










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.60 suppl.3 San José Nov. 2012
Apendicularias (Urochordata) y quetognatos (Chaetognatha) del Parque Nacional Isla del Coco, Costa Rica
Appendicularians (Urochordata) and chaetognaths (Chaetognatha) of Isla del Coco National Park, Costa Rica
Appendicularians (Urochordata) and chaetognaths (Chaetognatha) of Isla del Coco National Park, Costa Rica
Iván Castellanos-Osorio1*, Rosa María Hernández-Flores1, Álvaro Morales-Ramírez2*,3* & Marco Corrales-Ugalde3
*Dirección para correspondencia:
Abstract
Appendicularians and chaetognaths are important members of the zooplancton but little studied in Costa Rica and not at all at Isla del Coco Nacional Park, eastern tropical Pacific. Epipelagic (0-50m) zooplankton samples from the oceanic area off Isla del Coco, were collected in November 2001, and over patch reefs in 2009. We analized the species composition of appendicularians and chaetognaths present in 31 samples. Nine species of appendicularians, and eight species of chaetognaths were identified. Oikopleura longicauda was the most abundant in the surveyed area (217 specimens, 65.56% of total appendicularian numbers), followed by Oikopleura rufescens with 35 (10.57%). All the appendicularian species found in this study are common in warm ocean waters, but six species herein recorded for the first time from waters off Isla del Coco, and two species are new records for Costa Rica. Among the chaetognaths, Flaccisagitta enflata was the commonest species (85 specimens, 50.6% of total numbers) recorded in this study, followed by Serratosagitta pacifica with 38 organisms (20.62%). Zonosagitta bedoti was represented by a single specimen. These species have been recorded from tropical waters of the Eastern Pacific, but six species are new records for Isla del Coco. Data on the distribution and morphology of the species recorded are presented together with keys for the identification of both taxa in the surveyed area.
Key words: appendicularians, chaetognaths, Isla del Coco, Costa Rica, identification key.
Resumen
Las apendicularias y quetognatos son miembros importantes del zooplancton pero poco estudiados en Costa Rica y no del todo en el Parque Nacional Isla del Coco, Pacífico tropical este. Se recolectaron muestras epipelágicas de zooplancton de la zona oceánica (0-50m) de la Isla del Coco, en noviembre de 2001 y sobre parches arrecifales en 2009. Se analizó la composición de especies de apendicularias y quetognatos presentes en 31 muestras. Nueve especies de apendicularias, y ocho especies de quetognatos fueron identificadas. Oikopleura longicauda fue la más abundante en el área estudiada (217organismos, representando el 65.56% del número total de apendicularias), seguida por Oikopleura rufescens, con 35 (10.57%). Todas las especies de apendicularias encontradas en este estudio son comunes en las aguas cálidas de los océanos, seis especies se registran aquí por primera vez en aguas de la Isla del Coco y dos son nuevos registros para Costa Rica. Entre los quetognatos, Flaccisagitta enflata fue la especie más común (85 ejemplares, representando el 50.6% del número total) registrada en este estudio, seguida por Serratosagitta pacifica con 38 organismos (20.62%). Zonosagitta bedoti estuvó representada por un solo espécimen. Estas especies también se han registrado en las aguas tropicales del Pacífico oriental, pero seis especies son nuevos registros para Isla del Coco. Los datos sobre la distribución y morfología de las especies registradas se presentan junto con las claves para la identificación de los taxones en el área estudiada.
Palabras clave: apendicularias, quetognatos, Isla del Coco, Costa Rica, clave de identificación
En general, la investigación sobre organismos zoopláncticos en los mares de Costa Rica, ha estado limitada a ciertas zonas costeras e incluso, a ciertos taxa (Castellanos et al. 2009, Gasca 2009, Rodríguez-Sáenz & Gasca 2009); este desconocimiento no solo se restringe a zonas geográficas, sino también a diversos componentes de la comunidad zooplánctica, que aún no han sido aún estudiados. Además, mucha de la información sobre el zooplancton de Costa Rica ha sido generada por investigadores de otros países y en regiones de interés oceanográfico y económico, como lo es el Domo de Costa Rica (Suárez-Morales & Gasca 1989, FernándezÁlamo & Farber-Lorda 2006). La presencia de apendicularias y quetognatos ha sido reportada en diversos cuerpos de agua costeros, como: Bahía Culebra, Golfo Dulce y Golfo de Nicoya, entre otros sitios. Sin embargo, el número de publicaciones que incluyen datos de especies sobre apendicularias en Costa Rica se reduce a sólo tres: una observación realizada frente a la costa de Guanacaste (Barham 1979), en la costa del Pacífico; el de Morales-Ramírez (obs. pers.) quién analizó las tazas de filtración de las apendicularias en el Golfo de Nicoya y Morales-Ramírez (2008) reportó la presencia de ocho especies alrededor de Isla del Coco, pero solamente menciona a Oikopleura rufescens, Oikopleura fusiformis y al género Stegosoma. Castellanos et al. (2009) encontró Oikopleura rufescens, Oikopleura longicauda, Oikopleura fusiformis, Fritillaria pellucida y Fritillaria formica, en aguas de Bahía Culebra. Con respecto a los quetognatos, han sido más estudiados en aguas costarricenses, algunos trabajos son los de: Morones (1988) y Segura et al. (1992) quienes registraron 13 especies, en muestras oceánicas recolectadas en el Domo de Costa Rica; en ambientes costeros Hossfeld (1996), reportó 8 especies de quetognatos, tanto en el Golfo de Nicoya, como en Golfo Dulce, y Alvariño (1972) determinó la presencia de nueve especies en aguas de Costa Rica, al analizar muestras recolectadas en nueve cruceros oceanográficos. Suárez-Morales et al. (2009) registraron 19 en aguas del Pacífico costarricense y Morales-Ramírez (2008) encontró alrededor de Isla del Coco, siete especies de quetognatos, pero únicamente menciona a Flaccisagitta enflata y Serratosagitta pacifica. A partir de muestras de zooplancton recolectadas en los alrededores del Parque Nacional Isla del Coco se presentan datos taxonómicos y morfológicos de las apendicularias y los quetognatos. En este trabajo se amplía la información referente a estos dos grupos del zooplancton al incrementar el número de especies descritas en aguas del Parque Nacional Isla del Coco. Se incluyen claves para la identificación de las especies encontradas en esta área.
Material y Métodos
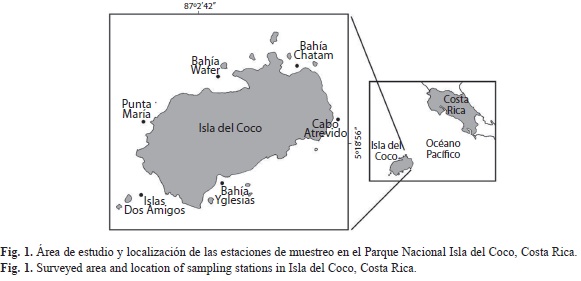
La Isla del Coco (5°32’N y 87°04’W), está ubicada en aguas oceánicas del Pacífico Tropical Oriental y conforma el núcleo del Área de Conservación Marina Isla del Coco. Este Parque Nacional fue decretado desde 1997 Patrimonio Mundial de la UNESCO (Cortés 2008).
Con el propósito de estudiar la composición del zooplancton de la Isla del Coco, se realizó un crucero oceanográfico en Noviembre de 2001. Se efectuaron muestreos de zooplancton alrededor de la isla, en seis estaciones: Bahía Chatham, Bahía Wafer, Punta María, Isla Dos Amigos, Bahía Yglesias y Cabo Atrevido (Fig. 1), y seis muestreos sobre parches de arrecifes, en 2009. Los arrastres de zooplancton fueron verticales en los primeros 50m de profundidad y horizontales con una duración de 3 a 4 minutos, en ambos casos se emplearon redes con un diámetro de boca de 0.49cm y aberturas de malla de 100, 200 y 500μm.
Las muestras fueron fijadas en formaldehido al 4% y preservadas en etanol al 70%. Se separaron los quetognatos y las apendicularias de un total de 31 muestras y se procesaron para su identificación. Los especímenes analizados y las muestras originales se encuentran depositados en el CIMAR, Universidad de Costa Rica, San Pedro, San José, Costa Rica.
Las apendicularias fueron identificadas empleando los trabajos de Tokioka y Suárez-Caabro (1956), Esnal (1981, 1996) y Fenaux (1993, 1998). Los quetognatos adultos con los trabajos de Bieri (1991), Pierrot-Bults (1996) y Casanova (1999).
Morfología general
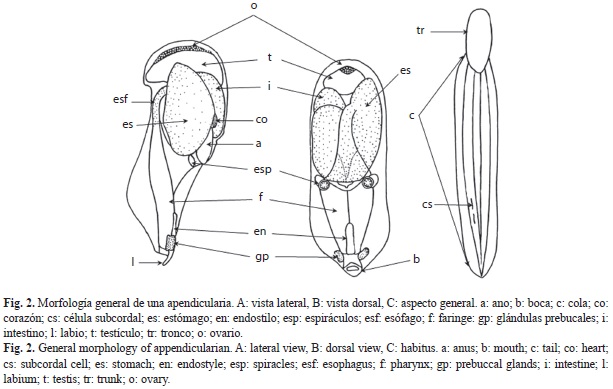
Apendicularias: El cuerpo de estos organismos consiste de un pequeño tronco frecuentemente menor de un milímetro de longitud y una cola, que usualmente no excede tres veces la longitud del cuerpo. La boca se abre a la cavidad faríngea, con dos aberturas branquiales que terminan en dos espiráculos. La región faríngea posee un endostilo corto, formado por células musculares largas. Algunas especies presentan un par de glándulas bucales cerca del extremo anterior del endostilo. El estómago es aproximadamente globular y puede tener lóbulos; antecede a un intestino más o menos oval y al recto, que termina en un ano abierto ventral o lateralmente (Fig. 2).
Son hermafroditas (excepto Oikopleura dioica); las gónadas ocupan una parte relativamente grande del cuerpo. La cola tiene un cordón nervioso y está cubierta con musculatura bien desarrollada. Las células subcordales y anficordales están ausentes en algunas especies (Tokioka & Suárez-Caabro 1956, Esnal 1981, Raymont 1983).
Las características más usadas en su taxonomía actual son: la presencia o ausencia de glándulas prebucales, forma de los espiráculos, forma y disposición de las gónadas, presencia o ausencia y número de células anficordales, desarrollo de un ciego en el lóbulo izquierdo del estómago y presencia o ausencia, número y disposición de células subcordales (Fenaux 1993).
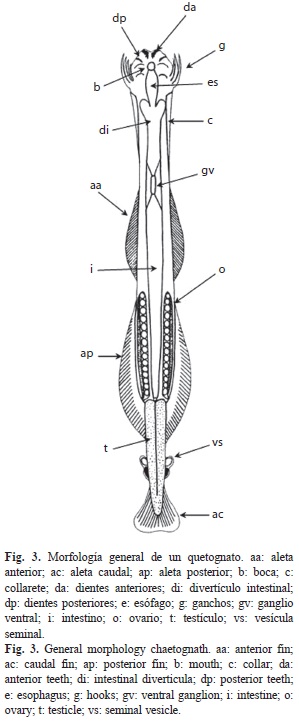
Quetognatos: Son organismos transparentes o translúcidos que están recubiertos de una cutícula. Tienen una cabeza, un tronco y una cola diferenciados. Presentan varias espinas quitinosas a ambos lados de la cabeza que guardan en una capucha cuando nadan; en la parte ventral está la boca y una o dos hileras de dientes pequeños a cada lado; en la parte dorsal hay una par de ojos con pigmentación característica en las diferentes especies. Todos los quetognatos presentan una aleta caudal, y dependiendo del género uno o dos pares de aletas laterales (Fig. 3). Todos son hermafroditas y se han descrito especies productoras de neurotoxinas, que usan para atrapar presas (Thuesen 1991, Pierrot-Bults 1996).
Carecen de sistemas respiratorio, circulatorio o excretor. Los únicos órganos dentro del tronco son el tracto digestivo, que consta de un esófago en la región del cuello y puede ser un tracto simple recto o con divertículos, y los ovarios, los cuáles son tubos con huevos que varían en tamaño, dependiendo de la especie.
La forma y posición de las vesículas seminales, ubicadas sobre la cola, tienen valor taxonómico pues es variable en las diversas especies.
Alcanzan una longitud que oscila entre 2mm y 12cm. Se han descrito más de 120 especies repartidas en 20 géneros, son organismos marinos pláncticos, excepto un género que es béntico (Spadella) (Bieri 1991, Pierrot-Bults 1996).
Resultados
A partir del análisis taxonómico de las apendicularias recolectadas en el Parque Nacional Isla del Coco, se determinó un total de nueve especies, pertenecientes a dos familias y tres géneros. Las especies: Oikopleura longicauda, O. dioica, O. gracilis, O. cophocerca, Fritillaria haplostoma y F. formica f. digitata y f. tuberculata son nuevos registros para la Isla del Coco. Su arreglo sistemático se basó en Fenaux (1993, 1998).
Familia Oikopleuridae (Lohmann, 1915)
Tronco ovoide, endostilo recto. Un par de pasajes branquiales con aros ciliados conecta la cavidad faríngea con el exterior, cada uno de estos termina en una apertura del espiráculo. Espiráculos ubicados en la región del recto. Pared del estómago compuesta por numerosas células muy pequeñas con una fila de pocas células grandes. Es la familia más diversa de apendicularias, con 11 géneros y 37 especies. Esta familia se encuentra ampliamente distribuida en todos los océanos.
Oikopleura longicauda (Vogt, 1854)
Esta especie fue la más abundante en el área de estudio con 217 organismos, representando el 65.56% del número total de apendicularias. Tronco compacto, superior a los 1.4mm de longitud. Se caracteriza por la presencia de un gran capuchón cuticular (Fig. 4a). El endostilo casi a la misma distancia entre la boca y el ano. La cola es larga, fuerte y de aspecto iridiscente.
Es una especie euritérmica y eurihalina; es la más común de las grandes regiones oceánicas, y su distribución alcanza las aguas antárticas. Es poco abundante en regiones con temperatura inferior a 15°C (Fenaux 1967). En Costa Rica está especie se observó en aguas de Bahía Culebra (Castellanos et al. 2009).
Oikopleura rufescens Fol, 1872
Apendicularia que se observó con gran frecuencia, aunque no en grandes números, fue la segunda especie más abundante (35 organismos), representando el 10.57% del total.
Se caracteriza por tener el tronco con el contorno dorsal curvado. El tronco alcanza tallas superiores a los 2.3mm. Glándulas prebucales esféricas y largas. El endostilo casi a la misma distancia entre la boca y el ano. La cola presenta una célula subcordal, grande y de forma fusiforme.
Especie euritérmica y eurihalina, ampliamente distribuida en los océanos Atlántico, Pacífico e Indico (Tokioka & Suárez-Caabro 1956, Esnal 1981). Oikopleúrido registrado en Bahía Culebra (Castellanos et al. 2009).
Oikopleura fusiformis f. typica Fol, 1872
Su abundancia oscilo entre 1 y 11 organismos, representando el 8.76% del total de apendicularias.
La longitud del cuerpo oscila entre 1 y 1.4mm. El tronco es alargado, fusiforme y angosto. No tiene glándulas prebucales y el endostilo es corto. El lóbulo del estómago se caracteriza por tener un ciego levemente curvado y dirigido oblicuamente hacia atrás. La cola es larga y angosta y carece de células subcordales. Es una especie eurihalina propia de regiones oceánicas y común en las aguas cálidas (Tokioka & Suárez-Caabro 1956, Fenaux 1967, Zoppi de Roa 1971). Especie de apendicularia observada en aguas de Bahía Culebra (Castellanos et al. 2009).
Oikopleura dioica Fol, 1872
Solo se observó un ejemplar de esta especie. Es de tamaño pequeño, la longitud del tronco oscila entre 0.5 y 1mm y la cola de 2 a 4mm. El tronco es ovoide. El extremo posterior del endostilo llega cerca del ano. Glándulas prebucales de contorno redondeado y pequeñas. Esta es la única especie de apendicularia que presenta los sexos separados, tanto el ovario como los testículos se encuentra en la parte posterior del tronco. La cola presenta en el lado derecho dos células subcordales fusiformes en una hilera (Fig. 4b).
Se ha observado tanto en ambientes costeros como oceánicos de regiones con aguas templadas y algunas veces puede encontrarse en regiones boreales, es por lo tanto, excepcionalmente entre las apendicularias, euritérmica y eurihalina. Esta especie se registra por vez primera para Costa Rica.
Oikopleura gracilis (Lohmann, 1896)
Se registro sólo en tres ocasiones, constituyendo el 4.53% de total de apendicularias. Glándulas prebucales ausentes. El endostilo está ubicado más cerca del ano que de la boca. El lóbulo izquierdo del estómago es redondeado y no presenta ciego digestivo. En individuos maduros, las gónadas se extienden dorsalmente hacia atrás, dándole una forma característica al tronco. La cola es casi cinco veces más grande que el tronco. Células subcordales ausentes.
Apendicularia de aguas cálidas (Fenaux 1967), común en la región oceánica del Atlántico sudoccidental (Esnal 1981). Aravena & Palma (2002) la han encontrado en sitios cercanos a la costa de Chile y Esnal y Castro (1977) reportaron O. gracilis en aguas oceánicas. Este es el primer registro para aguas de Costa Rica.
Oikopleura cophocerca (Gegenbaur, 1855)
Especie poco frecuente, observada en una sola estación con tres ejemplares. Tronco alargado y el contorno dorsal es ligeramente arqueado. Boca con un labio inferior desarrollado, glándulas prebucales redondas y bien desarrolladas. Lóbulo izquierdo del estómago generalmente alargado, con un ciego en forma de bolsillo en el margen posterior. Endostilo pequeño y situado entre la boca y el ano. Es una especie termófila cosmopolita, común en aguas cálidas y templadas y frecuente en la región oceánica (Fenaux 1967, Esnal 1981, Aravena & Palma 2002) y poco abundante en aguas superficiales (Flores-Coto 1965, Esnal & Castro 1977). Este es el primer registro para aguas de Costa Rica.
Stegosoma magnum (Langerhans, 1880)
Apendicularia poco frecuente en aguas de la Isla del Coco, registrándose solamente 12 ejemplares. Es la especie de mayor tamaño que se encontró (11mm), tiene el tronco alargado, comprimido y con el perfil dorsal arqueado, y alcanza una longitud superior a los 4mm. Glándulas prebucales esféricas y pequeñas (Fig. 4d). La cola es musculosa y presenta en el lado derecho 8 células subcordales en hilera (Fig. 4c, e).
Es una especie euritérmica que se ha observado en todos los océanos, con excepción del Ártico (Fenaux 1967). Es el primer registro para aguas de Costa Rica.
Familia Fritillariidae (Lohmann, 1915)
Tronco dorsoventralmente comprimido o en forma de espiga. Endostilo curvo hacia arriba. Los aros ciliados de los espiráculos conectan la cavidad faríngea con el exterior directamente sin un pasaje branquial tubular. Espiráculos situados distintamente en la parte anterior de la cavidad faríngea. Esta familia está representada por tres géneros y 30 especies, siendo el género Fritillaria el que tiene más especies. Generalmente estas apendicularias son más pequeñas que las de la familia Oikopleuridae.
Fritillaria formica
(Lohmann en Lohmann & Büchmann, 1926)
Esta especie representó el 5.14% del total de apendicularias, con 17 ejemplares distribuidos en tres estaciones. Tronco alargado y doblado hacia el dorso en la región digestiva. Los arcos ciliados de los espiráculos no entran en contacto con la línea ventral media. Musculatura de la cola gruesa en la parte media, se angosta distalmente. Fritillaria formica presenta dos variaciones que se identifican según la forma y los apéndices del lóbulo medio del labio superior y la forma de los testículos cuando el organismo está maduro (ver clave).
Se localiza en todos los mares cálidos y en zonas oceánicas (Zoppi de Roa 1971, Aravena & Palma 2002). Es una de las apendicularias más frecuentes en las aguas cálidas del Atlántico sudoccidental (Esnal 1981). Ambas formas de esta especie se registran por primera vez para aguas de Costa Rica.
Fritillaria haplostoma
(Fol, 1872; emend. Fol, 1874)
Especie rara, presente sólo en una estación con dos organismos. Tronco alargado en forma de espiga y ligeramente arqueado dorsalmente. La boca tiene solo un aro ciliado y un protuberante labio superior. Tracto digestivo longitudinal y espiráculos circulares. El ovario es esférico y posterior a este se encuentra la testes alargada. La cola es ancha y acuminada distalmente, presenta numerosas células glandulares pequeñas. La musculatura de la cola es angosta.
Es una especie que se ha encontrado en todos los mares cálidos, en aguas costeras y oceánicas (Tokioka & Suárez-Caabro 1956, Zoppi de Roa 1971, Esnal & Castro 1977).
Aunque la clave que se presenta a continuación representa las nueve especies identificadas en este estudio, es posible que existan más especies en la zona que no fueron recolectadas, debido a que se muestreó sólo el estrato superficial (0-50m).
Clave para la determinación de las especies de apendicularias en la Isla del Coco
Del análisis de las 31 muestras de zooplancton en el Parque Nacional Isla del Coco, se determinó un total de ocho especies de quetognatos, pertenecientes a siete géneros y tres familias. De estas especies, seis son nuevos registros para la Isla del Coco (Krohnitta pacifica, Aidanosagitta neglecta, Ferosagitta robusta, Zonosagitta bedoti, Z. pulchra y Pterosagitta draco). El arreglo sistemático de las especies identificadas se basó en Bieri (1991).
Familia Krohniitidae Tokioka, 1965
Krohnitta pacifica (Aida, 1897)
Fue la quinta especie en abundancia, registrándose con solamente 5 ejemplares, que corresponde al 2.98% del total de quetognatos. Especie de tamaño pequeño (8mm), los ganchos con placas finas en número de 8 a 11 y con una hilera de 16 dientes, en cada lado. El cuerpo es delgado, transparente. Cabeza poco grande con cuello bien marcado; las aletas laterales son cortas y parcialmente radiadas. Ojos pequeños con pigmento en forma de estrella; vesículas seminales angostas, con cabeza y tronco, tocando las aletas posteriores y la aleta caudal; ovarios largos, huevos pequeños, redondos. Es una especie epipelágica a mesopelágica somera, su distribución es circumglobal, 30°N-30°S (Bieri 1959, Alvariño 1972, Pierrot-Bults & Nair 1991). Esta especie ha sido registrada en aguas de Costa Rica (Morones 1988, Segura et al. 1992, Hossfeld 1996).
Familia Sagittidae Claus & Groben, 1905
Aidanosagitta neglecta (Aida, 1897)
Fue la tercera especie en abundancia, registrándose con 17 organismos, que corresponden al 10.12% del total de quetognatos. Especie de tamaño pequeño (8mm), con 3 a 7 ganchos, 7 dientes anteriores y 18 posteriores, en cada lado. El cuerpo es firme y opaco. Cabeza pequeña con cuello inconspicuo; las aletas anteriores y posteriores son redondeadas y completamente radiadas, las primeras poco más cortas. Collarete largo, con divertículo intestinal conspicuo; ojos redondeados y grandes, con pigmento en forma de estrella; vesículas seminales con cabeza y tronco bien marcados, tocan las aletas posteriores, pero separadas de la aleta caudal; ovarios largos, huevos redondos, grandes y arreglados en una hilera. Especie epipelágica, su distribución es Indo-Pacífica, 40°N-40 S (Bieri 1959, Alvariño 1972, Pierrot-Bults & Nair 1991). Esta especie con tendencias neríticas, ha sido registrada ampliamente en aguas de Costa Rica (Morones 1988, Segura et al. 1992, Hossfeld 1996).
Ferosagitta robusta (Doncaster, 1902)
Fue la sexta especie en abundancia, registrándose con solamente 3 organismos, que corresponde al 1.79% del total de quetognatos. Mide hasta 12mm de longitud, presenta de 7 a 8 ganchos, de 6 a 9 dientes anteriores y de 10 a 15 posteriores, en cada lado. El cuerpo es fuerte, firme y opaco. Cabeza grande con cuello fuerte; las aletas anteriores y posteriores están completamente radiadas, las primeras poco más cortas. Collarete angosto, con divertículo intestinal conspicuo; ojos con pigmento en forma de T; vesículas seminales en dos partes: cabeza y tronco bien marcados; ovarios largos, huevos poco grandes arreglados en una hilera. Especie epipelágica, su distribución es Indo-Pacífica, 30°N-30°S (Bieri 1959, Alvariño 1972, Pierrot-Bults & Nair 1991). Esta especie con tendencias neríticas, ha sido registrada ampliamente en aguas de Costa Rica (Morones 1988, Segura et al. 1992, Hossfeld 1996).
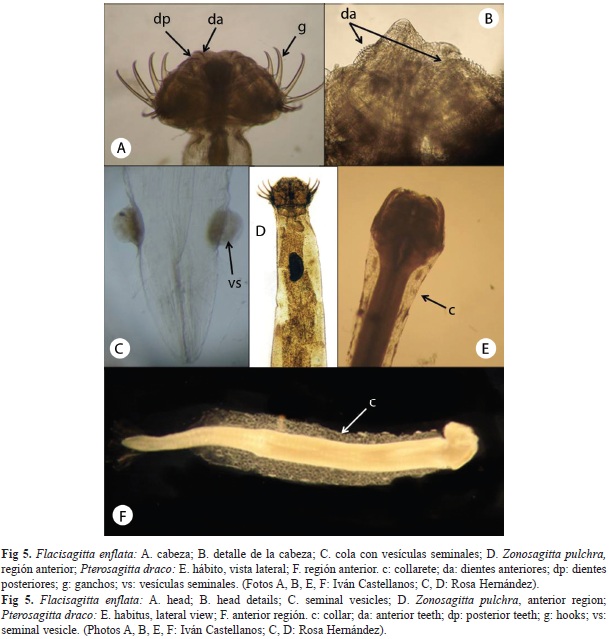
Flaccisagitta enflata (Grassi, 1881)
Fue la especie más frecuente y la más abundante, registrándose con 85 organismos, que corresponde al 50.6% del total de quetognatos, también tuvo la más amplia distribución en el área de estudio. Mide hasta 25mm de longitud, con 8 a 10 ganchos, 4 a 10 dientes anteriores y 4 a 15 posteriores en cada lado. El cuerpo es flácido, transparente y amplio hacia el centro. Cabeza pequeña con cuello bien marcado; las aletas anteriores son más cortas que las posteriores, ambos pares de aletas son redondeadas (Fig. 5a, b) y parcialmente radiadas. Ojos ovales, pequeños y con pigmento en forma de estrella; vesículas seminales esféricas, tocan la aleta caudal y bien separadas de las posteriores; ovarios cortos, huevos redondos arreglados en tres hileras (Fig. 5c). Especie epipelágica, su distribución es circumglobal, 40°N-40°S (Bieri 1959, Alvariño 1972, Pierrot-Bults & Nair 1991). Especie amplia y abundantemente registrada en aguas de Costa Rica (Morones 1988, Segura et al. 1992, Hossfeld 1996).
Serratosagitta pacifica (Tokioka, 1940)
Fue la segunda especie en abundancia, registrándose con 38 organismos, que corresponde al 22.62% del total de quetognatos; también tuvo amplia distribución en el área de estudio. Mide hasta 14mm de longitud, con 4 a 7 ganchos aserrados, 7 a 13 dientes anteriores y 16 a 24 posteriores, en cada lado. El cuerpo es translúcido, delgado y firme. Cabeza pequeña, con cuello bien marcado; las aletas anteriores son redondeadas, completamente radiadas (excepto por una pequeña región en la parte anterior), las aletas posteriores son angulares, casi totalmente radiadas. El collarete bien marcado en el cuello; ojos con pigmento en forma de T; vesículas seminales con cabeza y tronco bien conspicuos, presentan un escudo en la parte anterior con 5 a 10 dientes quitinosos; las vesículas tocan las aletas posteriores y están próximas a la aleta caudal; ovarios largos, huevos grandes arreglados en una hilera. Especie epipelágica, su distribución es Indo-Pacífica, 40°N-40°S (Bieri 1959, Alvariño 1972, Pierrot-Bults & Nair 1991). Esta especie con tendencias neríticas, ha sido registrada ampliamente en aguas de Costa Rica (Morones 1988, Segura et al. 1992, Hossfeld 1996).
Zonosagitta bedoti (Beraneck, 1895)
Fue la especie menos frecuente y abundante, con solamente 1 organismo, que corresponde al 0.6% del total de quetognatos. Mide hasta 15mm de longitud, con 6 a 8 ganchos, con 5 a 10 dientes anteriores y de 18 a 22 posteriores, en cada lado. El cuerpo es firme, opaco y amplio hacia el centro. Cabeza pequeña con cuello bien marcado; las aletas posteriores son triangulares y más cortas que las anteriores y sin radios en la parte anterior e interna. El collarete es corto o incipiente, ojos con pigmento en forma de estrella; vesículas seminales cónicas con una prominencia anterior que alcanza las aletas posteriores y caudal; huevos redondos o poliédricos, arreglados en tres hileras. Especie epipelágica, su distribución es Indo-Pacífica, 40°N-40°S (Bieri 1959, Alvariño 1972, Pierrot- Bults & Nair 1991). Esta especie con tendencias neríticas, ha sido registrada ampliamente en aguas de Costa Rica (Morones 1988, Segura et al. 1992, Hossfeld 1996).
Zonosagitta pulchra (Doncaster, 1902)
Fue la séptima especie en abundancia, registrándose con solamente 2 organismos, que corresponde al 1.19% del total de quetognatos, apareció únicamente en 2 estaciones de muestreo. Alcanza los 24mm de longitud, con 6 a 7 ganchos, de 8 a 10 dientes anteriores y con 8 a 16 posteriores, en cada lado. El cuerpo es delgado, firme y transparente. Cabeza pequeña con cuello marcado; las aletas anteriores son más largas que las posteriores, sin radios en la parte anterior e interna (Fig. 5d), las aletas posteriores son triangulares y radiadas en su parte externa. Collarete corto y conspicuo; ojos con pigmento en forma de estrella; vesículas seminales con cabeza y tronco angostos, próximas a las aletas posteriores y tocando la aleta caudal; los ovarios llegan al ganglio ventral, huevos redondos, pequeños, arreglados en dos hileras y en ocasiones una tercera hilera. Especie epipelágica, su distribución es Indo-Pacífica, 30°N-30°S (Bieri 1959, Alvariño 1972, Pierrot-Bults & Nair 1991). Esta especie con tendencias neríticas, ha sido registrada ampliamente en aguas de Costa Rica (Morones 1988, Segura et al. 1992, Hossfeld 1996).
Familia Pterosagittidae Tokioka, 1965
Pterosagitta draco (Krohn, 1853)
Fue la cuarta especie en abundancia, registrándose con 17 organismos, que corresponde al 10.12% del total de quetognatos. Especie que alcanza los 11mm de longitud, con 8 a 10 ganchos, 6 a 10 dientes anteriores y 8 a 18 posteriores en cada lado. El cuerpo es firme, opaco y amplio. Cabeza grande con cuello bien marcado; las aletas laterales son cortas, totalmente radiadas y localizadas sobre la cola. Collarete grande y voluminoso; ojos pequeños con pigmento en forma de T; las vesículas seminales son cónicas con una prominencia anterior (Fig. 5e, f), tocan las aletas laterales y próximas a la aleta caudal; ovarios largos, huevos redondos, arreglados en dos hileras. Especie epipelágica, su distribución es circumglobal, 40°N-40° S (Bieri 1959, Alvariño 1972, Pierrot-Bults & Nair 1991). Esta especie con tendencias neríticas, ha sido registrada ampliamente en aguas de Costa Rica (Morones 1988, Segura et al. 1992, Hossfeld 1996).
Agradecimientos
A Eduardo Suárez por sus valiosos comentarios y sugerencias que sirvieron para enriquecer este trabajo. Igualmente agradecemos el apoyo de la Vicerrectoría de Investigación de la Universidad de Costa Rica a través de los proyectos 808-3A-062 y 808-B0-060.
Referencias
Alvariño, A. 1972. Zooplancton del Caribe, Golfo de México y regiones adyacentes del Pacífico. Mem. IV. Congr. Nac. Ocean. México: 223-247. [ Links ]
Aravena, G. & S. Palma. 2002. Taxonomic identification of appendicularians collected in the epipelagic waters off northern Chile (Tunicata, Appendicularia). Rev. Chilena Hist. Nat. 75: 307-325 [ Links ]
Barham, E.G. 1979. Giant larvacean houses: observations from deep submersibles. Science 205: 1129-1131. [ Links ]
Bieri, R. 1959. The distribution of the planktonic Chaetognatha in the Pacific and their relationship to the water masses. Limnol. Oceanogr. 4: 1-28. [ Links ]
Bieri, R. 1991. Systematic of the Chaetognatha. XI, p. 122-136 In Q. Bone, H. Kapp & A.C. Pierrot-Bults (eds.). The Biology of Chaetognaths. Oxford Science Publications, New York, USA. [ Links ]
Casanova, J.P. 1999. Chaetognatha. p. 1353-1374 In D. Boltovskoy (ed.). South Atlantic Zooplankton. Backhuys Publ. Leiden, Holanda. [ Links ]
Castellanos-Osorio, I., A. Morales-Ramírez & E. Suárez- Morales. 2009. Appendicularians (Urochodata), p. 445-452. In I.S. Wehrtmann & J. Cortés (eds.). Marine Biodiversity of Costa Rica, Central America. Springer, Berlín. [ Links ]
Cortés, J. 2008. Historia de la investigación marina de la Isla del Coco, Costa Rica. . Rev. Biol. Trop. 56 (Supl. 2): 1-18. [ Links ]
Esnal, G.B. 1981. Apendicularia, p. 809-827 In D. Boltovskoy (ed.). Atlas del zooplancton del Atlántico sudoccidental y métodos de trabajo con el zooplancton marino. Pub. Esp. INIDEP, Mar del Plata, Argentina. [ Links ]
Esnal, G.B. 1996. Thaliacea y Apendicularia. XV, p. 597-630. In R. Gasca & E. Suárez (eds.). Introducción al Estudio del Zooplancton Marino. ECOSUR/CONACyT, Chetumal, Quintana Roo, México. [ Links ]
Esnal G. & R. Castro. 1977. Distributional and biometrical study of appendicularians from the west south Atlantic Ocean. Hydrobiologia 56: 241-246. [ Links ]
Fenaux, R. 1967. Les appendicularies des mers d’Europe et du Bassin Meditarraneén. Cent. Nat. Res. Sci. 2: 1-115. [ Links ]
Fenaux, R. 1993. The classification of Appendicularia (Tunicata): History and Current State. Mémories de l´Institut Oceanographique, Monaco, 17. Institut Océanographique, Monaco. 123 p. [ Links ]
Fenaux, R. 1998. The classification of Appendicularia. XVIII, p. 295-306. In Q. Bone (ed.). The Biology of the Pelagic Tunicates. Oxford Univ. Press, Londres. [ Links ]
Fernández-Álamo, M.A. & J. Färber-Lorda. 2006. Zooplankton and the oceanography of the eastern tropical Pacific: A review. Prog. Oceanogr. 69: 318-359. [ Links ]
Flores-Coto, C. 1965. Notas preliminares sobre la identificación de las apendicularias de las aguas veracruzanas. An. Inst. Biol. (México) 35: 293-296. [ Links ]
Gasca, R. 2009. Hyperiid amphipods, p. 275-282. In I.S. Wehrtmann & J. Cortés (eds.). Marine Biodiversity of Costa Rica, Central America. Springer, Berlín. [ Links ]
Hossfeld, B. 1996. Distribution and biomass of arrow worms (Chaetognatha) in Golfo de Nicoya and Golfo Dulce, Costa Rica. Rev. Biol. Trop. 44 (Supl. 3): 157-172. [ Links ]
Morales-Ramírez, A. 2008. Caracterización cualitativa del zooplancton del Área de Conservación Marina Isla del Coco (ACMIC), Océano Pacífico de Costa Rica. Rev. Biol. Trop. 56 (Supl. 2): 159-169. [ Links ]
Morones, L. 1988. Análisis cualitativo u cuantitativo de los quetognatos (Chaetognatha) en la región del Domo de Costa Rica (otoño, 1981). Tesis Profesional, Fac. Ciencias, UNAM, México. [ Links ]
Pierrot-Bults, A.C. 1996. Chaetognatha, p. 529-596. In R. Gasca & E. Suárez (eds.). Introducción al Estudio del Zooplancton Marino. El Colegio de la Frontera Sur (ECOSUR)/CONACYT, Chetumal, Quintana Roo, México. [ Links ]
Pierrot-Bults, A.C, & V.R. Nair. 1991. Distribution patterns in Chaetognatha, p. 86-116. In Q. Bone, H. Kapp & A.C. Pierrot-Bults (eds.). The Biology of Chaetognaths. Oxford Science Publ., New York, USA. [ Links ]
Raymont, J.E.G. 1983. Plankton and Productivity in the Oceans. Vol. 2. Zooplankton. Pergamon Press, Oxford, U.K. [ Links ]
Rodríguez-Sáenz, K. & R. Gasca. 2009. Siphonophores, p. 151-156. In I.S. Wehrtmann & J. Cortés (eds.). Marine Biodiversity of Costa Rica, Central America. Springer, Berlín. [ Links ]
Segura, P., R.M. Hernández & L. Morones. 1992. Distribución y abundancia de los quetognatos (Chaetognatha) en la región del domo de Costa Rica. Rev. Biol. Trop. 40: 35-42. [ Links ]
Suárez-Morales, E. & R. Gasca. 1989. Epiplanktonic calanoid copepods from the Costa Rica Dome (July-August, 1982). Cienc. Mar. 15: 89-102. [ Links ]
Suárez-Morales, E., R. M. Hernández-Flores & A. Morales. 2009. Chaetognaths or Arrow worms, p. 435-443. In I.S. Wehrtmann & J. Cortés (eds). Marine Biodiversity of Costa Rica, Central America. Springer, Berlín. [ Links ]
Tokioka, T. & J.A. Suárez-Caabro. 1956. Apendicularias de los mares cubanos. Mem. Soc. Cubana Hist. Nat. 23: 37-89. [ Links ]
Zoppi de Roa, E. 1971. Apendicularias de la región oriental de Venezuela. Stud. Fauna Curaçao Carib. Isl. 132: 77-109. [ Links ]
*Correspondencia a:
Iván Castellanos-Osorio. El Colegio de la Frontera Sur (ECOSUR) Unidad Chetumal, Av. Centenario km 5.5, A.P. 424, 77000, Chetumal, Quintana Roo, México; ivancast@ecosur.mx.
Rosa María Hernández-Flores. El Colegio de la Frontera Sur (ECOSUR) Unidad Chetumal, Av. Centenario km 5.5, A.P. 424, 77000, Chetumal, Quintana Roo, México; rosamahe@ecosur.mx.
Álvaro Morales-Ramírez. Centro de Investigación en Ciencias del Mar y Limnología (CIMAR), Universidad de Costa Rica, San Pedro, 11501-2060 San José, Costa Rica; alvaro.morales@ucr.ac.cr. Escuela de Biología, Universidad de Costa Rica, San Pedro, 11501-2060 San José, Costa Rica
Marco Corrales-Ugalde. Escuela de Biología, Universidad de Costa Rica, San Pedro, 11501-2060 San José, Costa Rica; mcugalde88@gmail.com.
1. El Colegio de la Frontera Sur (ECOSUR) Unidad Chetumal, Av. Centenario km 5.5, A.P. 424, 77000, Chetumal, Quintana Roo, México; ivancast@ecosur.mx; rosamahe@ecosur.mx
2. Centro de Investigación en Ciencias del Mar y Limnología (CIMAR), Universidad de Costa Rica, San Pedro, 11501-2060 San José, Costa Rica; alvaro.morales@ucr.ac.cr
3. Escuela de Biología, Universidad de Costa Rica, San Pedro, 11501-2060 San José, Costa Rica; mcugalde88@gmail.com
Recibido 09-III-2012. Corregido 05-VI-2012. Aceptado 24-IX-2012.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}