










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.60 n.4 San José Dec. 2012
Effects of short-term sediment nutrient enrichment and grazer (Neritina reclivata) removal on sediment microalgae in a shallow eutrophic estuary (Alabama, USA)
Just Cebrian1*,2*, Adrienne L. Stutes1,2,5*, Scott Phipps3*, Jason P. Stutes1,2,5, Bart Christiaen1,2 & Jonathan R. Pennock4*
*Dirección para correspondencia
Abstract
The olive snail (Neritina reclivata) is ubiquitous in tropical and sub-tropical systems of the Gulf of Mexico, however its impacts on sediment microalgae have been little studied. Many coastal systems around the world are being eutrophied due to human activities, and seemingly they will continue to be eutrophied to a further extent in the future. Exploring the single and combined impacts of further nutrient enrichment and grazing by the olive snail on sediment microalgae in such eutrophic systems is an important question for our understanding and management of these systems. Here we examine the effects of short-term nutrient enrichment and grazing by the olive snail N. reclivata on sediment microalgal biomass and composition in a shallow eutrophic estuary (Weeks Bay, Alabama, USA) of the Northern Gulf of Mexico. For this, we performed a series of factorial experiments adding or not nutrients and removing or not the snail, for a total of four treatments in each experiment: ambient grazing, ambient nutrients; ambient grazing, increased nutrients; no grazing, ambient nutrients; and no grazing, increased nutrients. We did not find any significant impact of nutrient addition in any of the eight short-term (i.e. four days) experiments carried out. Impacts by the snail were minor; we only found a decrease in biomass due to snail grazing in one of the eight experiments, and no impacts on microalgal (i.e. diatom) composition. High ambient nutrient concentrations in the sediment porewater and low snail abundances on the sediment could explain these findings. Our results suggest that ephemeral, short-term nutrient pulses into eutrophic coastal systems of the Northern Gulf of Mexico, such as Weeks Bay (Alabama, USA), should not greatly affect the abundance of sediment microalgae, even though those pulses occur in well-lit areas. The results further suggest the snail N. reclivata is not a major control of sediment microalgal populations in the subtidal sedimentary areas studied. Our findings contrast with the results of past work in sediments with well-lit and nutrient poor conditions, or sediments with high densities of other snail grazers. In conjunction this and other investigations indicate that the response of sediment microalgae to nutrient enrichment and modified grazer abundance depends to a large extent on the initial levels of nutrient availability and grazing before the system is altered.
Key words: sediment microalgae, eutrophication, Meridian reclivata, chlorophylla, herbivory, bottom-up, top-down.
Resumen
El caracol Neritina reclivata está presente en los sistemas tropicales y subtropicales del Golfo de México, sin embargo, su impacto en los sedimentos de microalgas ha sido poco estudiado. Muchos de los sistemas costeros de todo el mundo están siendo eutrofizados debido a actividades humanas, y al parecer van a seguir siendo eutrofizados en mayor grado en el futuro. La exploración de los efectos individuales y combinados de un mayor enriquecimiento de nutrientes y la herviboría por este caracol en microalgas de sedimentos en estos sistemas eutróficos es una cuestión importante para la comprensión y el manejo de estos sistemas. Aquí se examinan los efectos a corto plazo del enriquecimiento de nutrientes y herviboría del caracol de olivo sobre la biomasa y composición de microalgas de sedimentos en un estuario eutrófico superficial (Weeks Bay. Alabama, USA) del norte del Golfo de México. Para esto se llevaron a cabo una serie de experimentos factoriales añadiendo o no nutrientes y removiendo o no el caracol, para un total de cuatro tratamientos en cada experimento: ambiente con herviboría sin nutrimentos añadidos, ambiente con herviboría y nutrimentos añadidos, ambiente sin herviboría sin nutrimentos añadidos, y ambiente sin herviboría con nutrimentos añadidos. No se encontró ningún impacto significativo por la adición de nutrimentos en ninguno de los ocho experimentos a corto plazo (i.e. cuatro días). Los impactos debidos al caracol fueron menores, sólo se encontró un decrecimiento en la biomasa por herviboría del caracol en uno de los ocho experimentos, y no hubo impacto en la composición de microalgas (i.e. diatomeas). Ambientes con alta concentración de nutrimentos del sedimento y baja abundancia del caracol sobre el sedimento pueden explicar estos resultados. Los resultados obtenidos sugieren que pulsos efímeros y de corto plazo en sistemas costeros eutrofizados del Norte del Golfo de México tal como Weeks Bay (Alabama, USA), no deberían afectar de gran manera la abundancia de microalgas sobre el sedimento, a pesar de que los pulsos ocurran en áreas bien iluminadas. Además los resultados sugieren que el caracol N. reclivata no ejerce mayor control sobre las poblaciones de microalgas en las áreas submareales y sedimentarias estudiadas. Nuestros hallazgos contrastan con resultados de trabajos anteriores en sedimentos con buena iluminación y pobres condiciones nutritivas, o sedimentos con altas densidades de otros caracoles herbívoros. En conjunto, esta y otras investigaciones indican que la respuesta de microalgas en sedimentos al enriquecimiento de nutrientes y abundancia modificada de herbívoros depende en gran medida de los niveles iniciales de disponibilidad de nutrientes y herviboría antes de que el sistema se vea alterado.
Palabras clave: microalgas sedimentaria, eutroficación, Neritina reclivata, Clorofila-a, herviboría.
Sediment microalgae have important roles in shallow coastal systems. They often account for a significant fraction of the system’s total primary productivity, reaching up to 25% of the productivity by phytoplankton and macrophytes (Moncreiff et al. 1992, Schreiber & Pennock 1995). They also represent food resources for many herbivorous and detritivorous organisms.
Consumers of sediment microalgae range from microbes to invertebrate to vertebrate organisms, encompassing a wide variety of feeding modes and diet selectivity (Middleburg et al. 2000, Carman & Fry 2002). The mucilage that sediment microalgae excrete, besides being a food item for detritivores, helps stabilize the sediment and reduces coastal erosion (Miller et al. 1996, Cahoon 1999, Wolfstein & Stal 2002). Sediment microalgae are also key intermediaries in the cycling of organic and inorganic nutrients in shallow coastal systems, modulating nutrient fluxes between the bottom and overlying water-column (Sundbäck et al. 1991, Rizzo et al. 1992).
Some gastropod species are ubiquitous consumers of sediment microalgae. They ingest sediment and obtain most of their nutritional requirements from inhabiting microalgae (Fenchel & Kofoed 1976, Levinton & Bianchi 1981). However, experiments with gastropod grazers have shown variable levels of herbivory, from negligible to high consumption of sediment microalgae. For instance, Blanchard et al. (2000) reported considerable consumption of sediment diatoms by Hydrobia ulvae, with individual consumption rates averaging 22ng chlorophyll-a (Chla) per snail per hour for a snail density of three individuals per square cm, in laboratory experiments with samples collected from Aiguillon Bay (France). Similarly, Lever &Valiela (2005) found that Ilyanassa obsoleta could reduce by up to 50% the concentration of Chla in the sediment during field experiments done in estuaries of Waquoit Bay (Massachussets, USA). In contrast, Bennet et al. (1999) found almost no noticeable effect of Littoraria irrorata on sediment microalgal biomass in laboratory microcosm experiments done with sediments from Pass Fourchon (Lousiana, USA). In a number of field experiments done at the Swedish island of Väddö, Hillebrand & Kahlert (2002) found that macrograzers (where gastropod species, such as Hydrobia sp. and Potamopyrgus jenkinsi, were predominant) had no significant impact on sediment Chla concentration.
Here we study the olive snail (Neritina reclivata) (Say), a species of gastropod that is ubiquitous in shallow coastal systems of the Gulf of Mexico such as marshes, lagoons and estuaries (Russell 1941). In particular we focused on the shallow estuary of Weeks Bay (Alabama). This gastropod is abundant in the oligohaline reaches of the estuary and the only potentially important epifaunal macroinvertebrate grazer on sediment microalgae in these areas (Miller-Way et al. 1996). Studies on the life history, habitat selectivity and feeding behavior of the gastropod are scarce. The few existing reports indicate that N. reclivata resides preferentially in subtidal and intertidal vegetated stands (i.e. seagrass beds and marshes), and hard substrates such as stumps and rocks (Lehman & Hamilton 1980, Sheridan & Livingston 1983). The reports also indicate that these gastropods feed effectively on epiphytic microalgae that grow attached to the seagrasses, marshes and hard substrates, but not on sediment microalgae (Lehman & Hamilton 1980). On this basis, we can expect a rather modest role and low levels of herbivory of N. reclivata on the microalgae that live in the bare sediment adjacent to vegetated fringing habitats such as seagrass beds and marshes. Confirming this hypothesis is important to understand the ecological functions of this ubiquitous gastropod in shallow coastal systems of the Gulf of Mexico.
Along with grazing, nutrient availability may be an important factor for sediment microalgae. Under low ambient nutrient availability and favorable conditions of light, salinity and temperature, enhanced nutrient availability may lead to increased sediment microalgal biomass (e.g. Darley et al. 1981 Wulff et al. 2000, Cebrian et al. 2009). However, widespread transformation of coastal watersheds by humans is increasing the delivery of nutrients into bays and estuaries in many parts of the world (Lotze et al. 2006, Orth et al. 2006). Increased nutrient delivery into coastal waters often results in higher biomass of phytoplankton in the water-column, which reduces the quantity of light that reaches sediment microalgae (Stutes et al. 2007, Anton et al. 2011). Thus, due to high nutrient and low light ambient levels, the response of sediment microalgae to further nutrient enrichment in already-eutrophic coastal systems should be severely dampened in relation to pristine coastal systems.
That hypothesis has been confirmed in some eutrophic coastal systems (e.g. Hillebrand & Kahlert 2002, Cebrian et al. 2009), and it also seems to be the case at the relatively deeper reaches (>0.5m) of the Weeks Bay estuary. Weeks Bay receives high nutrient loads from farms and crop fields through groundwater and discharge from the Fish and Magnolia Rivers (Pennock et al. 2001, Lehrter 2006). Indeed, Stutes et al. (2006) found almost no impact of fertilization on the productivity of sediment microalgae at a number of locations deeper than 0.5m in the estuary and they attributed this to low light availability at the sediment surface due to high attenuation in the water column and high nutrient concentrations in the sediment that appeared saturating for sediment microalgal growth (for specific values see Stutes et al. 2006). It remains to be seen if the response of sediment microalgae to further nutrient enrichment is also much reduced in the shallower reaches (<0.5m) of this eutrophic estuary, where higher light availability at the sediment surface could promote that response to some extent. Determining this will contribute to our understanding of the interactions between sediment microalgae and nutrient inputs in this and other currently or soon-to-be eutrophic coastal systems.
In this paper, we examine whether there are any significant impacts by N. reclivata (i.e. top-down control) on the biomass of microalgae that grow in bare sediment adjacent to fringing marsh areas, and whether fertilization has any significant impacts on the biomass of those sediment microalgae where, due to their shallowness (<0.5m) and ameliorated light availability, adding additional nutrients may have an effect despite high background nutrient levels. By combining nutrient and gastropod manipulations we also address whether there are any interactions between both processes. For a small subset of experiments we also examine the impacts of grazing by N. reclivata on the structure of the sediment diatom community. The results contribute to assessing how N. reclivata and ephemeral nutrient pulses (i.e. short-term nutrient enrichment) affect sediment microalgae in eutrophic estuaries of the North Central Gulf of Mexico.
Materials and Methods
Study locations: The experiments were carried out at two locations in Weeks Bay, Alabama. The first location (hereafter referred to as Magnolia) is on the Southeastern shoreline near the mouth of Magnolia River (one of the two rivers discharging into Weeks Bay), and the second location (referred to as Reserve) is on the Northwestern shoreline within a little embayment near the mouth of Fish River. The locations are shallow, with water depth ranging 0.1-0.6m in both locations during our experiments. The mean tidal range in the Bay is 0.4m (Schroeder et al. 1990). Water flow in the study locations is dominated by tides, river discharge and wind (Pennock et al. 2001). The intertidal region is dominated by cordgrass (Spartina alterniflora Loisel) and black needlerush (Juncus roemerianus Scheele) in the two locations. There is no submerged aquatic vegetation present in the study locations. The subtidal sediment at Magnolia is mainly composed of sand, whereas at Reserve the sediment is dominated by highly-organic, unconsolidated mud. Sediment diatoms, such Achnanthes sp., Amphora sp., Navicula sp. and Nitzschia sp., are abundant in both locations (Miller-Way et al. 1996). Further description and a map of the study area are available in Stutes et al. (2006).
Experimental design: Experiments were carried out seasonally from October 2002 to July 2003 at the two locations. In each season 20 plots were set up at each location. The plots were set up on subtidal sediment adjacent to the shoreline. Ten of the plots were rectangular (37x22x25cm) transparent acrylic cages. The cages had mesh-covered (500μm) windows on the sides (one window per side for a total of four windows per cage) to allow water exchange while preventing access by N. reclivata. The windows were 8x6cm on the short side, and 18x6cm on the long side of the rectangle. The cages were secured with pieces of rebar bolted into the sediment through round braces that were attached to the corners. A rectangular (20x11cm) opening, which was kept shut in between sampling dates with a removable acrylic lid, was made on the top of each cage to allow for the addition of fertilizer (see “Sediment Nutrient Enrichment”) and the collection of samples inside the cages (see “Response Variables”). We carefully cleaned up the cages on each sampling day to prevent mud and slime build-up. The other ten plots were grazing controls, which consisted of an area of open sediment delineated with flags that was equivalent in shape and size to the area enclosed by the cages.
At each site, the area where the plots were set up appears homogeneous (i.e. area adjacent to fringing salt-marsh and completely covered with bare sediment, and similar sediment type, depth and water flow throughout the area). At any rate, in an effort to average out any possible large spatial heterogeneity that went unnoticed, the cages and controls were arranged in pairs, with each pair containing one cage and one control. Plots within a pair were half a meter apart and adjacent pairs were one meter apart. Before setting up the plots, the sediment enclosed by the cage was cleared of snails. We did this carefully, picking the snails one by one and minimizing sediment disturbance. The sediment of cage and control plots was fertilized in five randomly selected pairs (see “Sediment Nutrient Enrichment”). Therefore each experiment represented a two-factorial manipulation with four treatments, i.e. (one) natural nutrient availability and grazing by N. reclivata; (two) natural nutrient availability and no grazing by N. reclivata; (three) enhanced nutrient availability and grazing by N. reclivata and (four) enhanced nutrient availability and no grazing by N. reclivata, with each treatment having five replicates. Once during each of the experiments we counted N. reclivata densities within a 25x25cm2 quadrat tossed ten times haphazardly around the experimental plots. The mean values of these counts should correspond well with the average density of snails found in the non-caged plots (grazing controls) during the experiments.
Sediment nutrient enrichment: Our experiments were intended to examine the short-term impacts of grazing by N. reclivata and nutrient enrichment on sediment microalgae. To do that, we directly fertilized the sediment one time at the beginning of the experiment and surveyed the plots for four days, in accordance with other studies of short-term impacts (Downing et al. 1999). We directly spiked the fertilizer into the sediment because we sought to maximize the chances for inhabiting microalgae to take up and respond to the added nutrients. Other studies have shown that, when directly released in the water column, a large fraction of the fertilizer is taken up by primary producers above the sediment (i.e. phytoplankton, macroalgae and seagrasses) or advected out of the system before entering the sediment (Heck et al. 2006, Anton et al. 2011).
Prior to starting our experiments in fall 2002, we measured ambient nitrogen (N) and phosphorus (P) concentrations in the sediment porewater at the two study locations (see “Porewater Nutrient Analysis”). For each location, and based on the ambient concentrations and information gathered on sediment density and porewater content, we calculated a level of N addition per plot that represented a 70X increase relative to the average ambient concentration. This increase was well within the range of fertilization values applied in other experiments (Worm et al. 2000) and, based on human-induced increases in nitrogen loading observed for other coastal systems (Anton et al. 2011), it represents a realistic increase that could result in Weeks Bay from further human occupation. Then, based on the average ambient porewater P concentrations measured at the study locations, we calculated the level of P addition per plot needed to reach a 17:1 N:P molar ratio in fertilized plots, which corresponds to the internal ratio for sediment microalgae (Hillebrand & Sommer 1999). The calculated addition levels were 0.21g N and 0.03g P per plot in Magnolia, and 0.30g N and 0.04g P per plot in Reserve.
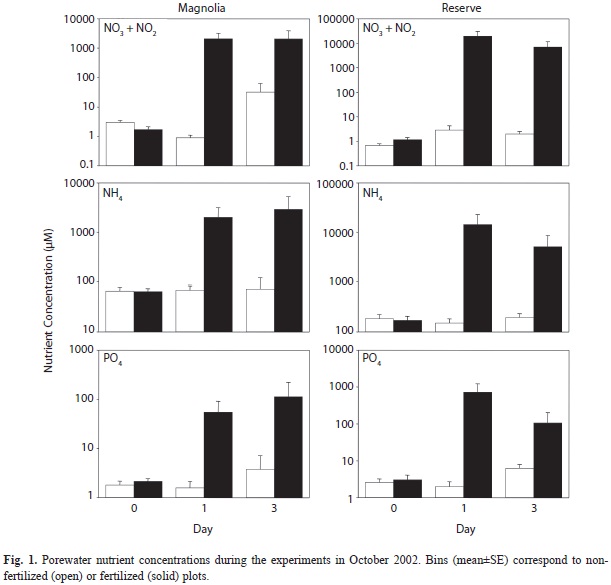
Those nutrient quantities were applied on day zero of the Fall 2002 experiments as a quick-release form of solid commercial-grade ammonium-nitrate (17% ammoniacal nitrogen, 17% nitrate nitrogen, Royster-Clark Inc., Norfolk, VA) and super phosphate (46% P2O5, Agribusiness Inc., Covington, LA). Nutrients were weighed and placed in nylon stockings. Three stockings were placed equidistantly in every fertilized plot, with each stocking having approximately the same amount of N and P. The bottom of the stocking was buried to a depth of approximately 10cm in an effort to maximize dispersion of the fertilizer into the top layer of the sediment while minimizing diffusion into the water column. For both non-fertilized and fertilized plots at each location, measurements of nitrate+nitrite, ammonium and phosphate concentrations in the sediment porewaters were done on days zero (prior to burying the packets), one and three (one measurement of each nutrient species at each plot on each day). The samples were taken and processed as explained below (“Porewater Nutrient Analysis”).
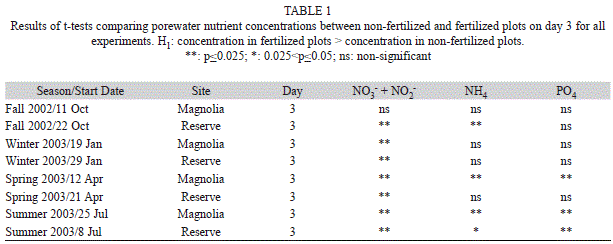
Examination of the porewater nutrient concentrations for the Fall 2002 experiments revealed the concentrations tended to remain higher in fertilized than in non-fertilized plots through day one of the experiment, but those differences tended to fade somewhat on day three (Fig. 1, Table 1). Therefore, in an effort to ensure elevated nutrient availability throughout the duration of the experiment in fertilized plots in relation to ambient conditions, we increased the levels of nutrient addition by three-fold in the other experiments. The new addition levels for the February 2003, May 2003 and July 2003 experiments were 0.63g N and 0.09g P per plot in Magnolia, and 0.9g N and 0.12g P per plot in Reserve. We measured porewater nutrient concentrations on days zero (before burying the packets) and three in all plots for each of these other experiments (one measurement of nitrate + nitrite, ammonium and phosphate concentrations at each plot on each of the two days). Porewater nutrient concentrations were similar between plots prior to fertilization, but were often significantly higher in fertilized than in non-fertilized plots on day three of these experiments (Table 1). Indeed, whereas we only found significantly higher sediment pore water nutrient concentrations in fertilized than in non-fertilized plots on day three in two out of six possible instances (i.e. nitrate+nitrite, ammonium and phosphate at Magnolia and Reserve) in the October 2002 experiments, we found higher concentrations in 12 out of 18 possible instances in the other three experiments (Table 1).
These results indicate that overall we maintained elevated nutrient availability in fertilized in relation to non-fertilized plots throughout the four days the experiment lasted, at least for one of the nutrient forms (e.g. nitrate+nitrite, ammonium or phosphate). However, most of the fertilization-enriched concentrations were observed with nitrate. We found higher ammonium concentrations in fertilized than in non-fertilized plots by day three in four out of the eight experiments done. Since practically all the nutrient pellets stocked were completely dissolved by the end of the experiment, this suggests substantial diffusion of ammonium from the porewater sediment spaces to the overlying water. We found higher phosphate concentrations in fertilized than in non-fertilized plots by day three in three out of the eight experiments done, suggesting substantial adsorption to sediment particles and/or diffusion to the water column.
Porewater nutrient analysis: Porewater nutrient samples were collected using a 60mL syringe attached to an eight cm-long piece of punctured rigid aquarium tubing (Stutes et al. 2006). In an effort to sample the porewater from the fertilized sediment layer (i.e. from 10cm deep to the surface, see above placement of the nutrient-replete nylon stockings), the tubing was completely pushed into the sediment and the porewater drawn up into the syringe. Samples were kept on ice in the field and brought back to the laboratory for processing. All samples were filtered through 0.45μm glass fiber filters and analyzed for nitrate, nitrite, ammonium and phosphate according to standard colorimetric methods (Strickland & Parsons 1972) using a Skalar SAN+Autoanalyzer.
Sediment microalgal biomass: We used chlorophyll a content as a proxy for microalgal biomass (de Jonge & Colijn 1994, Schreiber & Pennock 1995, Cebrian et al. 2008). We took sediment cores using a 2.5cm diameter coring tube. The top 1cm of sediment was transferred to a 50mL centrifuge tube and placed on ice for transportation to the laboratory. Samples were stored at -80oC until processing. The samples were cold extracted in 90% acetone for 24 hours and the pigment concentration measured fluorometrically according to the method of Parsons et al. (1984). In all experiments three chlorophyll a samples were taken per plot on days zero, one and three, and the three samples averaged as a single replicate. Sampled sediment areas were marked and avoided in subsequent visits.
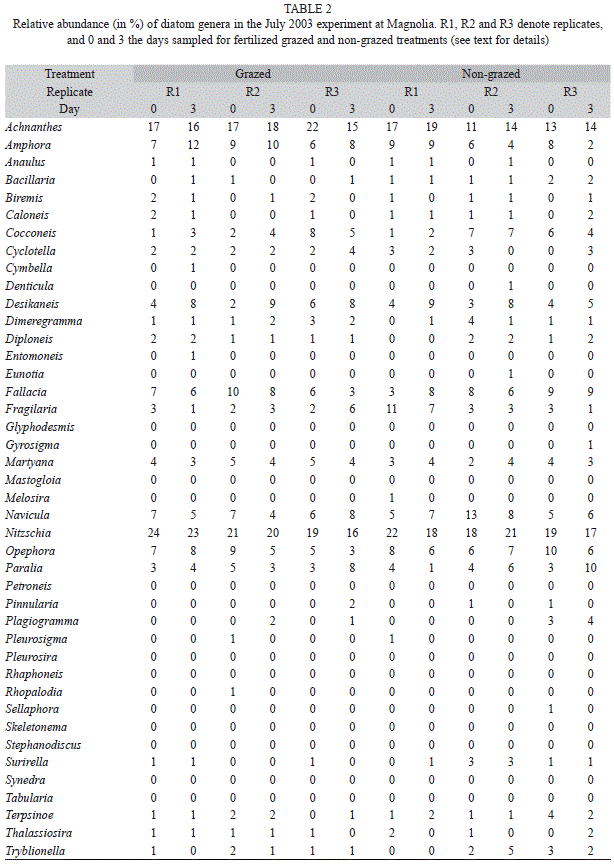
Sediment diatom community structure: On days zero and three of the July 2003 experiments, we took one extra sediment core in each of three randomly-selected fertilized pairs of plots. The cores were extracted with the same type of tube used for the chlorophyll a samples and the top 0.5cm of sediment transferred to a 20mL glass vial. The samples were covered with Lugol’s solution, fixed with 4% buffered formalin, placed on ice, brought back to the laboratory and stored in the refrigerator. During processing, the samples were digested with nitric acid in the presence of potassium dichromate and rinsed with distilled water. Sediment and diatom frustules were suspended through agitation and the supernatant suctioned to remove as much sediment as possible while preserving the diatom frustules in the sample. This process was reiterated until microscopic examination verified that diatoms frustules had been cleansed from most of the sediment (Parsons 1996).
Samples were then diluted 1/10 with distilled water and one drop of the diluted sample was suspended on a cover slip. The cover slips were air-dried slowly to achieve even settlement of the diatom frustules. Each cover slip was then upended onto a drop of Naphrax mounting medium put on a slide, and the slides heated briefly to produce a mount with a high diffraction index. For each sample (i.e. slide), the frustules in one full transect across the cover slip (i.e. from edge to edge of the slip) were counted and identified to the genus level. We completed as many full transects as necessary to identify at least 300 frustules per sample (Gesteira et al. 2003).
Potential side effects of the cages: Enclosure devices, such as the cages used here, may have a series of unwanted (i.e. spurious) impacts that may mask the effect of the factors researchers intend to test for. We examined three potential spurious impacts of the cages used: light attenuation, reduction of water flow, and alteration of the infaunal macro invertebrate community. When totally clean, the acrylic material that made up the cages absorbed ca. 5% of incident light. Since we cleaned up the cages on each sampling day, light attenuation by the cages should be inconsequential for our results.
We measured chlorine dissolution rates to estimate the reduction of water flow by the cages. On day zero, one pre-weighed chlorine tablet was secured onto the sediment inside each of three additional cages and three additional open plots adjacent to the experimental array. The tablets were located at the center of the cages or open plots. They were collected at the end of the experiment, carefully brought back the laboratory, air-dried and reweighed.
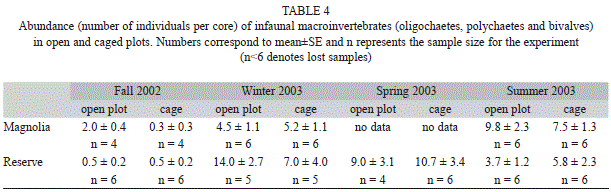
Additional sediment cores were taken to examine whether the community of infaunal macroinvertebrates differed between cages and open plots. The corer was a 7.6cm diameter PVC pipe that was buried to a depth of ca. 15cm. One core was taken in each of three randomly selected fertilized and nonfertilized pairs of plots on day three of each experiment. The samples were placed on ice, brought back to the laboratory and frozen at -4oC until processing. During processing, the sample was rinsed through a 500μm mesh sieve and the invertebrates remaining on the sieve counted and identified as oligochaetes, polychaetes or bivalves.
Hydrographic conditions: Water temperature, salinity and dissolved oxygen were measured with YSI Model 85 meter (Yellow Springs Instrument Co., Yellow Springs, OH, USA) on each sampling day just beneath the water surface and just above the sediment nearby the experimental plots. Water column nutrient (nitrate, nitrite, ammonium and phosphate) and chlorophyll-a concentrations were measured at the beginning and end of each experiment. Samples were taken at mid watercolumn nearby the experimental area, stored on ice and brought back to the laboratory for analysis. Samples were filtered through 0.45μm glass fiber filters, the filtrate collected in duplicate Nalgene bottles, and filters and bottles stored at -80oC until processing.Chlorophyll-a concentration was measured according to the fluorometric method of Parsons et al. (1984). Water column nutrient concentrations were analyzed according to standard colorimetric methods (Strickland & Parsons 1972) using a Skalar SAN+ Autoanalyzer. All these hydrographic data were collected as part of another study carried out simultaneously in nearby areas (Stutes et al. 2006) and are reported in table 2 of that study.
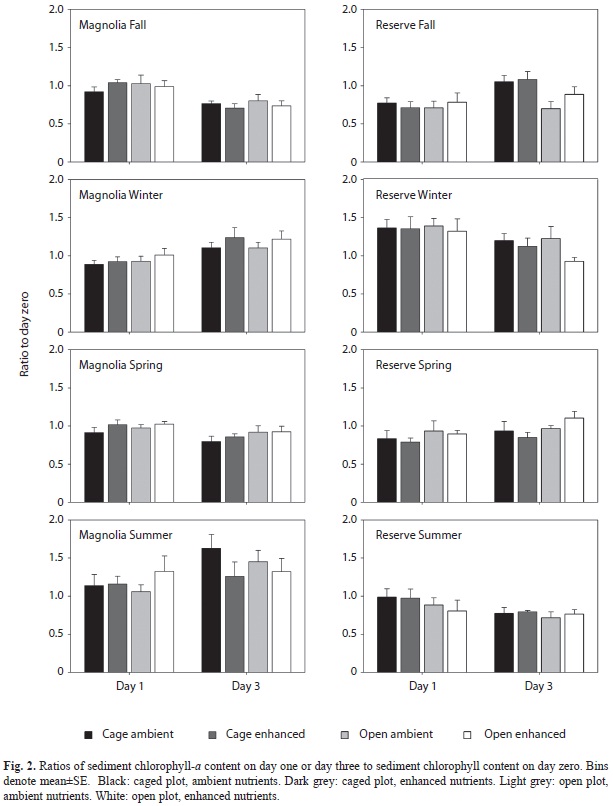
With statistical analysis we found substantial variability in initial sediment chlorophyll-a content among the different treatments-to-be before we actually started administering the treatments (i.e. calculating the mean value±SE for the five replicate plots per treatment, once the three samples in the plot had been averaged, and comparing the mean values±SE’s among treatments on day zero before the plots were fertilized and the cages set up). Thus, to ensure that these initial ambient differences among treatments-to-be did not mask the effects of enhanced nutrient availability and removal of grazing by N. reclivata, we calculated the ratio of chlorophyll-a content on day one or three to chlorophyll-a content on day zero for each replicate, with the content values corresponding to the average of the three samples obtained for the replicate on the given day. We then examined the impact of nutrient enrichment and grazing by N. reclivata with a two-way ANOVA on the ratio values for each sampling time (day one or three) and experiment, for a total of 16 ANOVA’s (two days per experiment x eight experiments). We used t-tests to compare chlorine dissolution rates and the total abundance of major groups of infaunal macroinvertebrates between cages and open plots for each experiment. All data employed in the ANOVA and t-tests were tested for normality and homogeneity of variance and, when necessary, transformed to meet these requirements.
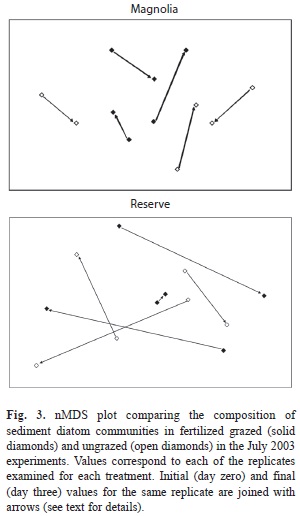
Changes in sediment diatom community structure over the course of the experiment were analyzed with the PRIMER statistical package (Clarke & Warwick 2001). We built the dissimilarity matrix using the Bray-Curtis index after square-rooting the relative abundances. In comparison with other indices, this index minimizes the impact of outliers while retaining high sensitivity (Bloom 1981, Ludwig & Reynolds 1988). We then plotted the data using techniques of non-metric multidimensional scaling (nMDS) and compared the resemblance of the diatom community between caged and open plots at the beginning (day zero) and end (day three) of the experiment using analysis of similarity (ANOSIM). This analysis allowed us to examine whether shifts in the structure of sediment diatom communities occurred over a four-day period in the fertilized plots as a result of grazing by N. reclivata.
Results
Hydrographic conditions: Water-column temperature and salinity showed clear seasonal oscillations, with salinity being lower during the spring and summer experiments (Table 2, Stutes et al. 2006). Dissolved oxygen concentrations in the water column were elevated (generally >7mg/L), tending to be highest during the winter experiments. Water-column chlorophyll-a and nutrient concentrations featured values typical of estuaries in the Northern Gulf of Mexico (Bianchi et al. 1999).
Sediment chlorophyll-a content: In relation to the values found before applying our treatments, sediment chlorophyll-a content did not vary largely as the experiment progressed. Indeed, most of the ratios of contents on days one or three to the content on day zero were close to one (Fig. 2). Interestingly, we found almost no significant impacts of nutrient enrichment and grazing by N. reclivata on that ratio. All the ANOVA’s displayed non-significant F values for the main effects and interaction of fertilization and grazing (p>0.05), except for a significant main effect of N. reclivata grazing on day three of the Fall 2002 experiment at Reserve where lower ratios were found in grazed plots in comparison with ungrazed plots (p<0.05).
Mean (±SE) values of sediment chlorophyll-a content were (in mg/m2): October 2002 experiment at Magnolia (all treatments and all days pooled together): 452.7±12.9; October 2002 experiment at Reserve: 36.98±1.14; February 2003 experiment at Magnolia: 295.17±6.56; February 2003 experiment at Reserve: 70.28±2.08; April 2003 experiment at Magnolia: 385.1±7.36; April 2003 experiment at Reserve: 43.27±1.05; July 2003 experiment at Magnolia: 176.53±6.20; July 2003 experiment at Reserve: 34.86±1.01.
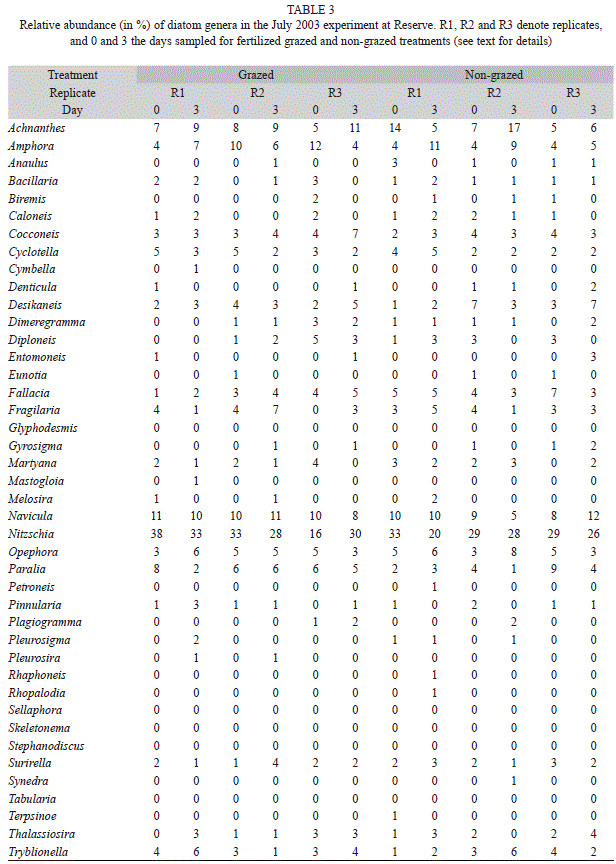
Sediment diatom community structure: Achnanthes, Amphora, Navicula and Nitzschia were the most abundant genera in the two locations at the time when the sediment diatom community was examined (July 2003). Together they accounted for ca. 40 to 60% of the genera found in any given replicate (Tables 2 and 3). Other common genera in both locations were Cocconeis, Cyclotella, Desikaneis, Fallacia, Fragilaria, Martyana, Opephora and Paralia.
As it can be inferred from tables 2 and 3, large differences in sediment diatom community structure were found among the three replicate plots, both for the grazed and ungrazed treatments, in the two study locations before starting the July 2003 experiment (i.e., samples taken on day zero; Fig. 3). The direction of the shift in community structure observed through the experiment did not follow any clear trends, neither among replicates of the same treatment nor between treatments, in any of the two locations. As a consequence, we did not find any significant differences in the structure of the sediment diatom community between grazed and ungrazed plots at days zero and three (Fig. 3; ANOSIM, one-way comparison between grazed plots at day zero, ungrazed plots at day zero, grazed plots at day three, and ungrazed plots at day three, p=0.60 for Magnolia, p=0.88 for Reserve).
Chlorine dissolution rates: Chlorine dissolution rates varied substantially among experiments and, in general, the location at Magnolia displayed higher dissolution rates than the location at Reserve (Fig. 4). Chlorine dissolution rates were significantly higher within open plots than within cages in three experiments (October 2002 at Magnolia, and February 2003 and May 2003 at Reserve; t-test comparing mean dissolution rates between open plots and cages for each experiment, p<0.05) out of the eight ones carried out.
Infaunal macroinvertebrate abundance: The total abundance of infaunal macroinvertebrates (oligochaetes+polychaetes+bivalv es) collected with the 7.6cm diameter corer showed large variability among experiments, with no clear trend between locations (Table 4). We only found significant differences between open and caged plots in the Fall 2002 experiment at Magnolia (t-test comparing mean abundance between open and caged plots, p<0.05) out of the seven experiments for which infaunal data were available (Table 4).
Discussion
With our fertilization procedure we made an effort to achieve realistic (i.e. within the range of fertilization values applied elsewhere and plausible in our study site following further human development of the watershed, Worm et al. 2000, Anton et al. 2011) and stochiometrically-balanced (i.e. having a 17:1 N to P atomic ratio, which is the internal ratio of benthic microalgae at optimal growth, Hillebrand & Sommer 1999) nutrient enrichment. We also applied enough nutrients to ensure persistent high concentrations (at least for nitrate+nitrite) through the duration of the experiments (i.e. four days). Yet, we found no significant effects of nutrient enrichment on sediment chlorophyll-a content in any of the eight experiments. These results suggest that short-term nutrient pulses should have little impact, if any, on the biomass of sediment microalgae at the locations studied.
Past experiments have shown that nutrient enrichment leads to higher benthic microalgal chlorophyll-a content in some instances (Hillebrand & Sommer 1997, 2000a, Lever &Valiela 2005, Cebrian et al. 2009), but not in others (Hillebrand & Kahlert 2002, Stutes et al. 2006, Cebrian et al. 2009). Potential reasons as to why nutrient addition may not result in increased sediment chlorophyll-a content include severe light limitation of photosynthesis (Barranguet et al. 1998, Meyercordt & Meyer-Reil 1999), ambient nutrient concentrations in the sediment that are saturating for growth/photosynthesis (Hillebrand & Kahlert 2002, Stutes et al. 2006), and loss due to horizontal advection and/or grazing (Blanchard et al. 2001, Cebrian 2004). Weeks Bay is a eutrophic estuary with poor light availability and high porewater nutrient concentrations in the deeper (>0.5m) reaches of the estuary (Schreiber & Pennock 1995, Pennock et al. 2001, Lehrter 2006). Stutes et al. (2006) carried out a number of fertilization experiments in areas nearby our study locations, but which were slightly deeper (>0.5m), and found no impact on the productivity of sediment microalgae. They attributed this to the limiting-light levels and saturating-nutrient levels for microalgal growth measured in the two areas. The locations studied here were adjacent to fringing marshes and shallower (most often <0.5m) than the areas examined by Stutes et al. (2006). We did not measure light irradiance at the sediment surface in our study locations, but based on depth and bottom light irradiance measurements in the areas studied by Stutes et al. (2006) bottom light irradiance in our study locations most likely exceeded 500µmol/m2.s photons during our experiments. These estimates appear to lie well over the range of published values of saturating light irradiance for benthic microalgal growth (i.e. 300 to 500µmol/m2.s photons; Pinckney & Zingmark 1991, 1993, Blanchard & Montagna 1992, Blanchard & Gall 1994, Wolfstein & Hartig 1998). This suggests bottom irradiance was not limiting for sediment microalgae in our study locations and probably not a reason why we did not find any fertilization impacts on their biomass.
The ambient sediment porewater nutrient concentrations found in our study locations may help explain why we did not find an effect of further nutrient addition. In Magnolia, concentrations of dissolved inorganic nitrogen (NH4+NO3+NO2) ranged 16.0-602.1μM (median=51.6μM) and concentrations of phos-phate ranged 0.1-8.3μM (median=1.1μM) in the sediment porewater of non-fertilized plots on day zero. In Reserve, those concentrations ranged 3.00-393.1μM (median=57.2μM) for dissolved inorganic nitrogen and 0.05-29.44μM (median=0.37μM) for phosphate. These values are high when compared with those measured in other coastal environments (Worm et al. 2000). Perhaps more importantly, relationships between nutrient availability and growth of benthic microalgae developed in the laboratory (Smayda 1997) suggest that the ambient sediment porewater nutrient concentrations in our study locations, with perhaps the exception of phosphate concentrations at Reserve, are saturating, or near-saturating at a minimum, for sediment microalgal growth given adequate conditions of light and temperature. If this is the case, adding extra nutrients for a few days would probably have no major impact on sediment microalgal biomass, as observed with our experiments. Other investigations have found significant short-term (i.e. within a few days since nutrient addition) responses of sediment microalgae to fertilization given poor ambient nutrient availability and adequate levels of light and temperature (e.g. Downing et al. 1999, Wulff et al. 2000, Cebrian et al. 2009), but our sediments appear to be saturated in nutrients for microalgal growth. However, prolonged nutrient dosage in our locations could certainly have indirect impacts on sediment microalgae through, for instance, changes in population structure and/or stimulation of phytoplankton growth and subsequent competition for light and nutrients (Hillebrand et al. 2000, Hillebrand & Sommer 2000b, Cloern 2001).
Resuspension and redistribution of sediment microalgae could also help explain why we do not find major impacts of nutrient addition (Cahoon 1999, Blanchard et al. 2001). The locations studied are shallow (<0.6m), and the sediment and inhabiting microalgae are often stirred up. Thus frequent resuspension and subsequent re-deposition could mask a significant effect of nutrient addition. Whatever the reason may be, here we report that short-term nutrient addition in shallow sediments adjacent to fringing marsh in Weeks Bay does not have a noticeable impact on the biomass of inhabiting microalgae, despite high light availability in those shallow sediments. The unresponsiveness of sediment microalgae to short-term nutrient addition may be a common feature in Weeks Bay.
We found little evidence for a significant role of grazing by N. reclivata on the biomass of microalgae in bare sediment adjacent to fringing marsh. Namely, out of 16 possible instances (i.e. eight experiments x two sampling days per experiment), we solely found a significant reduction in sediment chlorophyll-a content in open plots in comparison with caged plots on day three of the Fall 2002 experiment at Reserve. In addition, we did not find any significant impacts of exposure to the snail on the composition of the sediment diatom community in the two experiments where we tested for those effects (experiments at Reserve and Magnolia in July 2003). Shifts in diatom community composition were observed over the course of the experiments, but those shifts were independent of whether the plot was open or not to N. reclivata. Other than preventing access by the snail, our exclosures did not appear to have any major undesired (i.e. spurious) direct or indirect effects over the four days that the experiments lasted. Light interception by the exclosure was minimal (i.e., five % of incident light), chlorine dissolution rates (a proxy for flow rates and potential sediment resuspension) only differed significantly between open and caged plots in three out of the eight experiments, and the exclosure did not apparently change the abundance of major groups of infaunal macroinvertebrates.
Research done with other species of gastropod grazers has shown variable impacts that may range from negligible to large fractions of benthic microalgal biomass consumed (Pace et al. 1979, Branch & Branch 1980, Bennett et al. 1999, Blanchard et al. 2000). The low densities of N. reclivata recorded in our study locations (i.e. bare sediment adjacent to fringing marshes) in the fall, spring and summer experiments (mean±SE for the ten quadrats ranging from 0±0 to 8.0±3.6 snails/m among the six experiments) may explain the almost complete lack of significant impacts by the snail in these experiments. We recorded higher densities in the winter experiments (91.2±23.1 snails/m at Reserve; 17.6±6.5 snails/m at Magnolia). Potential reasons for higher densities on bare sediment adjacent to fringing marshes in winter in relation to other seasons are unclear, as few studies exist on the snail’s behavior and population dynamics (Frankael 1968, Lehman & Hamilton 1980). At any rate, the snail does not feed effectively on sediment microalgae (Lehman & Hamilton 1980) and this, in conjunction with the short duration of the experiments (i.e. four days), could help explain the lack of significant impacts by the snail found in the winter experiments. It seems unlikely a longer experiment could have led to more significant impacts by the snail in the others seasons given its extremely low densities during those seasons. These results are consistent with our hypotheses of low impact by N. reclivata on microalgae inhabiting bare sediment adjacent to vegetated stands due to low abundance of the snail in those habitats and inefficient grazing on the microalgae.
Overall our results suggest that ephemeral, short-term nutrient pulses into eutrophic coastal systems of the Northern Gulf of Mexico, such as Weeks Bay (Alabama, USA), should not affect greatly the abundance of sediment microalgae. For instance, based on our results we would expect that occasional nutrient inputs due to runoff would not drastically alter the standing stock of sediment microalgae in the locations studied. The impacts of short-lived pulses should be further lessened in deeper areas of eutrophic systems, which, unlike the well-lit locations studied here, typically feature severe light scarcity (Schreiber & Pennock 1995, Meyercordt & Meyer-Reil 1999, Stutes et al. 2006). Persistent nutrient delivery, however, could likely affect sediment microalgal standing stocks through mechanisms such as changes in the identity of dominant species (Hillebrand et al. 2000, Hillebrand & Sommer 2000b), and/or increased shading due to the buildup of phytoplankton and filamentous macroalgae (Cloern 2001, Hauxwell et al. 2001). Similarly, our results suggest the snail N. reclivata is not a major control of sediment microalgal populations in the subtidal sedimentary areas studied. Perhaps longer experiments in winter would have revealed significant effects, but it seems that overall the impact by the snail is modest at best in those areas.
Clearly the impacts of short-term nutrient pulses and snail grazers on sediment microalgae seem to be context-dependent. The results reported here may perhaps apply to other eutrophic, subtidal sediments with low ambient densities of this and other snail grazers, but the results would have likely been qualitatively different if we had done the same experiments in well-lit, nutrient poor sediments, or in sediments of highly vegetated intertidal or subtidal areas with much higher snail densities. In conjunction this and other investigations indicate that the response of sediment microalgae to nutrient enrichment and modified grazer abundance depends to a large extent on the initial levels of nutrient availability and grazing before the system is altered.
Acknowledgments
We thank M.J. Sullivan for assistance and valuable comments during the project. L. Linn provided technical assistance. R. Moody and H. MacIntyre provided valuable insights. L. G. Adams and the staff of Weeks Bay NERR helped with the field work. This research was supported by a grant from the Alabama Center for Estuarine Studies (ACES# 5-21828), as well as NOAA NERRS grant # NA17OR1177.
References
Anton, A., J. Cebrian, K.L. Heck, C.M. Duarte, K.L. Sheehan, M.E.C. Miller & C.D. Foster. 2011. Decoupled effects (positive to negative) of nutrient enrichment on ecosystem services. Ecol. Appl. 21: 991-1009. [ Links ]
Barranguet, C., J. Kromkamp & J. Peene. 1998. Factors controlling primary production and photosynthetic characteristics of intertidal microphytobenthos. Mar. Ecol. Prog. Ser. 173:117-126. [ Links ]
Bennet, A., T.S. Bianchi, J.C. Means & K.R. Carman. 1999. The effects of polycyclic aromatic hydrocarbon contamination and grazing on the abundance and composition of microphytobenthos in salt marsh sediments (Pass Fourchon, LA) I. A microcosm experiment. J. Exp. Mar. Biol. Ecol. 242: 1-20. [ Links ]
Bianchi, T.S., J. Pennock & R. Twilley. 1999. Biogeochemistry of Gulf of Mexico Estuaries. Wiley & Sons, New York, New York, USA. [ Links ]
Blanchard, G.F. & P.A. Montagna. 1992. Photosynthetic response of natural assemblages of marine benthic microalgae to shortand long-term variations of incident irradiance in Baffin Bay, Texas. J. Phycol. 28: 7-14. [ Links ]
Blanchard, G.F. & V.C. Gall. 1994. Photosynthetic characteristics of microphytobenthos on Marennes-Oleron Bay, France: preliminary results. J. Exp. Mar. Biol. Ecol. 182: 1-14. [ Links ]
Blanchard, G.F., J.M. Guarini, L. Provot, P. Richard & P.G. Sauriau. 2000. Measurement of ingestion rate of Hydrobiaulvae (Pennant) on intertidal epipelic microalgae: the effect of mud snail density. J. Exp. Mar. Biol. Ecol. 255: 247-260. [ Links ]
Blanchard, G.F., J.M. Guarini, F. Orvain & P.G. Sauriau. 2001. Dynamic behaviour of benthic microalgal biomass in intertidal mudflats. J. Exp. Mar. Biol. Ecol. 264: 85-100. [ Links ]
Bloom, S.A. 1981. Similarity indices in community studies: potential pitfalls. Mar. Ecol. Prog. Ser. 5: 125-128. [ Links ]
Branch, G.M. & M.L. Branch. 1980. Competition in Bembiciumauratum (Gastropoda) and its effect on microalgal standing stock in mangrove mudflats. Oecologia 46: 106-114. [ Links ]
Carman, K.R. & B. Fry. 2002. Small-sample methods for delta13 C and delta15 N analysis of the diets of marsh meiofaunal species using natural-abundance and tracer-addition isotope techniques. Mar. Ecol. Prog. Ser. 240: 85-92. [ Links ]
Cahoon, L.B. 1999. The role of benthic microalgae in neritic ecosystems. Ann. Rev. Ocean. Mar. Biol. 37: 47-86. [ Links ]
Clarke, K.R. & R.M. Warwick 2001. Change in marine communities: an approach to statistical analysis and interpretation. PRIMER-E, Plymouth. [ Links ]
Cebrian, J. 2004. Grazing on benthic primary producers, p. 153-185. In S.L. Nielsen, G.T. Banta & M.F. Pedersen. Estuarine nutrient cycling: the influence of primary producers. Kluwer Academic, Dordrecht, Boston, London. [ Links ]
Cebrian, J., C.D. Foster, R. Plutchak, K.L. Sheehan, M.E. Miller, A. Anton, K. Major, K. L. Heck & S.P. Powers. 2008. The impact of Hurricane Ivan on the primary productivity and metabolism of marsh tidal creeks in the North Central Gulf of Mexico. Aquat. Ecol. 42: 391-404. [ Links ]
Cebrian, J., A.A. Corcoran, A.L. Stutes, J.P. Stutes & J.R. Pennock. 2009. Effects of ultraviolet-B radiation and nutrient enrichment on the productivity of benthic microalgae in shallow coastal lagoons of the North Central Gulf of Mexico. J. Exp. Mar. Biol. Ecol. 372: 9-21. [ Links ]
Cloern, J.E. 2001.Our evolving conceptual model of the coastal eutrophication problem. Mar. Ecol. Prog. Ser. 210: 223-253. [ Links ]
Darley, W.M., C.L. Montague, F.G. Plumley, W.W. Sage & A.T. Psalidas. 1981. Factors limiting edaphic algal biomass and productivity in a Georgia salt marsh. J. Phycol. 17: 122-128. [ Links ]
de Jonge, V.N. & F. Colijn. 1994. Dynamics of microphytobenthos biomass in the Ems estuary. Mar. Ecol. Prog. Ser. 104:185-196. [ Links ]
Downing, J.A., C.W. Osenberg & O. Sarnelle. 1999. Metaanalysis of marine nutrient-enrichment experiments: variation in the magnitude of nutrient limitation. Ecology 80: 1157-1167. [ Links ]
Fenchel, T. & L.H. Kofoed. 1976. Evidence for exploitative interspecific competition in mudsnails (Hydrobidae). Oikos 27: 367-376. [ Links ]
Frankael, G. 1968. The heat resistance of intertidal snails at Bimini, Bahamas; Ocean Springs, Mississippi, and Woods Hole. Massachusetts. Phys. Zool. 41: 1-13. [ Links ]
Gainey, L.F. 1976. Locomotion in the Gastropoda: Functional morphology of the foot in Neritina reclivata and Thais rustica. Malacologia 15: 411-431. [ Links ]
Gesteira, J.L.G., J.C. Dauvin & M.S. Fraga. 2003. Taxonomic level for assessing oil spill effects on soft-bottom sublittoral benthic communities. Mar. Poll. Bull. 46: 562-572. [ Links ]
Hauxwell, J., J. Cebrian, C. Furlong & I. Valiela. 2001. Macroalgal canopies contribute to eelgrass (Zostera marina) decline in temperate estuarine ecosystems. Ecology 82: 1007-1022. [ Links ]
Heck, K.L., J.F. Valentine, J.R. Pennock, G. Chaplin & P.M. Spitzer. 2006. Effects of nutrient enrichment and grazing on shoalgrass (Halodule wrightii) and its epiphytes: results of a field experiment. Mar. Ecol. Prog. Ser. 326: 145-156. [ Links ]
Hillebrand, H. & U. Sommer. 1997. Response of epilithic microphytobenthos of the Western Baltic Sea to in situ experiments with nutrient enrichment. Mar. Ecol. Prog. Ser. 160: 35-46. [ Links ]
Hillebrand, H. & U. Sommer. 1999. The nutrient stoichiometry of benthic microalgal growth: Redfield proportions are optimal. Limnol. Oceanogr. 44: 440-446. [ Links ]
Hillebrand, H. & U. Sommer. 2000a. Effects of continuous nutrient enrichment on microalgae colonizing hard substrates. Hydrobiologia 426: 185-192. [ Links ]
Hillebrand, H. & U. Sommer 2000b. Diversity of benthic microalgae in response to colonization time and eutrophication. Aquat. Bot. 67: 221-236. [ Links ]
Hillebrand, H., B. Worm & H.K. Lotze. 2000. Marine microbenthic community structure regulated by nitrogen loading and grazing pressure. Mar. Ecol. Prog. Ser. 204: 27-38. [ Links ]
Hillebrand, H. & M. Kahlert. 2002. Effect of grazing and water column nutrient supply on biomass and nutrient content of sediment microalgae. Aquat. Bot. 72: 143-159. [ Links ]
Lehman, H.K. & P.V. Hamilton. 1980. Some factors influencing the distribution of the snail Neritinareclivata. Northeast Gulf Sci. 4: 67-72. [ Links ]
Lehrter, J.C. 2006. Effects of Land-Use and Land-Cover, Stream Discharge, and Interannual Climate on the Magnitude and Timing of Nitrogen, Phosphorus, and Organic Carbon Concentrations in Three Coastal Plain Watersheds. Water Environ. Res. 78: 2356-2368. [ Links ]
Lever, M.A. & I. Valiela. 2005. Response of microphytobenthic biomass to experimental nutrient enrichment and grazer exclusion at different land-derived nitrogen loads. Mar. Ecol. Prog. Ser. 294: 117-129. [ Links ]
Levinton, J.S. & T.S. Bianchi. 1981. Nutrition and food limitation of deposit-feeders. I. The role of microbes in the growth of mud snails (Hydrobiidae). J. Mar. Res. 39: 531-545. [ Links ]
Lotze, H.K., H.S. Lenihan, B.J. Bourque, R.H. Bradbury, R.G. Cooke, M.C. Kay, S.M. Kidwell, M.X. Kirby, C.H. Petterson & J.B.C. Jackson. 2006. Depletion, Degradation and Recovery Potential of Estuaries and Coastal Seas. Science 312: 1806-1809. [ Links ]
Ludwig, J.A. & J.F. Reynolds 1988. Statistical ecology: a primer on methods and computing. Wiley and Sons, New York, USA. [ Links ]
Meyercordt, J. & L.A. Meyer-Reil. 1999. Primary production of benthic microalgae in two shallow coastal lagoons of different trophic status in the Southern Baltic Sea. Mar. Ecol. Prog. Ser. 178:179-191. [ Links ]
Middleburg, J.J., C. Barranguet, H.T.S. Boschker, P.M.J. Herman, T. Moens & C.H.R. Heip. 2000. The fate of intertidal microphytobenthos carbon: an in-situ 13C-labeling study. Limnol.Oceanogr. 45:1224-1234. [ Links ]
Miller, D.C., R.J. Geider & H.L. MacIntyre. 1996. Microphytobenthos: The ecological roles of the “Secret Garden” of unvegetated, shallow-water marine habitats. II. Role in sediment stability and shallow-water food webs. Estuaries 19: 202-212. [ Links ]
Miller-Way, T., M. Dardeau & G. Crozier. 1996. Weeks Bay National Estuarine Research Reserve: an estuarine profile and bibliography. Tech. Rep. 96-01. Dauphin Island Sea Laboratory, Dauphin Island, Alabama, USA. [ Links ]
Moncreiff, C.A., M.J. Sullivan & A.E. Daehnick. 1992. Primary production dynamics in seagrass beds of Mississippi Sound: the contributions of seagrass, epiphytic algae, sand microflora, and phytoplankton. Mar. Ecol. Prog. Ser. 87:161-171. [ Links ]
Orth, R.J., T.J.B. Carruthers, W.C. Dennison, C.M. Duarte, J.W. Fourqurean, K.L. Heck, A.R. Hugues, G.A. Kendrick, W.J. Kenworthy, S. Olyarnik, F.T. Short, M. Waycott & S.L. Williams. 2006. A global crisis for seagrass ecosystems. Bioscience 56: 987-996. [ Links ]
Pace, M.I., S. Shimmel & W.M. Darley. 1979. The effect of grazing by a gastropod, Nassarius obsoletus, on the benthic microalgal community of a salt marsh mudflat. Estuar. Coast. Mar. Sci. 9:121-134. [ Links ]
Parsons, M.L. 1996. Paleoindicators of changing water conditions in Louisiana estuaries. Ph.D. Dissertation. Louisiana State University, Baton Rouge, Louisiana, USA. [ Links ]
Parsons, T.R., Y. Maita & C.M. Lalli. 1984. A manual of chemical and biological methods for seawater analysis. Pergamon, Oxford, England. [ Links ]
Pennock, J.R., J.H. Cowan, K.M. Shotts, J.L.W. Cowan & L.J. Gallagher. 2001. Weeks Bay data report: WBAY-2 to WBAY-56 Cruises (May 1996-May 2000). Tech. Rep. 2001-001. Dauphin Island Sea Laboratory, Dauphin Island, Alabama, USA. [ Links ]
Pinckney, J.L. & R.G. Zingmark. 1991. Effects of tidal stage and sun angles on intertidal benthic microalgal productivity. Mar. Ecol. Prog. Ser. 76: 81-89. [ Links ]
Pinckney, J.L. & R.G. Zingmark. 1993. Photophysical responses of intertidal benthic microalgal communities to in situ light environments: methodological considerations. Limnol. Oceanogr. 38: 1373-1383. [ Links ]
Rizzo, W.M., G.J. Lackey & R.R. Christian. 1992. Significance of euphotic, subtidal sediments to oxygen and nutrient cycling in a temperate estuary. Mar. Ecol. Prog. Ser. 86: 51-61. [ Links ]
Russell, H.D. 1941. The recent mollusks of the Family Neritidae of the Western Atlantic. Bull. Mus. Comp. Zool. 88: 347-403. [ Links ]
Sheridan, P.F. & R.J. Livingston. 1983. Abundance and seasonality of infauna and epifauna inhabiting a Halodule wrightii meadow in Apalachicola Bay, Florida. Estuaries 6: 407-419. [ Links ]
Schreiber, R.A. & J.R. Pennock. 1995. The relative contribution of benthic microalgae to total microalgal production in a shallow sub-tidal estuarine environment. Ophelia 42: 335-352. [ Links ]
Schroeder, W.W., W.J. Wiseman, Jr. & S.P. Dinnel. 1990. Wind and river induced fluctuations in a small, shallow tributary estuary, p. 481-493. In R.T. Cheng. Residual Currents and Long-Term Transport. Coastal and Estuarine Studies, Springer Verlag, New York, New York, USA. [ Links ]
Smayda, .T.J. 1997. Harmful algal blooms: their ecophysiology and general relevance to phytoplankton blooms in the sea. Limnol. Oceanogr. 42:1137-1153. [ Links ]
Strickland, J.D.H. & T.R. Parsons. 1972. A practical handbook of seawater analysis. Bull. Fish. Res. Board Can. 167:1-310. [ Links ]
Stutes, A.L., J. Cebrian & A.A. Corcoran. 2006. Effects of nutrient enrichment and shading on sediment primary production and metabolism in eutrophic estuaries. Mar. Ecol. Prog. Ser. 312: 29-43. [ Links ]
Stutes, J.P., J. Cebrian, A.L. Stutes, A. Hunter & A.A. Corcoran. 2007. Benthic metabolism across a gradient of anthropogenic impact in three shallow coastal lagoons in NW Florida. Mar. Ecol. Progr. Ser. 348: 55-70. [ Links ]
Sundbäck, K.L., V. Enoksson, W. Grane’Li & K. Pettersson. 1991. Influence of sublittoralmicrophytobenthos on the oxygen and nutrient flux between sediment and water: A laboratory continuous-flow study. Mar. Ecol. Prog. Ser. 74: 263-279. [ Links ]
Wolfstein, K. & P. Hartig. 1998. The photosynthetic light dispensation system: application to microphytobenthic primary production measurements. Mar. Ecol. Prog. Ser. 166: 63-71. [ Links ]
Wolfstein, K. & L.J. Stal. 2002. Production of extracellular polymeric substances (EPS) by benthic diatoms: effect of irradiance and temperature. Mar. Ecol. Prog. Ser. 236: 13-22. [ Links ]
Worm, B., T.B.H. Reusch & H.K. Lotze. 2000. In situ nutrient enrichment: methods for marine benthic ecology. Int. Revue Hydrobiol. 85: 359-375. [ Links ]
Wulff, A., S.A. Wängberg, K. Sundbäck, C. Nilsson & G.J.C. Underwood. 2000. Effects of UVB radiation on a marine microphytobenthic community growing on a sand-substratum under different nutrient conditions. Limnol. Oceanogr. 45: 1144-1152. [ Links ]
*Correspondencia: Just Cebrian: Dauphin Island Sea Lab, 101 Bienville Blvd., Dauphin Island, AL 36528; jcebrian@disl.org. Weeks Bay National Estuarine Research Reserve, 11300 U.S. Highway 98, Fairhope, AL 36532.
Adrienne L. Stutes: Dauphin Island Sea Lab, 101 Bienville Blvd., Dauphin Island, AL 36528. Department of Marine Sciences, University of South Alabama, LSCB 25, Mobile, AL 36688. Present address: Pentec Environmental/Hart Crowser, Inc. 120 Third Ave, So. Edmonds, WA 98020; adrienne.stutes@hartcrowser.com
Scott Phipps: Weeks Bay National Estuarine Research Reserve, 11300 U.S. Highway 98, Fairhope, AL 36532; scott.phipps@dcnr.alabama.gov
Jason P. Stutes: Dauphin Island Sea Lab, 101 Bienville Blvd., Dauphin Island, AL 36528. Department of Marine Sciences, University of South Alabama, LSCB 25, Mobile, AL 36688. Present address: Pentec Environmental/Hart Crowser, Inc. 120 Third Ave, So. Edmonds, WA 98020; jason.stutes@hartcrowser.com
Bart Christiaen: Dauphin Island Sea Lab, 101 Bienville Blvd., Dauphin Island, AL 36528; bchristiaen@disl.org. Department of Marine Sciences, University of South Alabama, LSCB 25, Mobile, AL 36688.
Jonathan R. Pennock: University of New Hampshire, 24 Colovos Road, Durham, NH 03824-3505; jonathan.pennock@unh.edu
1. Dauphin Island Sea Lab, 101 Bienville Blvd., Dauphin Island, AL 36528; jcebrian@disl.org, bchristiaen@disl.org
2. Department of Marine Sciences, University of South Alabama, LSCB 25, Mobile, AL 36688.
3. Weeks Bay National Estuarine Research Reserve, 11300 U.S. Highway 98, Fairhope, AL 36532; scott.phipps@dcnr.alabama.gov
4. University of New Hampshire, 24 Colovos Road, Durham, NH 03824-3505; jonathan.pennock@unh.edu
5. Present address: Pentec Environmental/Hart Crowser, Inc. 120 Third Ave, So. Edmonds, WA 98020; adrienne.stutes@hartcrowser.com, jason.stutes@hartcrowser.com
Received 13-X-2011. Corrected 23-III-2012. Accepted 23-IV-2012.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}