










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.60 n.3 San José Sep. 2012
Comportamiento reproductivo del pez Opistognathus rosenblatti (Perciformes: Opistognathidae) en cautiverio
Abstract
Reproductive behavior of Opistognathus rosenblatti (Perciformes: Opistognathidae) in captivity. The Blue Spotted Jawfish O. rosenblatti, is an endemic species from the Gulf of California, included in the local list of protected species. With few biological reports, this species is appreciated in the aquarium industry due to its coloration and digging behaviour, and has a considerable value. With the aim to generate valuable biological information, eight fishes were caught at Loreto Natural Marine Protected Area. Captured fishes were juveniles, and just three of them were kept in an aquarium conditioned with gravel, pieces of shells and coral as substrata. Temperature and photoperiod conditions were stable, and they were supplied with a variety of live and inert feeds. Fishes reached maturity in eight months, according to literature reports, displaying mature male courtship coloration and upward movements in the water column. Fishes spawned several times over more than two years in captivity. Presences of egg masses, or some evidence of egg shell or larvae, were registered in 50 occasions. Male took care of eggs in his shelter, but never in his mouth. Egg masses had an average of 3 592 eggs, with a hatching rate close to 99%. Eggs were apparently rounded having three diameter measurements with significant differences (1.17mm mean higher diameter, SD=0.054; 1.13mm mean lower diameter, 0.058; 0.99mm mean height, 0.045; n=125). Most of corion eggs had four, rarely six filaments; with a single oil drop (0.30mm mean diameter, 0.021, n=59). Incubation lasted 10.4 days (9-14), depending on water temperature (21.0-25.3°C). Egg hatching occurred after darkness, emerging newly hatched larvae of 4.51mm mean notochord length (0.082, n=30), with reserves exhausted, eyes pigmented and mouth opened, ready to eat. This study represents the first report on this species courtship displaying, spawning and some basic characteristics of eggs masses and larvae in captivity. Also, their flexibility and adaptability of individual behaviour to particular environment conditions, lead to consider worth their commercial culture, as already occur with other species of this family.
Key words: Opistognathus rosenblatti, jaw fish, ornamental fish, courtship, reproduction, spawning.
Resumen
El bocón manchas azules O. rosenblatti, es una especie endémica del Golfo de California con un elevado valor comercial, del cual existe poca información referente a su biología. Los peces alcanzaron la edad adulta después de ocho meses de cautiverio. Manifestaron ascensos en la columna de agua, típicos de los desplantes de cortejo en su hábitat natural. Asimismo se registro en 50 ocasiones la presencia de masas ovígeras o alguna evidencia de desgove. El macho cuidó los huevos dentro de su refugio, pero nunca en su cavidad bucal. En promedio, depositaron 3 592 huevos, con una viabilidad de eclosión superior al 99%. La eclosión se da de noche y se liberan larvas de 4.51mm de longitud promedio del notocordio. Este estudio presenta el primer registro de su cortejo, su desove e información sobre características elementales de las masas ovígeras y sus larvas, las cuales fueron obtenidas en cautiverio. De tal manera que, la flexibilidad y adaptabilidad de su comportamiento individual a condiciones particulares del ambiente, significan que vale la pena su cultivo comercial, como ya se realiza con otras especies de la misma familia.
Palabras clave: Opistognathus rosenblatti, bocón manchas azules, pez de ornato, cortejo, reproducción, desove.
La información científica referente al comportamiento reproductivo de peces marinos es escasa, y de forma sobresaliente la que se refiere a especies endémicas ornamentales. La familia Opistognathidae, está integrada por los géneros Opistognathus, Lonchopisthus y Stalix, que agrupa a 78 especies, cuya distribución es circumtropical (Nelson 1996).
Las observaciones realizadas en la naturaleza y en cautiverio, evidencian patrones generales de comportamiento y reproducción, coincidentes en señalar un cortejo elaborado y cuidados de la puesta, con incubación bucal por parte de los machos en todos los miembros de la familia (Colin 1973, Thresher 1984, Hess 1993). Es por ello que estos peces han captado la atención de naturalistas, científicos y aficionados al acuarismo marino, por las particularidades que manifiestan durante la construcción de refugios, por sus desplantes de cortejo y atractivo colorido. Las especies de pequeño tamaño pertenecientes a la familia Opistognathidae se acondicionan fácilmente al cautiverio y varias de ellas han logrado reproducirse de manera regular (Thresher 1984), lo que ha propiciado el desarrollo de investigaciones sobre su biología reproductiva, debido al potencial que representa desarrollar su cultivo (Walch 1994, Brons 1995).
En México, aspectos relacionados con la captura y reproducción de especies marinas con fines de ornato, no ha recibido la atención adecuada, como una actividad productiva y generadora de divisas, a pesar del enorme potencial que presenta con base en la elevada biodiversidad existente en sus costas (Flores & Gerez 1992, Anónimo 2004). El pez marino Opistognathus rosenblatti, nombrado regionalmente bocón manchas azules (Anónimo 2010), es endémico del Golfo de California, México (Thomson et al. 1979). Éste habita sobre planicies arenosas en la proximidad de arrecifes rocosos, entre los 18-24m de profundidad, dentro de agujeros a manera de refugios que ellos mismos construyen con una entrada estrecha de manera horizontal, reforzados con fragmentos de roca, coral y concha. El mismo se agrupa en colonias conformadas por pocos hasta cientos de individuos, con separación mínima de un metro entre los refugios (Allen & Robertson 1998).
El bocón manchas azules ha sido capturado como especie de ornato para su comercialización hacia los EEUU, principalmente. El único informe existente sobre la captura de esta especie fue de 713 ejemplares capturados, de un total autorizado de 1 800 organismos en el año 2000 (Piña 2005). A pesar del interés económico que esta especie tiene, con un precio al consumidor final de US $100 por individuo (Shultz III 2002), la información biológica existente es escasa, se limita al conocimiento de su distribución geográfica, coloración reproductiva y cortejo del macho en su hábitat natural. Por varias razones desde el año 2002 es considerada una especie protegida, incluida en la categoría de protección especial de acuerdo con la Norma Oficial Mexicana (D.O.F. 2002), por lo que requiere de permisos especiales para su captura, por parte del Instituto Nacional de Ecología (INE). Junto con el bocón cabeza amarilla del Atlántico (Opistognathus aurifrons), conforman los opistognátidos más solicitados para su manutención en cautiverio (Schultz III 2002). El presente trabajo amplia la información referente al patrón reproductivo de O. rosenblatti, además representa el primer registro de su cortejo y reproducción en cautiverio, los primeros registros de su fecundidad, del periodo de incubación, del tamaño y descripción de las larvas recién nacidas.
Materiales y métodos
Los organismos utilizados para el estudio fueron capturados en la Reserva del Parque Marino de Loreto, Baja California Sur, México, y cedidos en septiembre del año 2000 al Centro Interdisciplinario de Ciencias Marinas del Instituto Politécnico Nacional, por la compañía Marine Export Co. (Permiso de Pesca de Fomento No. 071299-213-03). Asimismo, su captura fue a 10m de profundidad con equipo de buceo autónomo, con una vara como extensión, amarrada en la punta la línea de nylon, con un anzuelo tipo robador (de tres puntas) del No.18. Al colocar el anzuelo dentro del refugios los peces lo engullen para expulsarlo, momento que se aprovecha para levantar al pez y capturarlo con una red de cuchara. De tal manera que se capturaron un total de ocho peces, los cuales se trasladaron en bolsas de plástico con agua, infladas con oxígeno y selladas con banda elástica, y contenidas en un recipiente térmico, por un lapso de 8hr. A su llegada al laboratorio fueron sometidos a un baño con agua dulce durante 15min y fueron colocados en cuarentena durante un mes, dentro de un acuario de 140L de capacidad, forrado con plástico negro, con flujo abierto a razón de 60L/hr de agua de mar, aireación y con tramos de tubo plástico como refugios provisionales. También, se acondicionó un acuario de 480L (152x53x60cm) de capacidad, con filtro de piso, constituido por trozos irregulares de coral pétreo (6cm), grava (2-4cm) y conchuela (0.52cm), con espesor de 4cm, 4cm y 6cm, respectivamente y estructuras decorativas de coral pétreo y conchas de moluscos. Dos elevadores de aire (air-lift) ubicados en extremos opuestos, permitieron la recirculación del agua a través del filtro de piso. La temperatura del agua se mantuvo entre 21.0-26.6°C con un calentador sumergible de 250w (Ebo-Jäger), la misma se registró con un termómetro digital (VWR) y la salinidad con un refractómetro entre 35-42ups (Aquatic Eco-Systems, Inc.). Con regularidad se agregó agua dulce sin cloro para mantenerla estable. Además, se proporcionó iluminación con cuatro lámparas fluorescentes de 39W, activadas por relojes controladores de encendido y apagado, en secuencia con 30min de desfase, para simular condiciones de amanecer y de crepúsculo, con un fotoperiodo de 12 L:12 O (horas de luz:oscuridad). La calidad aparente del agua en el acuario se mantuvo con recambios de 25-40% del volumen cada 60 días, y se removía el sustrato superficial con un sifón para después recuperar el nivel con agua de mar filtrada por cartucho de 30 micras.
Del número inicial de peces jóvenes, sólo tres fueron introducidos al azar (6.5-8.5cm de longitud patrón), alimentados diariamente con alguno o varios de los siguientes componentes de acuerdo con su disponibilidad: Artemia adulta, crías de poecílidos, misidáceos, camarón (Palaemon sp.) y/o peletizado comercial (Otto Marine Blend Premium). Durante tres años de cautiverio, se realizaron observaciones, registros en bitácora, tomas fotográficas y de video. También, se registró la periodicidad de cortejo y desove, las diferencias del patrón de coloración, la fecundidad, el periodo de incubación y el momento de la eclosión. Algunas masas ovígeras fecundadas y expulsadas fuera del refugio del macho, fueron usadas para determinar su fecundidad y su peso, y con algunas de ellas el porcentaje de viabilidad a la eclosión, al colocar la masa ovígera dentro de un tamiz suspendido en un acuario con agua de mar filtrada y esterilizada con UV, a la misma temperatura de los reproductores.
Con un microscopio estereoscópico, el cual contenía una reglilla calibrada, se realizó mediciones del tamaño del huevo y de la larva recién nacida, y con una cámara digital se obtuvieron fotografías de masas ovígeras, embriones y larvas.
Resultados
Construcción de refugios: Inmediatamente después de que los peces fueron introducidos al acuario se manifestaron desconcertados, pero transcurridas un par de horas, se comportaron inquietos, y comenzaron a remover a bocanadas fragmentos de grava y concha para cavar huecos expuestos en diversos sitios, los cuales se colapsaban fácilmente. Comenzaron por acumular los materiales hacia los lados hasta llegar a los fragmentos de coral pétreo de mayor tamaño sin lograr modificar su arreglo. En ese momento la excavación adquirió un aspecto de cráter amplio, en el cual reacomodaron con la boca cada fragmento y lo empujaron con los opérculos para afianzarlo y conformar las paredes del refugio. Asimismo, redujeron el área expuesta hasta construir refugios subterráneos independientes, apegados a las estructuras de coral y concha, que les permitió retener el material acumulado. Los refugios construidos por las dos hembras (reconocidas y diferenciadas entre si después de 11 meses de cautiverio) se ubicaron en triangulación a 46 y 53cm de distancia del macho, y sucesivamente expulsaron con regularidad fragmentos de material, como parte del arreglo y mantenimiento de sus refugios. Cuando se encontraban terminados, aparentaban pequeños montículos encima de donde sobresalió cada pez, a través de un orificio horizontal de aproximadamente 3cm de diámetro, el cual comunica por medio de un túnel, sea vertical o diagonal a una cámara pequeña y de arreglo sencillo. Habitualmente, los peces permanecieron posados con sus aletas pectorales y pélvicas en los límites de la entrada, con el cuerpo dentro del refugio y observaban lo ocurrido en su entorno. Durante las interacciones de cortejo y desove, la llegada de la hembra provocó daños en la estructura del refugio del macho, que se encargó de reparar en los días subsecuentes, e incluso ahuyentó a las mismas hembras al pretender tomar material de construcción de sus inmediaciones.
Periodo de actividad común y durante la incubación: Los peces manifestaron hábitos diurnos exclusivamente y transcurrieron la mayor parte del tiempo en actividades como asomar la cabeza y observar a su alrededor como a la espera de alimento, además, si se presentaba cualquier perturbación se retraían al interior. Por la tarde, cada pez sujetó con la boca su propio fragmento de roca, el cual acomodaron para cubrir la entrada de sus refugios, desde una hora previa al oscurecer hasta una hora posterior al amanecer. Fue así que permanecieron activos en el periodo de 07-17hr, y los eventos de cortejo y desove se registraron en el transcurso de la mañana (8-11hr). De manera particular ocurrió que cuando el macho cuidó alguna masa ovígera, rehusó a alimentarse y restringió su periodo de actividad a tan solo 0.5-3hr del día.
Fidelidad al refugio: Los peces y en especial el macho manifestaron una prolongada permanencia en sus refugios respectivos (8-26 meses). Únicamente los intercambiaron, al dar inicio los primeros desplantes de cortejo, y en dos ocasiones dada la insistencia de una hembra por introducirse al refugio para tratar de desovar, lo que ocasionó la salida del macho. No obstante, un par de días después se reubicaron en sus refugios respectivos.
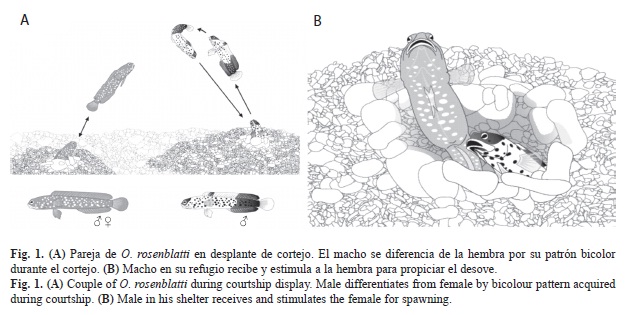
Patrón de desplazamiento durante el cortejo: Las hembras y el macho del bocón manchas azules limitaron sus movimientos a una distancia máxima de 20cm en las inmediaciones de su refugio, en esencia para capturar alimento y acarrear fragmentos de sustrato. Durante los eventos de cortejo y desove, realizaron incursiones a mayor distancia, tanto en sentido horizontal como vertical (Fig. 1A). El macho se distingue de las hembras por la rapidez de sus desplazamientos y por sus patrones de nado, descritos más adelante, en la sección de desplazamiento y desplantes.
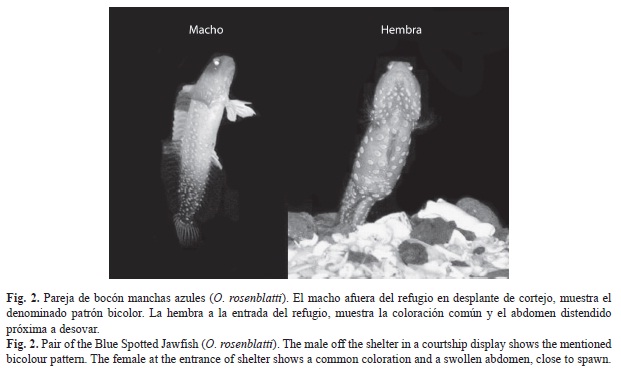
Patrón básico de coloración: Hembras y macho no se diferencian a simple vista, solo hasta que comenzaron a manifestar desplantes de cortejo. Antes de eso, presentaron coloración café-dorado en todo el cuerpo, con manchas azul rey de varios tamaños, desde atrás de los ojos hasta donde inicia la aleta caudal. Sin embargo, de una forma detallada se puede decir que el macho presenta el cuerpo ligeramente más brillante y el contorno anterior de la aleta dorsal de color ligeramente amarillo y las aletas pélvicas en negro desvanecido, comparado con el aspecto cristalino en todas las aletas de las hembras. Estas sutiles diferencias solo se aprecian en organismos vivos.
Coloración y desplantes del macho durante el cortejo: El macho manifestó cambios del patrón básico de coloración a partir del segundo evento de desove, con un patrón bicolor desvanecido, el cual adquirió durante el ascenso en la columna de agua al realizar un desplante de cortejo y perdió mientras permaneció en su refugio. A partir del cuarto evento de desove, el patrón bicolor fue bastante notorio y permaneció así por varios días hasta lograr el desove de alguna hembra y nuevamente perder dicho patrón bicolor durante el periodo de incubación. En cierto momento, mantuvo el patrón bicolor durante periodos prolongados (8-11 meses consecutivos), aún cuando no manifestaron desplantes de cortejo, ni desoves. En ocasiones el macho perdió el patrón bicolor, blanco y negro en apariencia, por lo cual los ascensos repetitivos en la columna de agua son considerados como criterios que evidencian la proximidad de un evento reproductivo, el cual se describe con mayor detalle a continuación:
1. Patrón bicolor en el macho: La región frontal de la cabeza y nuca mantuvieron la coloración café-dorada con manchas azul rey. El contorno inferior de la mandíbula se tornó de color negro. El tronco, detrás de los ojos hasta el ano, incluidas las aletas pélvicas y las espinas de la aleta anal, adquirieron un color blanco con tonalidad azul muy tenue, con las manchas azules desvanecidas, prácticamente imperceptibles por la rapidez del movimiento. La mitad anterior de la aleta dorsal fue ligeramente amarilla con un lunar de color amarillo-verde olivo entre las espinas segunda a sexta. La región caudal, que incluye la sección posterior de la aleta dorsal y la mayor parte de la aleta anal, predominó el color negro, con las manchas azules difíciles de apreciar por la falta de contraste y la rapidez de los desplazamientos. Las aletas pectorales y la caudal no presentaron coloración (Fig. 1A y 2).
2. Desplazamiento y desplantes: Durante los desplantes el macho se impulsó rápidamente con la región caudal y las pectorales; ascendió repentinamente y en el punto de máxima elevación (5-45cm), desplegó por completo sus aletas dorsal, anal y pélvicas; se mantuvo un instante (1-2s) con el aleteo de las pectorales, giro y descendió para introducirse de cabeza completamente al refugio. Los desplantes fueron rápidos y repentinos (duración total 2-4s). Después de un breve momento (2-10s), se asomó con cautela antes de realizar otro desplante.
Coloración y desplantes de las hembras en cortejo: Las dos hembras conservaron el patrón básico de coloración y los desplantes de cortejo consistieron en repeticiones de ascensos y descensos por encima de sus refugios (20-40cm), con el suave aleteo de las pectorales y ondulaciones de la región y aleta caudal. A diferencia de los machos, se mantuvieron durante más tiempo en el punto de mayor elevación (2-6s), con movimientos alternos de sus aletas pectorales. Invariablemente y sin girar, retrocedieron, pero solo se introdujeron parcialmente al refugio y permanecieron atentas, apoyadas en sus aletas pectorales o moviendo la región caudal dentro del refugio, hasta que volvieron a ascender. Sus desplantes fueron lentos y en apariencia aletargados (duración total 3-11s). En ningún momento desplegaron alguna de sus aletas, se movieron lento y por lo tanto realizaron menor cantidad de desplantes en comparación con el macho (Fig. 1A, Cuadro 1). Presentaron el abdomen distendido, como principal indicador de madurez gonádica así como evidencia de la proximidad del desove (Fig. 2).
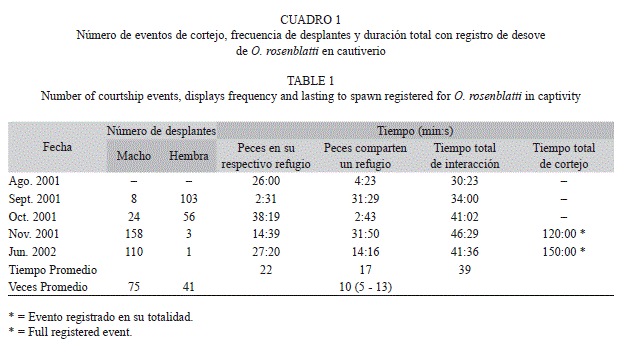
Interacciones durante el cortejo: El Cuadro 1 muestra que una pareja de peces, realizó diferentes repeticiones de ascensos de cortejo y los lapsos de tiempo que la hembra permaneció en su propio refugio o lo compartió con el macho. El macho realizó en promedio 75 ascensos (8-158) contra 41 de la hembra (1-103) y fue evidente como se intensifica la actividad que manifiesta la pareja conforme se aproxima el momento del desove. Ocasionalmente, la hembra se desplazó cautelosa y al ras del fondo hacia el refugio del macho y permaneció ahí durante un tiempo variable (2s-16min, n=49). Ante su presencia, el macho se retrajo, le permitió introducirse casi por completo, mientras la oprimió y empujó con la boca abierta sobre la región abdominal, cerca del ano (Fig. 1B). Esta manifestación dio la apariencia de tratar de sacarla, a lo que la hembra se resistió, situación interpretada como la estimulación directa del macho para lograr el desove. Las ocasiones en las que con certeza ocurrió el desove (n=5), compartieron el refugio en 10 oportunidades en promedio (5-13), durante lapsos que acumularon en promedio 39min (30min:23s-46min:29s) (Cuadro 1). De 50 eventos de desove registrados, únicamente dos fueron captados en su totalidad con 120 y 150min de duración. Fue imposible detectar alguna señal que motivara el acercamiento de las hembras al refugio del macho.
Desove: Los desoves ocurrieron en el intervalo de temperatura de 21.7-25.3°C y salinidad 36-41ups. La hembra pequeña (8cm aprox.) ubicada a mayor distancia del macho, (a 78cm contra 53cm de la hembra grande de 10cm aprox.) fue la primera en participar en actividades de cortejo y desovar de manera exitosa. La incorporación posterior, a las actividades reproductivas de la hembra relativamente más grande, provocó el intercambio de refugios entre ellas, con lo cual expresó su dominancia, territorialidad y agresividad reiterada, que en lo subsiguiente le permitió desovar con mayor frecuencia. En el momento preciso del desove, la hembra palidece ligeramente, mas no cambia su patrón básico. Inmediatamente sale del refugio del macho y se retira apresurada, al ras del fondo, con el abdomen hendido, señal inequívoca de haber liberado la masa ovígera. El macho por su parte, permanece oculto dentro del refugio, al parecer para fecundar los huevos y momentos después (40-80s) sale, se manifiesta agitado y realiza algunos desplantes (4-16), con menor vigor y escasa elevación (5cm). Un par de horas después, cesó toda actividad y el macho cubrió la entrada del refugio, mas temprano de lo habitual, y en los días subsecuentes, restringió sus periodos de actividad y de alimentación.
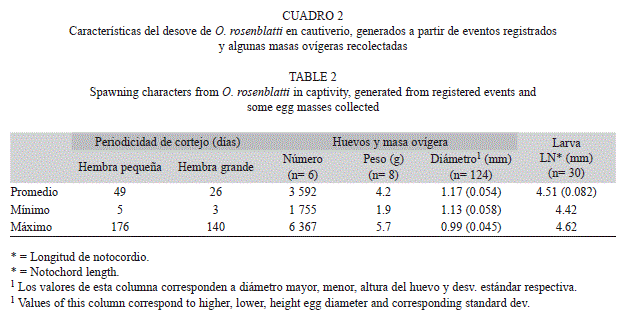
Periodicidad de desove: En total fueron captadas 50 situaciones relacionadas con eventos de cortejo del bocón manchas azules, la mayoría de manera parcial. En 33 de ellos se observó la masa ovígera o la presencia de larvas recién nacidas en días posteriores, lo que permitió asociarlas con el día exacto y el evento correspondiente de desove. A lo largo de 14 meses, la periodicidad promedio de desove fue de 49 y 26 días, para la hembra pequeña y grande, respectivamente (Cuadro 2). Las hembras llevaron a cabo el cortejo con evidencia de desove, tanto de manera simultánea como secuencial. De manera simultánea fueron registrados en siete ocasiones, pero solo se concretó el desove en tres de ellos, con el predominio de aquella hembra con mayor grado de distensión abdominal, insistencia en acudir con el macho, incluso logró desplazar a la otra con el cuerpo y mostrar agresividad, al grado de relegarla a su refugio. Esta última, expulsó una masa ovígera sin desarrollo embrionario, al detectar su presencia sobre el sustrato en la tarde del mismo día. Por otra parte, los eventos de cortejo secuencial, con escasos días de separación, fueron registrados en 11 ocasiones (desfase de 1-8 días). Cuando esto ocurrió, el macho logró incubar dos masas ovígeras a la vez, en cuatro ocasiones, que fueron constatadas a posteriori, por la presencia de larvas recién nacidas, con tan solo cuatro días de diferencia. Cuando ocurrió un desove secuencial con desfase menor a tres días, la hembra en turno se encargó de extraer la masa ovígera anterior, para desovar la propia, sin que el macho se lo impidiera. Al cabo de cierto tiempo, las hembras manifestaron canibalismo al abalanzarse sobre las masas ovígeras, en las escasas ocasiones que el macho entró, las tomó con la boca y las mostró a la entrada del refugio o enseguida de que fueron expulsadas fuera del mismo.
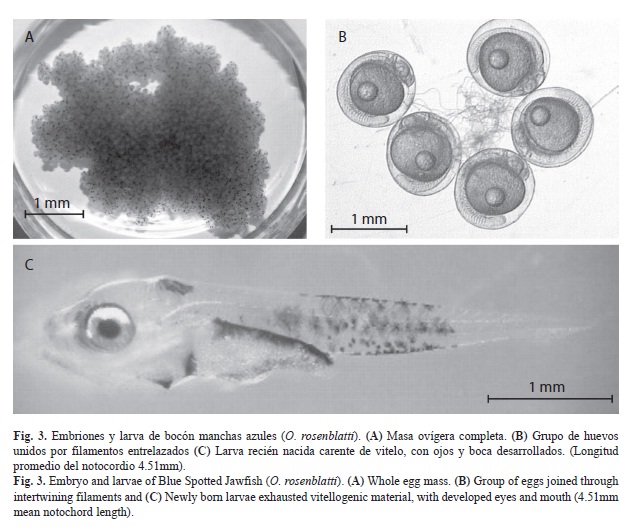
Incubación y fecundidad: Las masas ovígeras recién liberadas son de color blanquecino y conforme se desarrollan los embriones adquieren una tonalidad café-cobriza, debido a la pigmentación de los ojos. Presentan flotabilidad negativa y carecen de sustancias adhesivas que las fijan al sustrato, pero presentan filamentos del corion que los mantienen adheridos entre sí. Aun cuando realizó actividades de cortejo con una frecuencia de cada 14 días en promedio (1-119 días), las labores de incubación se presentaron cada 20 días en promedio (5-119 días), pero invariablemente el macho realizó la incubación de las masas ovígeras dentro del refugio, mas no en su cavidad bucal. Solamente, el macho manifestó cambios radicales de comportamiento y actividad, ya que durante la incubación descubrió la entrada del refugio por periodos breves (0.5-3hr), a diario o incluso en días alternos; perdió el patrón bicolor pero, en caso extremo, lo conservó hasta por 11 meses; al salir realizó desplantes de escasa elevación (3-6 desplantes, a 5cm de elevación), rechazó todo tipo de alimento y, finalmente, volvió a cubrir la entrada del refugio. El periodo de incubación promedio fue de 10.4 días (9-14 días), con variaciones debidas a la temperatura del agua (21.0-25.3°C). El cuadro 2 muestra información obtenida a partir de masas ovígeras recolectadas fuera del refugio, que el macho expulsó por razones desconocidas o por parte de alguna de las hembras para desovar una masa ovígera propia. Estas presentaron desarrollo embrionario sincrónico, con valor promedio de fecundidad de 3 592 huevos (n=6) y peso húmedo de 4.2g (n=8). Los huevos eran esféricos en apariencia, ya que presentaron tres medidas de diámetro con diferencias significativas (1.17mm promedio diámetro mayor, DE=0.054; 1.13mm promedio diámetro menor, 0.058; 0.99mm promedio de altura, 0.045; n=125) (Kruskall-Wallis, p<0.05). Al inicio carecen de pigmentos, presentan un glóbulo de aceite con 0.30mm de diámetro promedio (DE=0.021, n=59); la superficie del corion es lisa y en un sitio presentan generalmente cuatro (ocasionalmente seis) filamentos rizados, en torno al micrópilo (Nelson, 2006), los cuales se entrelazan con las de otros huevos para conformar la masa ovígera (Fig. 3A y B).
Eclosión: La eclosión de los huevos se da dentro de las primeras dos horas de oscuridad total y se liberan larvas de 4.51mm de longitud promedio de notocordio (DE=0.082, n=30) (Fig. 3C), que lograron salir a través de las ranuras de la entrada del refugio y se acumularon en la superficie del agua. Están pigmentadas profusamente, carecen de vitelo, presentan los ojos y la boca desarrollados, listas para comer. Al considerar el número de masas ovígeras liberadas por cada una de las hembras se estimó un éxito de incubación promedio del 70% (n=3). La recolecta y revisión, tanto de remanentes como de masas ovígeras expulsadas del refugio y colocadas en incubación de manera artificial en un acuario aparte con agua de mar esterilizada con UV a las misma temperatura de los adultos, permitió determinar que éstas presentaron elevados porcentajes de viabilidad a la eclosión, con valores superiores al 99% (n=4). A la mañana siguiente el macho expulsa el remanente de la masa ovígera, de aspecto algodonoso, debido a los restos de corion y sus filamentos, con escasos huevos y embriones en proceso de descomposición.
Discusión
Las características generales del cortejo del bocón manchas azules O. rosenblatti y su limitada capacidad de desplazamiento, son características que comparte con especies demersales que habitan en refugios, y que además manifiestan cuidado paterno, como sucede con representantes de otras familias como por ejemplo Chaenopsidae, Blennidae, Pomacentridae, Tripterygidae, Grammistidae (Thomson et al. 1979, Potts 1984, Thresher 1984, DeMartini 1999). Dichas cualidades, aunadas a su agresividad moderada, los hace atractivos para la industria del acuarismo marino, lo que ha propiciado investigaciones sobre la biología reproductiva de especies pertenecientes a la familia Opistognathidae, dado el potencial económico que representaría implementar su cultivo (Walch 1993, Walch 1994, Brons 1995).
La capacidad de O. rosenblatti para modificar el arreglo del sustrato, le permitió construir refugios estables para su protección, aún cuando el aspecto externo del refugio fue diferente del reportado en su hábitat natural por Allen & Robertson (1998), situación que también ha de haber ocurrido respecto del interior, al considerar lo reportado por Colin (1973) para O. aurifrons. Ello no impidió que manifestaran secuencias de comportamiento reproductivo, semejantes a las descritas en su hábitat natural (Thresher 1984, Walch 1994, Brons 1995, Allen & Robertson 1998). El presente estudio representa el primer registro de desove en cautiverio.
Los periodos prolongados de fidelidad al refugio, previos y subsecuentes al registro de los desoves (8-26 meses, respectivamente), permiten suponer que los peces eran jóvenes al momento de su captura (6cm) y alcanzaron la etapa adulta en cautiverio. Este hecho esta reforzado por el periodo de ocho meses, en que dos ejemplares convivieron en refugios contiguos y que posteriormente propició su separación al manifestar territorialidad y cortejo, además de la adquisición paulatina del patrón bicolor por parte del macho. La secuencia de manifestaciones de cortejo concuerda con los informes para la especie en su hábitat natural en cuanto al cambio a un patrón bicolor, el despliegue de aletas y suspensión en la columna de agua (3-5s), en secuencias repetitivas durante horas hasta lograr el desove (Thresher 1984). En nuestro caso, los primeros eventos de cortejo se prolongaron durante la mañana y parte de la tarde durante varios días, pero luego se restringieron considerablemente (2-2.5hr), lo cual es atribuible tanto a la transición de alcanzar la madurez reproductiva, así como a un proceso de aprendizaje, dicho esto último por el hecho extremo del macho en realizar un solo desplante y sin más preámbulo, propiciar que la hembra acudiera a desovar. Tal situación pudo deberse, a la ausencia de otros machos; no obstante, pone de manifiesto la flexibilidad y adaptabilidad del comportamiento de los individuos de Opistognathus a condiciones ambientales particulares (Potts, 1984).
Allen & Robertson (1998) reportan distancias breves entre refugios (<1m), como factor inhibidor de la reproducción en su hábitat natural, situación que contrasta con el éxito de desove registrado en este trabajo (46 y 63cm de distancia); sin embargo, fue evidente que la hembra más cercana al macho, recibió agresiones de manera insistente de parte del macho, pero sobre todo de la hembra, al grado de frustrar en varias ocasiones su participación en el desove. Los ascensos durante el cortejo propician el reconocimiento entre los organismos, pero se desconocen los mecanismos que favorecen la elección de pareja e incluso la señal precisa para acudir al refugio del macho a recibir la estimulación final que desencadena el desove, sin que la altura del acuario fuese limitante para la reproducción, al estar dentro del ámbito de ascensos reportado para O. rosenblatti (0.3-1.0m) (Thresher 1984).
El estupor o aletargamiento que manifiestan las hembras durante el proceso de cortejo, contrasta con la rapidez en los desplantes del macho, el cual se muestra inquieto y cauteloso, condición que ubica a las hembras en situación de mayor vulnerabilidad ante posibles depredadores (Johannes 1978), situación que no se da en cautiverio. Los opistognátidos son considerados incubadores bucales, en los que solo interviene el macho (Thresher 1984, Nelson 1996, Schultz III 2002); sin embargo, la peculiaridad observada en el macho de O. rosenblatti al optar por incubar las masas ovígeras dentro del refugio, pudo deberse a la desproporción de éstas, con respecto a su boca, aunque no se descarta la posibilidad que ocurra algo similar en organismos silvestres. De acuerdo con Hess (1993), solventar mediante este mecanismo las limitaciones que conlleva el cuidado paterno, le permite a O. rosenblatti, cortejar de manera secuencial ya sea con la misma u otra hembra y realizar la incubación simultánea, situaciones que representan el primer informe de poligamia, con capacidad de poliginia en esta especie, patrón que aún debe ser corroborado en la naturaleza, como está documentado para O. macrognathus y O. whitehursti (Hess 1993). Dicha particularidad, contribuye a incrementar el éxito reproductivo de las especies (Hess 1993) y podrían representar una ventaja para su crianza en cautiverio. La incubación exitosa de una segunda masa ovígera por parte de O. rosenblatti, fue confirmada por la aparición de larvas, desde cuatro hasta ocho días posteriores al desove mas reciente, al corroborar estos eventos con los registros de eclosión mas breves, que son de nueve días en condiciones estables de temperatura (25.3°C). Dicho periodo concuerda con tres de las cuatro especies de opistognátidos del Caribe (Hess 1993), no obstante que tal autor no proporciona valores de temperatura del agua. En el caso de O. rosenblatti, el éxito de incubación de masas ovígeras (70%) fue menor al reportado para otras especies (97%) (Hess 1993), sin dejar de lado que se contrastan eventos ocurridos en cautiverio contra otros en su hábitat natural.
La escasa información al respecto de la fecundidad de opistognátidos, con características similares en cuanto a talla, solo permite contrastar el promedio de 3 592 huevos obtenidos por desove de O. rosenblatti, contra los 500-1 000 huevos reportados en cautiverio para O. aurifrons (Walch 1994, Schultz III 2002). Hess (1993) en su trabajo con cuatro especies de opistognátidos, menciona la capacidad que tienen para engullir la masa ovígera y cerrar la boca, como medida de protección, es así que estima el volumen promedio de los huevos entre 0.57-0.81mm3, el cual depende de la especie, que para el caso de O. maxillosus es de 0.81mm3 y 1.15mm de diámetro (valor calculado por despeje de la incógnita en la formula de la esfera) de tal manera, se muestra muy semejante a O. rosenblatti (0.83mm3 y 1.17mm de diámetro). Al llevar a cabo la extrapolación al volumen de las masas ovígeras, ambas especies serían similares, con valores de 3 009.4 y 3 800.0mm3, respectivamente, con diferencias atribuibles al proceso de evaluación empleado por Hess (1993), consistente en rellenar con pequeñas esferas la cavidad bucal de ejemplares preservados.
Los ascensos que realiza el macho posteriores al desove y en días subsecuentes durante la incubación, podrían implicar la remoción de los desechos y garantizar la calidad del agua dentro del refugio, que junto con las propiedades asépticas del mucus segregado por los progenitores de especies que manifiestan cuidado paterno (Rasotto 1995, DeMartini 1999), les permite llevar la incubación a término de manera exitosa (éxito de incubación 70% y viabilidad a la eclosión superior al 99%), situación por demás interesante dada la prolongada permanencia del macho al interior del refugio. La sincronía del desarrollo embrionario se pone de manifiesto al ocurrir la eclosión en oscuridad, como una estrategia para disminuir la depredación y favorecer la dispersión de las larvas (Johannes 1978, Kohda 1988, Asoh & Yoshikawa 1996, Kohda 2002).
Al considerar la temperatura, el fotoperiodo y el alimento como los principales disparadores de la actividad reproductiva (Lam 1983), el hecho de mantener estables tales parámetros, aunado al ayuno prolongado durante los eventos de incubación, contribuyeron al desgaste fisiológico del macho (Hess 1993, Bromage 1995, Coleman 1999), que culminaron con su muerte.
Es factible suponer que las condiciones brindadas en cautiverio pudieron haber generado patrones de conducta diferentes e incluso anómalos, los cuales es necesario corroborar en la naturaleza, no obstante, al considerar las normas oficiales que regulan la captura de esta especie de ornato (NOM 059) y la capacidad de reproducirse en cautiverio, harían propicio enfocar esfuerzos tendientes a implementar su cultivo, como se ha hecho con otras especies (O. aurifrons y O. gilberti) (Tucker 1998) y contribuir de esta manera a la preservación de sus poblaciones naturales, principalmente por tratarse de una especie protegida y endémica del Golfo de California, de la cual no se tiene un seguimiento preciso de los registros de captura (Piña 2005), pero que se exhibe y oferta en medios electrónicos (Anónimo 2007, Anónimo 2008) y de acuerdo con Schultz III (2002), O. rosenblatti, junto con la especie del Atlántico O. aurifrons, son los opistognátidos más comúnmente solicitados por los aficionados al acuarismo mundial.
Agradecimientos
Al Instituto Politécnico Nacional (CGPI) por el apoyo financiero al proyecto: “Efecto del alimento vivo en el contenido de aminoácidos y ácidos grasos esenciales (polares y neutros) en larvas de Trambollito Vela (Emblemaria hypacanthus)” (clave 20061291). Armando Hernández por el apoyo brindado en la edición de las figuras. A los revisores, cuyas opiniones permitieron mejorar el manuscrito.
Referencias
Allen, G.R. & D.R. Robertson. 1998. Peces del Pacífico Oriental Tropical. Comisión Nacional para el Conocimiento y Uso de la Biodiversidad y Agrupación Sierra Madre, México, D.F., México. [ Links ]
Anónimo. 2004. Informe sobre la situación del Medio Ambiente en México. SEMARNAT, México, D.F., México. (Consultado: 27 enero 2011, http://app1. semarnat.gob.mx/dgeia/informe_04/04_biodiversidad/index_biodiversidad.html). [ Links ]
Anónimo. 2007. That Pet Place. Fish Net. Lancaster, Pensilvania, EEUU. (Consultado: 21 enero 2011, http:// www.thatpetplace.com). [ Links ]
Anónimo. 2008. Reef Hot Spot. Inglewood, California, EEUU. (Consultado: 21 enero 2011, http://www. reefhotspot.com). [ Links ]
Anónimo. 2010. FishBase. (Consultado: 1 agosto 2011, http://www.fishbase.org/Summary.php?ID=46578 & g enusname=Opistognathus & speciesname=rosenblatti & AT=Opistognathus+rosenblatti & lang=Spanish). [ Links ]
Asoh, K. & T. Yoshikawa. 1996. Nesting behaviour, male parental care, and embryonic development in the fairy basslet Gramma loreto. Copeia 1: 1-8. [ Links ]
Brons, R. 1995. Reproduction and Captive Breeding of Two Red Sea Dottybacks: Pseudochromis fridmani and P. flavivertex. Breeder’s Registry 3: 1-9. [ Links ]
Bromage, N. 1995. Broodstock Management and Seed Quality. General Considerations, p. 1-24. In N.R. Bromage & R.J. Roberts (eds.). Broodstock Management and Egg and Larval Quality. Blackwell, Oxford, Gran Bretaña. [ Links ]
Colin, P.L. 1973. Burrowing Behaviour of the Yellowhead Jawfish, Opistognathus aurifrons. Copeia 1: 84-90. [ Links ]
Coleman, R.M. 1999. Parental care in intertidal fishes, p 165-180. In M.H. Horn, K.L.M. Martin & M.A. Chotkowski (eds.). Intertidal Fishes. Life in Two Worlds. Academic, Londres, Inglaterra. [ Links ]
D.O.F. (Diario Oficial de la Federación). 2002. Norma Oficial Mexicana. NOM-059-ECOL-2001. México 96 p. [ Links ]
DeMartini, E.E. 1999. Intertidal Spawning, p. 143-164. In M.H. Horn, K.L.M. Martin & M.A. Chotkowski (eds.). Intertidal Fishes. Life in Two Worlds. Academic, San Diego, California, EEUU. [ Links ]
Flores, O. & P. Gerez. 1992. Biodiversidad y conservación en México: Vertebrados, vegetación y uso de suelo. CONABIO. UNAM, Distrito Federal, México. [ Links ]
Hess, H.C. 1993. Male mouth brooding in jawfishes (Opistognathidae): constraits on polygyny. Bull. Mar. Sci. 52: 806-818. [ Links ]
Johannes, R.E. 1978. Reproductive strategies of coastal marine fishes in the tropics. Env. Biol. Fish. 3: 65-84. [ Links ]
Kohda, M. 1988. Diurnal periodicity of spawning activity of permanent territorial damselfishes (Teleostei: Pomacentridae). Env. Biol. Fish. 21: 91-100. [ Links ]
Kohda, M. 2002. The role of temperature and embryo development time in the diel timing of spawning in a coral-reef damselfish with high-frequency spawning synchrony. Env. Biol. Fish. 64: 379-392. [ Links ]
Lam, T.J. 1983. Environmental influences on gonadal activity in fish, p. 65-116. In W.S. Hoar, D.J. Randall, E.M. Donaldson (eds.). Fish Physiology, IX B. Academic, Londres, Inglaterra. [ Links ]
Nelson, 1996. Fishes of the World. Wiley, Nueva York, EEUU. [ Links ]
Piña, E.R. 2005. La pesquería de especies marinas con fines de ornato en México. El parque marino de Loreto. Tesis de Maestría, Centro Interdisciplinario de Ciencias Marinas (IPN), La Paz, México. [ Links ]
Potts, G.W. 1984. Parental behaviour in temperate marine teleosts with special reference to the development of nest structures, p. 223-242. In G.W. Potts & R.J. Wooten (eds.). Fish Reproduction. Strategies and Tactics. Academic, Londres, Inglaterra. [ Links ]
Rasotto, M.B. 1995. Male reproductive apparatus of some Blennioidei (Pisces: Teleostei). Copeia 907-914. [ Links ]
Shultz, H.C. III. 2002. Lets Jaw About Jawfish. (Consultado: 1 agosto 2011, http://reefkeeping.com/issues/2002-03/hcs3/index.php). [ Links ]
Thresher, R.E. 1984. Reproduction in reef fishes. T.F.H. Neptune, Nueva Jersey, EEUU. [ Links ]
Thomson, D.A., L.T. Findley & A.N. Kerstitch. 1979. Reef Fishes of the Sea of Cortez. The Rocky-Shore Fishes of the Gulf of California. Wiley, Nueva York, EEUU. [ Links ]
Tucker, J.W. 1998. Marine Fish Culture. Kluwer, Norwell, Massachusetts, EEUU. [ Links ]
Walch, J.C. 1993. Breeding Jawfish. Breeder’s Registry 1: 1-4. [ Links ]
Walch, J.C. 1994. Reproduction of yellowhead jawfish in captivity. Sea Scope Aqua. Syst. 11: 1-2. Mentor, Ohio, EEUU. [ Links ]
*Correspondencia:
Mauricio Contreras: Laboratorio de Biología Experimental, Centro Interdisciplinario de Ciencias Marinas, Instituto Politécnico Nacional. Av. IPN s/n Col. Playa Palo de Santa Rita, La Paz, Baja California Sur, México. C.P. 23096 Apdo. Postal 592. contrero@hotmail.com
Benjamín Anguas: Laboratorio de Biología Experimental, Centro Interdisciplinario de Ciencias Marinas, Instituto Politécnico Nacional. Av. IPN s/n Col. Playa Palo de Santa Rita, La Paz, Baja California Sur, México. C.P. 23096 Apdo. Postal 592. banguas@ipn.mx.
Pedro G. González: Laboratorio de Biología Experimental, Centro Interdisciplinario de Ciencias Marinas, Instituto Politécnico Nacional. Av. IPN s/n Col. Playa Palo de Santa Rita, La Paz, Baja California Sur, México. C.P. 23096 Apdo. Postal 592. pgonzaljaiba@hotmail.com
Rodolfo E. Martínez: Laboratorio de Biología Experimental, Centro Interdisciplinario de Ciencias Marinas, Instituto Politécnico Nacional. Av. IPN s/n Col. Playa Palo de Santa Rita, La Paz, Baja California Sur, México. C.P. 23096 Apdo. Postal 592. rpecero@yahoo.com.
1. Laboratorio de Biología Experimental, Centro Interdisciplinario de Ciencias Marinas, Instituto Politécnico Nacional. Av. IPN s/n Col. Playa Palo de Santa Rita, La Paz, Baja California Sur, México. C.P. 23096 Apdo. Postal 592; contrero@hotmail.com, banguas@ipn.mx, pgonzaljaiba@hotmail.com, rpecero@yahoo.com.
Recibido 14-IV-2011. Corregido 09-IX-2011. Aceptado 07-X-2011.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}