










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.60 n.4 San José Dec. 2012
Intra and inter-annual structure of zooplankton communities in floodplain lakes: a long-term ecological research study
*Dirección para correspondencia
Abstract
Water flow management has significantly changed the natural dynamic of floods, which are responsible for the structure and dynamic of aquatic communities in river-floodplain systems. With the aim to elaborate a conceptual framework that describes the main ecological factors associated with zooplankton community structure in the Upper Paraná River, we investigated the mechanisms that regulate the communities structure and their response to inter-annual and hydro-sedimentological variations in the floodplain and the biological factors associated with species abundance in those communities. For this we conducted samplings every six months (potamophase in March and limnophase in September) to characterize intra and inter-annual variations in community structure between 2000 and 2008. The intra-annual differences on the species richness, abundance, Shannon diversity index, and evenness, were conducted using Bayesian procedures to show probabilistic predictions of the data fit to main variation sources. Non-metric multi-dimensional scaling (NMDS), multi-response permutation procedure (MRPP), and indicator species analysis (IndVal) were run to assess and characterize the seasonality of the community structure. During high water (potamophase), hydrologic connectivity favoured exchange and dispersal of species in some lakes, increasing local diversity; during low water (limnophase), higher local productivity favoured opportunistic taxa, increasing species dominance and decreasing local diversity. Food resources and density of small-size fish were biological factors associated with the seasonal dynamic of the zooplankton community; these factors were dependent on hydrosedimentological phase (potamophase or limnophase). Water levels and limnological modifications related to water flow management have promoted replacement and impoverishment of aquatic biota in affected lakes and have indicated the ecological importance of a natural dynamic flood, which displays regular flood pulses. The conceptual model presented encompassed interactions between diverse environmental variables to more understandable mechanisms of the main sources of community variation.
Key words: diversity, community structure, environmental monitoring, bayesian procedures, conceptual framework, wetlands.
Resumen
El manejo del régimen de inundación ha cambiado de manera significativa la dinámica natural de las inundaciones, que son responsables de la estructura y dinámica de las comunidades acuáticas en sistemas río-planicie de inundación. En este sentido, investigamos cómo la estructura de las comunidades zooplanctónicas responde a variaciones estacionales e interanuales en los sistemas de llanura de inundación, y los factores biológicos asociados con la abundancia de especies de las comunidades zooplanctonicas. Elaboramos también, un marco conceptual que describe los principales factores ecológicos asociados con la estructura de las comunidades para los sistemas del Alto río Paraná. Para ello se realizaron muestreos cada seis meses (potamophase en marzo y en septiembre limnophase) para caracterizar las variaciones intra e interanuales en la estructura de la comunidad entre 2000 y 2008. Las diferencias estacionales de la riqueza de especies, abundancia, índice de diversidad de Shannon y equitatividad, se llevaron a cabo utilizando procedimientos Bayesianos para mostrar predicciones probabilísticas de los datos ajustados para las principales fuentes variación. Análisis de ordenamiento no-métrico multi-dimensional (NDMS); procedimientos de permutación de respuestas múltiples (MRPP) y análisis de especies indicadoras (IndVal) fueron utilizados para evaluar y clasificar la estacionalidad de la estructura de la comunidad. Durante aguas altas (potamofase), la conectividad hidrológica favoreció el intercambio y dispersión de las especies en algunas lagunas, incrementando la diversidad local; durante aguas bajas (limnofase), la alta productividad local favoreció los táxones oportunistas, incrementando las especies dominante y disminuyendo la diversidad local. La disponibilidad de recursos alimenticios y la densidad de pequeños peces fueron los factores biológicos asociados con la dinámica estacional de la comunidad zooplanctonica; los cuales dependen de la fase hidrosedimentológica (potamofase o limnofase). Modificaciones del régimen hidrológico y limnológico relacionados con el manejo de inundación han promovido el reemplazo y empobrecimiento de la biota acuática, en las lagunas sin conexión afectadas, enfatizando así la importancia ecológica de la dinámica natural de inundaciones, que presenta pulsos regulares de inundación. El modelo conceptual que se presenta abarca desde interacciones entre diversas variables ambientales hasta mecanismos más comprensibles de las principales fuentes de variación de la comunidad.
Palabras clave: diversidad, estructura de la comunidad, manejo ambiental, procedimientos Bayesianos, modelo conceptual, humedales.
Diversity of zooplankton in river-floodplains system is frequently ascribed to interactions between habitat diversity and flood pulse (Robertson & Hardy 1984, Vásquez & Rey 1989, Rodrigo et al. 2003, De Paggi & Paggi 2007, Lansac-Tôha et al. 2009). Such interactions are caused by the dynamics of flooding, which is the driving force for variation in communities and their responses to spatial and temporal variations in river-floodplain systems (Junk et al. 1989, Neiff 1990). These community variations depend on the frequency, intensity (water volume), amplitude, and seasonality (period of occurrence) of the phases of the hydro-sedimentological pulse, which fluctuates between limnophase (low water) and potamophase (high water) (Neiff 1990). During limnophase, water bodies are disconnected because floodplain waters are restricted to rivers, channels, backwaters, and lakes; in potamophase, on the other hand, the increase in water volume promotes connections amongst water bodies. These hydro-sedimentological phases are characterised by distinct faunal assemblages, which show different functional properties related to the predominance of different ecological mechanisms in each phase.
Modifications of flow, due to human activities (e.g. reservoir management), and consequent changes in the flood dynamic, have placed floodplains amongst the most threatened habitats on Earth, significantly endangering biodiversity in these ecosystems (Tockner & Stanford 2002, Dudgeon et al. 2006). This occurs because reservoirs reduce the natural amplitude of water levels variation, consequently changing the dynamics of aquatic communities (Ward & Stanford 1995). The resulting ecological damage may have economic and social impacts in human communities, since recent studies indicated that biodiversity ensures the maintenance of ecosystem processes, buffering the effects of detrimental changes (Galat & Lipkin 2000, Joy & Death 2002, Naeem 2002, Srivastava & Vellend 2005).
An extensive literature exists showing the associations between local environmental variations and the structure of zooplankton communities (Dodson 1992, Schell et al. 2001, Hobaek et al. 2002) and the influence of human activities on these communities (e.g. Beaver et al. 1998, Dodson & Lillie 2001, Dodson et al. 2005, Angeler & Moreno 2007, Dodson et al. 2007). However, there is a paucity of information about zooplanktonic indicator species (such as those developed for phytoplankton by Reynolds et al. 2002) due to the wide niche ranges of most zooplanktonic species, despite several reports of species-specific responses to environmental variation (Cottenie et al. 2001, Cardoso & Marques 2004, Aoyagui & Bonecker 2004, Trevisan & Forsberg 2007).
The development of experimental approaches, and the capacity to control variables, have provided more accurate models for making inferences about the environmental effects of human activities on organisms. However, it is still difficult to reproduce environmental conditions that simulate the complexity and high degree of environmental variability in nature (Franklin 1987). Nevertheless, mediumand long-term data from long-term ecological research (LTER) studies have provided a bridge between empirical studies and the development of ecological models, informing social, economic, and political strategies for improved management of floodplain ecosystems (Baker et al. 2000, Barbosa et al. 2004).
The present study evaluates the temporal variability in the structure of zooplankton communities in lakes influenced by both natural flood pulse and anthropogenic operation of reservoirs. We tested the following hypotheses: (i) attributes of the zooplanktonic community are influenced by hydro-sedimentological phases; (ii) these attributes show patterns of covariation within communities; (iii) hydrosedimentological phases are characterized by different zooplanktonic assemblages (composition and dominance); and (iv) these phases distinctly influence the biotic factors (e.g. resource availability and predation patterns) associated with zooplankton abundance. Over a nine-year period, changes in the structure of the zooplankton community were evaluated to investigate associations between environmental factors and species response to environmental seasonality, as well as the influence of specific biotic factors (resource availability and predation) related to the hydro-sedimentological phases on zooplankton abundance. Finally, we formulated a conceptual framework and theoretical models to describe the structure of community patterns observed in floodplain zooplankton communities.
Materials and Methods
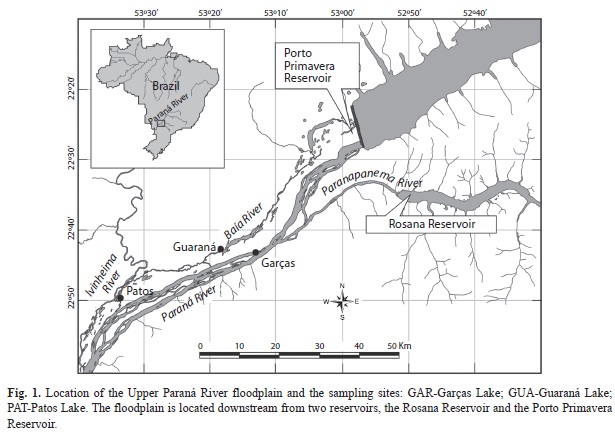
Study area: The Upper Paraná River floodplain (Fig. 1) comprises a variety of habits such as secondary channels, backwaters, tributaries, and temporary and permanent lakes (Agostinho et al. 2004). Currently, three conservation units are present in this area, indicating the relevance of the region for biodiversity conservation; however, their ecological integrity is threatened by a chain of upstream reservoirs.
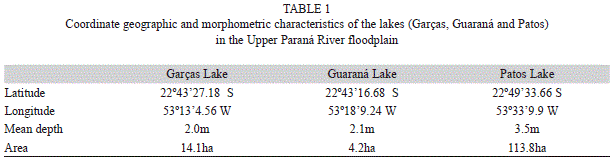
Sampling was conducted in three lakes (Table 1), each perennially connected to a different floodplain river, and influenced by variations in the water levels of the Paraná or Ivinheima rivers (Fig. 2). The level variation in the Garças Lake is directly associated with levels of the Paraná River, which are strongly related to operation of a reservoir located 30km upstream. Lake Guaraná is connected to the Baía River which follows a course parallel to the Paraná River. The hydrodynamics of the Baía River is influenced by floods on the Paraná River. Patos Lake is located in a dam-free region in the Ivinheima River State Park; consequently, this lake exhibits a hydrosedimentological dynamic distinct from the two other studied lakes (Rocha 2002).
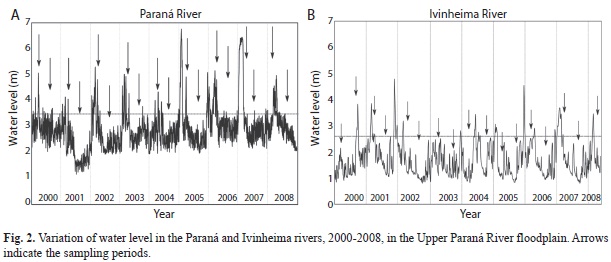
These two rivers showed different flood dynamics; strong floods occurred in 2005 and 2007 in the Paraná River, and in 2001 and 2005 in the Ivinheima River (Fig. 2). Extreme droughts occurred in 2001 and 2003, respectively, in the Paraná and Ivinheima rivers. Floods in the Paraná River were more often than in the Ivinheima River (Table 2), but most Paraná floods were shorter (amplitude) than in the Ivinheima River. The indicative reference levels for floods, which distinguish the limnophase and potamophase, were 3.5m (Paraná River) and 2.6m (Ivinheima River), measured in gauges in these two rivers (Rocha v2002, Thomaz et al. 2004). When river levels reach approximately these intensities, flood waters overflow marginal levels and inundate surrounding floodplains and associated aquatic systems (Rocha 2002). Generally, limnophase occurs during winter (ca. May-October) and potamophase occurs during summer (ca. November-March). The daily water levels of the Paraná and Ivinheima rivers were furnished by Itaipu Binacional.
Sampling of zooplankton and environmental variables: Samplings were conducted every six months (potamophase in March and limnophase in September) between 2000 and 2008 to characterize intra and inter-annual variations in community structure. Environmental data were water temperature (°C), concentration of dissolved oxygen (mg/L) (YSI oximeter), pH, electrical conductivity (µS/ cm) (DIGIMED potentiometer), total alkalinity (µeq/L)(Gran titration; Carmouze 1994), water transparency (cm)(Secchi depth), chlorophyll a concentration (µg/L) (Golterman et al. 1978), organic and inorganic suspended solids (mg/L), nutrients (µg/L) (nitrate Bergamin et al. 1978, ammonia Mackereth et al. 1978, total phosphorus Golterman et al. 1978), and density of fish (catch per unit effort, CPUE) with standard lengths <7cm. The fish sample represents small species and young individuals from medium and large species; although this size group includes non-planktivorous species, the young individuals of larger species are potential consumers of zooplankton. These fishes were captured using gillnets of different mesh sizes ranging from 2.4 to 4cm opposite knots. Further information about CPUE can be found in Fernandes et al. (2009); further information on methods of water analysis and their spatio-temporal dynamics can be found in Roberto et al. (2009).
Zooplankton was sampled in the pelagic region during the morning (between 7 and 11h), using a motorised pump with the boat moving at a constant velocity (to take a composite sampling from each lake), and plankton net (68µm) to filter 600L of water per sample (Lansac Tôha et al. 2009). The samples were preserved in formaldehyde (4%) buffered with calcium carbonate. Individuals were identified to the lowest taxonomic level possible using taxonspecific literature (Vucetich 1973, Koste 1978, Reid 1985, Matsumura-Tundisi 1986, Segers 1995, Velho & Lansac-Tôha 1996, ElmoorLoureiro 1997). Zooplankton abundance was determined using a Sedgewick-Rafter counting chamber under an optical microscope and results were given in ind/m3. At least 80 individuals were counted (Bottrell et al. 1976) in each of three sequential samples, obtained with a Hensen-Stempell pipette (2.5mL).
The zooplankton community was analysed in terms of species richness, abundance, Shannon diversity index, evenness (Krebs 1999), non-metric multi-dimensional scaling (NMDS), multi-response permutation procedure (MRPP), and indicator species analysis (IndVal) (Dufrêne & Legendre 1997).
Uni-dimensional dependent variables (species richness, abundance, Shannon diversity index, and evenness) were analysed with Bayesian procedures; these are more flexible than conventional statistics because traditional assumptions are not required, and they permit probabilistic predictions; conventional statistics, on the other hand, are restricted to statements about long-run averages obtained from hypothetical replicates of sampled data (McCarthy 2007). Furthermore, in general, Bayesian procedures provide more precise estimates of model parameters, because it is possible to incorporate knowledge acquired in previous studies into the structure of the model (Choy et al. 2009).
Local and hydro-sedimentological effects on community attributes (species richness, abundance, Shannon diversity index, and evenness) were evaluated with a Bayesian model analogous to factorial ANOVA, aiming to verify how hydro-sedimentological variations influence the attributes of the zooplankton community in the studied lakes. Hydro-sedimentological effects consisted of two levels (potamophase and limnophase), while local effect with three levels (corresponding to the three lakes). We assumed that the dependent variable, Yab (community attributes of phase a and locality b), were normally distributed (μab, σ2) with ε~N(0, σ2), where E(Yab)=µ0+αa+βb +αβab+ε. In this model, µ0 is the mean of the data, α is the effect of the hydro-sedimento-logical phase, β is the locality effect, αβ is the interaction effect, and the subscripts a and b are phases and locality levels, respectively. We assumed a priori non-informative, approximately normal distribution (0, 103; mean and standard deviation, respectively) for the parameters of this equation. The Bayesian credibility interval for the hydro-sedimentological and local effects was simulated using Markov chain Monte Carlo methods with 20 000 iterations and a burn-in of 1 000 iterations. All the chains analysed reached convergence. Both abundance and richness data were log transformed to decrease the range of data variation.
We used the Pearson correlation to determine how attributes covariate within communities; and if they were associated with water levels (mean of the 30 days before samplings; because it represent a temporal dynamic of the water level that can contribute to temporal variations of the assemblages due to time lag of the zooplankton responses).
Variations in the structure of the zooplankton community were summarised using NMDS, searching the best solution for representation in two dimensions. NMDS does not require assumptions about the distribution patterns of species abundance and is suitable for ecological data structures inflated by zeros (McCune & Grace 2002). Differences between limnophase and potamophase community structures were analysed using MRPP (Zimmerman et al. 1985) with 10 000 randomizations. This is a non-parametric permutation procedure applied to a previously defined similarity or dissimilarity matrix. We used the Bray-Curtis dissimilarity method, conducting the analysis with transformed data to reduce discrepancies amongst the abundances of different species [log2 (x+1), where x represents the abundance of individuals (m3)]. Rare species, defined as having a frequency of occurrence lower than 10%, were removed from the analyses to facilitate the observation of environmental patterns.
Associations of environmental variables with community structure were determined by Envfit. Envfit is an function of the Vegan package (R programming language) that determines the environmental vectors that best represent the distribution of community structures using a permutation procedure (Oksanen et al. 2008). To avoid collinearity, we removed some environmental variables that were highly correlated (r>0.70). Following this criterion, water temperature and pH were eliminated owing to their high correlations with dissolved oxygen.
Once we verified intra-annual patterns in the structure of the zooplankton community, we used a Bayesian multiple regression analysis to assess whether the same biotic factors influenced zooplankton abundance during the phases (limnophase and potamophase). In the multiple regression model, the following explanatory variables were selected to distinguish biological factors that regulate zooplankton abundance: chlorophyll a concentration, as an indicator of food resource availability, and fish density (standard length<7cm), as an indicator of predation pressure. We considered that the dependent variable (Y, community abundance) was normally distributed (μ, σ2) with ε~N(0, σ2), where E(Yi)=β0+β1xi+β2xi+ε. We also considered that the parameter β1 (fish den-sity) presented a priori a non-informative nor-mal distribution, β1~N(0, 103). Moreover, the parameters β0 (intercept) and β2 (chlorophyll a concentration) were normally distributed with positive values N (μ>0, 103); because, the food availability β2 always has a positive effect on zooplankton abundance. The Bayesian credibility interval for the regression coefficients was simulated using a Markov chain Monte Carlo method with 100 000 iterations, a burn-in of 1 000 iterations, and a thinning interval of 15 to minimise autocorrelation. All the analysed chains reached convergence.
The analyses were developed with R software version 2.8.1 (Development Core Team), using the VEGAN package (Oksanen et al. 2008) for the multivariate analyses and BRugs for the Bayesian analyses.
Results
Zooplankton composition and diversity: The zooplankton community was represented by 342 species, including 196 rotifers, 76 testate protozoans, 50 cladocerans, and 20 copepods. Guaraná Lake contained 267 species; Garças Lake, 242 species; and Patos Lake, 189 species. Seventy species were restricted to Garças Lake, 42 species to Guaraná Lake, and 13 species to Patos Lake. A total of 143 species were common to all three lakes.
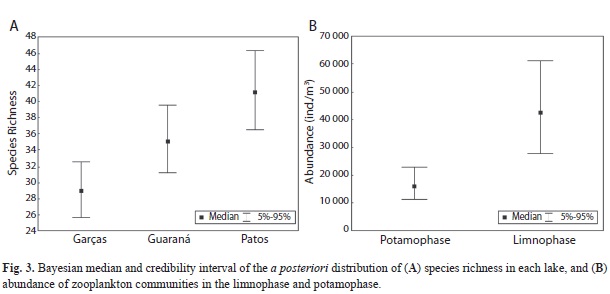
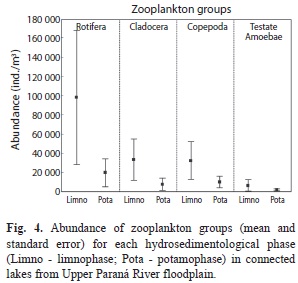
The mean species richness was highest in Patos Lake (mean=40, SD=12), followed by Guaraná Lake (35; 14) and Garças Lake (29; 12). The a posteriori credibility interval pointed to differences in the effects of locality on species number (Fig. 3a), mainly between Garças Lake, which had the lowest value, and Patos Lake, which had the highest value for species richness. Community abundance did not differ amongst localities, but was higher during limnophase and lower during potamophase (Fig. 3b). The a posteriori credibility interval of the hydro-sedimentological phase effects indicated a positive effect of the limnophase on community abundance. Rotifers were the most abundant group, and Testate amoebae was the least abundant one (Fig. 4). In general, the limnophase also showed higher variation of abundance than potamophase.
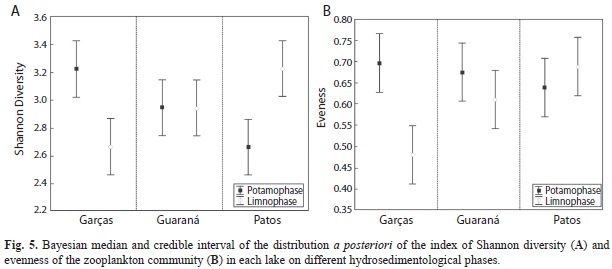
Species diversity and evenness presented patterns of variation similar to species richness, with a strong effect for the interaction between locality and seasonality (Fig. 5a and b); this interaction was characterized by lower values of evenness (mean=0.48) and Shannon diversity (mean=2.7bits/ind) during limnophase in Garças Lake, suggesting a negative effect of the limnophase in this locality. On the other hand, the same period showed a positive effect on species diversity (3.2bits/ind) and evenness (0.69) in Patos Lake. No hydrosedimentological phase effects were detected in Guaraná Lake.
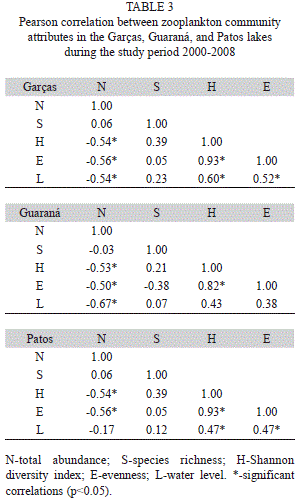
Relationship between community attributes and water level: Community attributes presented significant correlations with each other (Table 3). Increased species abundance was associated with decreased Shannon diversity index and evenness. Thus, at lower abundances, the species were distributed more uniformly, and, consequently, diversity was higher. Therefore, the most frequent association observed was between evenness and Shannon diversity index; i.e. homogeneous distributions amongst organisms favoured an increase in the Shannon diversity index. Elevation of water levels was associated with decreased zooplankton abundance, thereby increasing the Shannon diversity index and evenness in these communities.
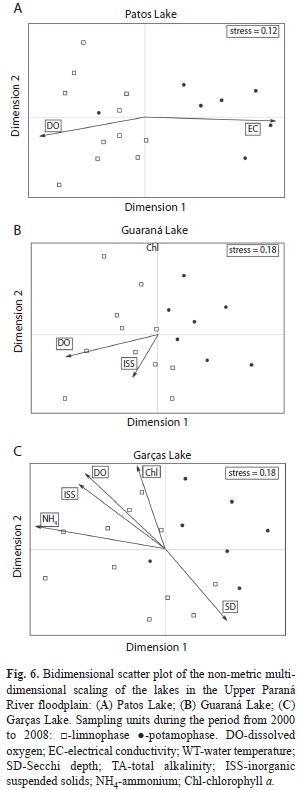
Structure of the zooplankton community: The hydro-sedimentological phases (potamophase and limnophase) were characterized by different structures of the zooplankton communities in the three lakes (Fig. 6; MRPP<0.01). IndVal results (Table 4) showed that 48 of 342 species significantly distinguish the hydrosedimentological phases, with 20 species characteristic of the limnophase and 28 species characteristic of the potamophase.
In Patos and Guaraná Lakes, the limnological features most associated with community structure were dissolved oxygen and inorganic suspended solids in the limnophase, and electrical conductivity in the potamophase. Lower values of dissolved oxygen (range=0.5-7mg/L) and inorganic suspended solids (0.1-3.8mg/L) were observed during potamophase. Higher values of dissolved oxygen (6.8-9.5mg/L) and inorganic suspended solids (0.2-8.8mg/L) were observed in the limnophase. Lower values of electrical conductivity were observed in the limnophase, while higher values occurred in the potamophase.
A large number of variables were associated with the structure of the zooplankton community in Garças Lake (dissolved oxygen, inorganic suspended solids, ammonium, chlorophyll a, and Secchi depth) showing a variation in the local productivity. During potamophase, an increase in water transparency (Secchi depth reaching 2.4m) was recorded and lower levels of dissolved oxygen (range=2.5-7.8mg/L), inorganic suspended solids (0.2-1.5mg/L), and chlorophyll a (1.2-9.1mg/L).
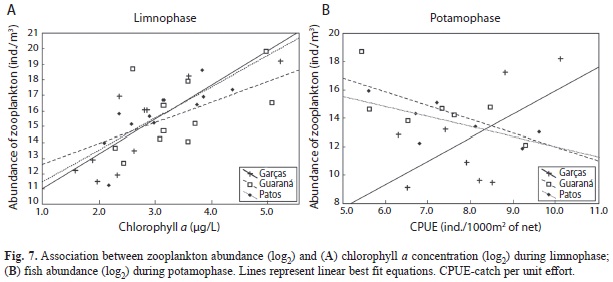
Intra-annual variability of trophic relationships in the zooplankton community: Biological factors likely responsible for variation in abundance were analysed with a Bayesian multiple regression model that suggested positive associations between zooplankton abundance and chlorophyll a concentration during limnophase (Fig. 7a). It is likely that an increase in resource availability during this phase favours an increase in total abundance. During potamophase, in Guaraná and Patos Lakes, community abundance was negatively associated with fish abundance (Fig. 7b), suggesting an effect of predation.
Bayesian a posteriori probabilities (0.99, 0.98, and 1.00; Patos, Guaraná, and Garças lakes, respectively) support the hypothesis that resource availability controls zooplankton abundance during limnophase. Regarding predation effects, Patos and Guaraná lakes presented high probabilities (0.94 and 0.82, respectively) of negative effect of predation on zooplankton abundance. Otherwise, the low probability in Garças Lake (0.06) suggested little evidence of a predatory effect in this locality.
Discussion
Zooplankton composition and diversity: Lansac-Tôha et al. (2009) recorded about 540 species of cladocerans, copepods, rotifers, and testate protozoans in the pelagic region from several localities in the Upper Paraná River floodplain. In the present study, we recorded 63% of these species in the three lakes studied; these lakes are permanently connected to rivers, thus enhancing diversity through constant exchange of fauna with other environments (Ward et al. 1999, Aoyagui & Bonecker 2004, Alves et al. 2005). In general, high species diversity in floodplains is a result of interactions between structural heterogeneity and functional diversity. Structural heterogeneity refers to the complexity of interconnected environments, formed by a mosaic of hydrologic landscapes including rivers, secondary channels, backwaters, and tributaries, each with distinct hydrodynamic features (lotic, semi-lentic, and temporary and permanent lakes) (Junk 2002, Agostinho et al. 2004). Ward & Tockner (2001) suggested that functional diversity in riverine environments is a consequence of the interaction promoted by natural disturbances (e.g. floods) on functional processes (e.g. nutrient cycling, energy flow, and biotic interactions) in the different water bodies. These disturbances (floods) contribute to an increase of biodiversity because it favours spatial and temporal turnover amongst species. Thus, however we sampled only connected lakes, this interaction functional contributed to the high local diversity.
The high accumulated species richness values in Guaraná Lake (267 species) and Garças Lake (242 species) are likely due to temporal replacements of local fauna in these lakes; species frequently observed at the beginning of the study gradually disappeared, and other species started to appear during the time frame of the study. An increase in the frequency of hydrodynamic disturbances in the Paraná River (many short flood pulses) might have negatively influenced the occurrence of common species, and favoured species resistant to disturbance. Furthermore, the operation of a reservoir, that began one year before this study, could contribute to these patters, since during the aging of reservoirs there is a change in its fauna (Agostinho et al. 1999), which also affects the downstream lakes.
Interestingly, high values of accumulated richness did not result in greater mean diversities. Mean diversity was higher in Patos Lake than in other lakes. In Patos Lake, the water level variability occurred naturally and human interventions, such as impoundments that control overland water flow, were not present. This suggests that sites with fewer human disturbances have a higher mean diversity and greater number of specialized niches (Ricklefs & Schluter 1993). Thus, the results indicate that water level and limnological modifications recorded after 2000 (Roberto et al. 2009) contributed to qualitative impoverishment and replacement of aquatic biota in Garças and Guaraná lakes (diminishing the local species richness, but increasing the accumulated species richness), highlighting the ecological importance of natural flood dynamics (with regular flood pulses), as observed in the Ivinheima River.
Some studies have suggested that floods lose substantial characteristics in localities where hydrological control is exerted by reservoirs (Kingsford 2000, Dudgeon et al. 2006, Steinberg et al. 2009); the controlled floods that occur in these managed localities do not produce the same effects as observed under natural conditions. Under regulated conditions, flood characteristics are modified by, for example, reductions in sediment load, large variations in the frequency of the pulse, and reduced amplitudes of water level variations. Thus, regulated rivers are subject to fewer pronounced floods and droughts and to fewer stochastic events.
Relationship between community attributes and water level: The associations of community attributes identified in this study (species richness, Shannon diversity index, evenness, and abundance) were also reported by Paidere et al. (2007) and Lindholm et al. (2007). These relationships characterize the behaviour of the zooplankton community according to two main scenarios: (i) at low abundances, fauna uniformly distributed, evenness increases, an increase in the diversity index is favoured, and species coexistence is promoted by minimizing competitive exclusion (Paidere et al. 2007); (ii) at high abundances, few species predominate, which decreases the evenness and the diversity of species. Water level influences the community dynamic, decreasing the total abundance but increasing the diversity of species.
Structure of the zooplankton community: Hydro-sedimentological phases were distinguished by the structures of zooplankton communities in the studied lakes (MRPP). Studies performed in South American floodplain systems have found higher abundances of some zooplanktonic species during dry periods or limnophase (Brandorff & Andrade 1978, Saunders & Lewis Jr. 1989, Paggi & De Paggi 1990, Lansac-Tôha et al. 1993, Lima et al. 1998, Rossa & Bonecker 2003, De Paggi & Paggi 2007; Henry et al. 2011), when the hydrodynamic conditions were more stable, thus promoting primary productivity which supports large populations of zooplanktonic organisms due to higher water residence time (Baranyi et al. 2002). In contrast, during potamophase, we observed a dilution effect in which organisms were more dispersed in the water column (Bozelli 2000, Lansac-Tôha et al. 2009). Furthermore, the environmental conditions were disadvantageous for the development of large zooplankton populations, due to higher hydrodynamic instability, decreases in autochthonous productivity (lower chlorophyll a concentration), and increases in predation pressure by fish.
Seasonal patterns in the structure of zooplankton communities were strongly correlated with environmental variables: dissolved oxygen, inorganic suspended solids, and electrical conductivity. These variables were indicative of processes that influence the zooplankton community during limnophase, such as higher concentration of dissolved oxygen during the limnophase is partially due to primary production which, in turn, may increase zooplankton densities.
Community structure in Garças Lake was characterised by inter-annual variations related to variations system productivity. The upstream reservoir reduced sediment transport and promoted increase in water transparency and a decrease in nitrogen and phosphorus (Roberto et al. 2009). In this lake, there was an increase in the number of environmental variables associated with the structure of the zooplankton community, indicating that this local community is more susceptible to sources of variation. This effect is attenuated in Guaraná Lake, because the particular biogeochemical composition of this site sustains nutrient levels higher than those present in the influx waters from the Paraná River.
Species that characterize limnophase occur at higher abundances and frequencies in this period. Amongst them, Keratella cochlearis has shown high abundances during dry periods in both natural and affected environments (Beaver et al. 1998, Rossa & Bonecker 2003, Cardoso & Marques 2004). The reproductive effort this species is highest in productive habitats in controlled experiments (Nagae unpublished data). The increased abundance of Bosminidae species (Bosmina hagmanni and Bosminopsis deitersi) and Daphnia gessneri in limnophase is also related to higher productivity (Matsumura-Tundisi 1984, Lopes et al. 1997, Lima et al. 1998, Branco et al. 2000, Rejas et al. 2005).
In potamophase, planktonic and littoral species are distributed in the pelagic region, because littoral species are displaced from the littoral region by hydrodynamic instability. Lecanidae, Epiphanes clavatula, and Dipleuchlanis propatula propatula (littoral rotifers), Chydoridae species (littoral cladocerans) and Moina reticulata (a pelagic cladoceran) were frequently observed during potamophase in other studies conducted in this same floodplain (Lima et al. 1998, Rossa & Bonecker 2003, Alves et al. 2005).
Intra-annual variability of trophic relationships in the zooplankton community: Data on chlorophyll a concentrations support the hypothesis that food availability increases community abundance during limnophase periods, reflecting a positive effect of resource availability; this pattern was observed in all three lakes, and in other studies conducted in floodplain environments over shorter time periods (Bozelli 1996, Beaver et al. 1998, Azevedo & Bonecker 2003, Trevisan & Forsberg 2007). A negative association of zooplanktonic abundance with fish density was observed in Guaraná and Patos Lakes, suggesting an effect of zooplanktivorous fishes. Zooplankton organisms are important components of the diets of small fish in lakes of the Upper Paraná River floodplain, mainly during potamophase (Russo & Hahn 2006, Crippa et al. 2009, Hahn & Crippa 2006). The evaluation of these biotic interactions is complex because relationships can be concealed by the ways data are presented, or due the association correlates with others variables (Bonecker et al. 2012)”. For example, zooplanktivorous predation may be significant only on large-size individuals (size-efficiency hypothesis, Brooks & Dodson 1965); variables can have confounded each other, e.g. an increase in water transparency may intensify predation pressure by fish (Scheffer 1998, Cottenie et al. 2001).
Thus, it is provable that trophic interactions, represented by the associations between zooplankton populations and resource availability, and zooplankton populations and predation, were evidently different in the two phases, showing a distinct trophic dynamic between limnophase and potamophase, as reported by Angeler et al. (2000). As suggested by Neiff (1996) and Thomaz et al. (2007), factors inherent to the hydro-sedimentological phases have different influences on aquatic communities. In potamophase, floods, which are regional-scale processes, result in dilution of the populations and homogenization of environments, whereas in limnophase, local processes, such as productivity, separately influence the succession of isolated communities.
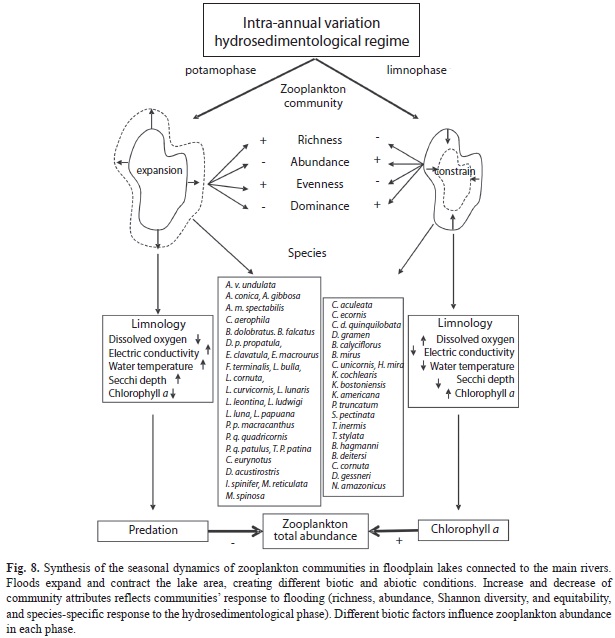
Conceptual framework for variability in the structure of zooplankton communities in floodplains: The main variation sources of zooplankton communities in connected lakes, that are influenced by the flood dynamic in the Upper Paraná River floodplain, were summarized as follows (Fig. 8):
a) Flood dynamic processes, which influence the zooplankton community structure physically and biologically: Physically, flood dynamic processes are responsible for the expansion and contraction of environments because they define the size and conditions of the habitat, as well as the quantity and quality of resources. Biologically, the floods change the structure and dynamic of the zooplankton community by defining the relationships between community attributes; an increase in connectivity favours the exchange and dispersion of species, which increases zooplankton diversity. During limnophase, biotic interactions are stronger because high abundances of a few dominant species decrease the evenness and the species diversity.
b) Variations in limnological conditions, which characterize distinct ‘assemblages’: During potamophase, an increase in water transparency and electrical conductivity and a decrease in dissolved oxygen and chlorophyll a were associated with increased abundance of some species. During limnophase, a reduction in the size of water bodies increases available food resources, because productivity in each locality is enhanced by increased concentrations of nutrients and chlorophyll a. These conditions support the growth of opportunistic taxa that were able to exploit habitat conditions, resulting in increased species dominance and decreased species richness.
c) Factors intrinsic to each phase, which are related to resource availability and predation, which, in turn, influence the total abundance of zooplanktonic organisms: In limnophase, the greater availability of food resources supported high zooplankton abundances. In potamophase, however, despite the dilution effect due to the greater volume of water, the decrease in zooplankton abundance may be in part due to fish predation, where the community structure presents marked seasonality.
These findings emphasise the complexity of interactions between physical, chemical, and biological factors in floodplain environments, as noted by other authors, and highlight the important contribution of the hydro-sedimentological phases to the seasonal dynamic of the community. Moreover, the present study overcomes the temporal limitations of previous studies, by considering inter-annual variability in the data, based on a long-term ecological research programme. Although the number of replicates was small, generalisations are based on the results of the present research combined with an extensive literature on floodplain systems.
The conceptual model presented above encompasses the interactions between diverse environmental variables to more complete understanding of the main sources of community variation. Although the model is schematically simple, it does not devalue the environmental complexity (spatial and temporal) that supports and maintains floodplain biodiversity, but instead it facilitates an understanding of system dynamics that is critical for eventual development of management and conservation proposals.
Acknowledgments
This work was supported by Conselho Nacional de Desenvolvimento a Pesquisa (CNPq) with financing of the Long Term Ecological Research; and Coordenação de Aperfeiçoamento de Pessoal de Ensino Superior (CAPES). We thank the Nupélia’s laboratories of Limnology and ichthyology for their assistance with the physical and chemical variables of water and provision of estimates of fish, respectively.
References
Agostinho, A.A., L.E. Miranda, L.M. Bini, L.C. Gomes, S.M. Thomaz & H.I. Suziki. 1999. Patterns of colonization in Neotropical reservoir, and prognoses on aging, p. 227-265. In J.G. Tundisi & M. Straskraba (eds.). Theoretical reservoir ecology and its applications. International Institute of Ecology, Brazilian Academy of Sciences and Buckhuys, São Carlos, Brazil. [ Links ]
Agostinho, A.A., S.M. Thomaz & L.C. Gomes. 2004. Threats for biodiversity in the floodplain of the Upper Parana River: effects of hydrological regulation by dams. Ecohydrol. Hydrobiologia 4: 267-280. [ Links ]
Alves, G.M., L.F.M. Velho, F.A. Lansac-Toha, B. Robertson & C.C. Bonecker. 2005. Effect of the connectivity on the diversity and abundance of cladoceran assemblages in lagoons of the upper Parana river floodplain. Acta Limnol. Bras. 17: 317-327. [ Links ]
Angeler, D. & J. Moreno. 2007. Zooplankton community resilience after press-type anthropogenic stress in temporary ponds. Ecol. Appl. 17: 1105-1115. [ Links ]
Angeler, D., M. Alvarez-Cobelas, C. Rojo & S. Sanchez- Carrillo. 2000. The significance of water inputs to plankton biomass and trophic relationships in a semiarid freshwater wetland (central Spain). J. Plankton Res. 22: 2075-2093. [ Links ]
Aoyagui, A. & C.C. Bonecker. 2004. Rotifers in different environments of the Upper Paraná River floodplain (Brazil): richness, abundance and the relationship with connectivity. Hydrobiologia 522: 281-290. [ Links ]
Azevedo, F. & C.C. Bonecker. 2003. Community size structure of zooplanktonic assemblages in three lakes on the upper River Parana floodplain, PR-MS, Brazil. Hydrobiologia 505: 147-158. [ Links ]
Baker, K., B. Benson, D. Henshaw, D. Blodgett, J. Porter & S. Stafford. 2000. Evolution of a multisite network information system: the LTER information management paradigm. BioScience 50: 963-978. [ Links ]
Baranyi, C., T. Hein, C. Holarek, S. Keckeis & F. Schiemer. 2002. Zooplankton biomass community structure in a Danube River floodplain system: effects of hydrology. Freshwater Biol. 47: 473-482. [ Links ]
Barbosa, F.A.R., F.R. Scarano, M.G. Sabará & F.A. Esteves. 2004. Brazilian LTER: Ecosystem and biodiversity information in support of decision-making. Environ. Monit. Assess. 90: 121-133. [ Links ]
Beaver, J.R., A.M. Miller-Lemke & J.K. Acton. 1998. Midsummer zooplankton assemblages in four types of wetlands in the Upper Midwest, USA. Hydrobiologia 380: 209-220. [ Links ]
Bonecker, C.C., Azevedo, F.A & Simões, N.R. 2012. Zooplankton body-size structure and biomass in tropical floodplain lakes: relationship with planktivorous fishes. Acta Limnol. Bras. 23(3): 217-228. [ Links ]
Bergamin, H., B.F. Reis & E.A.G. Zagatto. 1978. A new device for improving sensitivity and stabilization in flow injection analysis. Anal. Chim. Acta 97: 63-70. [ Links ]
Bottrell, H.H., A. Duncan, Z. Gliwicz, E. Grygierek, A. Herzig, A. Hillbricht-Illkowska, H. Kurasawa, P. Larsson & T. Weglenska. 1976. A review of some problems in zooplankton production studies. Norw. J. Zool. 24: 419-456. [ Links ]
Bozelli, R.L. 1996. The influence of bauxite tailings on the cladoceran populations of Lake Batata, Amazonia, Brazil. Int. Rev. Gesamten Hydrobiologia 81: 621-634. [ Links ]
Bozelli, R.L. 2000, Zooplâncton, p. 119-138. In R.L. Bozelli, F.A. Esteves & F. Roland (eds.). Lago Batata: impacto e recuperação de um ecossistema amazônico. IB-UFRJ/SBL, Rio de Janeiro, Brazil. [ Links ]
Branco, C., F.A. Esteves & B. Kozlowsky-Suzuki. 2000. The zooplankton and other limnological features of a humic coastal lagoon (Lagoa Comprida, Mace, RJ) in Brazil. Hydrobiologia 437: 71-81. [ Links ]
Brandorff, G. & E. Andrade. 1978. The Relationship between the water level of the Amazon River and the fate of the zooplankton population in Lago Jacaretinga, a várzea lake in the Central Amazon. Stud. Neotrop. Fauna Environ. 13: 63-70. [ Links ]
Brooks, J. & S.I. Dodson. 1965. Predation, body size, and composition of plankton. Science 150: 28-35. [ Links ]
Cardoso, L.S. & D.M. Marques. 2004. Structure of the zooplankton community in a subtropical shallow lake (Itapeva Lake-South of Brazil) and its relationship to hydrodynamic aspects. Hydrobiologia 518: 123-134. [ Links ]
Carmouze, J. 1994. Metabolismos dos ecossistemas aquáticos: fundamentos teóricos, métodos de estudo e análises quimicas. Edgard Blucher/Fapesp, São Paulo, Brazil. [ Links ]
Choy, S., R. O’Leary & K. Mengersen. 2009. Elicitation by design in ecology: using expert opinion to inform priors for Bayesian statistical models. Ecology 90: 265-277. [ Links ]
Cottenie, K., N. Nuytten, E. Michels & L. De Meester. 2001. Zooplankton community structure and environmental conditions in a set of interconnected ponds. Hydrobiologia 442: 339-350. [ Links ]
Crippa, V., N.S. Hahn & R. Fugi. 2009. Food resource used by small-sized fish in macrophyte patches in ponds of the upper Paraná river floodplain. Acta Sci. Biol. Sci. 31: 119-125. [ Links ]
De Paggi, J. & J.C. Paggi. 2007. Zooplankton, p. 229-245. In M.H. Iriondo, J.C. Paggi & M.J. Parma (eds.). The Middle Paraná River: Limnology of a Subtropical Wetland. Springer, New York, USA. [ Links ]
Dodson, S.I. 1992. Predicting crustacean zooplankton species richness. Limnol. Oceanogr. 37: 848-856. [ Links ]
Dodson, S.I. & R.A. Lillie. 2001. Zooplankton communities of restored depressional wetlands in Wisconsin, USA. Wetlands 21: 292-300. [ Links ]
Dodson, S.I., R.A. Lillie & S. Will-Wolf. 2005. Land use, water chemistry, aquatic vegetation, and zooplankton community structure of shallow lakes. Ecol. Appl. 15: 1191-1198. [ Links ]
Dodson, S.I., W.R. Everhart, A.K. Jandl & S.J. Krauskopf. 2007. Effect of watershed land use and lake age on zooplankton species richness. Hydrobiologia 579: 393-399. [ Links ]
Dudgeon, D., A.H. Arthington, M.O. Gessner, Z. Kawabata, D.J. Knowler, C. Lévкque, R.J. Naiman, A.H. Prieur-Richard, D. Soto, M.L Stiassny & C.A. Sullivan. 2006. Freshwater biodiversity: importance, threats, status and conservation challenges. Biol. Rev. 81: 163-182. [ Links ]
Dufrêne, M. & P. Legendre. 1997. Species assemblages and indicator species: the need for a flexible asymmetrical approach. Ecol. Monogr. 67: 345-366. [ Links ]
Elmoor-Loureiro, L.M.A. 1997. Manual de identificação de cladóceros Límnicos do Brasil. Universa, Brasília, Brazil. [ Links ]
Fernandes, R., A.A. Agostinho, E.A. Ferreira, C.S. Pavanelli, H.L. Suzuki, D.P. Lima & L.C. Gomes. 2009. Effects of the hydrological regime on the ichthyofauna of riverine environments of the upper Paraná River floodplain. Braz. J. Biol. 69: 669-680. [ Links ]
Franklin, J.F. 1987. Importance and justification of LongTerm Studies in Ecology, p. 3-20. In G.E. Likens (ed.). Long-Term Studies in Ecology: Approaches and Alternatives. Springer-Verlag, New York, USA. [ Links ]
Galat, D. & R. Lipkin. 2000. Restoring ecological integrity of great rivers: historical hydrographs aid in defining reference conditions for the Missouri River. Hydrobiologia 422/423: 29-48. [ Links ]
Golterman, H.L., R.S. Clymo & M.A.M. Ohmstad. 1978. Methods for Physical and Chemical Analysis of Fresh Waters. Blackwell Scientific, Oxford, England. [ Links ]
Hahn, N.S. & V. Crippa. 2006. Estudo comparativo da dieta, hábitos alimentares e morfologia trófica de duas species simpátricas, de peixes de pequeno porte, associados а macrófitas aquáticas. Acta Sci. Biol. Sci. 28: 359-364. [ Links ]
Henry, R., E.A. Panarelli, S.M.C. Casanova, D.C. Granado, R.C. Mortari & J. Abra. 2011. Plankton richness and abundance in several different hydrological situations in lakes lateral to a river: a case study in the mouth zone of a tributary into a tropical Reservoir. Oecol. Aust. 15: 537-558. [ Links ]
Hobaek, A., M. Manca & T. Andersen. 2002. Factors influencing species richness in lacustrine zooplankton. Acta Oecol. 23: 155-163. [ Links ]
Joy, M.K. & R.G. Death. 2002. Predictive modelling of freshwater fish as a biomonitoring tool in New Zealand. Freshwater Biol. 47: 2261-2275. [ Links ]
Junk, W.J. 2002. Long-term environmental trends and the future of tropical wetlands. Environ. Conserv. 29: 414-435. [ Links ]
Junk, W.J., P.B. Bayley & R.E. Sparks. 1989. The flood pulse concept in river-floodplain system. Can. Spec. Publ. Fish. Aquat. Sci. 106: 110-127. [ Links ]
Kingsford, R. 2000. Ecological impacts of dams, water diversions and river management on floodplain wetlands in Australia. Aust. Ecol. 25: 109-127. [ Links ]
Koste, W. 1978. Rotatoria die Rädertiere Mitteleuropas begründet von Max Voight. Monogononta. Gebrüder Borntaeger, Berlin, Germany. [ Links ]
Krebs, C. 1999. Ecological methodology. Harper & Row, New York, USA. [ Links ]
Lansac-Tôha, F.A., A. Lima, S.M. Thomaz & M.C. Roberto. 1993. Zooplâncton de uma planície de inundação do rio Paraná. II. Variação sazonal e influência dos níveis fluviométricos sobre a comunidade. Acta Limnol. Brasil. 6: 42-55. [ Links ]
Lansac-Tôha, F.A., C.C. Bonecker, L.F.M. Velho, N.R. Simões, J.D. Dias, G.M. Alves & E.M. Takahashi. 2009. Biodiversity of zooplankton communities in the Upper Paraná River floodplain: interannual variation from long-term studies. Braz. J. Biol. 69: 539-549. [ Links ]
Lima, A., F.A. Lansac-Tôha, L.F.M Velho & L. Bini. 1998. Environmental influence on planktonic cladocerans and copepods in the floodplain of the Upper River Paraná. Brazil. Stud. Neotrop. Fauna Environ. 33: 188-196. [ Links ]
Lindholm, M., D.O. Hessen, K. Mosepele & P. Wolski. 2007. Food webs and energy fluxes on a seasonal floodplain: the influence of flood size. Wetlands 27: 775-784. [ Links ]
Lopes, R.M., F.A. Lansac-Tôha, R. Vale & M. Serafim Jr.1997. Comunidade zooplanctônica do reservatório do segredo, p. 3-60. In A.A. Agostinho & L.L. Gomes (eds.). Reservatório de Segredo: Bases ecológicas para o manejo. EDUEM, Maringá, Brazil. [ Links ]
Mackereth, F., J. Heron & J. Talling. 1978. Water analysis: some revised methods for limnologists. Freshwater Biological Association, Sientific publication, London, England. [ Links ]
Matsumura-Tundisi, T.1984. Occurrence of species of the genus Daphnia in Brazil. Hydrobiologia 112: 161-165. [ Links ]
Matsumura-Tundisi, T. 1986. Latitudinal distribution of Calanoida copepods in freshwater aquatic systems of Brazil. Braz. J. Biol. 46: 527-553. [ Links ]
McCarthy, M. 2007. Bayesian Methods for ecology. Cambridge University, New York, USA. [ Links ]
McCune, B. & J.B. Grace. 2002. Analysis of ecological communities. MjM Software Design, Gleneden Beach, Oregon, USA. [ Links ]
Naeem, S. 2002. Ecosystem consequences of biodiversity loss: the evolution of a paradigm. Ecology 83: 1537-1552. [ Links ]
Neiff, J.J. 1990. Ideas para la interpretación ecológica del Paraná. Interciencia 15: 424-441. [ Links ]
Neiff, J.J. 1996. Large rivers of South America: toward the new approach. Verh. Internat. Verein. Limnol. 26: 167-180. [ Links ]
Oksanen, J., R. Kindt, P. Legendre, B. O’Hara & M. Stevens. 2008. vegan: Community Ecology Package. R package version 1.8-8 (available online: http://r-forge. r-project. org/projects/vegan). [ Links ]
Paggi, J.C. & J. De Paggi. 1990. Zooplâncton de ambientes lóticos e lênticos do rio Paraná médio. Acta Limnol. Bras. 3: 685-719. [ Links ]
Paidere, J., D. Gruberts, A. Skute & I. Druvietis. 2007. Impact of two different flood pulses on planktonic communities of the largest floodplain lakes of the Daugava River (Latvia). Hydrobiologia 592: 303-314. [ Links ]
Reid, J. 1985. Chave de identificação e lista de referências bibliográficas para as espécies continentais sulamericanas de vida livre da ordem Cyclopoida (Crustacea, Copepoda). Bol. Zool. 9: 17-143. [ Links ]
Rejas, D., S. Declerck, J. Auwerkerken, P. Tak & L. Meester. 2005. Plankton dynamics in a tropical floodplain lake: fish, nutrients, and the relative importance of bottom-up and top-down control. Freshwater Biol. 50: 52-69. [ Links ]
Reynolds, C.S., V. Huszar, C. Kruk, L. Naselli-Flores & S. Melo. 2002. Towards a functional classification of the freshwater phytoplankton. J. Plankton. Res. 24: 417-428. [ Links ]
Ricklefs, R. & D. Schluter. 1993. Species diversity: regional and historical influences, p. 350-363. In R. Ricklefs & D. Schluter (eds.). Species diversity in ecological communities. University of Chicago, Chicago, USA. [ Links ]
Roberto, M.C., N.F. Santana & S.M. Thomaz. 2009. Limnology in the Upper Paraná river floodplain: largescale spatial and temporal patterns, and the influence of reservoirs. Braz. J. Biol. 69: 717-725. [ Links ]
Robertson, B.A. & E.R. Hardy. 1984. Zooplankton of Amazonian lakes and rivers, p.337-352. In H. Sioli (ed.). The Amazon. Limnology and landscape ecology of a might tropical river and its basin. Junk publishers, The Hague, Holland. [ Links ]
Rocha, P.C. 2002. Dinâmica dos canais no sistema rio-planície fluvial do alto rio Paraná, nas proximidades de Porto Rico-PR. Ph.D. Thesis, Universidade Estadual de Maringá, Maringá, Brazil. [ Links ]
Rodrigo, M.A., C. Rojo & X. Armengol. 2003. Plankton biodiversity in a landscape of shallow water bodies (Mediterranean coast, Spain). Hydrobiologia 506: 317-326. [ Links ]
Rossa, D. & C.C. Bonecker. 2003. Abundance of planktonic and non-planktonic rotifers in lagoons of the Upper Paraná River floodplain. Amazoniana 17: 567-581. [ Links ]
Russo, M.R & N.S. Hahn. 2006. Importance of zooplankton in the diet of a small fish in lagoons of the upper Paraná River floodplain, Brazil. Acta Limnol. Brasil.18: 357-366. [ Links ]
Saunders III, J. & W. Lewis Jr.1989. Zooplankton abundance in the lower Orinoco River, Venezuela. Limnol. Oceanogr. 43: 397-409. [ Links ]
Scheffer, M. 1998. Ecology of shallow lakes. Chapman & Hall, London, England. [ Links ]
Schell, J.M., C.J. Santos-Flores, P.E. Allen, B.M. Hunker, S. Kloehn, A. Michelson, R.A. Lillie & S.I. Dodson. 2001. Physical-chemical influences on vernal zooplankton community structure in small lakes and wetlands of Wisconsin, USA. Hydrobiologia 445: 37-50. [ Links ]
Segers, H. 1995. Rotifera: the Lecanidae (Monogonta). The Hague: SPB Academic. Guides to the identification of the micro invertebrates of the continental water of the world. SPB, The Hague, Holland. [ Links ]
Srivastava, D.S. & M. Vellend. 2005. Biodiversity-ecosystem function research: is it relevant to conservation? Annu. Rev. Ecol. Syst. 36: 267-294. [ Links ]
Steinberg, A.J., J. Ejsmont-Karabin, J.R. Muirhead, C.T. Harvey & H.U. MacIsaac. 2009. Consistent, longterm change in rotifer community composition across four Polish lakes. Hydrobiologia 624: 107-114. [ Links ]
Thomaz, S.M., T.A. Pagioro, L.M. Bini, M.C. Roberto & R.R.A. Rocha. 2004. Limnology of the Upper Paraná Floodplain Habitats: Patterns of Spatio-Temporal Variation and influence of the water levels, p. 37-42. In A.A. Agostinho, L. Rodrigues, L.C. Gomes, S.M. Thomaz & L.E. Miranda (eds.). Structure and functioning of the Paraná River and its floodplain. EDUEM, Maringá, Brazil. [ Links ]
Thomaz, S.M., L.M. Bini & R.L. Bozelli. 2007. Floods increase similarity among aquatic habitats in riverfloodplain systems. Hydrobiologia 579: 1-13. [ Links ]
Tockner, K. & J. Stanford. 2002. Riverine flood plains: present state and future trends. Environ. Conserv. 29: 308-330. [ Links ]
Trevisan, G. & B. Forsberg. 2007. Relationships among nitrogen and total phosphorus, algal biomass and zooplankton density in the central Amazonia lakes. Hydrobiologia 586: 357-365. [ Links ]
Vásquez, E. & J. Rey. 1989. Longitudinal Study of Zooplankton Along the Lower Orinoco River and Its Delta (Venezuela). Ann. Limnol. 25: 107-120. [ Links ]
Velho, L.F.M. & F.A. Lansac-Tôha. 1996. Testate amoebae (Rhizopodea-Sarcodina) from zooplankton of the high Paraná river floodplain, state of Mato Grosso do Sul, Brazil: II. Family Difflugidae. Stud. Neo. Fauna. Environ. 31: 174-192. [ Links ]
Vucetich, M.C. 1973. Estudio de tecamebianos argentinos, en especial los del dominio pampasico. Rev. Mus. La Plata 118: 287-322. [ Links ]
Ward, J.V. & K. Tockner. 2001. Biodiversity: towards a unifying theme for river ecology.Freshwater Biol. 46: 807-819. [ Links ]
Ward, J.V. & J.A. Standford. 1995. The serial discontinuity concept: extending the model to floodplain river. Regul. Rivers 10:159-168. [ Links ]
Ward, J.V., K. Tockner & F. Schiemer. 1999. Biodiversity of floodplain river ecosystems: ecotones and connectivity. Regul. Rivers 15: 125-139. [ Links ]
Zimmerman, G.M., H. Goetz & P.W. Mielke. 1985. Use of an improved statistical method for group comparisons to study effects of prairie fire. Ecology 66: 606-611. [ Links ]
*Correspondencia: Nadson R. Simões: Universidade Estadual de Maringá-Nupélia/ Laboratory of zooplankton DBI-PEA, Av. Colombo, 5790, 87020-900, Paraná, Brazil; nadsonressye@yahoo.com.br
Fábio A. Lansac-Tôha: Universidade Estadual de Maringá-Nupélia/ Laboratory of zooplankton DBI-PEA, Av. Colombo, 5790, 87020-900, Paraná, Brazil; fabio@nupelia.uem.br
Luiz F. M. Velho: Universidade Estadual de Maringá-Nupélia/ Laboratory of zooplankton DBI-PEA, Av. Colombo, 5790, 87020-900, Paraná, Brazil; felipe@nupelia.uem.br
Claudia C. Bonecker: Universidade Estadual de Maringá-Nupélia/ Laboratory of zooplankton DBI-PEA, Av. Colombo, 5790, 87020-900, Paraná, Brazil; bonecker@nupelia.uem.br
1. Universidade Estadual de Maringá-Nupélia/ Laboratory of zooplankton DBI-PEA, Av. Colombo, 5790, 87020-900, Paraná, Brazil; nadsonressye@yahoo.com.br, fabio@nupelia.uem.br, felipe@nupelia.uem.br, bonecker@nupelia.uem.br
Received 25-X-2011. Corrected 25-III-2012.Accepted 30-IV-2012.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}