










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.60 n.3 San José Sep. 2012
Morfometría geométrica alar para la identificación de Chrysomya albiceps y C. megacephala (Diptera: Calliphoridae) de Venezuela
*Dirección para correspondencia
Abstract
Geometric wing morphometrics for Chrysomya albiceps and C. megacephala identification (Diptera: Calliphoridae) from Venezuela. Calliphoridae is one of the families with the greatest number of species with forensic importance, which immature stages feed and develop on decaying material. in Venezuela, there are few studies on this taxon reporting the dominance of Chrysomya over other carrion flies. The goal of this study was to analyze the variations on wing morphometrics, to support the identification of two forensic flies. For this we photographed a total of 168 wings from C. albiceps (n=111) and C. megacephala (n=57). Landmark coordinate (x, y) configurations were registered and aligned by Generalized Procrustes Analysis. Principal Component Analysis and shape significance test based on Procrustes distance were implemented. Statistical analysis of variance found significant differences in wing isometric size (Kruskal-Wallis). The PCA showed the separation between species, and shape test showed significant differences (F Goodall´s). The main differences between both species were localized on subcosta rupture, join of R2+3 with wing border, join of dm-cu, and join of Cu with bm-cu, suggesting that wing morphometrics is a suitable tool in the discrimination of blowfly species.
Key words: landmarks, procrustes, forensic entomology, Chrysomyinae.
Resumen
Calliphoridae es una de las familias con el mayor número de especies de importancia forense, donde sus fases inmaduras se alimentan y desarrollan sobre material en descomposición. En Venezuela, son pocos los estudios en este taxon, sin embargo se ha reportado la dominancia del género Chrysomya sobre los otros dípteros. El objetivo fue analizar la variación morfométrica en la arquitectura alar, como herramienta de apoyo a la identificación de dípteros de importancia forense. Se fotografiaron 168 alas de C. albiceps (n=111) y C. megacephala (n=57) y se registraron configuraciones de coordenadas (x, y), se alinearon mediante Análisis Generalizado de Procrustes. Se efectuaron Análisis de Componentes Principales y comparaciones pareadas entre distancia parcial de Procrustes. El análisis estadístico de varianza encontró diferencias en el tamaño isométrico del ala (Kruskal-Wallis). El ACP mostró la separación de ambas especies, y la prueba de configuraciones determinó diferencias significativas (F Goodall). Las principales diferencias entre ambas especies se encontraron en: ruptura de la subcosta, unión de R2+3 con el borde del ala, unión dm-cu, y unión de Cu con dm-cu, lo que confirma que la morfometría alar es una herramienta idónea en la discriminación de especies de Calliphoridae.
Palabras clave: puntos anatómicos de referencia, procrustes, entomología forense, Chrysomyinae.
Los insectos necrófagos, entre ellos Diptera y Coleoptera, se alimentan de tejido en putrefacción y forman sucesiones ecológicas asociadas con cuerpos en descomposición. Estos grupos constituyen la categoría más importante para el establecimiento del Intervalo Post Mortem (IPM), tiempo transcurrido desde el momento del deceso hasta el instante que se encuentra el cadáver (Cova 1974, Smith 1986, Queiroz et al. 1997, Magaña 2001, Liria 2006). Los insectos son atraídos al cadáver por
los gases desprendidos durante el proceso de degradación de glúcidos, lípidos, entre otros, generando amoníaco (NH3) ácido sulfúrico (SH2), nitrógeno (N2) y anhídrido carbónico (CO2) (Byrd & Castner 2001). Con ello, los diferentes estados de descomposición de un cadáver atraen selectivamente a grupos de insectos. Generalmente, las moscas son las primeras en colonizar manteniendo una sucesión, que varía solo en la composición de especies (Byrd & Castner 2001, Calderón-Arguedas et al. 2005). Basados en este principio, la entomología forense es la disciplina que estudia a insectos y otros artrópodos asociados a cadáveres, como herramienta de la medicina legal para datar y estimar las causas y lugar de una muerte (Smith 1986, Maldonado 2002). De esta manera, el reconocimiento de las especies vinculadas a la escena del crimen, proveen pruebas criminalísticas como: ubicación geográfica del suceso e IPM; las cuales pueden establecer la autoría, método utilizado, lugar y fecha de muerte (Smith 1986).
La estimación del IPM a partir de datos entomológicos, se realiza mediante: 1) estudios de sucesión (patrones de colonización y extinción de especies, no temporales, direccionales y continuos presentes en el cadáver), 2) comparando el desarrollo con tablas de crecimiento de la especie encontrada a una temperatura y humedad relativa específicas (Smith 1986, Von Zuben & Godoy 2001, Maldonado 2002). Es decir, el asiento para la determinación del IPM, es la identificación de especies, de los insectos encontrados en la escena del crimen.
Calliphoridae, es una de las familias con el mayor número de especies de importancia forense (Queiroz et al. 1997). En Venezuela, se ha reportado la dominancia del género Chrysomya sobre los otros dípteros. Particularmente especies como C. albiceps (Wiedemann 1819) y C. megacephala (Fabricius 1794), las cuales colonizan durante los primeros días de descomposición (Mavárez-Cardozo et al.2005, Liria 2006, Magaña et al. 2006). En esta familia, existen escenarios donde se presenta dificultad para la identificación, restringiendo la determinación de la especie a solo uno o muy pocos caracteres morfológicos, que en ocasiones pueden ser ambiguos (Queiroz et al. 1997). Por otro lado, la identificación puede ser crucial en el análisis, porque especies estrechamente relacionadas crecen en proporciones de tiempo diferentes: a una temperatura de 19ºC, el crecimiento de una especie puede tener un promedio de 47 días, mientras que otra especie del mismo género, a la misma temperatura puede tener un promedio de 84 días (Wells & Williams 2005). Por consiguiente, la irresolución taxonómica puede llevar a una incertidumbre en la estimación del IPM.
Adicionalmente, aún cuando la entomología forense es una ciencia utilizada en Estados Unidos, España y en países de América Latina como Brasil, Argentina y recientemente Colombia, en Venezuela el estudio de la entomología forense no ha recibido una atención sistemática por los profesionales del área entomológica o por expertos forenses. Como consecuencia, el científico criminalista se enfrenta a la escena de un crimen con poca formación en morfología de insectos y taxonomía de grupos, por lo que, su aporte en la labor de establecer la data de la muerte por datos entomológicos se dificulta. Por lo anterior, se intenta explorar y optimizar una nueva herramienta que permita de una manera rápida, fácil y eficiente, la identificación de especies, y que proporcione pruebas criminológicas a la investigación en el tiempo requerido.
Actualmente, los métodos de identificación no sólo deben ser rápidos sino también de bajo costo. En este sentido, la morfometría geométrica, es una herramienta matemática con fundamentos biológicos (Bookstein 1991), que permite descomponer la variación resultante de la fisiología de los individuos de aquellas más estables propias de la población, producto del componente genético. Permite identificar individuos a partir de la conformación de estructuras específicas, en especies estrechamente relacionadas (Dujardin 2000), basado en métodos de descripción y análisis estadísticos de la variación de la forma en estructuras particulares en y entre individuos (Rohlf & Marcus 1993). Esta técnica se ha utilizado en diversas clases de organismos vegetales y animales (vertebrados e invertebrados), y particularmente en insectos de importancia médica, como triatóminos (Triatominae), flebótomos (Phlebotominae) y mosquitos (Culicidae). Ha sido empleada en estudios de estructura poblacional (De La Riva et al. 2001, Belen et al. 2004, Schachter-Broide et al. 2004, Dvorak et al. 2006, Soto-Vivas et al. 2007), identificación de especies (Matías et al. 2001, Villegas et al. 2002, Calle et al. 2008), para detectar diferencias entre parentales de campo y descendientes de laboratorio (Jaramillo & Wolff 2002), así como en estudios filogenéticos (Soto-Vivas et al. 2011).
Este trabajo pretende comprobar la aplicabilidad de la morfometría geométrica en el análisis del tamaño y configuración del ala en dos especies de Chrysomya, con el fin de proporcionar al campo de la criminalística una nueva herramienta que permita discriminar e identificar moscas de importancia forense.
Materiales y métodos
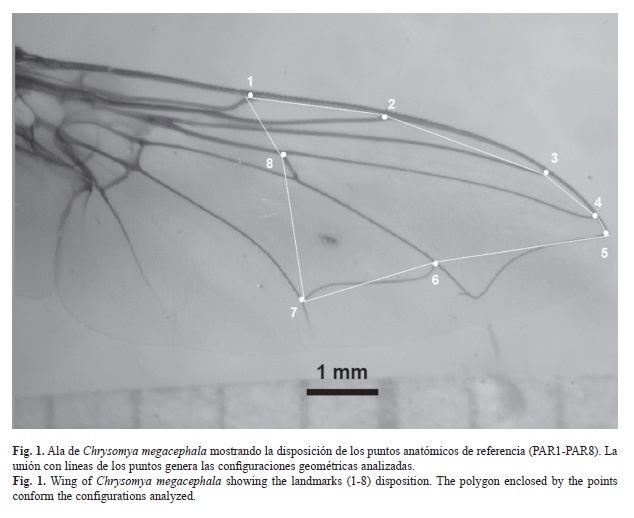
Adquisición de datos: Se realizaron muestreos sobre cadáveres de ratas SpragueDawley y cebo de bofe bovino (Corazón, hígado, páncreas), en el Campus Bárbula de la Universidad de Carabobo, estado Carabobo, Venezuela (10º18’15’’ N - 68º02’41’’ W). Las moscas fueron obtenidas durante el primer día de descomposición de las ratas y cebos. Los califóridos se identificaron siguiendo las claves taxonómicas de: Ribeiro & Carvalho 1998, Carvalho & Ribeiro 2000, Whitworth 2006 y Amat 2009. Un total de 168 alas: 111 de C. albiceps y 57 de C. megacephala, se disectaron y montaron en láminas, luego se fotografiaron, digitalizaron, y se ubicaron ocho puntos anatómicos de referencia en la nervadura de las venas del ala (PAR1-PAR8), los cuales corresponden con el tipo I sensu Bookstein (1991), siguiendo la nomenclatura de McAlpine (1987), ruptura de la subcosta (PAR1), unión de R1 con el borde del ala (PAR2), unión de R2+3 con el borde del ala (PAR3), unión de R4+5 con el borde del ala (PAR4), unión de M con el borde del ala (PAR5), unión dm-cu (PAR6), unión de Cu con dm-cu (PAR7) y unión de M con bm-cu (PAR8). La configuración de los ocho pares de coordenadas x y de cada ala (Fig. 1) se capturó mediante la ayuda del programa tpsDig (Rohlf 2006).
Análisis de morfometría geométrica: A partir de la matriz de 168 configuraciones de coordenadas geométricas de los ocho puntos (PAR1-PAR8), se realizó el Análisis Generalizado de Procrustes, con el programa CoordGen (Sheets 2005a) para una superimposición Procrustes y luego extraer una matriz con las variables de conformación (Partial warps=Pw) y tamaño centroide (CS). Esta matriz de Pw se usó para un Análisis de Componentes Principales (ACP) con el programa PCAGen (Sheets 2005b), mediante el cual se exploraron los principales ejes de variación morfológica en el espacio tangente, para observar las tendencias de variación entre y dentro de las muestras. Luego se evaluó la significancia entre las diferencias de configuraciones con el programa TwoGroup (Sheets 2005c) mediante la prueba de Goodall. El estadístico F está basado en el cociente de las distancias de Procrustes al cuadrado entre las medias de cada grupo, y la suma de las distancias de Procrustes al cuadrado entre cada espécimen y la media de su grupo. Sin embargo, esta prueba asume distribución de la variación alrededor de la media, por lo cual, se utilizó el método de Bootstrap para realizar 900 permutaciones a los datos y posterior cálculo del estadístico (Bookstein 1997, Zelditch et al. 2004). Las diferencias en los valores de CS entre las especies fueron analizadas en PAST (Hammer & Harper 2011) mediante la prueba de Kruskal-Wallis (p<0.05) con corrección de Bonferroni.
Resultados
Los análisis de morfometría geométrica se presentan a continuación.
Tamaño isométrico: Se encontraron diferencias significativas (Kruskal-Wallis: χ2=6.748, p<0.05) en el tamaño isométrico del ala, en donde C. albiceps presenta mayor talla (3.137mm±0.490) respecto a C. megacephala (2.971mm±0.409).
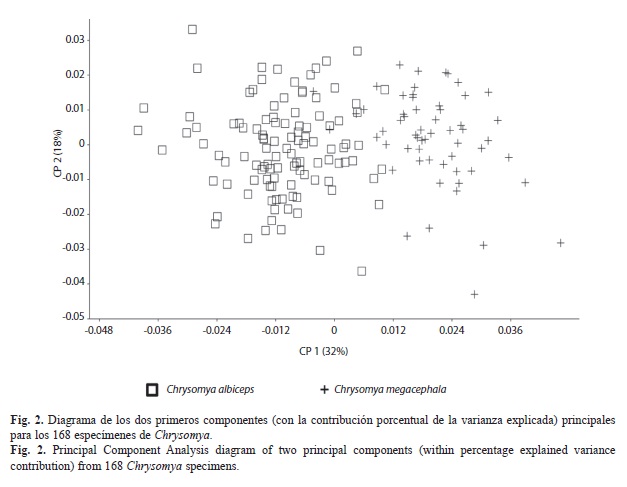
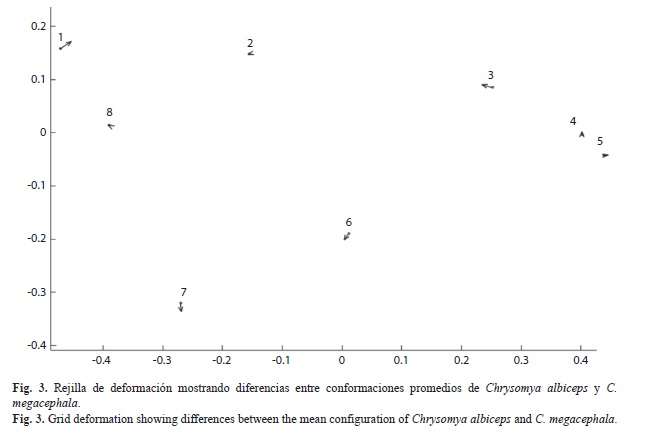
Diferencias de conformación alar: Se presentan los resultados del ACP y la comparación pareada entre las configuraciones de las especies. El diagrama de los dos primeros componentes principales (Fig. 2) muestra la separación entre C. albiceps y C. megacephala; estas diferencias fueron significativas (F=50.17, gl1=12, gl2=1992, p<0.001) y distancia parcial de Procrustes 0.0311. La función de interpolación de placas delgadas (thin plate spline=TPS), permitió inferir las diferencias en conformación como deformaciones de los ocho puntos en una rejilla, calculada entre las configuraciones promedios de ambas especies en el ACP (Fig. 3). Las diferencias más importantes se presentan en los PAR1, PAR3, PAR6 y PAR7. En C. albiceps en el PAR1 ocurre desplazamiento hacia arriba en el sentido diagonal a la derecha, el PAR3 se mueve en sentido diagonal hacia el PAR2, mientras que el PAR6 se acerca en sentido diagonal hacia el PAR7, y a su vez se desplaza verticalmente. Por otra parte en C. megacephala, el PAR1 se desplaza hacia abajo en sentido diagonal a la izquierda, el PAR3 se acerca diagonalmente a la derecha hacia el PAR4, el PAR6 se desplaza hacia arriba en sentido diagonal al PAR3, y el PAR7 presenta desplazamiento vertical.
Discusión
Tamaño isométrico: Los resultados de las variaciones de tamaño del ala entre C. albiceps y C. megacephala, estimada a través del tamaño isométrico, revelan diferencias significativas entre ambos táxones. Brown (1979), realizó uno de los primeros estudios morfométricos del ala en especies de Muscidae, Sarcophagidae y Calliphoridae. Aún cuando su investigación utilizó caracteres como distancia entre las venas r-m, m-cu, M, R2+3 y Sc, entre otras, la separación de los táxones se realizó mediante el tamaño del ala.
Por otro lado, Lomônaco & Germanos (2001) determinaron en Musca domestica (Linnaeus) el tamaño del ala y su relación con el tiempo de desarrollo larval y las densidades larvarias. Encontrando que tales variaciones son causadas por asimetrías fluctuantes (que denotan pequeñas desviaciones de la simetría perfecta de cualquier carácter de organismos con simetría bilateral), y que dichas fluctuaciones fueron inducidas por la competencia por el alimento. Asimismo, indican que los niveles de variaciones en la asimetría fluctuante pueden ser indicativos de stress o de la plasticidad fenotípica de la especie, que estarían promoviendo la sobrevivencia de un número mayor de individuos.
Estos resultados fueron confirmados por Reigada & Godoy (2005), que relacionaron el tamaño de alas y tibias en C. megacephala con su fecundidad estacional. Los autores determinan correlaciones negativas y significativas entre la temperatura mensual y los tamaños de ambas estructuras. De esta forma, el menor tamaño se presenta en los meses cálidos, cuando hay mayor abundancia de individuos (aumenta la competencia), por otro lado, durante los meses fríos se evidencian el mayor tamaño de las estructuras y a su vez la menor abundancia de individuos. Pero cabe destacar, que esto puede no cumplirse para todas las especies, puesto que, Gião & Godoy (2006) en estudios similares para Lucilia eximia (Wiedemann), encuentran que no existe correlación entre la temperatura y los tamaños de ala y tibia, y concluyen, que esto puede ser debido a la estabilidad en la abundancia de dicha especie durante el año.
En nuestra investigación, las diferencias del tamaño alar, donde C. albiceps presenta mayor talla respecto a C. megacephala, posiblemente se asocien a factores como la densidad larval, tipo de sustrato donde se criaron, variables ambientales como temperatura y humedad y competencia intraespecífica. Además, estudios sobre dinámica poblacional, dispersión y depredación en Calliphoridae, señalan la gran capacidad competitiva de C. albiceps, con respecto a especies como C. putoria (Wiedemann), C. megacephala y Cochliomya macellaria (Fabricius); así como, la depredación de C. albiceps sobre durante la fase de dispersión larvaria (Godoy et. al. 2001, Von Zuben & Godoy 2001, Bartholo de Andrade C. macellariaet al. 2002, Faria et al. 2004, Gomes et al. 2006). Debido a esto, las diferencias de talla encontradas entre C. albiceps y C. megacephala podrían asociarse con aspectos relacionados a colonización de sustrato y competencia entre ambas especies.
Diferencias de conformación alar y uso de la morfometría geométrica para la identificación de especies: La función de placas delgadas TPS permitió evidenciar las diferencias de conformación, en la configuración de ambas especies. Tradicionalmente, en la clasificación taxonómica de las especies de Calliphoridae y otras familias de calípterados, los caracteres utilizados son la quetotaxia del tórax, presencia de setas en la superficie posterodorsal de la sección basal de la vena radial y la inflexión de la vena medial (Shewell 1987, Ribeiro & Carvalho 1998, Carvalho & Ribeiro 2000, Whitworth 2006, Amat 2009). La visualización de estos caracteres requiere de conocimiento detallado de morfología de insectos y taxonomía del grupo. Por otro lado, algunas de estas setas podrían desprenderse durante la manipulación o recolecta de los especímenes, y en algunos casos la interpretación de estos caracteres puede ser ambigua. Debido a esto, la configuración de la venación alar podría considerarse como una herramienta adicional en la taxonomía de estos táxones. Concretamente los puntos anatómicos de referencia son: ruptura de la subcosta (PAR1), unión de R2+3 con el borde del ala (PAR3), unión dm-cu (PAR6), y unión de Cu con dm-cu (PAR7).
Los estudios de variación en la morfología del ala de dípteros han demostrado que factores como: clima, temperatura, fotoperiodo a lo largo de un gradiente latitudinal y competencia larval durante el desarrollo, influyen en el tamaño y conformación del ala (Aytekin et al. 2007, Bubliy et al. 2007, Jirakanjanakit et al. 2007, Loh et al. 2008). En otros casos, las diferencias en la conformación podrían ser consecuencia de interacciones más complejas, que contenienen así información que permitiría postular hipótesis filogenéticas (Soto-Vivas et al. 2011).
Por ello, los resultados de distintos estudios sugieren que la morfometría geométrica es particularmente apropiada para el análisis fenotípico de variación de la forma (Klingenberg & Zaklan 2000). Esta herramienta permite descomponer la variación resultante de la fisiología de los individuos (variación de tamaño) de aquellas más estables propias de la población, producto del componente genético (variación de la forma) (Dujardin 2000). La morfometría geométrica ha demostrado su utilidad para resolver problemas taxonómicos en insectos de importancia médica (Matias et al. 2001, Villegas et al. 2002, Calle et al. 2008), mediante la discriminación a partir de la configuración alar, de los integrantes de una especie en particular de otros que no lo son.
Calle et al. (2008) lograron distinguir las hembras de 11 especies del subgénero Nyssorhynchus de Anopheles presentes en Colombia mediante el uso de la información contenida en la conformación del ala. Estos resultados permiten proponer la utilización de los puntos anatómicos seleccionados como apoyo en la taxonomía de este grupo Recientemente, Lyra et al. (2010) sugieren que la morfometría alar representa un método sencillo y fiable para la identificación de C. hominivorax (Coquerel) y C. macellaria.
Los resultados descritos en este trabajo, demuestran que la morfometría geométrica del ala es una herramienta idónea en la discriminación de especies de Calliphoridae, siendo un método de fácil aplicación y con validación estadística. En este sentido, resulta interesante que utilizando ocho puntos anatómicos en el ala, a diferencia de los 16 empleados por Lyra (2010), se logren diferenciar las especies de Chrysomya. Sin embargo, aún es necesario realizar análisis adicionales que incluyan otras especies de los géneros Lucilia, Paralucilia, Cochliomya, entre otras, con el fin de evaluar la relevancia de los puntos anatómicos seleccionados. Por otro lado, esta investigación abre camino en estudios de especies similares como C. albiceps y C. rufifacies (Macquart), en las cuales, la diferenciación se realiza por la presencia de una seta proepimeral y 2-4 setas proespisternales en C. rufifacies y sin seta proepimeral y 4-6 setas proespisternales en C. albiceps (Amat 2009).
Agradecimientos
Deseamos expresar nuestro agradecimiento a todas las personas que facilitaron su apoyo en la realización de este trabajo. Darjaniva Molina de Fernández, Ana Soto-Vivas e Gianna Martiradona, del instituto de Altos Estudios en Salud Pública “Dr. Arnoldo Gabaldon” MppSalud. Al Departamento de Biología de la Universidad de Carabobo y FONACYT– MppCyT, Programa Misión Ciencias, por el apoyo y financiamiento de los estudios doctorales (MV). A los árbitros anónimos por los valiosos comentarios y sugerencias en mejora de la versión final del presente trabajo.
Referencias
Amat, E. 2009. Contribución al conocimiento de las Chrysomyinae y Toxotarsinae (Diptera: Calliphoridae) de Colombia. style="font-family: verdana;">Rev. Mex. Biodiv. 80: 693-708. [ Links ]
Aytekin, M., B. Alten, S.S. Caglar, Y. Ozbel, S. Kaynas, F.M. Simsek, O.E. Kasap & A. Belen. 2007. Phenotypic variation among local populations of phlebotomine sandflies (Diptera: Psychodidae) in southern Turkey. J. Vector. Ecol. 32: 226-234. [ Links ]
Bartholo de Andrade, J.F.A. Rocha, P. Rodrigues, G. Souza-Rosa, L. Faria, C.J. Von Zuben, M Rossi & W.A. Godoy. 2002. Larval dispersal and predation in experimental populations of Chrysomya albiceps and Chrysomya megacephala (Diptera: Calliphoridae). Mem. inst. Oswaldo Cruz 97:1137-1140. [ Links ]
Belen, A., B. Alten & A.M. Aytekin. 2004. Altitudinal variation in morphometric and molecular characteristics of Phlebotomus papatasi populations. Med. Vet. Entomol. 18: 343-350. [ Links ]
Bookstein, F.L. 1991. Morphometric tools for landmark data: Geometry and Biology. Cambridge, EEUU. [ Links ]
Bookstein, F.L. 1997. Landmark methods for forms without landmarks: localizing group differences in outline shape. Med. image Anal. 1: 225-243. [ Links ]
Brown, K.R. 1979. Comparative Wing Morphometrics of some Calyptrate Diptera. J. Aust. entomol. Soc. 18: 289-303. [ Links ]
Bubliy, O.A., K.S. Tcheslavskaia, A.M. Kulikov, O.E. Lazebny & V.G. Mitrofanov. 2007. Variation of wing shape in the Drosophila virilis species group (Diptera: Drosophilidae). J. Zool. Systemat. Evol. Res. 46: 38-47. [ Links ]
Byrd, J.H. & J.L. Castner. 2001. Forensic Entomology: The Utility of Arthropods in Legal investigations. CRC, EEUU. [ Links ]
Calderón-Arguedas, O., A. Troyo & M.E. Solano. 2005. Sucesión de larvas de muscoideos durante la degradación cadavérica en un bosque premontano húmedo tropical. Biomedica 16: 79-85. [ Links ]
Calle, D.A., M. Quiñónez, H. Erazo & N. Jaramillo. 2008. Discriminación por morfometría geométrica de once especies de Anopheles (Nyssorhynchus) presentes en Colombia. Biomedica 28: 371-385. [ Links ]
Carvalho, C.J.B. & P.B. Ribeiro. 2000. Chave de identificação das especies de Calliphoridae (Diptera) do Sul do Brasil. Rev. Bras. Parasitol. Vet. 9: 169-173. [ Links ]
Cova, G.P. 1974. Princípios Generales de Entomología. Fundación Venezolana para la salud y la educación, Caracas, Venezuela. [ Links ]
De La Riva, J., F. Le Pont, V. Ali, A. Matias, S. Mollinedo & J.P. Dujardin. 2001. Wing Geometry as a tool for studying the Lutzomyia longipalpis (Diptera: Psychodidae). Mem. inst. Oswaldo Cruz 96: 1089-1094. [ Links ]
Dujardin, J.P. 2000. introducción a la Morfometría, con énfasis en Triatominae y Phlebotominae. iRD, UMR CNRS-iRD 9226. (Disponible en: http://www.mpl.ird.fr/morphometrics/). [ Links ]
Dvorak, V., A.M. Aytekin, B. Alten, S. Skarupova, J. Votypka & P. Volf. 2006. A comparison of the intraspecific variability of Phlebotomus sergenti Parrot, 1917 (Diptera: Psychodidae). J. Vector. Ecol. 31: 229-238. [ Links ]
Faria, L.D.B., W.A.C. Godoy & L.A. Trinca. 2004. Dynamics of handling time and functional response by larvae of Chrysomya albiceps (Diptera: Calliphoridae) on different prey species. J. Appl. Entomol. 128: 432-436. [ Links ]
Godoy, W.A.C., F.J. Von Zuben, J. Von Zuben & S.F. Reis. 2001. Spatio-temporal dynamics and transition from asymptotic equilibrium to bounded oscillations in Chrysomya albiceps (Diptera, Calliphoridae). Mem. inst. Oswaldo. Cruz. 96: 627-34. [ Links ]
Gomes, L., G. Gomes, O.H. Gutierrez, S.M. Rogério & C.J. Von Zuben. 2006. influence of photoperiod on body weight and depth of burrowing in larvae of Chrysomya megacephala (Fabricius) (Diptera, Calliphoridae) and implications for forensic entomology. Rev. Bras. Entomol. 50: 76-79. [ Links ]
Gião, J. & W. Godoy. 2006. Seasonal population dynamics in Lucilia eximia (Wiedemann) (Diptera: Calliphoridae). Neotrop. Entomol. 35: 753-756. [ Links ]
Hammer, Ø. & D.A.T. Harper. 2011. PAST: Palaeontological Statistics, versión 2.10. (Disponible en: http://folk.uio.no/ohammer/past). [ Links ]
Jaramillo, N.D. & C.M. Wolff. 2002. Geometric Morphometric differences between Panstrongylus geniculatus from field and laboratory. Mem. inst. Oswaldo Cruz 97: 667-673. [ Links ]
Jirakanjanakit, N., S. Leemingsawat, S. Thongrungkiat, C. Apiwathnasorn, S. Singhaniyom, C. Bellec & J.P. Dujardin. 2007. Influence of larval density or food variation on the geometry of the wing of Aedes (Stegomyia) aegypti. Trop. Med. int. Health 12: 1354-1360. [ Links ]
Klingenberg, C.P. & S.D. Zaklan. 2000. Morphological integration between developmental compartments in the Drosophila wing. Evolution 54: 1273-1285. [ Links ]
Liria, J. 2006. insectos de importancia forense en cadáveres de ratas, Carabobo – Venezuela. Rev. Peru. Med. Exp. Salud Pública 23: 33-38. [ Links ]
Loh, R., J.R. David, V. Debat & B.C. Bitner-Mathé. 2008. Adaptation to different climates results in divergent phenotypic plasticity of wing size and shape in an invasive drosophilid. J. Genet. 87: 209-217. [ Links ]
Lomônaco, C. & E. Germanos. 2001. Variações fenotípicas em Musca domestica L. (Diptera: Muscidae) em resposta à competição larval por alimento. Neotrop. Entomol. 30: 223-231. [ Links ]
Lyra, M.L., L.M. Hatadani, A.M.L. De Azeredo-Espin & L.B. Klaczko. 2010. Wing morphometry as a tool for correct identification of primary and secondary New World screwworm fly. Bull. Entomol. Res. 100: 19-26. [ Links ]
Magaña, C. 2001. La entomología forense y su aplicación a la medicina legal. Data de la muerte. Bol. Soc. Entomol. Aragonesa 28: 49-57. [ Links ]
Magaña, C. et al. 2006. Estudio preliminar de la fauna de insectos asociada a cadáveres en Maracay, Venezuela. Entomotropica 21: 53-59. [ Links ]
McAlpine, J.F. 1987. Morphology and Terminology – Adults, p. 9-63. In J.F. McAlpine, B.V. Peterson, G.E. Shewell, H.J. Teskey, J.R. Vockeroth & D.M. Wood (eds.). Manual of Neartic Diptera. Research Branch Agriculture Canada, Otawa, Canada. [ Links ]
Maldonado, M.A. 2002. Entomología forense: Definición, generalidades y fauna relevante. Buenos Aires: Entomología Forense en Argentina. (Disponible en: www.entomologiaforense.unq.edu.ar/intro_es_htm). [ Links ]
Matías, A., J.X. De La Riva, M. Torrez & J.P. Dujardin. 2001. Rhodnius robustus in Bolivia identified by its Wings. Mem. inst. Oswaldo. Cruz 96: 947-950. [ Links ]
Mavárez-Cardozo, M.G., A.i. Espina de Fereira, F.A. Barrios-Ferrer & J.L. Fereira-Paz. 2005. La entomología forense y el neotrópico. Cuad. Med. Forense 11: 23-33. [ Links ]
Queiroz, M.M.C., R.P. Mello & M.M. Lima. 1997. Morphological aspects of the larval instars of Chrysomya albiceps (Diptera, Calliphoridae). Mem. inst. Oswaldo Cruz 92: 187-196. [ Links ]
Reigada, C. & W.A.C. Godoy. 2005. Seasonal fecundity and body size in Chrysomya megacephala (Fabricius) (Diptera: Calliphoridae). Neotrop. Entomol. 34: 163-168. [ Links ]
Ribeiro, P.B. & C.J.B. Carvalho. 1998. Pictorial key to Calliphoridae genera (Diptera) in southern Brazil. Rev. Bras. Parasitol. Vet. 7: 130-140. [ Links ]
Rohlf, F.J. & L. Marcus. 1993. A Revolution in Morphometrics. Trends Ecol. Evol. 8: 129-132. [ Links ]
Rohlf, F.J. 2006. TpsDig, program for Digitizing landmarks and outline for Geometric morphometric analyses, Version 2.04. Departamento de Ecologia y Evolución, Estado Universitario de Nueva York en Stony Brook. (Disponible en: http://life.bio.sunysb.edu/morph/index.html). [ Links ]
Schachter-Broide, J., J.P. Dujardin, U. Kitron & R. Gurtler. 2004. Spatial structuring of Triatoma infestans (Hemiptera, Reduviidae) populations from Northwestern Argentina using wing geometric Morphometry. J. Med. Entomol. 41: 643-649. [ Links ]
Sheets, H.D. 2005a. CoordGen6, Coordinate Generation program for calculating shape coordinates. (Disponible en: http://www3.canisius.edu/~sheets/morphsoft.html). [ Links ]
Sheets, H.D. 2005b. CVAGen6, Canonical Variates Analysis program for the analysis of shape, based on partial warp scores. (Disponible en: http://www3.canisius.edu/~sheets/morphsoft.html). [ Links ]
Sheets, H.D. 2005c. TwoGroup, program for testing significant differences in shape between two groups. (Disponible en: http://www3.canisius.edu/~sheets/morphsoft.html). [ Links ]
Shewell, G.E. 1987. Calliphoridae, p. 675-1332. In J.F. McAlpine, B.V. Peterson, G.E. Shewell, H.J. Teskey, J.R. Vockeroth & D.M. Wood (eds.). Manual of Nearctic Diptera. Research Branche Agriculture Canada, Otawa, Canada. [ Links ]
Smith, K.G. 1986. A manual of forensic entomology. The Trustees of the British Museum (Natural History) and Cornell University, Nueva York, EEUU. [ Links ]
Soto-Vivas, A., C. Rodríguez, R. Bonfante & E. Aldana. 2007. Morfometría geométrica de Triatoma maculata (Erichson, 1848) de ambientes doméstico y peridoméstico, estado Lara, Venezuela. Bol. Mal. Salud. Amb. 47: 231-235. [ Links ]
Soto-Vivas, A., J. Liria & E. De Luna. 2011 Morfometría geométrica y filogenia en Rhodniini (Hemiptera, Reduviidae) de Venezuela. Acta Zool. Mex. 27: 87-102. [ Links ]
Villegas, J., M.D. Feliciangeli & J.P. Dujardin. 2002. Wing shape divergence between Rhodnius prolixus from Cojedes (Venezuela) and Rhodnius robustus from Mérida (Venezuela). infect. Genet. Evol. 2: 121-128. [ Links ]
Von Zuben, C.J. & W.A.C. Godoy. 2001. Larval competition for patchy resources in Chrysomya megacephala (Dipt., Calliphoridae): implications of the spatial distribution of immatures. J. Appl. Entomol. 125: 537-541. [ Links ]
Wells, J.D. & D.W. Williams. 2005. Validation of a DNAbased method identifiying Chrisominae (Diptera: Calliphoridae) used in a death investigation. int. J. Legal Med. 19:1-8. [ Links ]
Whitworth, T. 2006. Keys to the genera and species of blow flies (Diptera: Calliphoridae) of America North of Mexico. Proc. Entomol. Soc. Wash. 108: 689-725. [ Links ]
Zelditch, M, D.L. Swiderski, H.D. Sheets & W.L. Fink. 2004. Geometric morphometrics for biologists: A primer. Elsevier, Boston, EEUU. [ Links ]
*Correspondencia:
Marianela Vásquez: Departamento de Estudios Ambientales, División de Ciencias Biológicas, Universidad Simón Bolívar, Caracas, Distrito Capital. Código Postal 1080, Venezuela. marianela.vasquez@gmail.com
Jonathan Liria: Departamento de Biología, Facultad Experimental de Ciencias y Tecnología, Universidad de Carabobo. Valencia, estado Carabobo. Código Postal 2005, Venezuela. jonathan.liria@gmail.com
1. Departamento de Estudios Ambientales, División de Ciencias Biológicas, Universidad Simón Bolívar, Caracas, Distrito Capital. Código Postal 1080, Venezuela; marianela.vasquez@gmail.com
2. Departamento de Biología, Facultad Experimental de Ciencias y Tecnología, Universidad de Carabobo. Valencia, estado Carabobo. Código Postal 2005, Venezuela; jonathan.liria@gmail.com
Recibido 20-Viii-2011. Corregido 06-ii-2012. Aceptado 07-iii-2012.


{kind=link}
{kind=link}
{kind=link}