










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.60 suppl.1 San José Mar. 2012
Benthic and fish population monitoring associated with a marine protected area in the nearshore waters of Grenada, Eastern Caribbean
*Dirección para correspondencia
Abstract
Annual benthic and fish population surveys were completed at five locations in the nearshore waters along Grenada´s southwest coast during 2008-2010. Two survey sites are located in a newly launched Marine Protected Area (MPA). Photo Quadrat (PQ) and Point Line Intercept (PLI) surveys were used to determine substrate cover. Algae was the primary live cover increasing significantly from 45.9% in 2008 to 52.7% in 2010 (PLI). Algae was also predominant (61.0%-59.3%) in the PQ surveys although annual variation was not significant. Hard coral cover ranged from 16.5% to 15.4% (PLI) and 11.4% to12.0% (PQ) with no significant differences between years. Branching and encrusting corals occurred more frequently than massive corals. In the three annual surveys neither algal cover nor hard coral varied significantly between MPA and non-protected areas (PLI). Relative abundance of fishes along 30x2m belt transects did not vary significantly among years however density of fishes decreased significantly across years for most major groups. Chromis spp. dominated the survey sites at 65.2% in 2008 and 49.8% in 2010, followed by territorial damselfish,11.1% and 15.5%, wrasse increased from 7.3% to 15.5%. Both the substrate cover and fish survey data analyses indicated a stable but degraded community. Annual surveys are planned at these sites for the foreseeable future. Existing and future data from this project will be valuable in determining the efficacy of MPA management, guiding resource management decisions and monitoring the health status of Grenada’s valuable reef systems.
Key words: benthic cover, coral, reef fish, monitoring, Grenada, Eastern Caribbean, marine protected area.
Resumen
Un estudio sobre poblaciones bentónicas y de peces fue realizado en cinco localidades en la zona costera en el suroeste de Grenada entre 2008 y 2010. Dos sitios se ubicaron en una Área Marina Portegida (AMP) recientemente creada. Para determinar la cobertura se utilizaron foto-cuadrantes (FQ) y transectos de intersección de puntos (TIP). Las algas fueron el principal componente del bentos, aumentando significativamente de 45,9% en 2008 a 52,7% en 2010 (TIP). Las algas también fueron predominantes (61,9%-59,3) en los FQ, aunque las diferencias anuales no fueron significativas. La cobertura de corales pétreos tenía un ámbito de 16,5% a 15,4% (TIP) y de 11,4% a 12,0% (FQ), sin diferencias significativas entre años. Los corales ramificados e incrustantes fueron más frecuentes que los corales masivos. En los tres años no hubo diferencias significativas entre las AMPs y las áreas no protegidas. La abundancia relativa de peces a lo largo de un transecto de 30x2m no varió significativamente entre los años, sin embargo, la densidad de peces decreció significativamente a través de los años, para los grupos principales. Chromis spp. predominó con 65,2% en 2008 y 49,8% en 2012, seguido por damiselas territoriales, 11,1% y 15,5%, y los lábridos aumentaron de 7,3% a 15,5%. Tanto la coberura del sustrato como los datos de peces indican una comunidad estable pero degradada. Sondeos anuales están planeados para el futuro. Los datos existentes y futuros de este proyecto serán muy útiles para determinar la eficacia de la gestión de las AMPs y el estado de salud de los sistemas arrecifales de Grenada.
Palabras clave: cobertura bentónica, coral, peces de arrecife, monitoreo, Grenada, Caribe Oriental.
The island nation of Grenada is part of the Eastern Caribbean region recently classified as being at “very high risk” by the Reefs at Risk in the Caribbean report (Bouchon et al. 2008). Of the 160 km2 of reef area in Grenada 41% were listed as having a high-risk threat index and 40% were listed as very high (Burke & Maidens 2004). The primary contributors to this rating were coastal development and fishing pressure.
Coral communities rely on large herbivorous fish species to manage levels of macroalgae (Burkepile & Hay 2010, Ceccarelli et al. 2011, Walsh 2011). In an analysis of the Grenadian demersal fish catch and fishing effort from 1986 to 1993, Jeffrey (2000) found that the number of boats employed in the demersal fishery off the west coast of Grenada increased by 200% however the catch declined by nearly 75% during this seven year period. Local overfishing often targets large herbivorous species reducing these fish stocks thus contributing to increased abundance of macroalgae on coral reefs (Hawkins & Roberts 2003). One impact of increased algal abundance is reduced growth and recruitment of coral polyps (Bascompte et al. 2005, Arnold 2007, Birrell et al. 2008, Mora 2008).
Introduction of excess nutrients to coral reef systems from coastal development further enhances overgrowth of algae (Lapointe et al.1997) and directly inhibits coral recruitment and growth (Littler et al. 2009). These local stressors weaken a coral community’s resilience (Hughes 1994, Hughes et al. 2003, Gardner et al. 2003, Wilkinson 2008) making it more vulnerable to global climate change and increased storm activities (Goldenberg et al. 2001, Eakin et al. 2010, Hughes et al. 2010). Grenada has been impacted by two major hurricanes in the past decade: Ivan in September 2004 and Emily in July 2005. Major storms such as these can result in devastating effects on reefs breaking down the basic structure and dislodging corals leaving leveled areas of rubble (Woodley et al. 1981).
Many countries have established Marine Protected Areas (MPAs) that restrict some uses of coastal reef systems with the hope that these sections will provide a source of biodiversity to adjacent or “down current” locations. Unfortunately, there is a paucity of data that demonstrates the effectiveness of specific management practices. While the Grenadian government established legislation for the Moliniere-Beausejour MPA on the southwest coast of the island in 2001, no significant management practices were implemented until 2010. Permanent mooring buoys were established in 2009, warden patrols began in 2010 and some fishing practices were restricted from September 2010.
A development plan for Grenada’s National Protected Areas System identified the need for external assistance in research and monitoring of Grenada’s protected areas (Mac Leod 2007). Initial surveys at nine sites off the southwest coast of Grenada in 2006 and 2007 identified macroalgae was the most abundant substrate cover ranging from 36.5% (± 0.8%) to 53.2 % (± 1.2%). Hard corals covered 23.8% (± 0.9%) to 38.1% (± 1.2%) (Bouchon et al. 2008). This 2008-2010 study builds on the initial survey and establishes a foundation upon which the effectiveness of the Moliniere-Beausejour MPA management techniques may be evaluated.
Study Area: Five sites ranging in depth from 5.2m-12.2m, located along Grenada’s southwest coast were established in 2008. Similar reefs both inside and outside the MPA that are frequently used by the dive industry were selected. Dragon Bay (12° 5’6.00”N 61°45’45.36”W) and Flamingo Bay (12° 5’30.36”N 61°45’30.60”W) are in MPAs, while Northern Exposure Shallow (12° 1’57.30”N 61°46’14.28”W), Northern Exposure Deep (12° 2’22.14”N 61°46’4.74”W) and Quarter Wreck (12° 1’40.98”N 61°47’0.84”W) are in non-protected areas. Four 30m parallel transects were set up at 5m intervals. Metal stakes mark the beginning and end of each permanent transect.
Materials and Methods
The substrate composition of Grenada’s southwest coast was surveyed with the Photo Quadrat (PQ) and Point Line Intercept (PLI) methods. The PQ method allows for careful identification of substrate types from a digital photograph. Although identification of substrate types is not always optimal based on digital photos this approach allowed more intense scrutiny of the substrate since time is not a factor in the sampling process. In addition using Coral Point Count with Excel extensions (CPCe) v.3.6 allowed a randomized sampling scheme for each picture (Kohler & Gill 2006). Since there are 60 pictures associated with each transect this increases the total number of observations. The PLI method developed by Crosby & Bruckner in 2002 based on Crosby & Reese (1996) was used to estimate relative abundance of major types of substrate cover and fish species associated with the coral reefs in Grenada’s nearshore waters. Four 30m permanent transects were surveyed at each of the five locations. Fish species, as well as Diadema antillarum, abundance occurring within a twometer wide belt (AGRRA Protocol v. 4.0) from the substrate to water surface along each transect were recorded. Benthic substrate was identified and recorded at a point directly below each half-meter mark along each transect.
Divers completed fish data collections along each transect in ten minutes.
Sixty photo quadrats from each transect were processed using Coral Point Count with Excel extensions (CPCe) v.3.6 (Kohler & Gill 2006). A Canon EOS Digital Rebel XTI camera in an Ikelite underwater housing was used to take a picture at every half-meter mark. Attached to the underwater housing was a tube with a calibrated scale used to maintain a consistent distance (60cm) from the substrate and to assist with scaling in the CPCe software program. The images were uploaded into the CPCe software program, and a 20cm by 20cm square was superimposed on the image. Eight points were randomly generated inside the square, and the substrate under each point was identified. Dumas et al. (2009) found that whether nine or ninety-nine points were used in a1m2 area, the difference for large categories was not significant. Thus for the 400 cm2 area in this study 8 points were deemed sufficient. Also, a Sony HDR-SR8 video camera in an Amphibico underwater housing was used to take video of each location to record a broader perspective of the survey sites.
For both the PLI and PQ data a repeated measure analysis of variance (ANOVAR) using transects as the sampling unit was used to monitor Grenada’s southwest benthic community and fish assemblage from 2008 to 2010. To satisfy the assumption of normality, proportional data was arcsine square root transformed and all non-proportional data was log transformed. The Shapiro-Wilk test, as well as skewness and kurtosis values were used to assess normality. Non-normal distributions were examined, and if appropriate outliers were removed (Zar 1999). Mauchly’s sphericity test was used to determine sphericity, if violated the Greenhouse-Geisser correction was used to determine significance. Additionally all cases of significance were verified with the multivariate analyses, which do not assume sphericity. A Bonferroni correction (significance value (0.05)/ number of comparisons made) was used when determining significance (Sokal and Rohlf 1995). Also, the Bonferroni correction multiple comparison test was used to determine among which years a significant difference occurred.
To identify interactions between the MPA and non-protected area from 2008 to 2010 a two-way ANOVAR was used. This was only done for the PLI data, because the PQ data had an insufficient sample size. In order to effectively make this comparison the same sample size needed to be used for the MPA and non-protected area. This was accomplished by selecting two of the three non-protected locations, Quarter Wreck and Northern Exposure Shallow. The above criteria for assessing normality, sphericity, and significance were used. When an interaction was found to be significant a follow up one-way analysis of variance (ANOVA) test was used to further examine the interaction. The same criteria for assessing normality were used, and Levene’s test of homogeneity was used to evaluate the equality of variances. If the results of Levene’s test were found to be significant, then a p<0.01 was used to determine significance. The Bonferroni correction was still used when determining significance as well (Sokal and Rohlf 1995).
Results
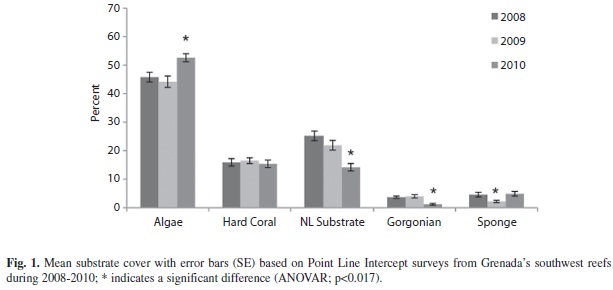
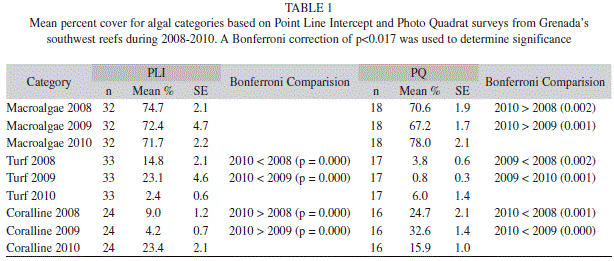
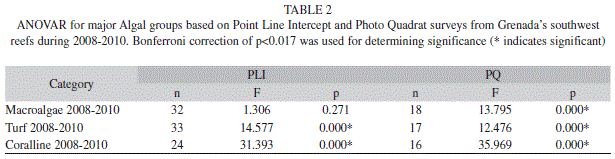
Substrate (PLI): Algae was the dominant substrate cover found at all locations off Grenada’s southwest coast (Fig. 1). Algae increased significantly from 45.9% (SE=1.7; n=35) in 2008 to 52.7% (1.4; 35) in 2010 (ANOVAR, F=7.431, p=0.001). Comparison of major algal groups (macroalgae, turf and coralline) showed that macroalgae consistently dominated the algal community. Turf algae decreased significantly in 2010 and coralline algae increased significantly in 2010 (Tables 1 & 2).
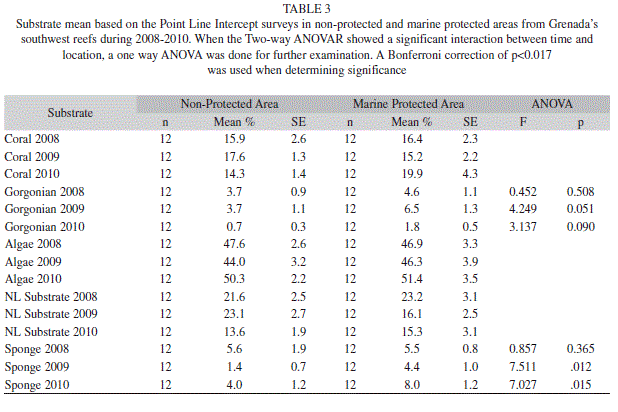
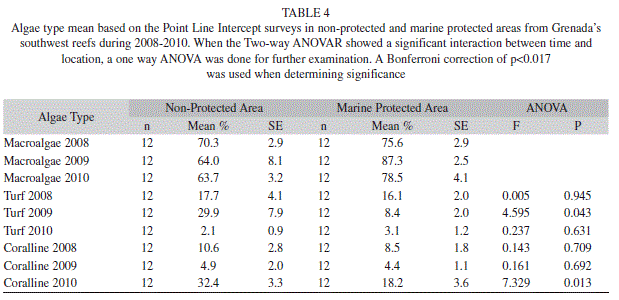
Algal cover in the MPA ranged from 46.3% (3.9; 12) to 51.4% (3.5; 12) over the three years, while in the non-protected area it ranged from 44.0% (3.2; 12) to 50.3% (2.2; 12); no significant interaction between time and location was found (Two-way ANOVAR, F=1.528, p=0.239) (Table 3). Yet the different types of algae experienced significant interaction. Turf algae did have a significant interaction (Two-way ANOVAR, F=6.738, p=0.005), but the follow up tests showed no significant differences between the MPA and non-protected area. Coralline algae also exhibited a significant interaction (Two-way ANOVAR, F=17.752, p=0.000), and for 2010 the 32.4% (3.3; 12) found in the non-protected area was significantly greater than the 18.2% (3.6; 12) in the MPA. Macroalgae did not exhibit any significant interaction (Two-way ANOVAR, F=1.581, p=0.237) (Table 4).
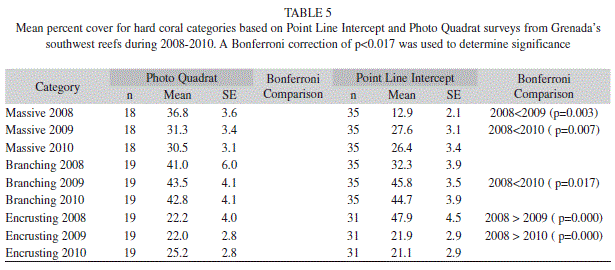
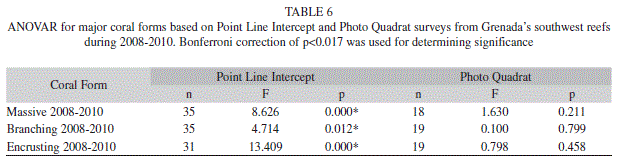
The hard coral cover did not vary significantly from year to year ranging from 16.5% (1.0; 35) to 15.4% (1.3; 35) (Fig. 1) (ANOVAR, F=0.531, p=0.591) (Fig. 1). However the type of hard coral (massive, encrusting and branching) observed did change across the years. Encrusting coral occurred most frequently in 2008 however by 2010 branching coral occurred most frequently (Tables 5 & 6).
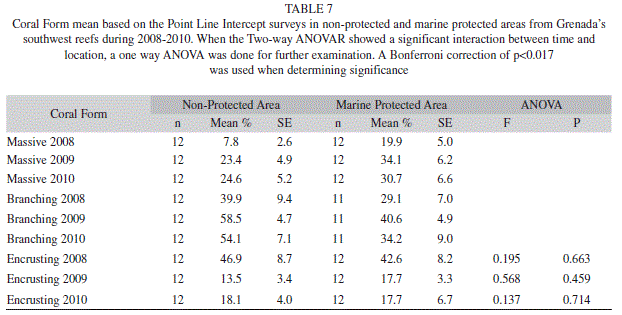
Hard coral cover ranged from 15.2% (2.2; 12) to 19.9% (4.3; 12) in the MPA, and 14.3% (1.4; 12) to 17.6% (1.3; 12) in the non-protected area (Table 3). Although hard coral did not differ significantly (Two-way ANOVAR, F=0.072, p=0.931) in overall percent between the MPA and non-protected area encrusting coral did have a significant interaction between time and location (Two-way ANOVAR, F=7.049, p=0.004). Yet in follow up analyses no significant differences between the MPA and non-protected area was found. Massive (Two-way ANOVAR, F=3.555, p=0.046) and branching (Two-way ANOVAR, F=3.170, p=0.091) coral had no significant interaction between time and location (Table 7).
While hard coral cover remained stable, gorgonian cover significantly dropped from 3.7% (0.4; 32) and 4.0% (0.6; 32) in 2008 and 2009 to 1.8% (0.3; 32) in 2010 (ANOVAR, F=19.609, p=0.000).
Other significant changes in the substrate were seen in the sponge and non-living categories. Sponge cover saw a sudden decrease from 4.6% (0.8; 35) in 2008 to 2.2% (0.4; 35) in 2009, but recovered to 4.9% (0.8; 35) in 2010 (ANOVAR, F=6.212, p=0.005). Also non-living substrate significantly decreased from 25.2% (1.7; 35) and 21.9% (1.9; 35) in 2008 and 2009 to 14.2% (1.3; 35) in 2010 (ANOVAR, F=14.745, p=0.000) (Fig. 1).
Further comparisons of percent cover in the MPA to non-protected areas revealed that gorgonian cover did have a significant interaction (Two-way ANOVAR, F=13.005, p=0.000). Additional tests of gorgonian cover in the MPA and non-protected areas showed no significant difference among years. Sponge cover also exhibited a significant interaction (Two-way ANOVAR, F=8.654, p=0.002). The sponge cover in the MPA was not significantly different from the non-protected area in 2008; however sponge cover in 2009 and 2010 showed that the MPA sponge cover was significantly greater than the non-protected area (Table 3).
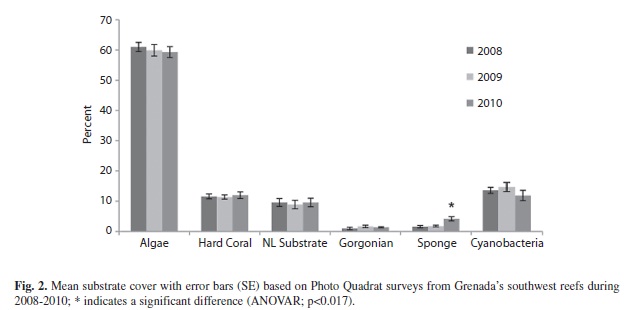
Substrate (Photo Quadrat): Algae, the dominant substrate cover, ranging from 61.0% (1.5; 19) in 2008, 59.9% (1.9; 19) in 2009 and 59.3% (1.8, 19) in 2010 showed no significant annual differences (ANOVAR, F=0.373, p=0.616) (Fig. 2). Although the percent cover of algae did not change across years, the type of algae observed did. Macroalgae which occurred more frequently than other types of algae increased significantly in 2010. Turf algae dipped significantly in 2009 and coralline algae decreased significantly in 2010 (Table 1 & 2).
Percent hard coral cover remained stable across years ranging from 11.4% (0.7; 18) to 12.0% (1.1; 18) (ANOVAR, F=0.037, p=0.964). Of the three hard coral forms recorded branching coral occurred most frequently with no significant annual variation (Table 3 & 4). Cyanobacteria which was not recorded over the three year sampling period with the PLI method was similar in percent cover to hard coral ranging from 14.7% (1.5; 19) to 11.9% (1.7; 19) (ANOVAR, F=1.314, p=0.277). Percent sponge cover did increase significantly from 1.6% (0.4; 19) in 2008 to 4.2% (0.7; 19) in 2010 (ANOVAR, F=9.478, p=0.002, Bonferroni p=0.004) (Fig. 2).
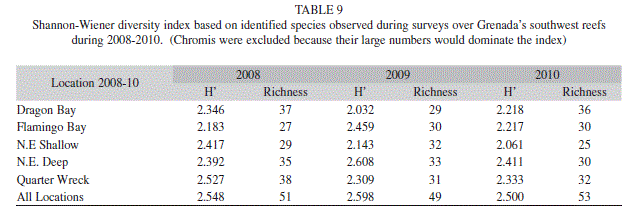
Fish: A total of 62 fish species were observed at the five sampling locations from 2008 to 2010 (Table 8). The major groups of fish analyzed included Chromis spp., damselfishes, parrotfishes, surgeonfishes, and wrasse. Chromis spp. were separated from the damselfishes because of their large number. Diversity indices were quite high and similar across all sites (Table 9).
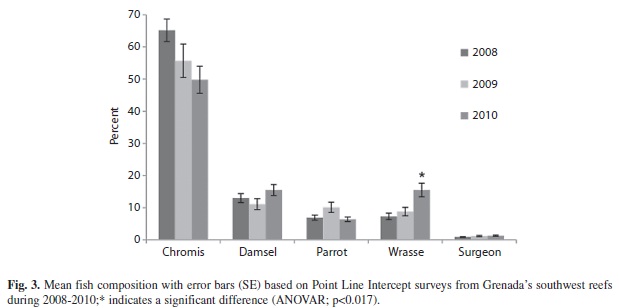
Relative abundance of all but one of the most frequently occurring groups of fishes did not vary significantly across the three years of this study. Chromis spp., the largest group observed, showed a downward trend going from 65.2% (3.5; 34) to 49.8% (4.2; 34); however the difference was not significant (ANOVAR, F=3.611, p=0.032). Damselfishes ranged from 11.1% (1.7; 32) to 15.5% (1.7; 32) (ANOVAR, F=3.531, p=0.035) and parrotfishes from 10.1% (1.6; 36) to 6.4% (0.7; 36) (ANOVAR, F=1.732, p=0.184) (Fig. 3). Surgeonfishes also remained stable between 0.9% (0.1; 31) and 1.3% (0.2; 31) (ANOVAR, F=0.146, p=0.864). Wrasse however showed a significant increase from 7.3% (1.0; 35) to 15.5% (2.1; 35) (ANOVAR, F=7.341, p=0.001) (Fig. 3).
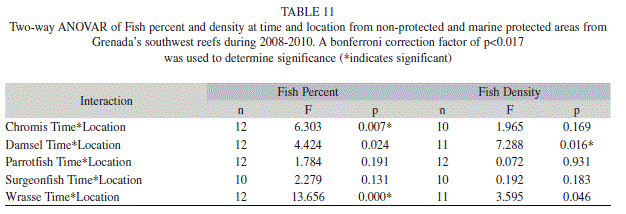
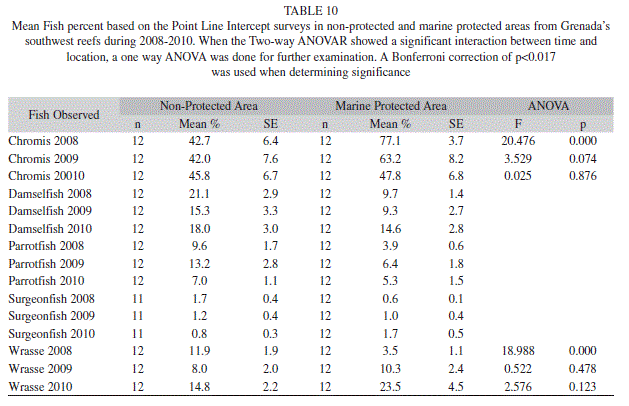
In comparing the MPA to the non-protected area, a significant interaction between time and location was observed for the chromis, which ranged from 47.8% (6.8; 12) to 77.1% (3.7; 12) in the MPA and 42.0% (6.4; 12) to 45.8% (6.7; 12) in the non-protected area (Two-way ANOVAR, F=6.303, p=0.007). Additional tests revealed that percent chromis observed in 2008 was significantly lower in the nonprotected area then the MPA (Table 10). The wrasse group also had a significant interaction between time and location (Table 11). During 2008 the wrasse were significantly higher in the non-protected area at 11.9% (1.9; 12), whereas the MPA only had 3.5% (1.1; 12) wrasse (Table 10). None of the other fish groups showed a significant interaction at time and location (Table 11).
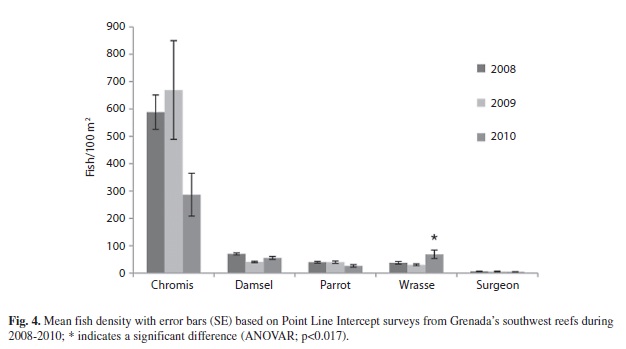
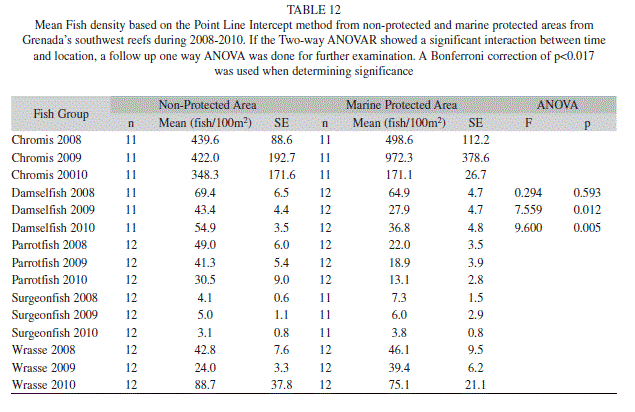
The density of fishes on the other hand did show significant differences for most groups over the years of the study. Chromis spp. decreased significantly from 669.3 fish/100m2 (180.5; 30) to 286.6 fish/100m2 (78.3; 30) (ANOVAR, F=9.215, p=0.000). Damselfishes density also significantly decreased from 70.3 fish/100m2 (3.7; 34) in 2008 to 40.6 fish/100m2 (2.7; 34) in 2009 (Bonferroni, p=0.000) and 55.3 fish/100m2 (5.2; 34) in 2010 (Bonferroni, p=0.015) (ANOVAR, F=17.994, p=0.000). The density of parrotfishes significantly decreased from 39.5 fish/100m2 (3.2; 30) and 39.7 fish/100m2 (4.2; 30) in 2008 and 2009 to 26.3 fish/100m2 (4.5; 30) in 2010 (ANOVAR, F=10.786, p=0.000). Wrasse density however showed an increase from 37.6 fish/100m2 (4.6; 34) and 30.2 fish/100m2 (3.5; 34) in 2008 and 2009 to 68.6 fish/100m2 (15.4) in 2010 but the change was not significant (ANOVAR, F=3.525, p=0.035). The density of surgeonfishes did not significantly change, however it showed a downward trend going from 6.3 fish/100m2 (0.8; 25) in 2008 to 5.9 fish/100m2 (1.4; 25) in 2009, and finally to 4.5 fish/100m2 (0.5; 25) in 2010 (ANOVAR, F=1.859, p=0.179) (Fig. 4). The only fish group that experienced a significant interaction between time and location for density was damselfish (Two-way ANOVAR, F=7.288, p=0.016) (Table 11). Damselfish in 2010 were significantly higher in the non-protected area at 54.9% (3.5; 11), while only 36.8% (4.8; 12) were observed in the MPA (ANOVA, F=9.600, p=0.005) (Table 12).
The observed fish assemblage was divided into feeding groups based on the classification of Sadin (2008b). Combined data from all sites across years (2008-2010) showed the dominant feeding group of the assemblage to be planktivores at 81.3% (1.8%; 35) to 74.7 (3.5%; 31). Herbivores represented 9.7% (1.0%; 35) to 13.6 (2.0%; 36), while carnivores comprised 10.9 (1.3%; 30) to 14.9% (2.2; 34). Fish feeding groups were not significantly different between years. In addition there was no significant difference between the MPA and non-protected areas in the percent planktivores (Two-way ANOVAR, F=3.891, p=0.036), herbivores (Two-way ANOVAR, F=2.900, p=0.078) or carnivores (Two-way ANOVAR, F=2.490, p=0.111) since none exhibited a significant interaction between time and location.
Combined Diadema antillarum density for Grenada’s southwest coast exhibited a significant downward trend having 3.1 urchins/100m2 (0.5; 36) in 2008, to 1.9 urchins/100m2 (0.5; 36) in 2009 and to only 0.2 urchins/100m2 (0.1; 36) in 2010 (ANOVAR, F=6.078, p=0.004). It should be noted even after log transformation the data did not fulfill the assumption of normality, however sphericity could be assumed. There was also no significant interaction at time and location for diadema (Two-Way ANOVAR, F=1.853, p=0.197).
Discussion
Data collected during three annual surveys indicates benthic cover in the nearshore waters off the southwest coast of Grenada was similar to many reported findings from across the Caribbean. Algae dominated the substrate (45.9% to 61.0%) and live hard coral coverage (16.5% to 11.4%) was quite low. Algae dominated systems have been reported for many nearshore communities in the Caribbean (Hughes 1994, Gardner 2003, Burke & Maidens 2004, Bouchon et al. 2008, Wilkinson 2008, Mumby 2009, Walsh 2011). Algae has been the dominant substrate cover on Caribbean reefs since a major ecological phase shift occurred in the 1980s. Overfishing, hurricane damage and a disease-induced die-off of D. antillarum have been proposed as major factors in this shift (Hughes 1994, Gardner et al. 2003).
Low densities of D. antillarum in Grenadian nearshore waters may be one of the key factors in the high algal component of this benthic community. The mean D. antillarum density found in 2x30m belt transects off the coast of Grenada during this study ranged from 0.002/m2 to 0.031/m2 which is much lower than the 4.25/m2 densities measured in 2003 by Carpenter & Edmunds (2006) for these waters and the 1.7-8.9/m2 they found associated with reefs of six countries around the Caribbean. Based on general surveys across a spectrum of western Atlantic reefs between 1998-2000 Kramer (2003) reported mean D. antillarum densities of 0.029/m2. Newman et al. (2006) found mean densities of 0.019/m2 at similar depths in the western and northern Caribbean. In both studies fleshy macroalgae generally dominated the reef benthic communities where these low D. antillarum densities occurred.
Given the importance of D. antillarum in the coral reef community reestablishment of D. antillarum may have potential as a management tool to enhance coral growth in algal dominated systems. This potential became apparent when a phase shift reversal was noted on Jamaica’s north coast. Coral cover increased from 23% in 1995 to 54% in 2004 with higher growth rates of juvenile corals and higher densities of small juvenile recruits in “dense urchin zones” (Idjadi et al. 2006, 2010, Bechtel et al. 2006). The potential impact of increased numbers of D. antillarum on coral recovery sparked introductions of additional D. antillarum into Grenada’s MPA from adjacent populations in 2011 (Nimrod personal communication). These relocations will hopefully result in significant increases in local populations of D. antillarum that will reduce macroalgae and facilitate an increase in coral recruitment and growth.
Understanding the composition of Grenada’s southwest coastal nearshore fish community will also inform existing and future fisheries management practices. Heavy fishing pressure has been identified as one of the key factors in transformation of coral reefs to algal dominated systems (Hawkins & Roberts 2003). Fishing methods in Grenada include beach seining, trap nets, hand lines and spearing. Target species are mainly carnivores such as Lutjanidae (snapper), Serranidae (groupers) and Carangidae (jacks, pompanos and mackerels and scad) (Finlay 2000). Large herbivores such as Scaridae (parrotfish) and Acanthuridae (surgeonfishes and tang) are also frequently seen in Grenadian fish markets. Observations during 2008-2010 indicated that planktivores (74.7-81.3%) dominated the nearshore Grenadian fish community followed by herbivores (9.7-13.6%), carnivores (10.9-13.9%). The herbivore component of Grenada’s nearshore fish assemblage seems low when compared to other studies. Toller et al. (2010) found 65% herbivores off Saba Island in habitat types similar to those in Grenada. In a synthesis of Caribbean wide surveys between 1998 and 2000, Kramer (2003) found herbivores made up 64.6% of the fish community sampled. Simply determining that the herbivore component of the Grenadian fish community is low compared to other locations does not allow a full understanding of the impact this has on substrate cover. Burkepile & Hay (2010) pointed out the importance of species level identification in reef fish monitoring. Each herbivorous species can have unique impacts on algal succession and coral growth. A diverse assemblage of herbivorous fishes can reduce development of macroalgae communities and thereby enhance recruitment of coral to open substrates. Ceccarelli et al. (2011) divides herbivorous fishes into roving herbivores or “foragers” (parrotfish Scarus spp. and surgeonfish Acanthurus spp.) and “farmers” (territorial damselfish Stegastes spp.) in order to evaluate their potential influence on algal succession and coral reef recovery. “Farmers” tend to suppress algal succession preventing development of the fleshy macroalgae stage. “Foragers” have an intermediate effect allowing development of some macroalgae but not a late-successional assemblage (Ceccarelli et al. 2011). In Grenada’s nearshore fish community herbivores were dominated by parrotfishes (Scaridae) at 70.2% followed by territorial damselfish fish (Pomacentrus spp., Stegastes spp., Microspathodon spp.) at 17.9% and surgeonfish (Acanthurus spp.) at 11.5%. Thus “farmers” comprised only 17.9% in the Grenadian nearshore herbivorous fish community while “foragers” made up 82.1%. It is understandable therefore that turf algae comprised such a small portion of the algal community and fleshy macroalgae made up the majority. Arnold (2007) demonstrated that grazing by scrapers such as parrotfish and urchins facilitate coral recruitment more than territorial damselfishes that maintain low levels of turf algae. Since the species composition of herbivores in Grenada’s nearshore waters is primarily comprised of “foragers” rather than “farmers” potential benefits are likely for future coral recruitment if the overall number of herbivores can be increased. It is hoped that newly implemented fishing restrictions in the Moliniere-Beausejour MPA will facilitate increased abundance of herbivorous fishes.
In addition to low numbers of herbivores, algal dominance is also driven by increases in nutrients in nearshore waters. Littler et al. (2009) described the importance of taking into consideration the complex interaction of herbivory, nutrient levels and stochastic events in understanding existing conditions and developing management strategies for coral reef communities. Lapointe et al. (1997) argued that nutrient input from non-point source pollution related to development and population increases on the island of Jamaica was a major factor in driving the shift from a coral dominated system to an algal dominated community. In a comparative study of reef communities Sandin et al. (2008a) saw a shift from dominance by a few large top predator fish species to dominance by small lower trophic level consumers, primarily planktivores, in areas of increasing human populations. The dominance of planktivores (primarily Chromis spp.) in Grenadian nearshore waters may be an indicator of excess nutrients into these waters. Two major rivers (St. Johns and Beausejour), flow into the nearshore waters of Grenada’s southwest coast. These rivers drain heavily populated areas as well as agricultural lands and have the potential of delivering excess nutrients into the reef communities. These nutrients have the potential of enhancing macroalgal growth and inhibiting the recruitment and growth of coral (Littler et al. 2009).
The three years of monitoring at permanent transects in this study provide a basis for future trend analysis and evaluation of management practices. Hughes et al. (2010) advocates long term monitoring of important taxonomic groups as well as identification of mechanisms and feedbacks in order to detect indicators of phase shifts. He also encourages agencies involved in research and management of reefs to take a proactive integrative approach through education of grassroots constituencies, enhancing access to existing information and expertise and strengthening regulations associated with harvesting important species from these communities. This approach is beginning to be implemented in Grenada through the work of the Grenada Government Fisheries Division and the Moliniere-Beausejour Marine Protected Area Stakeholder Group.
This study establishes a baseline of information but long term and more specific monitoring is needed to better understand the trajectory of Grenada’s reef communities. Gardner et al. (2003) indicated that areas of coral recovery are often dominated by non-framework builders such as Agaricia and Porites rather than framework builders such as Acropora and Montastrea. These framework builders that formerly dominated reefs in the Caribbean are essential to surviving the destructive forces of major storms. The coral community in Grenada’s nearshore waters is comprised primarily of branching coral much of which is Agaricia and Porites. Given the importance of framework builders to the resilience of coral reef communities identification of coral species will be added to the monitoring program to better understand the coral community.
The similarity between the MPA and non-protected areas seen in this study may be due to the fact that the Moliniere-Beausejour MPA management plan was not fully implemented until September 2010. After full implementation of the plan wardens began to patrol the area and prevent fishing from boats and enforce the use of permanent mooring buoys by divers and snorkelers in the MPA. Future monitoring efforts will be able to use the results of this study as a basis for comparison in order to assess the impact of the newly implemented management practices in the MPA. Expansion of current studies will allow a better understanding of mechanisms and feedbacks in these reef systems. Video and photographs of transects and surrounding habitats are being incorporated into public presentations for Grenadian resource managers and the general public to encourage a broader understanding of the importance of careful resource management.
In addition to focusing on local environments it is important to connect these studies to broader ecosystem wide analyses. Ogden (2010) encourages moving toward an ecosystem-based management plan for the Caribbean. Ogden cites plans for regional management inspired by the CARICOMP network of marine laboratories and encourages going beyond local problems and addressing issues like the D. antillarum die-off, wide spread white band disease and the annual plume of discharge from Venezuela’s Orinoco River. Efforts are ongoing to strengthen connections of this ongoing monitoring effort to the network of Caribbean marine laboratories and provide information that will assist regional management.
Acknowledgements
Funding for this project was provided by the Fischer Family Foundation and Mr. Gary Stimac and is greatly appreciated. A special thanks to Jacob Krause for playing a major role in developing this program. Thanks are also offered to Jillian Groeschel, Kyle Foster, Svetlana Bornschlegl, Victoria Krueger, Thomas Dietrich, Ben Hermanson, Laurelyn Dexter, Angela Majeskie, Emily Bolda, Allison Page, and Angela Blasezyk for data collection. Kayli Giertych and Stephen Vandenberg are thanked for assistance with the manuscript. Billie Harrison of the Racine Zoo is also thanked for field assistance.
References
Arnold, S.N. 2007. Running the gauntlet to coral recruitment through a sequence of local multiscale processes. MSc Thesis, Univ. Maine, Orono, Maine, USA. [ Links ]
Bascompte, J., C.J. Melian & E. Sala. 2005. Interaction strength combinations and the overfishing of a marine food web. Proc. Natl. Acad. Sci. USA 102: 5443-5447. [ Links ]
Bechtel, J.D., P. Gayle & L. Kaufman. 2006. The return of Diadema antillarum to Discovery Bay: patterns of distribution and abundance. Proc. 10th Int. Coral Reef Symp., Okinawa 1: 367-375. [ Links ]
Birrell, C.L., L. J. McCook, B.L. Willis & G.A. DiazPulido. 2008. Effects of benthic algae on the replenishment of corals and the implications for the resilience of coral reefs. Oceanogr. Mar. Biol. Annu. Rev. 46: 25-63. [ Links ]
Bouchon, C., P. Portillo, Y. Bouchon-Navaro, M. Louis, P. Hoetjes, K. De Meyer, D. Macrae, H. Armstrong, V. Datadin, S. Harding, J. Mallela, R. Parkinson, J. van Bochove, S. Wynne, D. Lirman, J. Herlan, A. Baker, L. Collado, S. Nimrod, J. Mitchell, C. Morrall, C. Isaac. 2008. Chapter 19. Status of coral reefs of the Lesser Antilles: The French West Indies, The Netherlands Antilles, Anguilla, Grenada, Trinidad and Tobago, p. 265-279 In: Wilkinson, C. (Ed.). Status of coral reefs of the world 2008. Global Coral Reef Monitoring and Reef and Rainforest Research Centre, Townsville, Australia. [ Links ]
Burke, L. & J. Maidens. 2004. Reefs at Risk in the Caribbean. World Resources Institute, Washington D.C., USA. [ Links ]
Burke, L., K. Reytar, M. Spalding & A. Perry. 2011. Reefs at Risk Revisited. World Resource Institute, Washington, D.C., USA. [ Links ]
Burkepile, D.E. & M.E. Hay. 2010. Impact of herbivore identity on algal succession and coral growth on a Caribbean reef. PLoS ONE 5: e8963. [ Links ]
Carpenter, R.C. & P.J. Edmunds. 2006. Local and regional scale recovery of Diadema promotes recruitment of scleractinian corals. Ecol. Lett. 9: 268-277. [ Links ]
Ceccarelli, D.M., G.P. Jones & L.J. McCook. 2011. Interactions between herbivorous fish guilds and their influence on algal succession on a coastal coral reef. J. Exp. Mar. Biol. Ecol. 399: 60-67. [ Links ]
Crosby, M.P. & E. S. Reese. 1996. A manual for monitoring coral reefs with indicator species: butterfly fishes as indicators of change on the Indo-Pacific reefs, Office of Ocean and Coastal Resource Management, NOAA, Silver Spring, Maryland, USA. [ Links ]
Dumas, P., A. Bertaud, C. Peignon, M. Leopold & D. Pelletier. 2009. A “quick and clean” photographic method for the description of coral reef habitats. J. Exp. Mar. Biol. Ecol. 368:161-168. [ Links ]
Eakin, C.M., J.A. Morgan, S.F. Heron, T.B. Smith, G. Liu, L. Alvarez-Filip, B. Baca, E. Bartels, C. Bastidas, C. Bouchon, M. Brandt, A. Bruckner, L. Bunkley-Williams, A. Cameron, B.D. Causey, M. Chiappone, T.R.L. Christensen, M.J.C. Crabbe, O. Day, E. de la Guardia, G. Díaz-Pulido, D. DiResta, D.L. Gil-Agudelo, D. Gilliam, R. Ginsburg, S. Gore, H.M. Guzman, J.C. Hendee, E.A. Hernández-Delgado, E. Husain, C.F.G. Jeffrey, R.J. Jones, E. Jordán-Dahlgren, L. Kaufman, D.I. Kline, P. Kramer, J.C. Lang, D. Lirman, J. Mallela, C. Manfrino, J.P. Maréchal, K. Marks, J. Mihaly, W.J. Miller, E.M. Mueller, E. Muller, C.A. Orozco-Toro, H.A. Oxenford, D. Ponce-Taylor, N. Quinn, K.B. Ritchie, S. Rodríguez, A. Ramírez, S. Romano, J.F. Samhouri, J.A. Sánchez, G.P. Schmahl, B. Shank, W.J. Skirving, S.C.C. Steiner, E. Villamizar, S.M. Walsh, C. Walter, E. Weil, E. H. Williams, K. W. Roberson & Y. Yusuf. 2010. Caribbean corals in crisis: record thermal stress, bleaching, and mortality in 2005. PLoS ONE 5: e13969. [ Links ]
Finlay, J. 2000. Grenada: National Biodiversity Strategy and Action Plan: Assessment and Analysis of Fisheries Marine and Coastal Areas, Consultants Report. United Nations Development Programme; Global Environmental Facility. Project No.: GRN/98/ G31/A/1G/99. [ Links ]
Gardner, T.A., I.M. Côté, J.A. Gill, A. Grant & A.R. Watkinson. 2003. Long term region-wide declines in Caribbean corals. Science 301: 958-960. [ Links ]
Goldenberg, S.B., C.W. Landsea, A.M. Mestas-Nuñez & W.M. Gray. 2001. The recent increase in Atlantic hurricane activity: causes and implications. Science 293: 474-479. [ Links ]
Hawkins, J.P. & C.M. Roberts. 2003. Effects of artisanal fishing on Caribbean coral reefs. Conserv. Biol. 18: 215-226. [ Links ]
Hughes, T.P. 1994. Catastrophes, phase shifts, and large-scale degradation of a Caribbean coral reef. Science 265: 1547-1551. [ Links ]
Hughes, T.P., A.H. Baird, D.R. Bellwood, M. Card, S.R. Connolly, C. Folke, R. Grosberg, O. Hoegh-Guldberg, J.B.C. Jackson, J. Kleypas, J.M. Lough, P. Marshall, M. Nystrom, S.R. Palumbi, J. M. Pandolfi, B. Rosen & J. Roughgarden. 2003. Climate change, human impacts, and the resilience of coral reefs. Science 301: 929-933. [ Links ]
Hughes, T.P., N. Graham, J.B.C. Jackson, P.J. Mumby & R.S. Steneck. 2010. Rising to the challenge of sustaining coral reef resilience. Trends Ecol. Evol. 25: 633-642. [ Links ]
Idjadi, J.A., S.C. Lee, J.F. Bruno, W.F. Precht, L. Allen-Requa & P.J. Edmunds. 2006. Rapid phase shift reversal on a Jamaican coral reef. Coral Reefs 25:65-68. [ Links ]
Idjadi, J.A., R.N. Haring & W.F. Precht. 2010. Recovery of the sea urchin Diadema antillarum promotes scleractinian coral growth and survivorship on shallow Jamaican reefs. Mar. Ecol. Prog. Ser. 403: 91-100. [ Links ]
Jeffrey, C.F.G. 2000. Annual, coastal and seasonal variation in Grenadian demersal fisheries (1986-1993) and implications for management. Bull. Mar. Sci. 66: 305-319. [ Links ]
Kohler, K.E. & S.M. Gill. 2006. Coral Point Count with excel extensions (CPCe): A visual basic program for the determination of coral and substrate coverage using random point count methodology. Comput. Geosci. 32: 1259-1269. [ Links ]
Kramer, P.A. 2003. Synthesis of coral reef health indicators for the western Atlantic: results of the AGRRA program (1997-2000). Atoll. Res. Bull. 496: 1- 57. [ Links ]
Lapointe, B.E., M.M. Littler & D.S. Littler. 1997. Macroalgal overgrowth of fringing coral reefs at Discovery Bay, Jamaica: bottom-up versus top-down control. Proc. 8th Int. Coral Reef Symp., Panama. 927-932. [ Links ]
Littler, M.M., D.S. Littler & B.L. Brooks. 2009. Herbivory, nutrients, stochastic events, and relative dominances of benthic indicator groups on coral reefs: a review and recommendations. Proc. Mar. Sci. Network Symp., Washington, DC, USA. Smithsonian Contr. Mar. Sci. 38: 401-414. [ Links ]
Mora, C. 2008. A clear human footprint in the coral reefs of the Caribbean. Proc. Royal Soc. B. 275: 767-773. [ Links ]
Mumby, P.J. 2009. Phase shifts and the stability of macroalgal communities on Caribbean coral reefs. Coral Reefs 28: 761-773. [ Links ]
Newman, M.J.H., G.A. Paredes, E. Sala & J.B.C. Jackson. 2006. Structure of Caribbean coral reef communities across a large gradient of fish biomass. Ecol. Lett. 9: 1216-1227. [ Links ]
Ogden, J. 2010. Marine spatial planning (MSP): A first step to ecosystem-based management (EBM) in the wider Caribbean. Rev. Biol. Trop. 58 (Suppl. 3): 71-79. [ Links ]
Sandin, S.A., J.E. Smith, E.E. DeMartini, E.A. Dinsdale, S.D. Donner, A.M. Friedlander, T. Konotchick, M. Malay, J.E. Maragos, D. Obura, O. Pantos, G. Paulay, M. Richie, F. Rohwer, R.E. Schroeder, S. Walsh, J.B.C. Jackson, N. Knowlton & E. Sala. 2008. Baselines and degradation of coral reefs in the Northern Line Islands. PLoS ONE 3: e1548. [ Links ]
Sokal, R.R. and J.F. Rohlf. 1995. Biometry. 3rd ed. W.H. Freeman, New York. [ Links ]
Toller, W., A.O. Debrot, M.J.A. Vermeij & P. C. Hoetjes. 2010. Reef fishes of Saba Bank, Netherlands Antilles: assemblage structure across a gradient of habitat types. PLoS ONE 5: e9207. [ Links ]
Walsh, S.M. 2011. Ecosystem-scale effects of nutrients and fishing on coral reefs. Mar. Biol. 2011: 1-13. [ Links ]
Wilkinson, C. 2008. Status of Coral Reefs of the World: 2008. Global Coral Reef Monitoring Network and Reef and Rainforest Research Centre, Townsville, Australia. [ Links ]
Woodley, J. D., E.A Chronesky, P.A. Clifford, J.C.B. Jackson, L.S. Kaufman, N. Knowlton, J.C. Lang, M.P. Pearson, J.W. Porter, M.C. Rooney, K.W. Rylaarsdam, V.J. Tunnicliffe, C.M. Wahle, J.L. Wulff, A.S. . Curtis, M.D. Dallmeyer, B.P. Jupp, M.A.R. Koehl, J. Neigel & E.M. Sides. 1981. Hurricane Allen’s impact on Jamaican coral reefs. Science 214: 749-755. [ Links ]
Zar, J.H. 1999. Biostatistical Analysis. 4th ed. Prentice Hall, New Jersey. [ Links ]
*Correspondencia:
Robert Anderson: Wisconsin Lutheran College, 8800 W. Bluemound Rd., Milwaukee, WI 53226, USA; banderson@wlc.edu.
Clare Morrall: St. George’s University, P.O. BOX 7, St. George’s, Grenada, West Indies; cmorrall@sgu.edu.
Steve Nimrod: St. George’s University, P.O. BOX 7, St. George’s, Grenada, West Indies; snimrod@sgu.edu.
Robert Balza: Wisconsin Lutheran College, 8800 W. Bluemound Rd., Milwaukee, WI 53226, USA; rob.balza@wlc.edu.
Craig Berg: Milwaukee County Zoo, 10001 W Bluemound Road, Milwaukee, WI 53226, USA; craig.berg@milwcnty.com.
Jonathan Jossart: Wisconsin Lutheran College, 8800 W. Bluemound Rd., Milwaukee, WI 53226, USA; jossart1@gmail.com.
1. Wisconsin Lutheran College, 8800 W. Bluemound Rd., Milwaukee, WI 53226, USA; banderson@wlc.edu, rob.balza@wlc.edu, jossart1@gmail.com
2. St. George’s University, P.O. BOX 7, St. George’s, Grenada, West Indies; cmorrall@sgu.edu, snimrod@sgu.edu
3. Milwaukee County Zoo, 10001 W Bluemound Road, Milwaukee, WI 53226, USA; craig.berg@milwcnty.com
Received 15-VII-2011.Corrected 1 XII-2011.Accepted 20-XII 2011.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}