










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.60 n.3 San José Sep. 2012
Defense response of susceptible and resistant Biomphalaria alexandrina snails against Schistosoma mansoni infection
*Dirección para correspondencia
Abstract
In Egypt, Biomphalaria alexandrina is the intermediate host for Schistosoma mansoni. The fates of Schistosoma miracidia in the snails varies between different species of Biomphalaria. The internal defense system is one of the factors that influence the susceptibility pattern of the snails. The interaction between Biomphalaria snails and S. mansoni needs to be identified for each species, and even between the members of the same species with different degrees of susceptibility. In the present study, the first generation of susceptible and resistant parents of B. alexandrina was examined histologically at the 30th day post exposure. The study includes the characterization of the immune response, as expressed by tissue reactions, of susceptible and resistant B. alexandrina snails against S. mansoni. It was also designed to determine the impact of the resistance increase in parent snails, on the mechanisms of interaction of their offspring against infection. The results showed that the infection rate of the offspring from the susceptible parents was 92%. No susceptible offspring was produced from the resistant parents. When the parents were of equal number of susceptible and resistant snails, they gave an offspring with an infection rate of 20%. Susceptible snails that had susceptible parents showed a higher degree of susceptibility than those that had both susceptible and resistant parents.
A common feature of the resistant snails was the absence of any viable parasites. The tissue reactions of the resistant snails having only resistant parents occurred at the site of miracidial penetration. In resistant snails for which susceptible ones were included in their parents, the reactions occurred in the deep tissues. These results characterized the immune response of B. alexandrina snails against Schistosoma infection which was found to occur by two different mechanisms. One type of defense occurs in highly resistant snails, and employs direct miracidial destruction soon after parasite penetration. The other type occurs in less resistant snails where a delayed resistance development occurs after the dissemination of the sporocysts in the snail tissues. It seems that B. alexandrina snails respond more or less similar to B. glabrata. The results also proved that the immune response of the internal defense system increased with increasing the number of the inherited resistant genes.
Key words: Biomphalaria alexandrina, Schistosoma mansoni, susceptible, resistant, immune response, hemocyte, histopathology.
Resumen
En Egipto, Biomphalaria alexandrina es el huésped intermediario de Schistosoma mansoni. La supervivencia de los miracidios de Schistosoma en los caracoles varía entre las especies de Biomphalaria. El sistema de defensa interno es uno de los factores que influyen en el patrón de susceptibilidad de los caracoles. La interacción entre los caracoles Biomphalaria y S. mansoni requiere ser identificada para cada especie e incluso, entre los miembros de la misma especie con diferente grado de susceptibilidad.
En el presente estudio, la primera generación de padres susceptibles y resistentes de B. alejandrina fue examinada histológicamente al día 30, después de la exposición. El trabajo fue realizado tanto para caracterizar la respuesta inmune, según las reacciones de los tejidos, de los caracoles susceptibles y resistentes de B. alejandrina contra S. mansoni. También, el estudio se diseñó para determinar el impacto en el aumento de la resistencia en los caracoles padres, en los mecanismos de interacción de sus crías contra la infección. Los resultados mostraron que la tasa de infección para las crías, de padres susceptibles, fue del 92%. No se originaron crías susceptibles de los padres resistentes. Cuando los padres incluían un número igual de caracoles susceptibles y resistentes, dieron como resultado crías con una tasa de infección del 20%. Los caracoles susceptibles que tuvieron padres susceptibles mostraron un mayor grado de susceptibilidad que los que tenían tanto padres sensibles como resistentes. Una característica común de los caracoles resistentes fue la ausencia de parásitos viables. Las reacciones en los tejidos de los caracoles resistentes de sólo padres resistentes ocurrió en el sitio de penetración del miracidio. En los caracoles resistentes, para los que variedades susceptibles fueron incluídas entre sus padres, las reacciones se produjeron en tejidos profundos.
Los resultados caracterizaron la respuesta inmune de los caracoles B. alexandrina contra la infección por Schistosoma, que ocurre por dos mecanismos diferentes. El primer tipo de defensa la cual se produce en los caracoles con alta resistencia, utiliza la destrucción directa del miracidio poco después de la penetración de los parásitos. El segundo tipo se produce en los caracoles menos resistentes, en el cual se después de la difusión de los esporocistos en los tejidos del caracol. Parece que los caracoles B. alexandrina responden de una manera más o menos similar a B. glabrata. Los resultados también demostraron que la respuesta inmune del sistema de defensa interna aumentó cuando en el número de genes de resistencia heredados es mayor.
Palabras clave: Biomphalaria alexandrina, Schistosoma mansoni, susceptibilidad, resistencia, respuesta inmune, hemocitos, histopatología.
Biomphalaria alexandrina is the common aquatic snails that serve as an intermediate host for Schistosoma mansoni in Egypt. The fates of Schistosoma miracidia that penetrate different species of Biomphalaria vary from destruction within hours to productive infections that yield human-infective cercariae several weeks later. To understand the mechanisms used by the resistant snails to destroy the parasites, or those used by the parasites to ensure their survival in their suitable hosts, one must comprehend the internal defense mechanisms that are available to the host (Bayne 2009). The internal defense system (IDS) is one of the factors that influence the susceptibility pattern of the snails. This system is stimulated by the excretory/secretory products of the penetrating miracidia (Zahoor et al. 2010). Its main action is mediatedby the phagocytic hemocytes in cooperation with humoral components (Negrão-Corrêa et al. 2007). The interaction between S. mansoni and the IDS of the snail suggests that the hemocytes could be the main effecter element in the destruction of the parasite. The hemocytes are directly involved in the death of some encapsulated parasites (Van der Knaap & Loker 1990) or in the production of soluble factors which could be cytotoxic (Connors et al. 1995). However, the parasite can escape the IDS by two mechanisms, molecular mimicry and antigenic masking. In the molecular mimicry, the parasite expresses glycoprotein epitopes on its surface that mimic host molecules, while the antigenic masking is the absorption and incorporation of the snail agglutinins and hemolymph soluble components to the parasite surface (Thompson 2001).
Understanding the host-parasite interactions represents a major challenge in biology and may help in finding a novel control method against Schistosoma in different snail species. Differences in the immune responses were represented by different species of Biomphalaria against Schistosoma infection and even between members of the same species with different degrees of susceptibility (de Souza et al. 1997, Borges et al. 1998). So, the mechanisms involved in these interactions need to be specified for each species.
The outcome of the interplay between Biomphalaria snail defense system and the invading Schistosoma parasite is determined by the level of concordance of the genetically determined characters (Basch 1976). Classical genetics showed, unsurprisingly, that the genotypes of both partners determine the outcome of each infection (Basch 1976, Abou El Naga et al. 2010). The present work was designed to determine the impact of increasing the resistance of B. alexandrina parent snails on the mechanisms of interaction and the tissue reactions of their offspring against S. mansoni infection. This will help with the characterization of the immune response of B. alexandrina species against S. mansoni infection.
Material and Methods
The S. mansoni cycle was propagated at the laboratory of the Parasitology department, Faculty of Medicine, Alexandria University, between B. alexandrina snails and Swiss strain albino mice. Initially, the snails were collected from the water channels at Alexandria Governorate, and the miracidia of S. mansoni were obtained from eggs in the feces of untreated children from the same area.
The miracidia were collected from S. mansoni eggs obtained from the liver of infected mice, seven-eight weeks post infection (El-Gindy et al. 1985). The eggs were exposed to direct sunlight for approximately 30min to stimulate miracidial hatching.
A total of 200 juvenile snails at the age of two months, and the size of 3-4mm in diameter were challenged by exposure of each snail to ten active miracidia for 4-5hr, at day light and at room temperature (Henning & Youssef 1976). Groups of 25 snails were transferred to separate containers, each contained 2.5L of dechlorinated tap water (DTW). They were kept in the darkness at 26ºC for about four weeks. Fresh lettuce leaves were supplied as food every couple of days and the dead snails were regularly removed (Smithers & Terry 1965).
The live exposed snails were examined microscopically every week for the presence of the larval stages of S. mansoni during four weeks. 30 days post infection; the snails were checked individually for the cercarial shedding (McClelland 1965). The snails that shed cercariae were considered susceptible, while those that failed to shed cercariae were examined alive under the microscope for detection of any delay in larval migration. In absence of any delay, these snails were considered resistant.
To obtain the first generation (F1), 45 susceptible and 45 resistant snails were crossed in different proportions, to get the three experimental groups, as follows: Group I: 30 susceptible snails, Group II: 30 resistant snails, and Group III: 15 susceptible and 15 resistant snails. The snails of group I and II were reared together, each group in a separate aquarium.
Those of group III were reared in beakers; each containing one susceptible and one resistant snail. Each aquarium and beaker contained DTW and pieces of foam that served as the substratum for the egg deposition. To avoid fertilized eggs before the beginning of the experiment, we discarded egg batches from each of the three groups during the first four weeks. After the fourth week, the newly deposited batches of each group were regularly collected using a scalpel, transferred to a separate container containing DTW and inspected daily until hatching. Baby snails were reared for one month until they became juveniles, and were fed boiled lettuce leaves during one month. White chalk pieces were added as a source of calcium for the growth of the snail shell (Dettman et al. 1989).
A total of 100 F1 juvenile snails were used from each experimental group excluding the dead snails during the experiment. Three uninfected snails from each group were used as a control and the remaining snails were exposed to ten S. mansoni miracidia. This study included the end of the experiment from each group. Susceptibility/resistance was determined as mentioned before.
Light microscopic study: Five susceptible and five resistant snails form each group, if present, together with the three uninfected snails were subjected to histological examination. They were relaxed with methanol crystals (approximately 2x10-4M), separately crushed between two microscope slides, and then the broken shell was pulled away from the body. The columellar muscle was separated from the shell, and the snail was extracted intact. The snails were fixed in Bouin’s fixative for at least 24hr and then placed in gradually increasing concentrations of ethanol. Hematoxyline and Eosin stained 5μm sections that were examined microscopically. The grading of the diffuse cellular infiltration and the granuloma formation were carried out by two independent observers.
Ethical approval: All animal studies presented here have been approved by the local government based on national regulations for animal experimentation.
Results
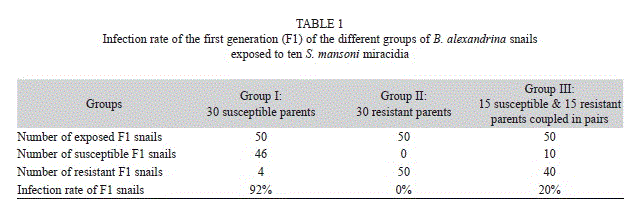
All the snails that showed larval developmental stages of S. mansoni during the microscopic inspection shed cercariae at the 30th day post exposure. No delay in the larval migration was observed. Table 1 shows the infection rate of the first generation (F1) of the different studied groups infected with ten S. mansoni miracidia. The infection rate of the offspring from the susceptible parents was 92%. No susceptible offspring was produced from the resistant parents. When the parents originated on equal numbers of susceptible and resistant snails, they gave an offspring with an infection rate of 20%.
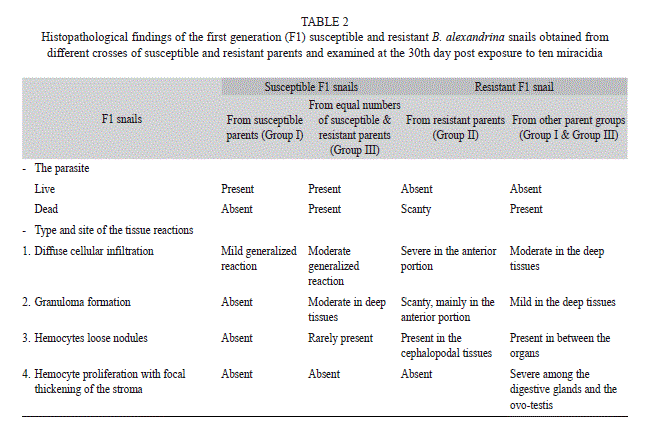
Histopathological findings of the infected F1 offspring shedding cercariae at the 30th day post exposure (Table 2): The infected susceptible F1 generation that had susceptible parent snails (Group I) showed a high degree of susceptibility. Large numbers of live parasites were present with normal development (Fig. 1). The parasites were widely spread in different organs specially the digestive glands and ovotestis. No cellular reaction was present around the parasite. Mild generalized diffuse cellular infiltration was present in the tissues and in between the organs.
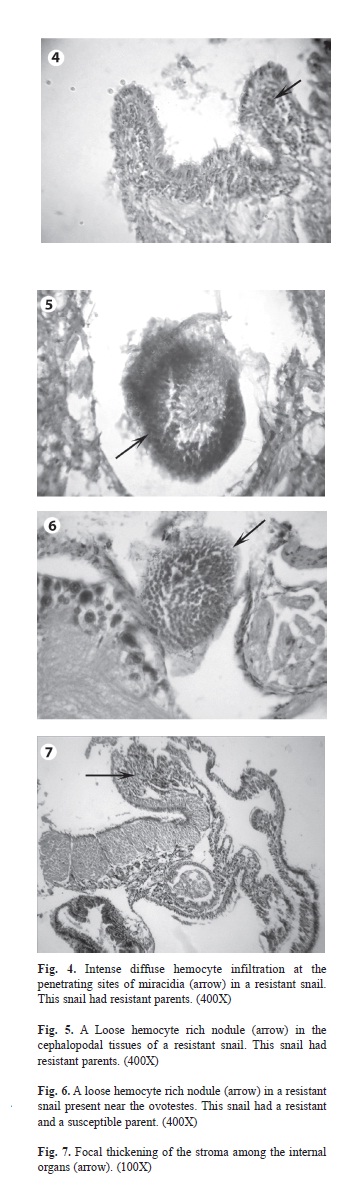
Histopathological findings of the infected F1 offspring failed to shed cercariae at the 30th day post exposure (Table 2): A common feature of these snails was the absence of viable only (Group II) contained very few numbers of dead parasites and granulomata. These snails showed an intense diffuse hemocytic reaction at the penetrating sites of miracidia (Fig. 4). Loose hemocyte rich nodules were detected in the cephalopodal tissues (Fig. 5).
Dead parasites were present in the resistant infected F1 snails that had parents from the other groups (Group I and Group III). The most common type of the dead parasites appeared as round eosinophilic masses surrounded by several layers of flattened hemocytes. The tissue reaction was in the form of moderate diffuse cellular infiltration in between the organs. The two types of granulomata with the same structure described before surrounding the dead parasites were present in between the organs. Loose hemocyte rich nodules were present in between the organs (Fig. 6).
Hemocyte proliferation with focal thickening of the stroma was present among the digestive glands, the albumin glands and the ovo-testis (Fig. 7). The control non infected snails from different groups showed normal histology.
Discussion
The present work is a comparative histopathological study verifying the tissue reactions of B. alexandrina snails with different degrees of resistance against the development of S. mansoni. The studied snails were the offspring of resistant and susceptible parents crossed in different proportions.
The number of the susceptible and the resistant F1 progeny in table 1 showed that in B. alexandrina, the susceptibility/resistance to S. mansoni is inherited as shown in different Biomphalaria species (Lewis et al. 2003, Rosa et al 2005, Abou El Naga et al. 2010)
In this study, histopathological examination of the susceptible offspring having only susceptible parents showed a normal development of the parasites with wide spread of cercariae in the different organs. There were neither dead parasites nor cellular reactions around the living ones. It seems that there is a tolerance in the snail tissue to the presence, growth and multiplication of the larval stages of the parasite (Newton 1952). However, some degree of the host tissue reactions was exhibited by the susceptible offspring having an equal number of susceptible and resistant parents. This was presented by the cellular infiltration surrounding the living parasites, and by the formation of the granuloma around some dead parasites. In spite of the presence of these host reactions, the susceptible B. alexandrina snails and those of other species were not able to clear the infection (de Souza et al. 1997, El-Assal et al. 2001, Soomro et al. 2005). Parasitological studies of different Biomphalaria species showed that within the same species a smaller number of cercariae was shed from the susceptible offspring, having a proportion of resistant snails as parents, than those having susceptible parents only (Rosa et al 2005, Abou El Naga et al. 2010). These parasitological results could be explained by the previous immune reactions in the snail tissues.
Our results also showed a generalized hemocytic infiltration in the susceptible snails at the 30th days post exposure. This period represents the early phase of massive colonization of the snail tissue by the cercariae; the very pathogenic mobile larvae that are capable of directly ingesting the host tissues. The the migrating cercariae which themselves may be protected by masking the snail antigens may be responsible for this reaction (Van der Knaap & Loker 1990). At that time, it was hypothesized that the tissue reactions eliminate the waste products resulting from the parasites, and participate in the healing of the injured tissues (Vasquez & Sullivan 2001). By this time the hemocytes have been exposed for a long time to “non-self” and “changed self” (damage tissue caused by escaping cercariae) resulting in a higher state of responsiveness of the hemocytes (Bayne 1990, Cooper et al. 1994).
The resistant snails showed many forms of tissue reactions against the parasite, including diffuse cellular infiltration with phagocytosis, granuloma formation, hemocyte rich nodules and focal thickening of the stroma. Phagocytosis was evident by the absence of viable parasites and the presence of remnants of dead forms. As the number of dead parasites was scanty in the resistant snails having only resistant parents, this implied that a more vigorous phagocytosis had occurred. Loker et al. (1986) found that parasite-amebocyte contact occurred and led to phagocytosis and by 48hr only scattered remnants of sporocysts remained.
The two types of granulomata formed in the present study around the dead parasites were described by Lie & Heyneman (1976) as type 1 and type 2 granulomata. Basch (1975) attributed this variability to the individual difference in both the host and the parasite. Both types of granulomata appeared in the routinely stained slides as a mixture of hemocytes and fibers. In snails, the participation of the extracellular tissue in the granulomatous lesions is controversial (Yoshino 1976, Krupa et al. 1977). Borges & Andrade (2003) found that the hemocytes in the granuloma had expanded cytoplasmic processes that gave the appearance under light microscope as containing fibers.
The hemocyte rich nodules are another form of tissue reactions detected in the present study, mainly in the resistant snails and occasionally in the susceptible ones. They were found in the anterior portion of the snails and in between the organs. These nodules were also present in the strongly resistant B. glabrata and B. tenagophila (Godoy et al. 1997, Borges et al. 1998, Negrão-Corrêa et al. 2007). After doing experiments with different snail crosses, Lewis et al. (2001) suggested that a single gene controlled this reaction. They expected that certain hemocyte functions are under genetic control, independent of their response to schistosomiasis.
By observing the tissue reactions of B. alexandrina snails in this study, we can suggest that the immune responses occurred by two different mechanisms. One type of defense employs direct miracidial destruction soon after penetration of the parasite. This type occurred in the resistant offspring having only resistant parents. In these snails, the intense hemocytic infiltration and the hemocyte rich nodules were present at the site of the penetration of the miracidia. These reactions may lead to direct miracidial destruction soon after their penetration. In B. tenagophila Taim strain, a diffuse and focal hemocytic infiltration were observed in the cephalopodal tissue of the infected highly resistant snails and were found to be associated with rapid parasite destruction after penetration (Negrão-Corrêa et al. 2007).
The other type of the immune reactions occurred in the resistant snails in which their parents included susceptible snails. Their histopathological examination showed that the diffuse cellular infiltration, the hemocyte rich nodules and the focal thickening of the stroma were present in the deep tissues. Godoy et al. (1997) and Borges et al. (1998) considered these reactions as a delayed development of resistance that occurred after dissemination of sporocysts in the snail tissues. They considered this type of delayed developed resistance represents an alternative kind of host defense mechanism against S. mansoni miracidia. It occurs in spite of the evidence suggesting that S. mansoni sporocysts can sometimes develop their ability, in a better way, to interfere with the defense mechanism of the snail as they grow older (Lie et al. 1980).
It was found that when B. tenagophila and B. straminea were exposed to S. mansoni they utilized the first mechanism of resistance described before. This is because of their strong resistance and their efficient defense system that destroys miracidia once they have penetrated (de Souza et al. 1997, Grassi et al. 2001, Negrão-Corrêa et al. 2007). On the other hand, B. glabrata which is more susceptible to S. mansoni than the other two snail species used the second mechanism of defense (de Souza et al. 1997). However, when B. glabrata was challenged by an incompatible strain of S. mansoni, the parasites were immediately recognized and killed, mainly by the strong phagocytic activity of the cells (Lie et al. 1980, Roger et al. 2008).
Our results characterized the immune response, as expressed by the tissue reactions, of susceptible and resistant B. alexandrina snails against the exposure to S. mansoni. It seems that B. alexandrina snails respond more or less similar to B. glabrata. Moreover, the present work is designed to determine the impact of increasing the resistance of the parent snails on the mechanisms of interaction of their offspring against B. alexandrina S. mansoni exposure. The results proved that the immune response of the IDS increased with increasing the number of the inherited resistant genes.
Acknowledgments
Thanks to Samia Ibrahim, for her technical assistance in the snail rearing.
References
Abou El Naga, I.F., M.M. Eissa, S.F. Mossallam & S.I. Abd El-Halim. 2010. Inheritance of Schistosoma mansoni infection incompatibility in Biomphalaria alexandrina snails. Mem. Inst. Oswaldo Cruz 105: 149-154. [ Links ]
Basch, P.F. 1975. An interpretation of snail-trematode infection rates: specificity based on concordance of compatible phenotypes. Int. J. Parasitol. 5: 449-452. [ Links ]
Basch, P.F. 1976. Intermediate host specificity in Schistosoma mansoni. Exp. Parasitol. 39: 150-169. [ Links ]
Bayne, C.J. 1990. Phagocytosis and non-self recognition in invertebrates. BioScience 40: 723-731. [ Links ]
Bayne, C.J. 2009. Successful parasitism of vector snail Biomphalaria glabrata by the human blood fluke (trematode) Schistosoma mansoni: A 2009 assessment. Mol. Biochem. Parasitol. 165: 8-18. [ Links ]
Borges, C.M. & Z.A. Andrade. 2003. Extra-cellular matrix changes in Schistosoma mansoni-infected Biomphalaria glabrata. Mem. Inst. Oswaldo Cruz 98: 135-139. [ Links ]
Borges, C.M., C.P. de Souza & Z.A. Andrade. 1998. Histopathologic features associated with susceptibility and resistance of Biomphalaria snails to infection with Schistosoma mansoni. Mem. Inst. Oswaldo Cruz 93: 117-121. [ Links ]
Connors, V.A., I. de Buron & W.O. Granath Jr. 1995. Schistosoma mansoni: interleukin-1 increases phagocytosis and superoxide production by hemocytes and decreases output of cercariae in schistosome-susceptible Biomphalaria glabrata. Exp. Parasitol. 80: 139-148. [ Links ]
Cooper, L., C. Richards, F. Lewis & D. Minchella. 1994. Schistosoma mansoni: relation between low fecundity and reduced susceptibility to parasite infection in the snail, Biomphalaria glabrata. Exp. Parasitol. 79: 21-28. [ Links ]
de Souza, C.P., C.C. Borges, A.G. Santana & Z.A. Andrade. 1997. Comparative histopathology of Biomphalaria glabrata, B. tenagophila and B. straminea with variable degrees of resistance to Schistosoma mansoni miracidia. Mem. Inst. Oswaldo Cruz 92: 517-522. [ Links ]
Dettman, C.D., S.B. Higgins-Opitz & A. Saikoolal. 1989. Enhanced efficacy of paddling method for schistosoma infection of rodents by a four-step pre-soaking procedure. Parasitol. Res. 76: 183-184. [ Links ]
El-Assal, F.M., N.M. Shoukry, M.H. Abdalla & A.H. Saad. 2001. Cellular response to Schistosoma mansoni infection in Biomphalaria alexandrina strains selected for susceptibility and resistance. J. Egypt Soc. Parasitol. 31: 915-938. [ Links ]
El-Gindy, M.S., M.S. Arafa, M.K. Makled, M.M. Ismail, M.M. Elridi, M.M. Atia & R.L. el-Gamal. 1985. Effect of some factors upon the susceptibility of Biomphalaria alexandrina to infection with Schistosoma mansoni. J. Egypt Soc. Parasitol. 15: 231-235. [ Links ]
Godoy, A., C.P. de Souza, C.T. Guimarães & Z.A. Andrade. 1997. Unusual histological findings in Biomphalaria glabrata with high degree of resistance to Schistosoma mansoni miracidia. Mem. Inst. Oswaldo Cruz 92: 121-122. [ Links ]
Grassi, L., M. Torres Jordá, Z. Andrade & S.M. González- -Cappa. 2001. Short report: Schistosoma mansoni miracidia are killed by the defense system of an Argentine strain of Biomphalaria straminea. Am. J. Trop. Med. Hyg. 65: 290-292. [ Links ]
Henning, J. & G. Youssef. 1976. Influence of diet on breeding and infectivity in mass cultivation of Biomphalaria glabrata. Egypt J. Bilharz. 3: 45-55. [ Links ]
Krupa, P.L., L.M. Lewis & P. Del Vecchio. 1977. Schistosoma haematobium in Bulinus guernei: electron microscopy of hemocyte-sporocyst Interactions. J. Invertebr. Pathol. 30: 35-45. [ Links ]
Lewis, F.A., C.N. Patterson & C. Grzywacz. 2003. Parasite- susceptibility phenotypes of F1 Biomphalaria glabrata progeny derived from interbreeding Schistosoma mansoni-resistant and -susceptible snails. Parasitol. Res. 89: 98-101. [ Links ]
Lewis, F.A., C.N. Patterson, M. Knight & C.S. Richards. 2001. The relationship between Schistosoma mansoni and Biomphalaria glabrata: genetic and molecular approaches. Parasitology 123: 169-179. [ Links ]
Lie, K.J. & D. Heyneman. 1976. Studies on resistance in snails. 3. Tissue reactions to Echinostoma lindoense sporocysts in sensitized and resensitized Biomphalaria glabrata. J. Parasitol. 2: 51-58. [ Links ]
Lie, K.J., K.H. Jeong & D. Heyneman. 1980. Tissue reaction induced by Schistosoma mansoni in Biomphalaria glabrata. Ann. Trop. Med. Parasitol. 78: 157-166. [ Links ]
Loker, E.S., C.J. Bayne & M.A. Yui. 1986. Echinostoma paraensei: hemocytes of Biomphalaria glabrata as targets of Echinostoma mediate interference with host snails resistance to Schistosoma mansoni. Exp. Parasitol. 62: 149-154. [ Links ]
McClelland, W.J. 1965. The production of cercariae by Schistosoma mansoni and Schistosoma haematobium and methods for estimating the numbers of cercariae in suspension. Bull. World Health Organ 3: 270-276. [ Links ]
Negrão-Corrêa, D., C.A.J. Pereira, F.M. Rosa, R.L. Martins- Souza, Z.A. Andrade & P.M.Z. Coelho. 2007. Molluscan response to parasite: Biomphalaria and Schistosoma mansoni interaction. J. Invert. Survival 4: 101-111. [ Links ]
Newton, W.L. 1952. The comparative tissue reaction of two strains of Australorbis glabratus to infection with Schistosoma mansoni. J. Parasitol. 38: 362-366. [ Links ]
Roger, E., G. Mitta, Y. Moné, A. Bouchut, A. Rognon, C. Grunau, J. Boissier, A. Théron & B.E. Gourbal. 2008. Molecular determinants of compatibility polymorphism in the Biomphalaria glabrata/Schistosoma mansoni model: New candidates identified by a global comparative proteomics approach. Mol. Biochem. Parasitol. 157: 205-216. [ Links ]
Rosa, F.M., A.L. Godard, V. Azevedo & P.M. Coelho. 2005. Biomphalaria tenagophila: dominant character of the resistance to Schistosoma mansoni in descendants of crossbreedings between resistant (Taim, RS) and susceptible (Joinville, SC) strains. Mem. Inst. Oswaldo Cruz 100: 19-23. [ Links ]
Smithers, S.R. & R.J. Terry. 1965. The infection of laboratory hosts with cercariae of Schistosoma mansoni and the recovery of the adult worms. Parasitology 55: 695-700. [ Links ]
Soomro, N.M., A.G. Arijo, T.A. Qureshi, N.W. Runham & M.J. Doenhoff. 2005. Pathology of Schistosome infection on host tissue during developmental stages of parasite in vector snails. Int. J. Agri. Biol. 7: 133-141. [ Links ]
Thompson, R.C. 2001. Molecular mimicry in schistosomes. Trends Parasitol. 17: 168. [ Links ]
Van der Knaap, W.P. & E.S. Loker. 1990. Immune mechanisms in trematode-snail interactions. Parasitol. Today 6: 175-182. [ Links ]
Vasquez, R.E. & J.T. Sullivan. 2001. Effect of miracidial dose on adoptively transferred resistance to Schistosoma mansoni in the snail intermediate host, Biomphalaria glabrata. J. Parasitol. 87: 460-462. [ Links ]
Yoshino, T.P. 1976. Encapsulation response of the marine prosobranch Cerithidea californica to natural infections of Renicola buchanani sporocysts (Trematoda: Renicolidae). Int. J. Parasitol. 6: 423-431. [ Links ]
Zahoor, Z., A.J. Davies, R.S. Kirk, D. Rollinson & A.J. Walker. 2010. Larval excretory-secretory products from the parasite Schistosoma mansoni modulate HSP70 protein expression in defence cells of its snail host, Biomphalaria glabrata. Cell Stress Chaperones 15: 639-650. [ Links ]
*Correspondencia:
Iman F. Abou-El-Naga: Medical Parasitology Department, Faculty of Medicine, Alexandria University, Egypt. imanabouelnaga@hotmail.com
Eman H. Radwan: Zoology Department, Faculty of Science, Damnhour University, Egypt. dr_emanhashem@yahoo.com
1. Medical Parasitology Department, Faculty of Medicine, Alexandria University, Egypt; imanabouelnaga@hotmail.com
2. Zoology Department, Faculty of Science, Damnhour University, Egypt; dr_emanhashem@yahoo.com
Received 18-VIII-2011. Corrected 08-I-2012. Accepted 07-II-2012.


{kind=link}
{kind=link}
{kind=link}
{kind=link}