










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.60 n.3 San José Sep. 2012
Selection of perching site background color by Hamadryas feronia (Lepidoptera: Nymphalidae) in Costa Rica: Implications for industrial melanism
Luis Ricardo Murillo-Hiller1*
*Dirección para correspondencia
Abstract
Observations of the increased frequency of melanic forms in moths of the genus Biston in Great Britain after the industrial revolution lead to the development of the theory of industrial Melanism. Nonetheless, arguments against that interpretation of the experimental evidence have polarized acceptance of the concept. New evidence based on diurnal butterflies is more credible because it involves behavior that can be seen in action, during daylight, and because the natural history of the selected species is well known. An experiment was carried out in which three substrate colors (white, black, and gray) were employed to test the landing preferences of Hamadryas feronia. A marked preference was observed for landing on white and gray, and a chi-square (N=644 tests) showed evidence of a preference by males to land on white, and for females to land on gray. Black was rejected perhaps because it provides very little background matching with the butterfly’s colors. The butterfly habit of perching selectively on particular color substrates is a genetically fixed behavior, where the males possibly choose white as a tactic to be noticed by females and attract them, whereas females prefer gray to enhance crypsis and avoid attracting predators.
Key words: crypsis, disruptive coloration, camouflage, industrial melanism, behavior.
Resumen
Observaciones en el incremento de la frecuencia de las formas melánicas de la polilla Biston de Gran Bretaña después de la revolución industrial, llevó al desarrollo de la teoría del melanismo industrial. Sin embargo, se originaron argumentos en contra de la interpretación experimental de dicho fenómeno que llevaron a polarizar su aceptación general. Nueva evidencia basada en mariposas diurnas genera nuevas perspectivas puesto que incluye el comportamiento, que puede ser apreciado durante el día. Además, la especie seleccionada es bien conocida desde el punto de vista de su historia natural. El experimento que desarrolle consiste en tres sustratos de diferente color (blanco, negro y gris) en donde se pone a prueba la preferencia de Hamadryas feronia para posarse. Se encontró una marcada tendencia a posarse sobre el blanco y el gris, chi-cuadrado (N=644), donde los machos prefirieron el blanco y las hembras el gris. El negro fue rechazado probablemente porque ofrece muy poca defensa visual al disminuir el efecto de la coloración críptica de esta especie. El habito por parte de esta mariposa de posarse selectivamente sobre sustratos de distinto color esta fijado genéticamente, los machos posiblemente prefieren blanco como una táctica para atraer hembras y las hembras prefieren gris para aumentar el efecto de su coloración críptica y así evitar ser detectadas por los depredadores.
The term camouflage has been used for years even though it generally has been confused with other terms such as disruptive coloration and background matching (Webster et al.2008, Stevens & Merilaita 2009a, Stevens & Merilaita 2009b). Stevens & Merilaita (2009a) proposed that the term camouflage be used to mean all strategies involved in concealment, and Cott (1940) presented circumstantial evidence that concealing coloration plays a major role in secure the safety of otherwise defenseless animals.
The manner in which predators detect their prey depends upon the physical characteristics of the habitat in which they live and the amount and quality of available light (Cot 1940). in general, the visual systems of diurnal animals are adapted to permit the detection of distant preys that are best identified by their shapes (Cott 1940, Troscianko et al. 2009).
Crypsis, therefore, is a key form of camouflage that initially prevents detection, as disruptive coloration, background matching and masquerade do. Another defense mechanism is for the prey to avoid being caught once it has been detected, for example through motion dazzle (Hanlon et al. 2008, Stevens & Merilaita 2009a).
The concepts above presume intense coevolution in visual interactions between predators and prey (Hanlon et al. 2008) because the resting place selected by the prey must be perceived in the same way by both predator and prey. To enhance crypsis during resting, prey often search for backgrounds that mach their body patterns and color reflectance (Kettlewell 1958a, Sargent 1968, Sargent 1969, Steward 1976, Hanlon et al. 2008). in many cases, the chosen backgrounds consist of small to medium-sized objects of moderate contrast, and for that reason mottled camouflage is extremely common in many animals (Hanlon et al. 2008).
One of the most studied and a polemic case of crypsis is the industrial Melanism described in Great Britain for moths of the genus Biston Leach, 1815. According to Kettlewell (1958b) Biston betularia (Geometridae) is one of about 70 species of British macrolepidoptera whose wing patterns fluctuate from light to dark, depending upon the degree of air pollution in their area, in order to enhance crypsis. Nevertheless, a problem is posed by experimental results indicating that melanic individuals of several species prefer light over dark backgrounds (Sargent 1968). Monge-Nájera et al. (1998) obtained data for a diurnal butterfly for which it was not possible to combine a preference for perch color with a preference for bark texture, in fact, he discovered that occasionally the butterflies perched on inappropriate backgrounds, probably when attracted to them by food or to be more visible to females.
Sargent (1985) suggested that some melanic moths may be adapted to exploit white backgrounds that are variegated with black, rather than the uniformly dark backgrounds suggested by the traditional explanation of industrial melanism (Kettlewell 1958a).
Experimental studies of additional species that show cryptic coloration might help to understand in what way some butterflies exploit background matching coloration. Diurnal species are the best experimental subjects because it is possible to observe them in the act of choosing a background, either experimentally or in the wild. Members of the genus Hamadryas are well known butterflies that change their perches many times a day, during daylight hours, and which chose apparently matching substrates. Protection from predators is the reason most accepted for that behavior, until now (Young 1974, Jenkins 1983, Monge-Nájera et al. 1998). These butterflies are influenced by natural selection to choose an appropriate subtract to land on, and if their ability to select it is shown to be deliberate, then new valuable evidence may be available to support the concept of industrial melanism. Hamadryas feronia farinulenta (Fruhstorfer 1916) is a good example of a butterfly that is known to be selective with respect to its perching site (Young 1974, Jenkins 1983, Monge-Nájera et al. 1998). The goal of this research is to test the color preference of H. feronia butterflies experimentally with regard to their choice of resting sites, to investigate how that relates to their biology and behavior, and perhaps to be able to provide a new example of the industrial melanism concept.
Materials and methods
The experiment was carried out between October 2009-July 2010 in San José, Costa Rica, in an open area in the vicinity of the University of Costa Rica campus, at 1 200m above sea level and an average temperature of 27°C. A greenhouse 3m long, 2.5m wide and 2m high, and with black screening allowing 80% light penetration was built in the selected area.
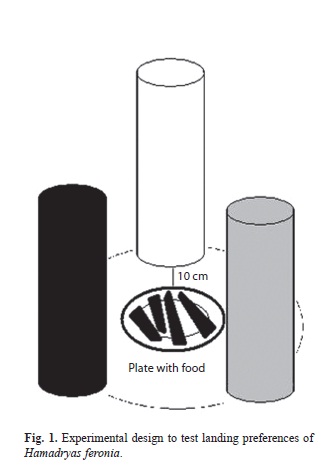
Three iron cylinders 17cm in diameter and 40cm high were built and painted, each with one of three different colors in order to resemble vertical tree trunks. The colors were White 3500 Glidden brand glide-on paint, Black Dark Secret 05/000 Glidden glide-on and gray prepared by mixing the first two colors, 50% each.
The cylinders were set inside the greenhouse, one at each point of an equilateral triangle, and a plate with rotting bananas (food) was placed in the center of the triangle. The rotting fruits were 10cm from each of the cylinders, and for every individual butterfly tested, the cylinders position were changed randomly to reduce the influence of external factors (Fig. 1).
The H. feronia used for the experiments were caught in second growth forest in La Guácima of Alajuela, Costa Rica, 9°56’36.88” N - 84°15’37.53” W; at 750m where they were very common from October 2009-July 2010. The butterflies were caught and kept alive in a temporary cage for two days without food to accelerate their decision making during the experiments. Finally, the butterflies were released one by one into the greenhouse, and their behavior was observed for 5-20 minutes until they chose a resting place. in total, 301 females and 343 males were tested, with an average observation time of 85 hours.
Results
When each butterfly was released into the testing cage, they flew around as patrolling the area. Most commonly, they would fly for few minutes until attempting to land on any of the cylinders. The specimens reacted equally to all the cylinders independently of the position or color; sometimes they landed at once, and some other times they landed for less than a second, and flew again until selecting a definitive landing place. For this experiment, if the butterfly landed for more than ten seconds, i recorded it as a valid data for the respective substrate color, and then i removed the insect from the greenhouse.
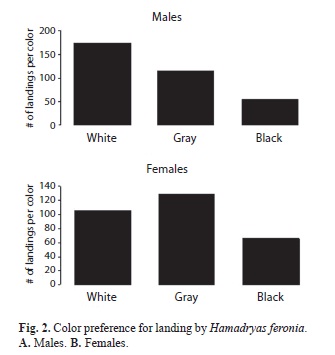
Males landed mostly on the white background, and secondly on the gray background; (X2, p<0.001, Fig. 2a). Females landed mostly on gray and secondly on white (X2, p<0.001, Fig. 2b).
Discussion
A preference for selecting a certain background color to land has been demonstrated in the past for several moth species (Kettlewell 1958a, Sargent 1966, Sargent 1968, Sargent 1969, Steward 1976), although, in the case of diurnal butterflies very few studies have experimentally proven this conclusion (Monge- Nájera et al. 1998). The results in this paper are supported by the field observations of other researchers who also concluded that Hamadryas species select certain background colors that better match their appearance (Young 1974, Jenkins 1983, Monge-Nájera et al. 1998).
According to the results presented, it is clear that H. feronia can distinguish between black, white and gray and actively reject black, which offers no crypsis to either gender, a result consistent with those of Monge-Nájera et al. (1998), which showed that perching on cryptic backgrounds was more frequent. Besides, these tests suggest that males and females have different preferences at the time of selecting a place to land. Statistical analysis showed a marked preference for females to perch on gray, which makes sense because of the increased crypsis and makes females employ more energy in reproduction: eggs represent a biggest investment than sperm and they emphasize protection over finding a mate quickly (Hickman et al. 1998). Males on the other hand, are more compelled to find a mate despite a possible decrease in their fitness (Hickman et al. 1998) because living without reproducing is like dying young (Freeman & Herron 2002). An additional possibility is that this behavior reflects sexual selection: maybe males prefer to land on white where they are more visually-evident to females, so with predators. This may evolve if females select males with faster reaction responses to predator attacks (Hickman et al. 1998). This phenomenon was reported in butterflies that have brilliant coloration, sacrificing crypsis and increasing the predation risk (Scoble 1992).
The colors employed in the experiments were solid, white, gray or black, in contrast to the mottled tree trunks found in the wild. This is important because in nature a butterfly’s decision to land is easier because mottled surfaces represent better matches with their wing patterns and thus provides better protection (Hanlon et al. 2008).
Methodological improvements are important to consider for future experiments. A weak point of the experiment is that the black color absorbs more radiation from de sun light than the other two colors, this could have an effect on the substrate surface temperature, favoring butterflies avoid it. This variable is important to keep in consideration for future studies; nevertheless, i did not observe any particular behavior that supports this argument; besides, during cloudy days when the temperature was lower in all of the cylinders, the behavior was similar to the observed in sunny conditions.
Color perception is also important to consider when studying substrate selection in diurnal butterflies. Butterflies do not perceive the colors the same way we do, since they are also sensible to ultraviolet wavelength (Menzel & Backhaus 1991). in any case, this is not a weak point of the experimental design, because the tree trunks where Hamadryas has evolved to land on, has no ecological reason to hide ultraviolet colors. The color we see in the bark of a tree is the same the butterflies perceive, even when they can see a wider spectrum of wavelength.
Another aspect of these results is their relevance to the concept of industrial melanism. Hamadryas feronia may be considered a better example to explain the process of background selection than Biston betularia from Great Britain, because its natural history and taxonomy are well known and its behavior can be seen during daylight (Jenkins 1983, Monge-Nájera & Hernandez 1991, Monge-Nájera 1992, Monge-Nájera et al. 1998).
Some unclear points in previous research include melanic forms perching on white surfaces (Sargent 1968), which could be attributed to the fact that the melanism is not genetically linked to the behavior, which needs more time to be fixed by natural selection (Sargent 1985). Melanic forms of the moths behave as normal because they do not know they are black as has been shown by Sargent (1968), and contrary to Kettlewell (1955) assumptions. instead, H. feronia is a well known species that has fixed both, genetically and morphological together, and therefore evidence of behavior increasing fitness related to wing coloration support the industrial melanism theory without the methodological problems that B. betularia presents (Monge-Nájera et al. 1998).
Acknowledgments
I thank J. Monge-Nájera for important discussion and for providing helpful literature, to Federico Bolaños for statistical support and also to Paul Hanson and Adrea González- Karlsson for providing critical comments and a review of the manuscript.
References
Cott, H.B. 1940. Adaptive coloration in animals. Methuen, London, United Kingdom. [ Links ]
Freeman, S. & J.C. Herron. 2002. Análisis Evolutivo. Pearson, Madrid, España. [ Links ]
Hanlon, R., C. Chiao, L. Mäthger, A. Barbosa, K. Buresch & C. Chubb. 2008. Cephalopod dynamic camouflage: bridging the continuum between background matching and disruptive coloration. Phil. Trans. R. Soc. B. 364: 429-437. [ Links ]
Hickman, C.P., L.S. Roberts & A. Larson. 1998. Principios integrales de zoología. McGraw-Hill, interamer, Madrid, España. [ Links ]
Jenkins, D.W. 1983. Neotropical Nymphalidae. i. Revision of Hamadryas. Bull. Allyn. Mus. 81: 1-146. [ Links ]
Kettlewell, H.B.D. 1955. Selection experiments on industrial melanism in the Lepidoptera. Heredity 9: 323-342. [ Links ]
Kettlewell, H.B.D. 1958a. The importance of the microenviroment to evolutionary trends in the Lepidoptera. Entomologist 91: 214-224. [ Links ]
Kettlewell, H.B.D. 1958b. A survey of the frequencies of Biston betularia L. (Lep.) and its melanic forms in Britain. Heredity 12: 51-72. [ Links ]
Menzel, R. & W. Backhaus 1991. Colour vision in insects, p. 262-293. In P. Gouras (ed.). vision and visual dysfunction: perception of colour, vol. 7. Macmillan, Houndsmills, United Kingdom. [ Links ]
Monge-Nájera, J., F. Hernandez, M.i. Gonzáles, J. Soley, J. Araya & S. Zolla. 1998. Spatial distribution, territoriality and sound production by tropical cryptic butterflies (Hamadryas, Lepidoptera: Nymphalidae): implications for the ¨industrial melanism¨ debate. Rev. Biol. Trop. 46: 297-330. [ Links ]
Monge-Nájera, J. 1992. Clicking Butterflies, Hamadryas, of Panama: Their biology and classification (Lepidoptera, Nymphalidae), p. 567-572. In D. Quintero & A. Aiello (eds.). insects of Panama and Mesoamerica: Selected Studies. Oxford University, Oxford, United Kingdom. [ Links ]
Monge-Nájera, J. & F. Hernández. 1991. A morphological search for the sound mechanism of Hamadryas butterflies. J. Res. Lepid. 30: 196-208. [ Links ]
Sargent, T.D. 1966. Background selection of geometrid and noctuid moths. Science 154: 1674-1675. [ Links ]
Sargent, T.D. 1968. Cryptic moths: effects on background selections of painting the circumocular scales. Science 159: 100-101. [ Links ]
Sargent, T.D. 1969. Behavioural adaptations of cryptic moths. i. Preliminary studies on bark-like species. J. Lepid. Soc. 23: 1-9. [ Links ]
Sargent, T.D. 1985. Melanism in Phigalia titea (Cramer) (Lepidoptera: Geometridae) in southern New England: a response to forest disturbance? J. New York Entomol. Soc. 93: 1113-1120. [ Links ]
Scoble, M. 1992. The Lepidoptera: form, function and diversity. Oxford University, New York, USA. [ Links ]
Stevens, M. & S. Merilaita. 2009a. Animal camouflage: current issues and new perspectives. Phil. Trans. R. Soc. B. 364: 423-427. [ Links ]
Stevens, M. & S. Merilaita. 2009b. Defining disruptive coloration and distinguishing its functions. Phil. Trans R. Soc. B. 364: 481-488. [ Links ]
Steward, R.C. 1976. Experiments on resting site selection by the typical and melanic forms of the moth, Allophyes oxyacanthae (Caradrinidae). J. Zool. Lond. 178: 107-115. [ Links ]
Troscianko, T., C. Benton, P. Lovell, D. Tolhurst & Z. Pizlo. 2009. Camouflage and visual perception. Phil. Trans. R. Soc. B. 364: 449-461. [ Links ]
Webster, R., A. Callahan, J. Godin & T. Sherratt. 2008. Behaviourally mediated crypsis in two nocturnal moths with contrasting appearance. Phil. Trans. R. Soc. B. 364: 503-510. [ Links ]
Young, A.M. 1974. On the biology of Hamadryas februa (Lepidoptera: Nymphalidae) in Guanacaste, Costa Rica. Z. Ang. Ent. 76: 380-393. [ Links ]
*Correspondencia:
Luis Ricardo Murillo-Hiller: Universidad de Costa Rica, Mariposario de la Escuela de Biología. 2060 San José, Costa Rica. murillo.luis@lycos.com
1. Universidad de Costa Rica, Mariposario de la Escuela de Biología. 2060 San José, Costa Rica; murillo.luis@lycos.com
Received 11-VII-2011. Corrected 20-I-2012. Accepted 27-II-2012.


{kind=link}
{kind=link}