contr. Zool. 504: 1-33. [ Links ]
Cairns, S.D. 1991b. New records of Stylasteridae (Hydrozoa: Hydroida) from the Galápagos and Cocos Islands. Proc. Biol. Soc. Washington 104: 209-228. [ Links ]
Clark, M.R., A. A. Rowden, T. Schlacher, A. Williams, M. Consalvey, K.I. Stocks, A.D. Rogers, T.D. O’Hara, M. White, T.M. Shank & J.M. Hall-Spencer. 2010. The ecology of seamounts: Structure, function, and Human impacts. Ann. Rev. Mar. Sci. 2: 253-278. [ Links ]
Cortés, J. 2008. Historia de la investigación marina de la Isla del Coco, Costa Rica. Rev. Biol. Trop. 56 (Suppl. 2): 1-18. [ Links ]
Cortés, J. & S. Blum. 2008. Life to 450 m depth at Isla del Coco, Costa Rica. Rev. Biol. Trop. 56 (Suppl. 2): 189-206. [ Links ]
Greene, H.G., M.M. Yoklavich, R.M. Starr, V.M. O’Connell, W.W.Wakefield, D.E. Sullivan, J.E. McRea & G.M. Cailliet. 1999. A classification scheme for deep sea- floor habitats. Oceanol. Acta 22: 663-678. [ Links ]
Koslow J.A., G.W. Boehlert, J.D.M. Gordon, R.L. Haedrich, P. Lorance & N. Parin. 2000. Continental slope and deep-sea fisheries: Implications for a fragile ecosystem. ICES J. Mar. Sci. 57: 548-557. [ Links ]
Koslow J.A., K. Gowlett-Holmes, J.K. Lowry, T. O’Hara, G. Poore & A. Williams. 2001. Seamount benthic macrofauna off southern Tasmania: Community structure and impacts of trawling. Mar. Ecol. Prog. Ser. 213: 111-125. [ Links ]
Lizano, O.G. 2008. Dinámica de aguas alrededor de la Isla del Coco, Costa Rica. Rev. Biol. Trop. 56 (Suppl. 2): 31-48. [ Links ]
Lizano, O.G. 2012. Rasgos morfológicos alrededor de la Isla del Coco y de sus montes submarinos vecinos, Pacífico de Costa Rica. Rev. Biol. Trop. 60 (Suppl.
3): 43-51. [ Links ]
O’Hara T.D., A.A. Rowden & A. Williams. 2008. Cold- water coral habitats on seamounts: Do they have a specialist fauna? Biodivers. Lett. 14: 925-934. [ Links ]
Mendoza, J.C.E., T. Naruse, S. Tan, T. Chan, D.F. Richer & P.K.L. Ng. 2010. Case studies on decapod crustaceans from the Philippines reveal deep, steep underwater slopes as prime habitats for ‘rare’ species. Biodivers. Conserv. 19: 575-586. [ Links ]
Reiswig, H.M. 2010. A new species of Tretodictyum (Porifera: Hexactinellida: Tretodictyidae) from off Cocos Island, tropical eastern Pacific Ocean. Proc. Biol. Soc. Washington 123: 242-250. [ Links ]
Ricciardi, A. & E. Bourget. 1999. Global patterns of macroinvertebrate biomass in marine intertidal communities. Mar. Ecol. Prog. Ser. 185: 21-35. [ Links ]
Richer de Forges, B., J.A. Koslow & G.C.B. Poore. 2000. Diversity and endemism of the benthic seamount fauna in the southwest Pacific. Nature 405: 944-947. [ Links ]
Rojas, W. & G.E. Alvarado. 2012. Geología y contexto geotectónico de la Isla del Coco y la zona marítima frente al Pacífico Central de Costa Rica. Rev. Biol. Trop. 60 (Suppl. 3): 15-32. [ Links ]
Rowden, A.A., T.A. Schlacher, A. Williams, M.R. Clark, R. Stewart, F. Althaus, D.A. Bowden, M. Consalvey, W. Robinson & J. Dowdney. 2010a. A test of the seamount oasis hypothesis: Seamounts support higher epibenthic megafaunal biomass than adjacent slopes. Mar. Ecol. 31: 95-106. [ Links ]
Rowden, A.A., J.F. Dower, T. A. Schlacher, M. Consalvey & M.R. Clark. 2010b. Paradigms in seamount ecology: fact, fiction and future. Mar. Ecol. (Suppl.) 131: 226-241. [ Links ]
Samadi, S., L. Bottan, E. Macpherson, B. Richer De Forges & M. Boisselier. 2006. Seamount endemism questioned by the geographic distribution and population genetic structure of marine invertebrates. Mar. Biol. 149: 1463-1475. [ Links ]
SINAC, MINAET. 2008. GRUAS II: Propuesta de Ordenamiento Territorial para la conservación de la biodiversidadde Costa Rica. Volumen 3: Análisis de Vacíos en la Representatividad e Integridad de la biodiversidad marina y costera. Sistema Nacional de Áreas de Conservación (SINAC), Ministerio de Ambiente, Energía y Telecomunicaciones (MINAET), San José, Costa Rica. 60 p. [ Links ]
Starr, R.M. & M.M. Yoklavich. 2008. Monitoring MPAs in Deep Water off Central California: 2007 IMPACT Submersible Baseline Survey. California Sea Grant College Program Publication T-067. 24 p. [ Links ]
Starr, R.M., K. Green & E. Sala. 2012. Deepwater fish assemblages at Isla del Coco National Park and Las Gemelas Seamounts, Costa Rica. Rev. Biol. Trop. 60 (Suppl. 3): 347-362. [ Links ]
Thoma, J.N., E. Pante, M.R. Brugler & S.C. France. 2009. Deep-sea octocorals and antipatharians show no evidence of seamount-scale endemism in the NW Atlantic. Mar. Ecol. Prog. Ser. 397: 25-35. [ Links ]
Tissot, B.N., M.A. Hixon & D.L. Stein. 2007. Habitat- based submersible assessment of macro-invertebrate and groundfish assemblages at Heceta Bank, Oregon, from 1988 to 1990. J. Exp. Mar. Biol. Ecol. 352: 50-64. [ Links ]
Yoklavich, M.M. & V. O’Connell. 2008. Twenty Years of Research on Demersal Communities Using the Delta Submersible in the Northeast Pacific. In J.R. Reynolds & H.G. Greene (eds.). Marine Habitat Mapping Technology for Alaska. Alaska Sea Grant College Program, University of Alaska Fairbanks. doi:10.4027/mhmta.2008.10. [ Links ]
Zar, H. 1999. Biostatistical Analysis. 4th Ed. Prentice Hall, Upper Saddle River, New Jersey. [ Links ]
*Correspondencia a:
Richard M. Starr. University of California Sea Grant Program and Moss Landing Marine Laboratories, 8272 Moss Landing Road, Moss Landing, California 95039, USA; starr@mlml.calstate.edu
Jorge Cortés. Centro de Investigación en Ciencias del Mar y Limnología (CIMAR), Ciudad de la Investigación, Universidad de Costa Rica, San Pedro, 11501-2060 San José, Costa Rica; jorge.cortes@ucr.ac.cr
Cheryl L. Barnes. University of California Sea Grant Program and Moss Landing Marine Laboratories, 8272 Moss Landing Road, Moss Landing, California 95039, USA.
Kristen Green. Alaska Department of Fish and Game, 304 Lake Street, Room 103, Sitka, Alaska, 99835; kristen.green@alaska.gov
Odalisca Breedy. Centro de Investigación en Ciencias del Mar y Limnología (CIMAR), Ciudad de la Investigación, Universidad de Costa Rica, San Pedro, 11501-2060 San José, Costa Rica
1. University of California Sea Grant Program and Moss Landing Marine Laboratories, 8272 Moss Landing Road, Moss Landing, California 95039, USA; starr@mlml.calstate.edu
2. Centro de Investigación en Ciencias del Mar y Limnología (CIMAR), Ciudad de la Investigación, Universidad de Costa Rica, San Pedro, 11501-2060 San José, Costa Rica; jorge.cortes@ucr.ac.cr
3. Alaska Department of Fish and Game, 304 Lake Street, Room 103, Sitka, Alaska, 99835; kristen.green@alaska.gov
Received 05-III-2012. Corrected 06-IX-2012. Accepted 24-IX-2012.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}