










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.60 n.4 San José Dec. 2012
Darkedge midshipman Porichthys analis (Batrachoidiformes: Batrachoididae), a common shrimp-bycatch fish from the Gulf of California
*Dirección para correspondencia
Abstract
Shrimp fishery bycatch of the Gulf of California constitutes a wide variety of highly unknown fish, crustacean, and mollusk species with very low or null economic value, in contrast to those of commercial interest. However, there are no studies yet on the role of these low economic valued species have in the community structure and function, together with their possible effect on commercial populations. With the aim of contributing to the knowledge of Darkedge Midshipman fish Porichthys analis, the most common waste species in this fishery, we estimated some population characteristics. For this purpose, we obtained shrimp-bycatch fish during the 2004-2005 fishing season and performed some research surveys. A total of 1 725 Darkedge Midshipman were captured from 350 bycatch samples. Individual size ranged from 43-352mm SL. The weight to standard length relationship was determined as W=0.000092SL3.0509. Von Bertalanffy’s growth coefficient indicated a moderate growth rate (K=0.5) with an estimated asymptotic length ofL∞=352mm standard length, and a longevity of 6yr. Natural mortality was estimated as 0.97 and total mortality as 4.67. The recruitment pattern, as estimated by ELEFAN II, was extended over the year, peaking during spring and summer seasons. Sexual proportion of male:female was 1.65:1. Mature organisms appeared from August to March, and length at maturity was 157mm SL, which is larger than the mean size at capture 135mm SL. We concluded that the species is indirectly protected by its own bathymetric distribution and the off-shrimping season. This is the first study that considers population characteristics from this common but still unknown species.
Key words: population biology, Porichthys analis, growth, mortality, recruitment.
Resumen
A diferencia de las especies de interés comercial, la historia de vida de los peces no comerciales es casi desconocida a pesar de su potencial importancia en la estructura y función de la comunidad. Para contribuir al conocimiento de estas especies se estimaron algunas características poblacionales del chupalodo Porcihchty sanalis. La fauna de acompañamiento de la pesquería del camarón en el Golfo de California está constituida por especies poco conocidas y de escaso valor comercial. Las especies de la familia Batrachoididae son frecuentemente utilizadas como animales experimentales, pero poco se sabe de P. analis. De esta manera a partir de la fauna de acompañamiento se recolectaron 1 725 organismos y se determinó la relación talla-peso W=0.000092SL3.0509. Mediante ELEFAN I, se estimó un crecimiento moderado (K=0.5/año) con una longitud asintótica mayor que la observada reportada anteriormente (L∞=352mm SL) y una longevidad de 6 años. La mortalidad natural se estimó en M=0.97 la total en Z=4.67. El patrón de reclutamiento fue mayor durante primavera-verano. La proporción sexual M:F fue 1.65:1 y la talla media de primera madurez fue de 157mm SL mayor que la talla media de captura 135mm SL. Este es el primer reporte que considera las características poblacionales de esta especie común, pero poco conocida.
Palabras clave: biología poblacional, Porichthys analis, crecimiento, mortalidad, reclutamiento.
Shrimp fishing is important in the Gulf of California (Sierra-Rodríguez et al. 2005), but most shrimp bycatch species have low or null economic value and are normally discarded or underused (Kelleher 2005). Some studies have described aspects on distribution, taxonomy, or abundance of economically important fish species reasonably well; nevertheless, we have not yet understood their functions in the ecosystems. In contrast, information on noncommercial species is still scarce and some ignored, for example, life histories, seasonal variability, or growth rates (González-Ochoa et al. 2009, Rábago-Quiroz et al. 2008, 2011, López-Martínez et al. 2011, Morales-Azpeitia 2007).
Nowadays, species population characteristics, combined with community descriptors to evaluate aquatic communities, are considered of outmost importance (Piet & Jennings 2005, Rochet et al. 2005). Within this scheme, non-commercial species may function as predators, prey, or competitors, affecting community structure and influencing economically important populations (Horn 1980). Models, such as predation models, could be tested using size-based multispecies to explore community functions or to evaluate fishing effects (Rochet et al. 2005). In addition, the proportion of noncommercial species has been suggested as an indicator of fishing effects in marine communities (Trenkel & Rochet 2003). Besides, the population dynamics of various species is a useful element for community-based model integration. Population parameters such as growth, mortality, recruitment, and morphometric relationships, among others, are considered key pieces of information (Petrakis & Stergiou 1995, Froese 1998, Moutopouolos & Stergiou
2002, López-Martínez et al. 2011).
Shrimp bycatch in the Gulf of California constitutes mainly fish (80%) and in a minor proportion crustaceans and mollusks (López-Martínez et al. 2010). The Darkedge Midshipman fish Porichthys analis (Batrachoididae) (Hubbs & Shults 1939) is one of the most common species in shrimp bycatch (≅14%) (López-Martínez et al. 2010) and is practically endemic in the Gulf of California region. This family comprises benthic and temperate to tropical species (Robertson & Allen 2002). P. analis is one of the biggest species of the genera and catalogued as a potential fishery resource (Collette 1995) lacking economic importance. Additionally, Batrachoididae species have often been used as experimental animals in biomedicine and evolutionary and ecophysiological research (Wang & Walsh 2000, Walsh et al. 2004). However, practically nothing is known about P. analis yet. Therefore, the main objective of this study was to contribute to the knowledge of the species by estimating some population characteristics obtained from shrimp trawl catches in the Gulf of California.
Material and Methods
Study area: The Northern side limits with the Southern end of the Biosphere Reserve of the Upper Gulf of California and the Colorado River Delta; to the South with Rio Fuerte, Sinaloa at the continental shore; to Bahia San Luis Gonzaga at the peninsular shore (21°1’37’’-31°24’35’’ N and 105°16’06’’-114°22’51’’ W). Sampling stations ranged from 10-70m in depth. Two sources of information were available: (1) A program of 10-onboard observers was implemented during the 2004-2005 shrimpfishing seasons, during which 20kg of bycatch samples were taken from commercial trawling and were kept frozen until analysis. Authorized bottom trawling nets were 37.5mm in mesh size at the cod-end and no less than 44.45mm at the square. Sea turtle excluding devices were used in all nets. We took records from depth, trawling speed, crossed distance, and geographical position for every trawl, among others. The species P. analis was found in 91 samples out of 2 439 (2). During the shrimp-spawning season, we obtained additional bycatch samples from research surveys performed in July and August 2004 and 2005. For each trawling, we registered geographic starting and ending position, initial and ending time, depth, speed, sea surface and air temperature, among others (Herrera-Valdivia 2004).
Morphometric measurements: Once in the laboratory, bycatch species were separated and identified (Fischer et al. 1995, Collette 1995) from each sample. The geographic origin of P. analis specimens was recorded as well as total length (L), standard length (SL) (mm) (±1mm) and total weight (W) (g) (±0.1). Sex and sexual maturity were determined according to a gonadal morphochromatic scale (Nikolsky 1963).
Population parameter estimation: Total length-standard length (L-SL) relationship was determined by the least-squares method to fit a linear model L=a+bSL, and weight-standard length relationship (W-SL) to fit a potential model W=aSLbby non-linear estimation. We constructed monthly length frequency distributions (SL), and in using these, the von Bertalanffy growth equation Lt=L∞(1-e (- K (t - t0 ) ) was fitted, where Lt is standard length at age t, L∞ asymptotic length, K instantaneous growth rate, and t0 the theoretical age at zero length. When fitting the von Bertalanffy equation, an initial L∞ value estimation was attempted by the Powell-Wetherall method. With the L∞ value, Shepherd’s method (Sparre & Venema 1998) was used to obtain an initial estimate of K; the final growth parameter estimation was made using ELEFAN I method. By using Pauly’s empirical equation, t0 was obtained as t0=1x10[-0.3922- (0.2752LogL∞)-(1.038LogK)] (Sparre & Venema 1998).
Longevity was estimated as proposed by Taylor (1958) tmax=t0+3/K, where tmax is the longevity (yr) and other parameters as described above. Total mortality rate (Z) was estimated by the linearized length-converted catch curve (Sparre & Venema 1998) Ln(Ni/∆t)=a+bti; where Ni is the number of fish in length class i; ∆t is the time needed for the fish to grow through length class i; t is the relative age corresponding to the mid-length of class i; and b is an estimate of Z (sign changed). Natural mortality (M) was estimated using Pauly´s empirical equation LnM=-0.0152-0.279LnL∞+0.6543LnK+0.463LnT, where T is the mean annual sea surface temperature in the species habitat (ºC). Fishing mortality rate (F) can be calculated by a difference of Z=M+F, and exploitation rate (E) was determined according to Jones (1984), E=Z/F (Sparre & Venema 1998).
Recruitment pattern was estimated by the ELEFAN II method. We also estimated both, length at first capture and length at sexual maturity using only all mature females (stages III-V). Organisms were grouped in 11mm length intervals and cumulated relative frequency calculated by interval. The nonlinear least-square fitting method, which is available in the FISHPARM suite (Prager et al. 1987), was used for fitting the logistic function P=1/ (1+e-a(SL-SL50)), where P is the proportion of caught organisms or mature females in case of sexual maturity: SL50the standard length at which 50% of the population is caught (SLc50) or attains reproductive maturity (SLm50) according to the case.
Results
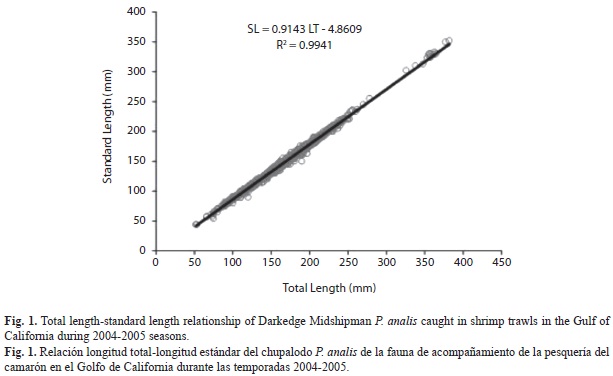
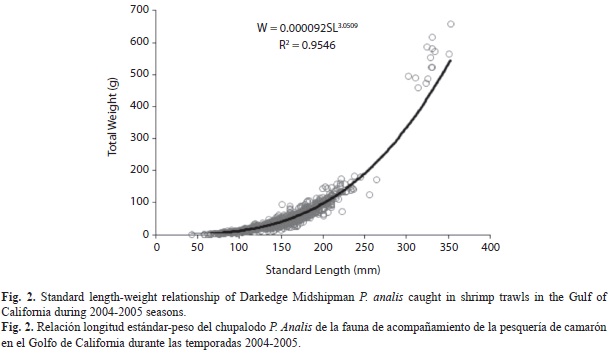
A total of 1 725 P. analis were obtained from commercial and research surveys. Spatial distribution of P. analis showed that it is present along the East coast, from the upper Gulf of California to Sinaloa. Individual sizes ranged from 43-352mm (SL). The L-SL relationship was described by the equation SL=0.9143L-4.8609 (Fig. 1). Given the close correlation between both lengths, post-hoc analyses were performed considering only SL. The SL-W relationship was described by the function W=0.000092SL3.0509 (Fig. 2).
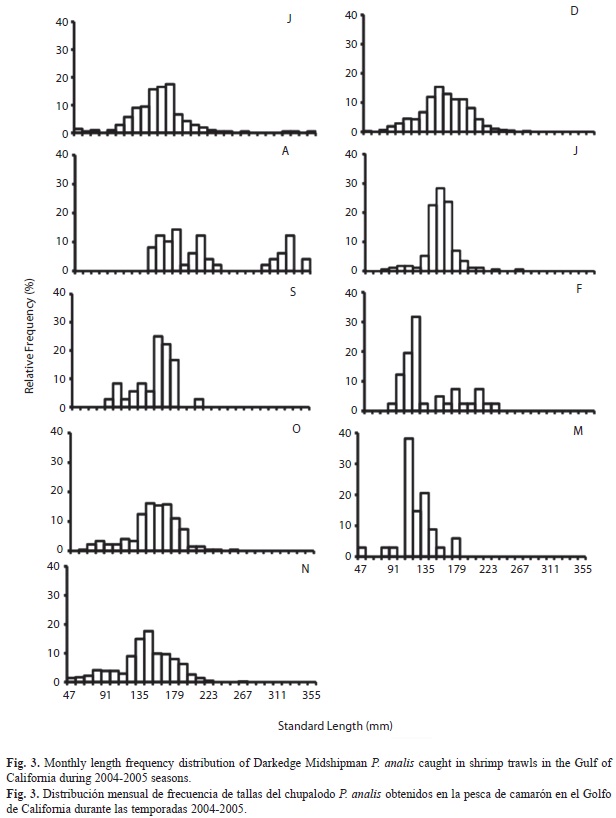
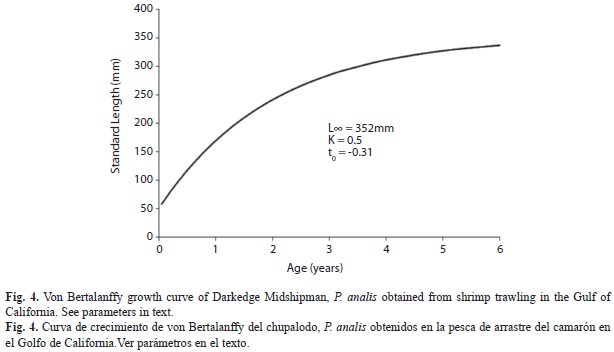
Availability of organisms was restricted to the shrimp-fishing season (September-March) and research survey months (July-August).We used monthly length frequency distributions shown in figure 3 to estimate the von Bertalanffy growth parameters and adjusted the corresponding curve to the data. From our length distribution data, we identified at least four cohorts; only few larger size individuals appeared in July-September surveys. In L∞ estimation by the Powel-Wetherall method, we got an underestimation, so we used the maximum observed length as the L∞ seminal value in the K calculation by the Shepherd (1987) method. Both values were later used for the joint re-estimation by ELEFAN I, giving the adjusted growth equation SL=352(1-e (-0.5(t+0.31))).
We used these parameters to trace the growth curve (Fig. 4); longevity was estimated in 6.31yr.Therecruitment pattern practically extended over the year and showed two peaks: one in February-May and another in July-October (Fig. 5).
Natural mortality M was 0.97 considering a 23ºC temperature, and total mortality Z was 4.67 (±0.49); therefore, fishing mortality F was 3.7 and exploitation rate E was 0.79.
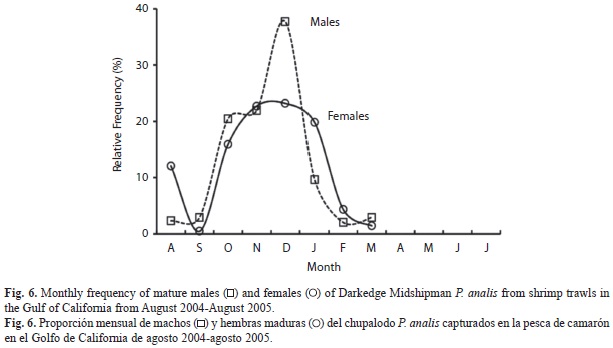
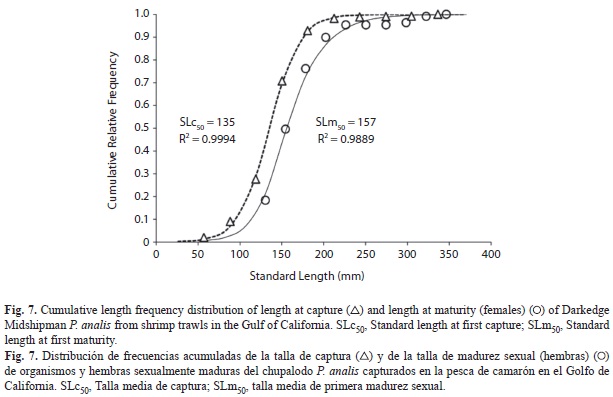
The number of males exceeded the number of females and sexual proportion, M:F was 1.65:1 (X2 α/2, 0.025,7=24.322, p<0.0000). Mature organisms (stages II-IV) appeared from August to March, and frequencies were apparently higher from October to January (Fig. 6). Length at first capture was estimated considering cumulative frequency of all individuals. Adjusted sigmoid function was P=1/ (1+e -0.055(SL-135)), where length at first capture (SLc50) was 135mm SL (Fig. 7). We estimated mean length at first sexual maturity (SLm50) using SL records and considering cumulative frequency of mature females (stages III-V). The adjusted sigmoid curve function was P=1/ (1+e-0.051(SL-157)), so mean length at first maturity was attained at 157mm SL (Fig. 7). Both sizes (curves) were different when compared by Kolmogorov-Smirnov test (D0.05 = 0.1341), with length at capture being smaller than length at first sexual maturity (Sokal & Rohlf 1981).
Discussion
Up to now, papers on P. analis have documented the sites where this species has been recorded. Our sampling comprised those areas from the Gulf of California where commercial trawling takes place, i.e. soft bottom between 9-80m in depth. Only few larger size individuals appeared in July-September surveys. Considered as a demersal benthic species from a visual inspection of body form, it may be inferred that the fish uses body-fin caudal transient locomotion. Species with this shape show low sustained cruise propulsion with sudden rapid acceleration (Webb 1984). Therefore, bigger individuals may have a better swimming capacity, evading trawling gear. Additionally, bathymetric distribution may also contribute because the species can be seen in up to 360m in deeper shrimp (Crustacea: Solenoceridae) research surveys using a similar fishing method (Rábago-Quiroz et al. 2011). Shrimp commercial vessels operate at shallower waters and mainly on the continental shore (López-Martínez et al. 2000, Morales-Azpeitia 2007), while the species distributional so comprises the peninsular side where the platform is much narrower or almost absent (Roberson & Allen 2002).
The weight-length relationship showed an isometric growth, and despite the scarcity of larger sizes in our samples, it resulted similar to other studies. Pearl Spot Midshipman P. margaritatus showed values of a=0.0069, and b=3.19 (Ruiz-Ramírez et al. 1997), and Atlantic Midshipman P. Porosissimus showed a=0.005 and b=3.148 (Vianna et al. 2000). Froese (2006) suggested that only a few individuals in the whole range size and throughout all seasons were enough to get representative results in this relationship.
A major number of larger organisms may lead to a decrement in K estimation. However, the observed maximum size was larger than that previously reported (Collette 1995, Robertson & Allen 2002) as well as the estimated one. In relation to theoretical values Oyster Toad Fish Opsanustau showed an L∞=37.9cm and K=0.16; Gulf Toadfish O. beta L∞=30cm, K=0.22 (Serafy et al. 1997). The maximum observed size of Plainfin Midshipman P. notatus was 33cm attaining longevity of 7yr (Garrison & Miller 1982), which is very close to our P. analis estimates.
Considering the distribution area, the fishing mortality estimate in this work may also be lower than the one obtained because only a proportion (smaller sizes) of the population was caught by shrimp vessels. Moreover, the method used (length converted catch curve) considered death of organisms from the population were due to fishing throughout the time. This problem is showing the mortality evaluation in those species populations where we did not have access to the whole population, as well as in migratory species populations (López-Martínez et al. 2005).
From SL50 and using the inverse von Bertallanfy growth equation, age at maturity is 0.9yr (≅0.11months), which is earlier than Plainfin Midshipman P. notatus (3.5yr) (Garrison & Miller 1982). Length at capture was smaller than length at maturity, so fishing effort may be causing a detrimental effect on the population by recruitment overfishing. Nevertheless, length at maturity may be larger than mentioned because of the few large size organisms in our estimations. At least one species, Daisy Midshipman P. margaritatus shows a higher abundance at 60m during the warmest bottom water season and moves slightly to shallower waters during colder months (Godínez-Domínguez et al. 2001). Although considered as a resident species, our data suggests a low scale reproductive migratory pattern. As mentioned earlier, commercial vessels operate at shallower waters (López-Martínez et al. 2000, Morales Azpeitia 2007), mainly in the continental shore. Therefore, the latitudinal and bathymetric distribution of the species lets a considerable proportion of the population be outside trawling effects, and fishing mortality should be lower.
On the other hand, although an apparently higher frequency of Darkedge Midshipman fish P. analis mature organisms occurred during winter, the following must be considered: The spawning season in P. notatus occurs in springsummer (Garrison & Miller 1982), and in our study there were months without any samples (April to June). Shrimp off-season is from March to August when we had less represented samples from a few research surveys and the few mature organisms appeared from October to January. Considering that recruitment pattern by length back calculation from various cohorts estimates possible hatching periods, a bimodal extended spawning appeared in our work from March to April and a less higher one from August to September. Then the observed mature organisms (Fig. 6) may have corresponded to the second spawning peak (although more accurate data might be obtained from reproductive biology during the shrimp closed season). The open shrimping season is usually from August-September to March-April each year, and then an associated benefit from the off-season is the indirect protection of P. analis spawners, additional to the bathymetric and latitudinal distribution mentioned earlier.
Acknowledgments
We would like to express our thanks to SAGARPA (SAGARPA-CONACYT Project-2003-C01-089) and Centro de Investigaciones Biológicas del Noroeste (CIBNOR Projet EP012) for their support in this study; to Instituto Nacional de Pesca of Mexico for financing and providing samples from research surveys; to Consejo Nacional de Ciencia y Tecnología of Mexico for the fellowship granted to the first author (Beca CONACyT); to E. Herrera-Valdivia, A.M. Siaruki-Quijano, and R. Morales-Azpeitia from Laboratorio de Pesquerías, Campus Guaymas (CIBNOR) for specimen identification, biometrics, and data base conformation; and to D. Dorantes for English edition.
References
Collette, B.B. 1995. Batrachoididae. Peje-sapos, sapos, p. 910-918. In W. Fischer, F. Krupp, W. Schneider, C. Sommer, K.E. Carpenter & V.H. Niem (eds.). Guía FAO para identificación de especies para los fines de la Pesca. Pacífico Centro-Oriental. V. 2. Vertebrados. FAO, Roma. [ Links ]
Fisher, W., F. Krupp, W. Schneider, C. Sommer, K.E. Carpenter & V.H. Niem. 1995. Guía FAO para la identificación de especies para los fines de la pesca: Pacífico Centro-Oriental. Vol. III. Roma, Italia. [ Links ]
Froese, R. 1998. Length-weight relationships for 18 lessstudied fish species. J. Appl. Ichthyol. 14: 117-118. [ Links ]
Froese, R. 2006. Cube law, condition factor and weightlength relationships: history, meta-analysis and recommendations. J. Appl. Ichthyol. 22: 241-253. [ Links ]
Garrison, K.J. & B.S. Miller. 1982. Review of the early life history of Puget Sound fishes. Unpubl. Report FRIUW-8216, Fisheries Research Institute University Washington, Washington, Seattle, USA. [ Links ]
Godínez-Domínguez, E., B. Aguilar-Palomino & G. González-Sansón. 2001. Abundance variation of Porichthys margaritatus (Pisces: Batrachoididae) throughout thermic and bathymetric gradients in the central Mexican Pacific. Rev. Biol. Trop. 49: 643-646. [ Links ]
González-Ochoa, O.A., J. López-Martínez & N.Y. Hernández-Saavedra. 2009. Population characteristics of spotted rose snapper Lutjanus guttatus caught as shrimp bycatch in the Gulf of California. Interciencia 34: 808-811. [ Links ]
Jones, R. 1984. Assessing the effects of changes in exploitation pattern using length composition data (with notes on VPA and cohort analysis). FAO Fish. Tech. Pap. 256, Roma, Italia. [ Links ]
Herrera-Valdivia, E. 2004. Manual para observadores a bordo de la flota comercial. Laboratorio de Pesquerías. Non editeddocument. CIBNOR, Campus Guaymas. Guaymas, Sonora, México. [ Links ]
Horn, M.H. 1980. Diversity and ecological roles of noncommercial fishes in California marine habitats. CalCOFI Rep. 21: 37-47. [ Links ]
Kelleher, K. 2005. Discards in the world’s marine fisheries. An update. FAO Fish. Tech. Pap. 470. Roma, Italia. [ Links ]
López-Martínez, J., M.E. Morales-Bojórquez, F. Paredes-Magallón, D. Luch-Belda & C. Cervantes-Valle. 2000. La pesquería de camarón de altamar de Sonora. In D. Luch-Belda, J. Elourdy-Garay, S.E. Luch-Cota & G. Ponce-Díaz (eds.). B.A.C. Centros de actividad biológica del Pacífico Mexicano. CIBNOR, CICIMAR, CONACYT, La Paz, Baja California Sur, México. [ Links ]
López-Martínez, J., C. Rábago-Quiroz, M.O. NevárezMartínez, A.R. García-Juárez, G. Rivera-Parra & J. Chávez-Villalba. 2005. Growth, reproduction, and size at first maturity of blue shrimp, Litopenaeus stylirostris (Stimpson, 1874) along the east coast of the Gulf of California, Mexico. Fish. Res. 71:93-102. [ Links ]
López-Martínez, J., E. Herrera-Valdivia, J. Rodríguez- Romero & S. Hernández-Vázquez. 2010. Composición taxonómica de peces integrantes de la fauna de acompañamiento de la pesca industrial de camarón del Golfo de California, México. Rev. Biol. Trop. 58: 925-942. [ Links ]
López-Martínez, J., J. Rodríguez-Romero, N.Y. Hernández-Saavedra & E. Herrera-Valdivia. 2011. Population parameters of the Pacific flagfin mojarra Eucinostomus currani (Perciformes: Gerreidae) captured by the shrimp trawling fishery in the Gulf of California. Rev. Biol. Trop. 59: 887-897. [ Links ]
Morales-Azpeitia, R. 2007. Variación espacio temporal de la relación camarón: FAC en la pesquería de camarón del Golfo de California y su uso como medida de manejo. Implicaciones socioeconómicas. Tesis de Doctorado, Universidad Autónoma de Nayarit, Nayarit, México. [ Links ]
Nikolsky, G. 1963. The ecology of fishes. Academic, New York, USA. [ Links ]
Moutopoulos, D.K. & K.I. Stergiou. 2002. Length-weight and length-length relationships of fish species from the Aegean Sea (Greece). J. Appl. Ichthyol. 18: 200-203. [ Links ]
Petrakis, G. & K. Stergiou. 1995. Weight-length relationships for 33 fish species in Greek waters. Fish. Res. 21: 465-469. [ Links ]
Piet, G.J. & S. Jennings. 2005. Response of potential fish community indicators to fishing. ICES J. Mar. Sci. 62: 214-225. [ Links ]
Prager, M.H., C.W. Recksiek & S.B. Saila. 1987. FISHPARM: A microcomputer program for parameter estimation of nonlinear models in fishery science. Old Dominion Univ. Res. Fundation, Oceanogr.Tech. Rep. 87-10. Norfolk, Virginia, USA. [ Links ]
Rábago-Quiroz, C.I., J. López-Martínez, M. Nevárez-Martínez & J. Rodríguez. 2008. Specific composition, spatial distribution, and population dynamics of flatfish species in the shrimp trawl bycatch in the Gulf of California. Hidrobiológica 18: 193-202. [ Links ]
Rábago-Quiroz, C.I., J. López-Martínez, J.E. Valdéz-Olguin & M. Nevárez-Martínez. 2011. Distribución latitudinal y batimétrica de las especies más abundantes y frecuentes en la fauna acompañante del camarón del Golfo de California, México. Rev. Biol. Trop. 59: 255-267. [ Links ]
Robertson, D.R. & G.R. Allen. 2002. Shorefishes of the Tropical Eastern Pacific: an information system. (CD-ROM) Version 1.0.0. Smithsonian Tropical Research Institute, Balboa, Panamá [ Links ].
Rochet, M.J., V. Trenkel, R. Bellail, F. Coppin, O. Le Pape, J.C. Mahe, J.J. Morin, C. Poulard, I. Schlaich, A. Souplet, Y. Ve´rin & J. Bertrand. 2005. Combining indicator trends to assess ongoing changes in exploited fish communities: diagnostic of communities off the coasts of France. ICES J. Mar. Sci. 62: 1647-1664. [ Links ]
Ruiz-Ramírez, S., G. Lucano-Ramírez & J. Mariscal--Romero. 1997. Length-weight relationships of softbottom demersal fishes form Jalisco and Colima states, Mexico. Naga 20: 62-63. [ Links ]
Serafy, J.E., T.E. Hopkins & P.J. Walsh. 1997. Field studies on the ureogenic gulf toadfish in a subtropical bay. I. Patterns of abundance, size composition and growth. J. Fish. Biol. 50: 1258-1270. [ Links ]
Shepherd, J.G. 1987. A weakly parametric method for estimating growth parameters from length composition data, p. 113-119. In D. Pauly & G.R. Morgan (eds.). Length-based methods in fisheries research. ICLARM Conf. Proc. 13. Manila, Philippines, and Kuwait Institute for Scientific Research, Safat, Kuwait. [ Links ]
Sierra-Rodríguez, P., C. Acosta-Castañeda, J.A. García-Borbón, A.R. García-Juárez, A. Liedo-Galindo, J.M. Melchor-Aragón, S. Ramos-Cruz, A. Rosas-Cota, M.P. Toledo Díaz-Rubín & E. Zárate-Becerra. 2005. [ Links ]
Pesquerías del Océano Pacífico. Camarón. INP. Sustentabilidad y pesca responsable en México 1999-2000. (available on-line http://cripens.inp.gob.mx/sust_pesc_resp_ev_manejo_1999_2000/). [ Links ]
Sokal, R.R. & F.J. Rohlf. 1981. Biometry: The principles and practice of statistics in biological research. W.H. Freeman, San Francisco, California, USA. [ Links ]
Sparre, P. & S.C. Venema. 1998. Introduction to tropical fish stock assesment.Part 1.Manual. FAO Fish. Tech. Pap. 306.1 Rev. 2. FAO, Roma, Italia. [ Links ]
Taylor, C.C. 1958. Cod growth and temperature. J. Cons. Int. Explor. Mer. 23: 266-370. [ Links ]
Trenkel, V.M. & M.J. Rochet. 2003. Performance of indicators derived from abundance estimates for detecting the impact of fishing on a fish community. Can. J. Fish. Aquat. Sci. 60: 67-85. [ Links ]
Vianna, M., A.R.G. Tomas & J.R. Verani. 2000. Aspects of the biology of the Atlantic midshipman Porichthys porisissimus (Teleostei, Batrachoididae): an important by-catch species of shrimp trawling off southern Brazil. Rev. Bras. Oceanogr. 48: 131-140. [ Links ]
Walsh, P.J., Z. Wei, C.M. Wood, A.M. Loong, K.C. Hiong, S.M.L. Lee, W.P. Wong, S.F. Chew & Y.K. Ip. 2004. Nitrogen metabolism and excretion in Allen batrachus grunniens (L): effects of variable salinity, confinement, high pH and ammonia loading. J. Fish Biol. 65: 1392-1411. [ Links ]
Wang, Y. & P.J. Walsh. 2000. High ammonia tolerance in fishes of the family Batrachoididae (Toadfish and Midshipmen) Aquat.Toxicol. 50: 205-219. [ Links ]
Webb, P.W. 1984. Body form, locomotion and foraging in aquatic vertebrates. Amer. Zool. 24: 107-120. [ Links ]
*Correspondencia: Oscar A. González-Ochoa: Centro de Investigaciones Biológicas del Noroeste. Km. 1 Carretera a San Juan de La Costa “El Comitán”. Apdo. Postal 128, La Paz, B.C.S. 23097, México; ocibnor@yahoo.com.mx
Juana López-Martínez: Centro de Investigaciones Biológicas del Noroeste, Unidad Sonora. Km. 2.35 Carretera a Las Tinajas, Col. Tinajas, Guaymas, Sonora, México. C.P. 85460; jlopez04@cibnor.mx
Norma Y. Hernández-Saavedra: Centro de Investigaciones Biológicas del Noroeste. Km. 1 Carretera a San Juan de La Costa “El Comitán”. Apdo. Postal 128, La Paz, B.C.S. 23097, México; nhernan04@cibnor.mx
1. Centro de Investigaciones Biológicas del Noroeste, Unidad Sonora. Km. 2.35 Carretera a Las Tinajas, Col. Tinajas, Guaymas, Sonora, México. C.P. 85460; jlopez04@cibnor.mx
2. Centro de Investigaciones Biológicas del Noroeste. Km. 1 Carretera a San Juan de La Costa “El Comitán”. Apdo. Postal 128, La Paz, B.C.S. 23097, México; ocibnor@yahoo.com.mx, nhernan04@cibnor.mx
Received 30-IX-2011. Corrected 10-V-2012. Accepted 18-VI-2012.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}