










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.60 n.3 San José Sep. 2012
A new species of Urocopitid land snail from Haiti and a discussion of the genus Autocoptis (Gastropoda: Urocoptidae)
Fred G. Thompson1*
*Dirección para correspondencia
Abstract
Autocoptis paulsoni n. sp. is described from Haiti. it is characterized by its large size, its cylindricaltapered shape, its fine costate sculpture on the teleoconch, a distinct but weak circum basal keel and its abbreviate conical juvenile shell. it is most similar to Autocoptis gruneri (Dunker 1844), which is redescribed, and its distribution is reviewed. The taxonomic status of the genus Autocoptis Pilsbry 1902 and its subgenus Urocoptola Clench, 1 935 are reviewed. The genus is endemic to Hispaniola and satellite islands.
Key words: Gastropoda, Urocoptidae, Autocoptis, Urocoptola, land snail, Hispaniola, Haiti.
Resumen
Para Haití se describe Autocoptis paulsoni n. sp. Esta especie se caracteriza por su gran tamaño, su forma cilíndrica-cónica, teleoconcha en forma de costillas, una clara pero débil quilla circumbasal y concha juvenil cónica y abreviada. Es muy similar a Autocoptis gruneri (Dunker, 1844), que se describe de nuevo y su distribución se revisa. También se presenta el estado taxonómico del género Autocoptis y del subgénero Urocoptola. El género es endémico de La Española y las islas satélites.
Palabras clave: Gastropoda, Urocoptidae, Autocoptis, Urocoptola, caracol terrestre, La Española, Haití.
The Urocoptidae of Hispaniola remain poorly known. Most species have received little notice in the literature since their original descriptions, except for repeated citations of previous information. Autocoptis Pilsbry (1903) is one such genus. it occurs exclusively on Hispaniola and satellite islands. Clench (1966) proposed the subgenus Urocoptola for those species of Autocoptis that do not have a crenulated suture as opposed to other species of Autocoptis, in which the suture is crenulated by the upper ends of the costate sculpture.
Pilsbry (1903) monographed Autocoptis when he named the genus. Later, Pilsbry (1933), and Pilsbry (1938) described four additional species, and Pilsbry & Olsson (1954) described a Miocene species that was questionably referred to Autocoptis. Bartsch (1932) described a new species from the off-shore Beata island. Clench (1935) described three new species and two new subspecies, and established the subgenus Urocoptola. Later, Clench (1966) described three additional species of Urocoptola. Thompson & Franz (1976) described three new species of Autocoptis. Richardson 1991, without discussion, treated Urocoptola as a synonym of the Jamaican genus Urocoptis.
Twenty-six species, 9 subspecies and 3 species of uncertain status currently are placed in the genus Autocoptis (Appendix 1).
In 1962 Dennis R. Paulson visited Haiti for the purpose of collecting natural history specimens, including mollusks. Paulson gave the molluscan specimens to the author. Among several novelties that he found is a new species of Autocoptis. At the time I prepared a description of the new species, completion of a manuscript was postponed because of other priorities.
I described the new Autocoptis to complement recent investigations of urocoptoid families from Mexico and Central America (Thompson 1968, Thompson 1976, Thompson & Correa 1991, Thompson in press). i am pleased to name the new species after its discoverer, Dennis R. Paulson.
Material and methods
This study is based on 31 dry specimens and two preserved specimens of the new species. Live specimens were preserved in the field in 75% ethanol. Dissections were made in 75% ethanol. Shell measurements are given in mm and are standard for elongate-cylindrical shells. The length of the shell is measured from the tip of the apical plug to the base of the peristome. The width is the width of the spire perpendicular to the shell length but does not include the peristome. The columella is viewed by filing open the shell to expose internal structure. Anatomical drawings were made with the aid of a Bausch & Lomb dissecting microscope and a camera lucida.
Results
Autocoptis (Urocoptola) paulsoni new species Figs. 1-6
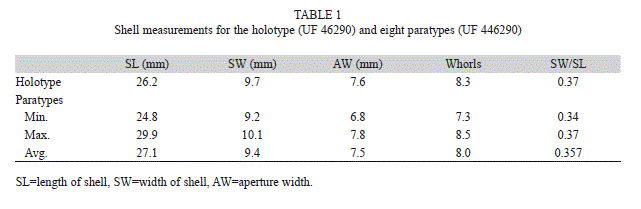
Diagnosis: A species related to Autocoptis (Urocoptola) gruneri (Dunker 1863), because of size, its sculpture, coloration, and a circumumbilical keel on the base of the last whorl. it is distinguished from A. gruneri by its cylindrical-tapered shape, by its narrower juvenile shell, by having a lusterless white shell with more numerous fine, flat, thread-riblets, by having a weaker umbilical keel, and by the following measurements; standard length, 24.8-2; standard width, 9.2-10.3mm; aperture width, 6.8-7.8mm; 7.3-8.5 whorls retained in decollated adult shells.
Shell (Figs. 1-6): Mature shells are lusterless white, are up to nearly 30mm in standard length, are moderately stout, being 0.34-0.37 times as wide as long, and are decollated with 7.3-8.5 whorls remaining below the apical plug (vestiges of earlier whorls in the holotype and paratypes were removed to expose the plug). The shape is cylindrical-tapered with the tapered portion of the upper spire consisting of 4-6 whorls which regularly increase in size. The first whorl at the point of decollation is 4.5-5.0mm wide. initially the juvenile shell is weakly concave or straight sided in outline (Fig. 5). The whorls nearly uniformly increase in size. The rate of growth of the juvenile shell indicates that 5-7, but usually 6 whorls are lost above the point of decollation at the apical plug (Fig. 4). The apical plug is elongate and laterally flattened as a tongue-like extension upward along the columella. The external color is dull-white. The base of the last whorl is tinged reddish due to the internal coloration. The peristome is white, fading into the rustcolored aperture deeper in. The septum of the first remaining whorl is pinkish. Below the first septum the interior of the whorls are white down to the penultimate whorl. internally the last whorl is tinged pink to dull red due to one band that begins along the suture, and another band that begins on the columella. The bands spread to color the columella and the last whorl internally dark rust-colored or red. The first 1.75 whorls of the protoconch are smooth, after which very fine riblets develop into distinct uniform riblets by the end of 2.5 whorls. The teleoconch whorls are slightly convex and shiny and are crossed by obsolescent, oblique, flat, thread-riblets. The penultimate whorl has 57-85 riblets. On the cylindric part of the shell the riblets increase in width on successive whorls where they are flat and close, and the interspaces are indicated only by incised grooves. The riblets become more pronounced on the periphery and base of the last whorl, and again become very fine in the umbilical chink. The suture is slightly impressed and is not crenulated by the upper ends of the riblets on the last whorl or two. The base of the last whorl has a poorly developed circum-umbilical keel (Fig. 3). The umbilicus is imperforate. The aperture is circular in shape. it is prosocline at an angle of about 22-25º to the shell axis (Fig. 2), so that in a frontal view it appears wider than high (Fig. 1). The peristome is nearly uniformly reflected along the columellar, basal and palatal margins of the aperture, and is attached to the preceding whorl across the parietal wall.
The columella is solid, straight and slender, but increases in diameter in the last three whorls, and is off-set in the last whorl (Fig. 6). A low, medium-placed columellar lamella is present in the last two whorls, but it does not enter into the aperture.
Measurements of the holotype (UF 46290) and eight paratypes (UF 446290) are presented in table 1.
Anatomy (Figs. 12-14): The sole is unipartite. The margin is bordered by a row of high pilaster-like folds. The dorsal surface of the foot is pebbled. The posterior-dorsal margin is rounded.
The salivary glands (not figured) are about 5mm long, elongate-ovate in shape and are loosely connected above the esophagus. The salivary ducts are long and slender.
Retractor muscle system (Figs. 13, 14): The columellar retractor muscle (Fig. 13) is large and stout, and attaches to the columella at about two whorls into the shell. The retractor muscle gives rise to three branches from about the same point, the dorsally located pharyngeal retractor and the two laterally placed pedal retractors. The pharyngeal retractor divides into three short branches that insert at the apex of the pharynx (Fig. 14). An elongate muscle slip originates on the dorsal surface of the pharyngeal retractor and inserts at the end of the odontophore sac. The right pedal retractor passes through the genital atrium, attaches at its midpoint to the apex of the penis and continues forward to the foot. The right ocular retractor originates on the outer margin of the right pedal retractor. The left ocular retractor originates on the outer margin of the left pedal retractor.
Reproductive system (Fig. 12): The genital atrium is short, about 0.6mm long. The penis is large and club-shaped, 7.5mm long. A slip of muscular tissue connects the apex of the penis to the vagina. The penis bears a large internal fold. An epiphallus is absent. The vas deferens is simple, extending from the apex of the penis as a narrow duct, and increases in size near the prostate where it is imbedded at the base of the oviduct. The albumen gland is large, reniform in shape and is 9mm long. The ovotestis consists of three lobes; each lobe consists of about 10-20 elongate digitiform lobules. The hermaphroditic duct is about 11mm long, slender and simple near the ovotestis, and becomes thicker and convoluted midway to the albumen gland where it enters near the junction with the prostate-oviduct. The prostate-oviduct is long, simple and tapering at the lower end. The free oviduct is 7mm long, slender and tapered. The spermathecal duct lacks an appendix, is about 11mm long, slender, and enters the free oviduct shortly above the atrium to form a short vagina.
Type locality: Haiti, Dept. de l’Oest, 3km Southeast of Trou Forban, (18.92º N - 72.63º W), ca. 700m alt. Holotype UF 46290; collected by Dennis R. Paulson; August 14, 1962. Paratypes: UF 446290 (20 adults), UF 446789 (10 juveniles); same data as the holotype.
Distribution: Haiti; known only from the type locality.
A diagnosis of Autocoptis gruneri is given for comparison with Autocoptis paulsoni.
Cylindrella gruneri Dunker 1844,in, Philippi 1842-1845; Abbildungen und Beschreibungen neuer oder wennig gekannter Conchylien unter Mithülfe mehrer deutscher Conchyliologen, no. 1: 185; pl. 1, fig. 20- Pfeiffer 1848; Monographia heliorum viventium 2: 370.- Crosse 1891; Journal de Conchyliologie 29: 135.
Urocoptis (Autocoptis) gruneri (Dunker). Pilsbry 1903; Manual of Conchology, 15: 157; pl. 39, Figs. 16-19.
Urocoptis (Urocoptola) gruneri (Dunker). Clench 1935; Proc. Bost. Soc. Nat. Hist., 41: 3.
Diagnosis (Figs. 7-11): This is a large robust species with a conical-ovate shell. it is about 27-30mm long, and is about 0.37-0.42 times as wide as long. Adults retain about 7-8 whorls. The suture is moderately impressed. The rate of growth of juvenile shells indicates that about six whorls are lost above the point of decollation (Fig. 10a). The shell bears a large, wide aperture surrounded by a wide, thick, reflected peristome that may be free from (Fig. 7) or narrowly attached to (Fig. 8) the preceding whorl. The upper whorls are broadly conical and slightly concave in outline. The protoconch consists of two smooth whorls. The teleoconch bears fine, low, closely spaced thread-ribs which become stronger on the lower whorls. The base of the last whorl bears a strong circum-basal crest which is crossed by strong, well –defined ribs (Fig. 9). The umbilicus is imperforate. The shell is grayish-white with the last one or two whorls livid colored. The interior of the aperture and the last two whorls are dark rust-colored. The slender columella increases in diameter to the last whorl (Fig. 11). it is solid and is slightly twisted in each whorl. in the last whorls the columella is off-set. in the last two whorls it bears a low, medium-placed, rounded lamella that ends in the last half-whorl behind the aperture.
There appears to be no significant difference between populations from ile Gonave and from the mainland near St. Marc.
Type locality: Haiti, Port-au-Prince. The type locality is questionable. it has not been reported from near Port-au-Prince since it was described in 1844. it has not been found near there during extensive searches by John B. Henderson in 1900, by Dennis R. Paulson in 1962, nor by the author in 1979, 1980 or 1984. Henderson (1901) did not include a list of species he found near Port-au-Prince, but his urocoptid material from his collection was reported by Pilsbry (1903).
Distribution: This species is common on the Southeast side of ile Gonave, and on the Haitian mainland near St. Marc at elevations of up to 350m. Early records for ile Gonave and St. Mark are given by Pilsbry (1903).
Specimens examined. HAITI, Ile Gonave: (UF 50979. 5 spec., UF 179190. 7 spec.); Anse-A-Galets (UF 179187. 58 spec., UF 179188. 69 spec., UF 280107 1 spec.); Trou Zombie, Anse-A-Galets (UF 290098. 6 spec.); 2km SE of Anse-A-Gelets, 300m alt. (UF 23078. 23 spec.); 3km SSW of Anse-A-Galets, 200m alt. (UF 23116. 2 spec.). 2 spec.; 5km WNW of Anse-A-Galets (UF 23091. 7 spec.); 4km S of Anse-A-Galets (UF 23120. 2 spec.); Anse-A-Galets (UF 290098. 6 spec.); 3km S Etroits, 100m alt. (UF 179186. 48 spec.). Grand Source, 340m alt. (UF 179189. 1 spec.); Dept. Artibonite: St. Marc (UF 179191. 10 spec.); 5km NNE of St. Marc (UF 33087. 8 spec.); 10km ESE St. Marc (UF 32721. 21 spec.); 7km SW of St. Marc (UF 32954. 24 spec., UF 32955. 18 spec.); 7km WSW St. Marc, Amaniy-Les Bains (UF 32665. 22 spec., UF 32666,. 19 spec., UF 32667. 8 spec.).
Discussion
Autocoptis (Urocoptola) paulsoni n. sp. (Figs. 1-6) is most similar to Autocoptis (Urocoptola) gruneri (Dunker, 1844), the type species of Urocoptola. it is placed in the subgenus because the axial sculpture does not crenulate the suture, unlike typical Autocoptis, which has a strongly crenulated suture. Autocoptis paulsoni is similar to A. gruneri in size, sculpture, color and by having a prominent circum-umbilical crest.
Autocoptis paulsoni differs from A. gruneri by the following combination of characters. The shell is cylindrical-conical in shape. it is narrower and not as robust. The juvenile shell is narrower. The sculpture on the teleoconch is weaker, and the suture between the whorls is weaker. The basal crest on the last whorl is weaker, and is crossed by finer, more closely spaced ribs. The aperture is proportionally smaller. The peristome is adnate across the parietal wall.
Autocoptis gruneri has a robust, conicalovate shell with tumid lower whorls. The juvenile shell is more broadly conical. The teleoconch sculpture is stronger and the suture is more deeply impressed. The prominent basal crest is crossed by stronger, more widely spaced ribs. The aperture is proportionally larger. The peristome is thicker and is adnate to or free from the previous whorl (the attachment of the peristome to the previous whorl is individually variable, unlike other species of Autocoptis).
Anatomical comparisons are limited to four recently described species from the Dominican Republic. The penis retractor muscle is attached to the mid-point of the right pedal retractor, as it does in Autocoptis eburate Thompson & Franz 1976, A argiphrix Thompson & Franz 1976, A. stibe Thompson & Franz 1976, Allocoptis nebrius Thompson & Fanz 1976 and in Jamaican Urocoptinae (Paul 1983). The spermathecal duct lacks an appendix, as in all species of Urocoptidae (Pilsbry 1902, Paul 1980).
The genus Autocoptis is characterized as follows. The shell is cylindrical and rather large, up to about 30mm long. The columella is straight, but may be moderately twisted in the last whorl or two. in the penultimate and last whorl a columellar lamella may be present which may extend downward to the throat of the aperture. The shell is decollated in adults. About six juvenile whorls are lost above the point of decollation. Normally the adult shell retains fragmented portions of the earlier whorls. Usually, the apical plug is concealed behind fragments of earlier whorls. The juvenile shell is broadly conical and relatively short compared to other genera. The first two or three protoconch whorls are smooth. The following whorls are sculptured with prosocline riblets. The teleoconch is costate or smooth. The suture may be crenulated or smooth. The central tooth of the radula is relatively broad and is almost as wide as the lateral teeth.
The genus Autocoptis differs from Urocoptis, a Jamaican genus, by characteristics of the juvenile shell and the radula. in Autocoptis the juvenile sell is abbreviated and broadly conical, and the central tooth is about as wide as the lateral teeth (Pilsbry 1902, Thompson & Franz 1976). in Urocoptis the juvenile shell is attenuated and slender, and the central tooth is narrow, being about half a wide as the lateral teeth.
The available data, scant as it may be, support recognition of Autocoptis as a genus distinct from Urocoptis, and of Urocoptola as a subgenus distinct from typical Autocoptis.
Acknowledgments
For assistance in the field relating to this study i thank Ronald i. Crombie (formerly USNM), Roger W. Portell (Florida Museum of Natural History) and Dennis R. Paulson (Seattle Washington).
References
Bartsch, P. 1932. A newly discovered West indian Mollusk faunula. Proc. U. S. Nat. Mus. 70: 1-13. [ Links ]
Clench, W.J. 1935. Some new Urocoptidae from Hispaniola. Proc. Bost. Soc. Nat. Hist. 41: 1-12. [ Links ]
Clench, W.J. 1966. Notes and descriptions of new Urocoptidae from Cuba and Hispaniola (Mollusca: Pulmonata). Breviora 245: 1-14. [ Links ]
Henderson, J.B. 1901. Collecting in Haiti. Nautilus 15: 13-16. [ Links ]
Paul, C.R.C. 1983. The comparative anatomy and systematics of Jamaican Urocoptidae (Mollusca: Pulmonata). Jour. Linn. Soc. 78: 1-28. [ Links ]
Pilsbry, H.A.1902. Manual of Conchology. Academy of Natural Sciences of Philadelphia, Philadelphia, Pennsylvania, USA. [ Links ]
Pilsbry, H.A. 1903. Manual of Conchology. Philadelphia Academy of Natural Sciences of Philadelphia, Philadelphia, Pennsylvania, USA. [ Links ]
Pilsbry, H.A. 1933. Santo Domingo land mollusks collected by Daniel C. Pease, 1932 and by A.A. Olsson, 1916. Proc. Acad. Nat. Sci. Phila. 85: 121-162. [ Links ]
Pilsbry, H.A. 1938. New species of Urocoptis and Euglandina. Nautilus 52: 15-16. [ Links ]
Pilsbry, H.A. & A.A. Olsson. 1954. Miocene land shell fossils from the Dominican Republic. Not. Nat. 266: 1-4. [ Links ]
Richardson, C.L. 1991. Urocoptidae: Catalog of species. Tryonia 22: 1-245. [ Links ]
Thompson, F.G. 1968. Some Mexican land snails of the family Urocoptidae. Bull. Fla. St. Mus. 12: 125-183. [ Links ]
Thompson, F.G. 1976. The genus Epirobia in Chiapas, Mexico. Nautilus 90: 41-46. [ Links ]
Thompson, F.G. & A. Correa. 1991. Mexican land snails of the genus Hendersoniella. Bull. Fla. Mus. Nat. Hist. 36: 1-23. [ Links ]
Thompson, F.G. & R. Franz. 1976. Some urocoptid land snails from Hispaniola. Rev. Biol. Trop. 24: 7-33. [ Links ]
*Correspondencia:
Fred G. Thompson: Florida Museum of Natural History, University of Florida, Gainesville, Florida USA 32611. fgt@flmnh.ufl.edu
1. Florida Museum of Natural History, University of Florida, Gainesville, Florida USA 32611; fgt@flmnh.ufl.edu
Received 11-vii-2011. Corrected 04-i-2012. Accepted 03-ii-2012.


{kind=link}
{kind=link}
{kind=link}