










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.60 n.3 San José Sep. 2012
Faunistic analysis of the caridean shrimps inhabiting seagrasses along the NW coast of the Gulf of Mexico and Caribbean Sea
Everardo Barba Macías1*
*Dirección para correspondencia
Abstract
Seagrass meadows are highly productive and ecologically important habitats in estuaries and coastal lagoons, and contain a variety of faunal communities, from which the caridean shrimps are a dominant component. The purpose of this work was to analyze the environmental parameters of water and sediments, with the biological components in seagrass epifaunal communities, from the Western Gulf of Mexico and the Caribbean Sea. For this, density and diversity of caridean shrimps were analyzed and correlated with environmental parameters and seagrass biomass, and zoogeographic affinities were determined. The spatial distribution of caridean shrimps was recorded for 12 localities with Halodule wrightii and Thalassia testudinum monospecific seagrass meadows. A total of 72 158 individuals of 16 taxa were collected. Among results, the Hippolytidae resulted the most abundant group (92.3%) with eight species, and was followed by Palaemonidae with 7.6% of the abundance and seven species, and the Alpheidae with only one genus. From the total of collected carideans, a 37.3% was found in H. wrightii and 62.7% in T. testudinum. The dominant species were Hippolyte zostericola (12.39ind./m2), Tozeuma carolinense (9.5ind./m2), Thor dobkini (4.84ind./m2) and Palaemonetes vulgaris (4.87ind./m2). The zoogeographic distribution of the carideans presented two groups: species of the Virginian-Carolinean province representing its Southern limit (43.75%) and species of the Brazilian-Caribbean province representing its Northern limit (56.25%). The species H. zostericola, T. carolinense, P. vulgaris, P. pugio and P. intermedius are widely distributed along the Western Atlantic coast. This study has base line information for seagrass habitats, the community of epifaunal carideans and their ecological affinities, previous to the oil spill in the Gulf of Mexico.
Key words: caridean shrimp, Western Atlantic coast, tropical, seagrass meadows, baseline.
Resumen
Las praderas de pastos marinos son hábitats altamente productivos y ecológicamente importantes a lo largo de las costas y estuarios, en estas se encuentra una gran variedad de comunidades faunísticas, donde los camarones carideos son un componente dominante por su densidad. Los parámetros ambientales del agua y sedimento y los componentes biológicos de biomasa de pastos marinos y comunidad epifaunística fueron recolectados a lo largo del occidente del Golfo de México y Mar Caribe. La densidady diversidad de los camarones carideos fueron analizadas y correlacionadas con los parámetros ambientales y biomasa de pastos, también se determinaron las afinidades zoogeográficas de las especies. La distribución espacial de los camarones carideos fue registrada en 12 localidades con praderas monoespecíficas de los pastos Halodule wrightii y Thalassia testudinum. Un total de 72 158 individuos pertenecientes a 16 taxa fueron recolectados. La familia Hippolytidae incluyó a ocho especies y representó el 92.3% de la abundancia, la familia Palaemonidae comprendió a siete especies y el 7.6%, y la familia Alpheidae estuvo representada por un solo género. Del total de carideos recolectados, el 37.3% se capturó en H. wrightii y el 62.7% en T. testudinum. Las especies dominantes fueron Hippolyte zostericola (12.39ind./m2), Tozeuma carolinense (9.5ind./m2), Thor dobkini (4.84ind./m2) y Palaemonetes vulgaris (4.87ind./m2). La composición zoogeográfica de los carideos estudiados estuvo representada por dos grupos: el primero constituido por especies afín a la provincia Virginiana- Carolineana en su límite más sureño (43.75%), y el segundo por especies de la provincia Brasileña-Caribeña con su límite más norteño (56.25%). Las especies H. zostericola, T. carolinense, P. vulgaris, P. pugio y P. intermedius tienen una amplia distribución a lo largo de la costa noreste Atlántica.
Este estudio constituye la línea base de información sobre los hábitats de pastos marinos, la comunidad de camarones carideos epifaunales y sus afinidades ecológicas de recolectas previas al derrame de petróleo en el noreste del Golfo de México.
Palabras clave: camarones carideos, costa noreste Atlántica, tropical, praderas de pastos marinos, estudio base.
The salinity gradient, habitat complexity and heterogeneity are among the most important ecological factors that regulate the distribution of estuarine fauna (McLusky 1989, Gilmore 1995, Sánchez et al. 1996, Barba 1999, Sheridan & Minello 2003), and interactions such as predation, competition and food and space availability determine the presence or absence of species (Bulger et al. 1993). Estuarine decapod crustaceans, particularly the infraorder Caridea, are a numerically important component of the communities of invertebratesassociated with submerged aquatic vegetation (SAV) (Bauer 1985, Sánchez et al. 1996, Sheridan & Minello 2003, Barba et al. 2005). Several studies have indicated the importance of crustaceans as a primary food source for fish (Greening & Livingston 1982, Llansó et al. 1998), as mesoherbivores controlling periphytic algae (Kitting 1984, Jernakoff 1996, Barba et al. 2000), in reducing seagrass mortality as a vehicle of energy transference from epiphytes through caridean biomass to upper trophic levels (Llansó et al. 1998, Mazzella et al. 1992), in recycling nutrients through the fecal pellets of resident populations (Meyer et al. 1983) and as migratory populations (Thayer et al. 1984).
The distribution patterns of the caridean shrimp in the large estuarine systems of the Mexican Gulf of Mexico, including Laguna Madre and Laguna de Términos, are related to the environmental heterogeneity that is determined mainly by the salinity and type of habitat, with 80% of the abundance and more than 55% of the species richness collected in habitats with SAV substrates (Barba et al. 2005). The Gulf of Mexico and Caribbean Sea have extensive areas with SAV substrates, where the most abundant species are: Thalassia testudinum Banks & Sol. ex K.D. Koenig, Syringodium filiforme Kütz and Halodule wrightii Aschers (de la Lanza & Tovilla 1986, Sánchez et al. 1996). Halophila engelmani Aschers and H. decipiens Ostenfel (van Tussenbroek 1995) are also present in the Caribbean Sea. These SAV areas present a complex spatial arrangement, compared with bare substrates that do not provide food and protection against predation (Heck & Orth 1980, Vose & Bell 1994, Llansó et al. 1998).
The possible impacts of diverse origins on the benthic communities of these areas, particularly the last oil spill in the Northern Gulf of Mexico, have not been evaluated. The purpose of this study is to describe the composition, density and diversity of caridean shrimp in coastal and estuarine SAV substrates along a spatial gradient in the Gulf of Mexico and Caribbean Sea, as baseline study of anthropogenic disturbances such as oil spill effects on benthic communities.
Materials and methods
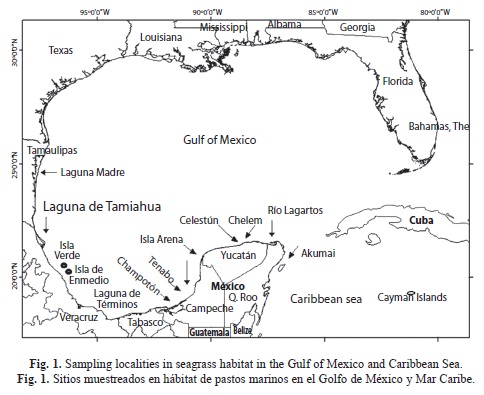
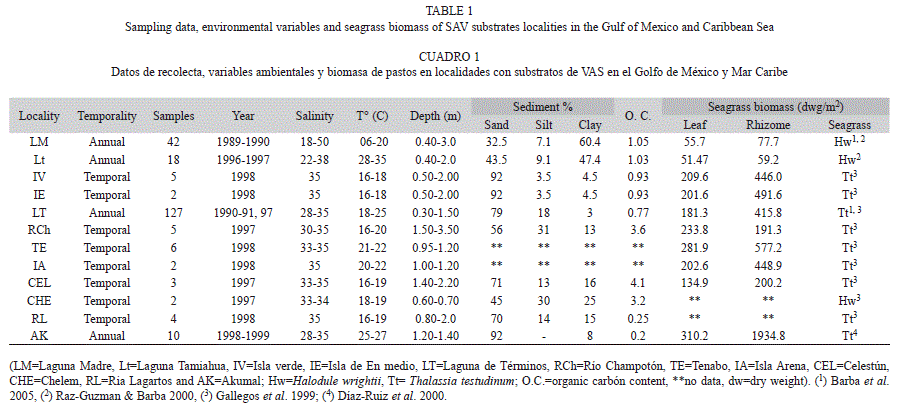
Environmental characterization and clustering: 12 localities were selected for sampling in coastal and estuarine seagrass meadows in the Gulf of Mexico and Caribbean Sea: Laguna Madre (LM), Laguna de Tamiahua (Lt), Isla verde (IV), Isla de Enmedio (IE), Laguna de Términos (LT), Río Champotón (RCh), Tenabo (TE), Isla Arena (IA), Celestún (CEL), Chelem (CHE), Ría Lagartos (RL) and Akumal (AK) (Fig. 1). Water parameters of temperature, salinity and depth were registered in each locality. Sediment size was analyzed following Folk (1974) technique; sediments were dried in oven at 40ºC during 48hr and sieved to 0.25mm mesh, according to Gaudette et al. (1974) to determine organic carbon content, from duplicated samples and two blanks. Sand was quantified as total fraction.
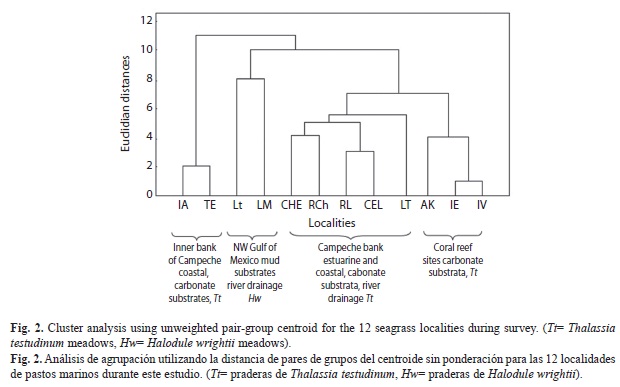
Sampling years during 1996-1997 and 1998 were occasional and those from Laguna Madre, Laguna Tamiahua, Laguna de Términos and in the Caribbean were conducted annually. The environmental parameters of water temperature and salinity, habitat type, percentage of gravel, sand, mud and organic carbon content, together with the species, were used to define affinities and to group species and localities in a cluster analysis using unweighted pairgroup centroid (UPGW) and Euclidian distances (STATISTICA 2000).
Seagrass biomass: Submerged aquatic vegetation (SAV) and sediments were sampled with a metallic core of 20.5cm of diameter and 20cm of length (sampling area 0.03m2). Seagrasses were separated into leaves, rhizomes and roots, and oven dried at 105°C for 12hr to determine biomass as dry weight (Fourqurean et al. 2001).
Faunistic composition and density: Caridean shrimp were collected with a Colman-Seagrove net (sampling area 38.5m2) (Eleftheriou & Holme 1985) in Laguna de Términos, and with a beam trawl net (sampling area 50m2) (Renfro 1962) in the other localities. Sampling was carried out during daylight in monospecific Halodule wrigthii and Thalassia testudinum meadows. The selectivity of these two sampling nets has been found to be similar (Álvarez et al. 1996), and comparing samples from a quantitative viewpoint is considered valid. Caridean shrimp species were identified following the taxonomic criteria of Holthuis (1952), Chace (1972) and Williams (1984), and arranged following De Grave et al. (2009) criteria. Density was determined as number of individuals per m2.
The density of each species was transformed to ln (x+1), the dominance of the caridean shrimp was determined considering the density (ind./m2) and frequency of eachspecies, using the non-parametric association of Olmstead-Tukey (Sokal & Rohlf 1981). Ordinations were conducted using principal component analysis (PCA) to explore spatial variability of carideans shrimps in localities (PRIMER 6 & ERMANOVA).
Diversity: Species richness and diversity were determined by computing the species richness index (D), the Shannon diversity index (H) and the evenness index (E) were used to describe community (Krebs 1989, Ramírez 1999, McCune & Grace 2002). The values obtained for these indices in the different samples were automatically significant, and no statistical analysis was required (Magurran 1988).
Zoogeographic affinity: Zoogeographic distributions were arranged in agreement with the American Zoogeographic provinces proposed by Boschi (2000) for decapod crustaceans.
Results
Environmental characterization and clustering: Four groups of localities were determined using salinity, temperature, depth, sediment composition, organic carbon content and seagrass biomass data. The first group included LM and Lt, with a high salinity, a marked variation in temperature, a high content of fine sediments and organic carbon and intermediate values of H. wrightii biomass. The second group included CHE, RCH, RL, CEL and LT, all located in the transitional region of terrigenous-carbonate substrates, in euhaline environments, with high temperature, 40-80% of sand, a high content of organic carbon and intermediate to high values of T. testudinum biomass. The third group IA and TE, corresponded with euhaline conditions, high temperature values, and a high T. testudinum biomass. The fourth group corresponded to the euhaline coral reef sites of AK, IE and IV with 90% sand, carbonate substrate, a low organic content and a high T. testudinum biomass (Table 1, Fig. 2).
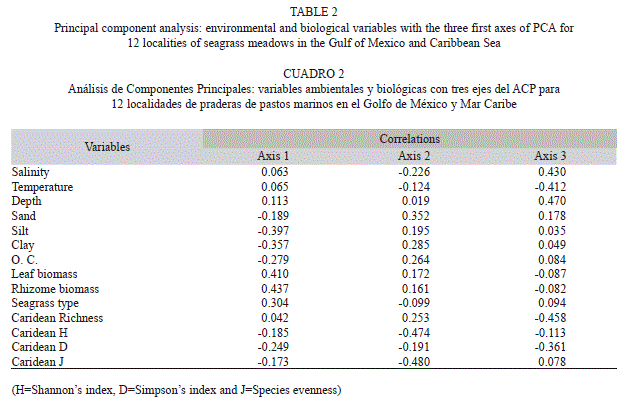
Principal component analysis indicated that the first three axes explained 73.1% of the cumulative spatial variability in this study (Fig. 3), and accounted for 30.3, 23.4 and 19.4% of variability, respectively. From the correlations (Table 2), it is concluded that the component axes representing environmental variables were explained by multivariate space determined by axes 1 and 2 (Fig. 3), because axis 1 was five times, and axes 2 three times more strongly correlated with the environmental and biological variables. The first principal component was strongly and negatively correlated with lime and clay, sites with solid substrates and trongly and positively correlated with seagrass biomass (leaf and rhizome). This group included sites LM, Ak, RCh, IV, IE, LT, TE and IA. The second principal component was strongly correlated with diversity indexes of carideans, grouping sites CEL, CHE, Lt and RL (Fig. 3).
Seagrass biomass: Monospecific meadows of H. wrightii dominated in the LM and Lt systems, in subtropical eu-hypersaline conditions and terrigenous sediments. Thegreatest values of leaf (55.73g/m2) and rhizome (77.67g/m2) biomass were recorded for LM. The other sites were characterized by T. testudinum, with maximum leaf and rhizome biomass values recorded for AK (310.2 and 1 934.8g/m2) and the minimum values for CEL (134.9 and 200.2g/m2) (Table 1).
Faunistic composition and density: A total of 72 158 individuals belonging to 16 taxa of caridean shrimps were collected from 12 localities in the Gulf of Mexico and Caribbean Sea: 8 species of the Hippolytidae, 7 species of Palaemonidae and 1 species of Alpheidae (Table 3). A total of 37.3% of the global caridean density occurred in the H. wrightii habitat in LM, Lt and CHE, and 62.7% in the T. testudinum habitat in the other sampling sites (Table 3).
The 92.3% of the total density was represented by the Hippolytidae family, and was followed by Palaemonidae (7.6%). The greatest total densities were recorded for the hippolytids H. zostericola (12.39ind./m2), T. carolinense (9.5ind./m2), T. dobkini (4.84ind./m2) and thepalaemonid grass shrimp Palaemonetes vulgaris (4.87ind./m2) (Table 3).
On the spatial scale, the highest density values were recorded for RL (18.82ind./ m2), CHE (13.45ind./m2) and Lt (10ind./m2), and corresponded to the collection of Thor dobkini, Hippolyte zostericola and Tozeuma carolinense, respectively. The lowest density values were recorded for IE and LM with 0.39 and 0.48ind./m2, respectively (Table 3).
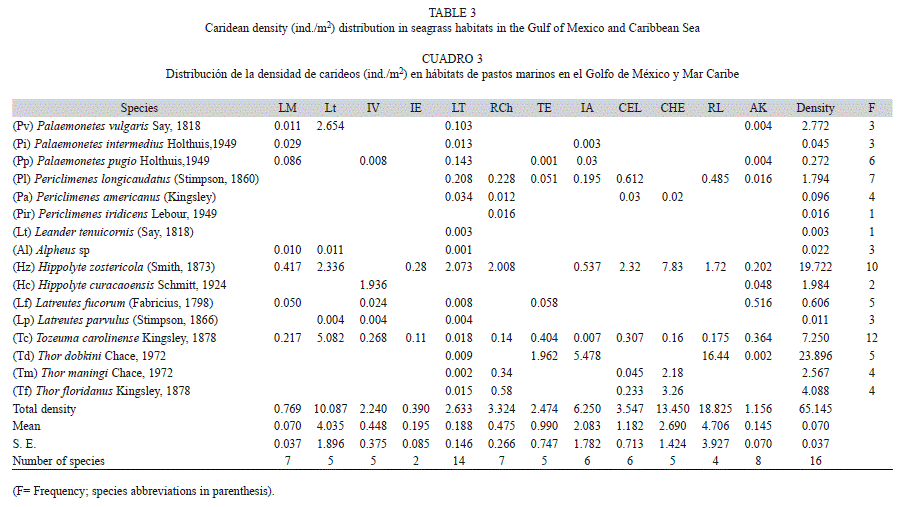
The dominant species H. zostericola represented more than 50% of the total density per sampling site in LM, IE, LT, RCh, CEL and CHE, whereas T. carolinense in Lt and T. dobkini in TE, IA and RL represented 80%. Tozeuma carolinense was collected in the 12 sampling sites, H. zostericola in ten sites (except Isla Verde and Tenabo), ericlimenes longicaudatus in seven, Palaemonetes pugio in six, Periclimenes iridiscens and Leander tenuicornis only in CHA and LT, and Hippolyte curaςaoensis in the IV and AK reef habitats (Table 3). The dominant species according to abundance were: T. carolinense, H. zostericola, T. dobkini and P. longicaudatus, the frequent species were Latreutes fucorum and P. pugio, and the abundant species were: P. vulgaris, T. floridanus and T. maningi (Fig. 4).
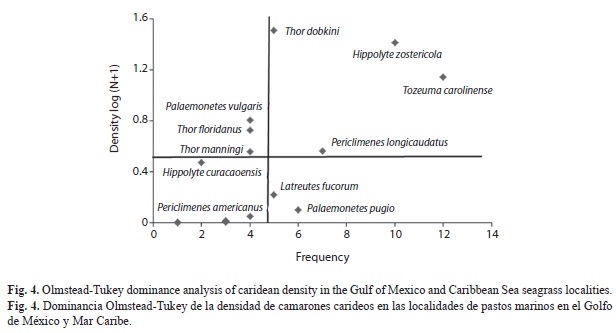
Based on the distribution of the density data, four groups were formed: 1) T. carolinense and H. zostericola, the dominant species associated with poly-euhaline conditions throughout the spatial range; 2) T. dobkini and P. longicaudatus, dominant species recorded from LT southward; 3) Palaemonetes intemedius, T. floridanus and T. maningi, abundant species with an intermediate distribution; 4) the frequent and rare species (Fig. 5).
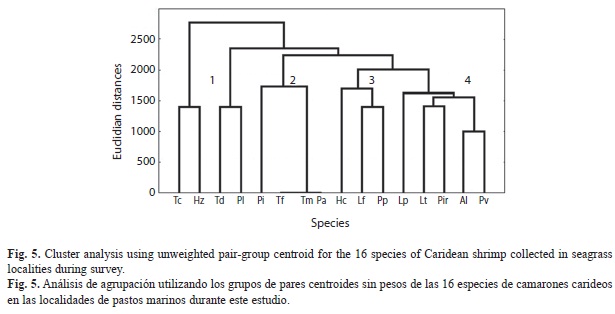
Diversity: The maximum number of species per site was 14 in LT and the minimum was two species in IE. The maximum Simpson index values were those of IV, IA and RL (5-6 species). The greatest Simpson indices indicate that a community is dominated by one or two species. These were H. curacaoensis in IV and T. dobkini in IA and RL, where they represented more than 85% of the total abundance (Fig. 6).
The hannon-Weinner index considers the richness and distribution of abundance of each species. The greatest values were recorded for LM, RCh and AK, together with the greatest richness. Evenness measures the abundance of the dominant species. The greatest values were those of IE, and the lowest were those of IVand IA, where the most abundant species were: H. curacaoensis and T. dobkini (Fig. 6).
Zoogeographic affinity: The zoogeographic distribution of the caridean shrimp presents an affinity with the Virginian (6.5%), Carolinean (75%), Texan (81.2%), Caribbean (56%) and Brazilian (12.5%) provinces.
Discussion
The seven species of Palaemonidae and eight species of Hippolytidae collected during this study, account the 70% of palemonids recorded for the Caribbean coast (Román-Martínez 2010) and 57.1% of hippolytids recorded by Markham et al. (1990) in the Caribbean coast. The low representation of the Alpheidae family was thought to be due to sampling exclusion. They are known for their infaunal burrowing habits (Bauer 1985). This low representation from using selective sampling gear and sampling in monospecific SAV areas, thus excluding other habitats such as coral and rocky substrates, Sargassum mats, sandy and muddy bare substrates, calcareous algae and anemones (Román- Martínez 2010).
Most of the carideans were widely distributed and associated with SAV substrates, where the conditions of salinity, temperature, type of habitat and predation regulate the abundance and diversity of the group (Barba et al. 2005). Briggs (1974) argued that the widespread distribution of many species is mainly associated with temperature, which controls distribution in the ocean. This study area is under the influence of subtropical and tropical water masses, which explains the convergence of different types of fauna.
The dominant species Tozeuma carolinense is distributed in SAV in subtropical and tropical euhaline waters and, in the case of Hippolyte zostericola, it is absent in Laguna de Alvarado in response to its low tolerance to mesohaline waters (Sánchez et al. 1996). These are numerically dominant epifaunal species associated with SAV throughout the Western Atlantic, both in temperate (Zostera and Halodule) and tropical (Thalassia, Halodule and Syringodium) seagrass beds, as well as in drift macroalgae (Dugan 1982, Greening & Livingston 1982, Livingston 1984, Sánchez et al. 1996). The distribution of the arrow shrimp spans from Massachusetts to Curaςao, including Yucatán and Panama (Chace 1972, Román-Contreras & Martínez-Mayén 2009). It is abundant in Halodule beds in Lower Laguna Madre, Texas (Sheridan & Minello 2003), and in this study it was abundant in LM and Lt in Halodule meadows. The hippolytids L. fucorum and L. parvulus are distributed in euhaline waters, and are associated with T. testudinum and red macroalgae. Both species have an amphi-Atlantic tropical distribution and are nocturnal (Bauer 1985). They occur in tropical marine ecosystemsin Puerto Rico and Panama, and have been recorded in low numbers in temperate estuaries (Greening & Livingston 1982). Another hippolytid, T. dobkini, was collected from SAV and red macroalgae in the NC-NE subsystem of Laguna de Términos in poly-euhaline waters. It is distributed widely in subtropical-tropical regions from North Carolina to Yucatán (Williams 1984, Román-Contreras & Martínez-Mayén 2009). Latreutes fucorum, L. parvulus and T. dobkini were collected with low density values. Both are nocturnal species, that hide from predators among macroalgae during the day (Greening & Livingston 1982, Bauer 1985).
Species of Palaemonetes, inhabit a variety of substrates including Zostera, Halodule, Thalassia, Syringodium and macroalgae, and have a wide distribution that responds to their ample physiological tolerance to salinity and temperature conditions and spans from the Northern Atlantic latitudes of Massachusetts and North Carolina to Yucatán, and from Bermuda to Trinidad and Curaςao (Chace 1972, Dugan 1982, Lewis 1984, Livingston 1984, Zupo & Nelson 1999, Sheridan & Minello 2003, Román-Contreras & Martínez-Mayén 2010). They are also found along the Pacific coast of Colombia (Wicksten 1989). In contrast, their abundance decreases and the dominance of P. pugio is favoured in Laguna de Alvarado, Veracruz, attributed to the oligomesohaline conditions (Sánchez et al. 1996, Barba 1999, Barba et al. 2005).
Palaemonetes pugio is the most studied caridean of the Western Atlantic where it is commonly found in SAV, smooth cordgrass Spartina alterniflora Loisel, soft substrates, logs and among mangrove roots (Posey & Hines 1991, Everett & Ruiz 1993, Cross & Stiven 1999). The three dagger-blade grass shrimp species of the genus Palaemonetes were placed by Barba et al. (2005) in the “widely distributed” and “not-associated with the habitat” distribution pattern. Of these three species, the brackish grass shrimp P. intermedius and the grass shrimp P. pugio are the more tolerant/resistant species to low salinities (Knwolton & Williams 1970). Palaemonetes pugio is most commonly foundin 10-20ppm, while the marsh grass shrimp P. vulgaris is common in the higher salinities of 15-40ppm (Williams 1984) and does not survive in less than 15ppm (Nagabhushanam 1961).
The coexistence of P. pugio and P. vulgaris in intermediate salinities responds to predator-prey interactions, competition or differences in behaviour, rather than to differences in physiological tolerance (Thorp & Hoss 1975). When both species coincide in oyster beds, P. vulgaris displaces P. pugio as a result of competition for shell substrates that supply refuge against fish predation and for food from the continual tidal influx of detritus (Thorp 1976). Consequently, when both species are exposed to fish predation, P. pugio suffers a greater mortality in spite of being more resistant to low salinities, poor flushing, a high organic content and anoxic conditions (Williams 1984). Palaemonetes intermedius is distributed from Massachusetts to Texas (Holthuis 1952, Sheridan & Minello 2003) and South to Quintana Roo (Chace 1972). The three palaemonid species are widely distributed with an affinity for the Virginian-Carolinean province, as well as a more Southern distribution range along the Yucatán Peninsula into the Caribbean province (Holthuis 1952, Williams 1984, Román-Contreras & Martínez-Mayén 2010).
Periclimenes longicaudatus and P. americanus are tropical euhaline species. They were collected in poly-euhaline waters and were associated with T. testudinum, H. wrightii, S. filiforme, macroalgae and carbonate substrates in the NC-NE subsystem of Laguna de Términos (Sánchez & Raz-Guzman, 1997, Barba et al. 2005). These two species are distributed from North Carolina to Southwestern Florida, and from the West Indies to São Paulo, Brazil (Williams 1984, Román-Contreras & Martínez-Mayén 2010).
Although, the structural elements of seagrass ecosystems have been reasonably well described, the basic functional relationships of the component species remain only partly understood (Livingston 1984). Seagrasses and their associated epiphytes form a habitat and provide an organic matter matrix for diverse assemblages of organisms (Heck & Orth 1980). Caridean shrimp are numerically dominantcomponents in vegetated substrates (Dugan 1982, Greening & Livingston 1982, Sánchez et al. 1996, Barba 1999), and are important links between primary production and the higher trophic levels (Llansó et al. 1998, Barba et al. 2000). The grass shrimp H. zostericola, T. carolinense and P. pugio are among the most abundant species in SAV habitats, and are also prey of fish associated with these habitats (Llansó et al. 1998, Cross & Stiven 1999).
Seagrasses, in general terms, appear to be relatively tolerant to many anthropogenic chemicals. Several species have been shown to be less sensitive to chemicals than other species of marine flora, corals and macroinvertebrates. In addition, some seagrasses bioaccumulate chemicals at lower concentrations (Lewis & Devereux 2009). Damage to seagrass communities from oil exposure includes acute mortality resulting from physical impacts and chemical toxicity; indirect mortality as the result of light loss, death of food sources, or the destruction or removal of habitat; destruction of sensitive juvenile fishes and invertebrates; and accumulation of potentially carcinogenic or mutagenic substances in the food chain (Zieman et al. 1984). Fauna and flora associated with seagrasses are also affected by oil. Infauna, nearly completely killed by oil exposure, gradually returned to abundances above prespill levels. However, only species with high reproductive potential or planktonic stages recovered quickly. The effects of oil on tropical fish have not been extensively studied (Peters et al. 1997).
Other human impacts are associated with increase of nutrients producing bad effects on the structure and functions of the seagrass ecosystem, as eutrophication leads directly to a large reproduction of plankton and epiphytic algae on seagrasses, thus reducing light flux of water body and dissolved oxygen contents due to decomposition of the algae. Increased sediment loads can smother seagrasses, change the redox potential of rhizome-penetrated sediments, or cause increased water column turbidity, robbing seagrasses of the high levels of incident light required for metabolic functions. Such high light requirements mean seagrassesare affected by even minor increases in turbidity, which is now recognized as the predominant threat to seagrass communities (Lewis & Devereux 2009).
The results obtained in this study show that caridean shrimps are resident and representative fauna of SAV substrates, and the salinity, temperature and type of habitat regulate the dominance of the species. Understanding the distribution and ecological affinities of the caridean shrimp as dominant epifaunal components in SAV substrates requires research on the trophic contribution and energy transfer of these communities, as well as on the impact of environmental phenomena such as the recent oil spill in the Northern Gulf of Mexico and also by human impacts as tourism activities, nutrient upload, increased of sediment load and turbidity as threats for seagrass habitats in the area. This study contributes with baseline information on the benthic communities and distributional patterns of caridean shrimps in seagrass habitats in the Gulf of Mexico and Caribbean Sea.
Acknowledgments
Thanks are extended for the information obtained through the projects “Regional Evaluation of Seagrass Production in the Mexican Atlantic Coasts”, CONACyT-Universidad Autónoma Metropolitana-Unidad Iztapalapa, Margarita E. Gallegos Martínez, and “Ecological and Biological Evaluation of Dominant Ichthyofaunistic Resources in Coral Reefs of Quintana Roo”, Universidad Autónoma Metropolitana- Unidad Iztapalapa, Silvia Díaz-Ruiz.
Referencias
Álvarez, F., A.J. Sánchez & L.A. Soto. 1996. Comparison of two nets as samplers of estuarine macrofauna in a tropical meadow. Rev. Inv. Mar. 17: 17-27. [ Links ]
Barba, E. 1999. Variación de la densidad y la biomasa de peces juveniles y decápodos epibénticos de la región central de Laguna Madre, Tamaulipas. Hidrobiológica 9: 101-114. [ Links ]
Barba, E., A. Raz-Guzman & A.J. Sánchez. 2005. Distribution patterns of estuarine caridean shrimps in the southwestern Gulf of Mexico. Crustaceana 78: 709-726. [ Links ]
Barba, E., A.J. Sánchez, A. Raz-Guzman & M.E. Gallegos. 2000. Dieta natural y tasa de forrajeo del carideo Hippolyte zostericola Smith sobre epífitas de Thalassia testudinum Banks et Solander ex König. Hidrobiológica 10: 139-146. [ Links ]
Bauer, R.T. 1985. Diel and seasonal variation in species composition and abundance of caridean shrimps(Crustacea, Decapoda) from seagrass meadows on the north coast of Puerto Rico. Bull. Mar. Sci. 36: 150-162. [ Links ]
Boschi, E.E. 2000. Species of decapod crustaceans and their distribution in the American marine zoogeographic provinces. Rev. Inv. Des. Pesquero 13: 7-136. [ Links ]
Briggs, J.C. 1974. Marine zoogeography. McGraw-Hill, New York, USA. [ Links ]
Bulger, A.J., B.P. Hayden, M.E. Monaco, D.M. Nelson & M.G. McCormick-Ray. 1993. Biologically-based estuarine salinity zones derived from a multivariate analysis. Estuaries 16: 311-322. [ Links ]
Chace, F.A., Jr. 1972. The shrimps of the Smithsonian- Bredin Caribbean Expeditions with a summary of the West Indian shallow-water species (Crustacea: Decapoda: Natantia). Smith. Contrib. Zool. 98: 1-179. [ Links ]
Cross, R.E. & A.E. Stiven. 1999. Size-dependent interactions in salt marsh fish (Fundulus heteroclitus Linnaeus) and shrimp (Palaemonetes pugio Holthuis). J. Exp. Mar. Biol. Ecol. 242: 179-199. [ Links ]
Díaz-Ruiz, S. 2000. Evaluación Ecológica y Biológica de los Recursos Ictiofaunísticos Dominantes en Arrecifes Coralinos de Quintana Roo. Segunda Etapa. Informe final. [ Links ]
de la Lanza, G. & C. Tovilla. 1986. Una revisión sobre la taxonomía y distribución de los pastos marinos. Univ. Cienc. 3: 17-38. [ Links ]
De Grave, S., N.D. Pentcheff, S.T. Ahyong, T.Y. Chan, K.A. Crandall, P.C. Dworschak, D.L. Felder, R.M. Feldmann, C.H.J.M. Fransen, L.Y.D. Goulding, R. Lemaitre, M.E.Y. Low, J.W. Martin, P.Ng.K. Lin, C.E. Schweitzer, S.H. Tan, D. Tshudy & R. Wetzer, 2009. A classification of living and fossil genera of decapod crustaceans. The Raffles Bull. Zool. Suppl. 21: 1-109. [ Links ]
Dugan, P.J. 1982. Seasonal and geographic distribution of seven decapod crustaceans in Apalachee Bay, Florida. Contrib. Mar. Sci. 26: 65-79. [ Links ]
Eleftheriou, A. & N.A. Holme. 1985. Macrofaunal Techniques, p. 1-349. In N.A. Holme & A.D. McIntyre (eds.). Methods for the study of marine benthos. Blackwell Scientific Publications, Oxford, United Kingdom. [ Links ]
Everett, R.A. & G.M. Ruiz. 1993. Coarse woody debris as refuge from predation in aquatic communities: an experimental test. Oecología 93: 475-486. [ Links ]
Fourqurean, J.W., A. Willsie, C.D. Rose & L.M. Rutten. 2001. Spatial and temporal pattern in seagrass community composition and productivity in south Florida. Mar. Biol. 138: 341-354. [ Links ]
Gallegos-Martínez, M. 1999. Evaluación regional de la producción de pastos marinos de las costas del Atlántico Mexicano. Proyecto de CONACyT-UAMI 1997- 1998. Informe final. [ Links ]
Gilmore, G.R. 1995. Environmental and biogeographic factors influencing ichthyofaunal diversity: Indian River Lagoon. Bull. Mar. Sci. 57: 153-170. [ Links ]
Greening, H.S. & R.J. Livingston. 1982. Diel variation in the structure of seagrass-associated epibenthic macroinvertebrate communities. Mar. Ecol. Progr. Ser. 7: 147-156. [ Links ]
Heck, K.L. & R.J. Orth. 1980. Structural components of eelgrass (Zostera marina) meadows in the lower Chesapeake Bay-Decapod Crustacea. Estuaries 3: 289-290. [ Links ]
Holthuis, L.B. 1952. The subfamily Palaemonidae. Part II. A general revision of the Palaemonidae (Crustacea, Decapoda, Natantia) of the Americas. Occ. Papers, Allan Hancock Foundation 12: 1-396. [ Links ]
Jernakoff, P., A. Brearlely & J. Nielsen. 1996. Factors affecting grazer-epiphyte interactions in temperate seagrass meadows. Oceanogr. Mar. Biol. Annu. Rev. 34: 109-162. [ Links ]
Karplus, I. 1987. The association between gobid fishes and burrowing alpheid shrimps. Oceanogr. Mar. Biol. Annu. Rev. 25: 507-562. [ Links ]
King, S.P. & P. Sheridan. 2006. Nekton of new seagrass habitats colonizing a subsided salt marsh in Galveston Bay, Texas. Estuaries 29: 286 -296. [ Links ]
Kitting, C.L. 1984. Selectivity by dense populations of small invertebrates foraging among seagrass blade surfaces. Estuaries 7: 276-288. [ Links ]
Kitting, C.L., B. Fry & M.D. Morgan. 1984. Detection of inconspicuous epiphytic algae supporting food webs in seagrass meadows. Oecologia 62: 145-149. [ Links ]
Knolton, R.E. & A.B. Williams. 1970. The life history of Palaemonetes vulgaris (Say) and P. pugio Holthuis in coastal North Carolina. Jour. Elisha Mitchell Sci. Soc. 86: 185. [ Links ]
Krebs, C.J. 1989. Ecological methodology. Harper Collins, New York, USA. [ Links ]
Lewis, F.G. III. 1984. Distribution of macrobenthic crustaceans associated with Thalassia, Halodule and bare sand substrata. Mar. Ecol. Progr. Ser. 19: 101-113. [ Links ]
Lewis, M.A. & R. Devereux. 2009. Nonnutrient anthropogenic chemicals in seagrass ecosystems: fate and effects. Environ. Toxicol. Chem. 28: 644-661. [ Links ]
Livingston, R.J. 1984. The relationship of physical factors and biological response in coastal seagrass meadows. Estuaries 7: 377-390. [ Links ]
Llanso R.J., S.S. Bell & F.E. Vose. 1998. Food habits of red drum and spotted seatrout in a restored mangrove Impoundment. Estuaries 21: 294-306. [ Links ]
Magurran, A. E.1988. Ecological diversity and its measurement. Croom Helm, London, United Kingdom. [ Links ]
Markham, J.C., F.E. Donath, J.L. Villalobos & A.C. Díaz. 1990. Notes on the shallow-water marine Crustacea of the Caribbean coast of Quintana Roo, Mexico. Anal. Inst. Biol. Univ. Nac. Autón. Méx., Ser. Zool. 61: 405-446. [ Links ]
Mazzella, L. & G.F. Russo. 1989. Grazing effect of two Gibbula species (Mollusca, Archaeogastropoda) on the epiphytic community of Posidonia oceanic leaves. Aq. Bot. 35: 357-373. [ Links ]
McCune, B. & J.B. Grace. 2002. Analysis of ecological communities. MjM Software Design. Oregon, USA. [ Links ]
McLusky, S.D. 1989. The Estuarine Ecosystem. Chapman & Hall, Glasgow, Ireland. [ Links ]
Meyer, J.L., E.T. Schultz & G.S. Helfam. 1983. Fish schools: An asset to corals. Science 220: 1047-1049. [ Links ]
Nagabhushanam, R., 1961. Tolerance of the prawn Palaemonetes vulgaris (Say) to waters of low salinity. Sci. Cul. 27: 1-43. [ Links ]
Peters, E.C., N.J. Gassman, J.C. Firman, R.H. Richmond & E.A. Power. 1997. Ecotoxicology of tropical marine ecosystems. Environ. Tox. Chem. 16: 12-40. [ Links ]
Pielou, E.C. 1984. The interpretation of ecological data: a primer on classification and ordination. John Wiley & Sons, New York, USA. [ Links ]
Posey, M.H. & A.H. Hines. 1991. Complex predator-prey interactions within an estuarine benthic community. Ecology 72: 2155-2169. [ Links ]
PRIMER –E Ltd. 2008. PRIMER 6 & PERMANOVA + versión 1.01. Multivariate statistics for ecologists. United Kingdom. [ Links ]
Renfro, W.C. 1962. Small beam net for sampling postlarval shrimp. Galveston Biological Lab. U. S. Fish. Wildl. Serv. Circ. 161: 86-67. [ Links ]
Ramírez, A. 1999. Ecología Aplicada. Diseño y análisis estadístico. Fundación Universitaria Jorge Tadeo Lozano, Bogotá, Colombia. [ Links ]
Raz-Guzman, A. & E. Barba. 2000. Seagrass biomass, distribution and associated macrofauna in southwestern Gulf of Mexico coastal lagoons. Biol. Mar. Medit. 7: 271-274. [ Links ]
Román-Contreras, R. 1988. Características ecológicas de los crustáceos decápodos de la Laguna de Términos. In A. Yáñez-Arancibia & J.W. Day (eds.). Ecología de los ecosistemas costeros del sur del Golfo de México: la región de la Laguna de Términos. Instituto de Ciencias del Mar y Limnología, UNAM and Coastal Ecology Institute, Editorial Universitaria, D.F. México. [ Links ]
Román-Contreras, R. & M. Martínez-Mayén. 2009. Shallow water hippolytids from the Mexican Caribbean. Hidrobiológica 19: 119-128. [ Links ]
Román-Contreras, R. & M. Martínez-Mayén. 2010. Palaemonidae (Crustacea: Decapoda: Caridea) from the shallow waters of Quintana Roo, Mexican Caribbean coast. Rev. Mex. Bio. 81: 43-51. [ Links ]
Sánchez, A.J., A. Raz-Guzman & E. Barba. 1996. Habitat value of seagrasses for decapods in tropical coastal lagoons of the southwestern Gulf of Mexico: an overview. In J. Kuo, R.C. Phillips, D.I. Walker & H. Kirkman (eds.). Seagrass Biology, Proceedings of an International Seagrass Workshop. Rottnest Island, Western Australia. [ Links ]
Sánchez, A.J. & A. Raz-Guzman. 1997. Distribution patterns of tropical estuarine brachyuran crabs in the Gulf of Mexico. J. Crust. Biol. 17: 609-620. [ Links ]
Sheridan, P.F. & T.J. Minello. 2003. Nekton use of different habitat types in seagrass beds of lower Laguna Madre, Texas. Bull. Mar. Sci. 72: 37-61. [ Links ]
Sokal, R.R. & F.J. Rohlf. 1981. Biometry. The principles and practice of statistics in biological research. W.H. Freeman and Co., San Francisco, California, USA. [ Links ]
STATSOFT. 2004. STATISTICA. Data analysis software system. Version 7. Tulsa, Oklahoma, USA. [ Links ]
Thayer, G.W., K.A. Bjorndall, J.C. Ogden, S.L. Williams & J.C. Zieman. 1984. Role of larger herbivores in seagrass communities. Estuaries 7: 351-376. [ Links ]
Thorp, J.H. & D.E. Hoss. 1975. Effects of salinity and cyclic temperature on survival of two sympatric species of grass shrimp (Palaemonetes), and their relationship to natural distributions. J. Exp. Mar. Biol. Ecol. 18: 19-28. [ Links ]
Thorp, J.H. 1976. Interference competition as a mechanism of coexistence between two sympatric species of the grass shrimp Palaemonetes (Decapoda: Palaemonidae). J. Exp. Mar. Biol. Ecol. 25: 19-35. [ Links ]
van Tussenbroek, B. 1995. Thalassia testudinum leaf dynamics in a Mexican Caribbean reef lagoon. Mar. Biol. 122: 33-40. [ Links ]
Vosé, E.E. & S.S. Bell. 1994. Resident fishes and macrobenthos in mangrove-rimmed habitats: Evaluation of habitat restoration by hydrologic modification. Estuaries 17: 585-596. [ Links ]
Wicksten, M.K. 1989. Hippolyte zostericola (Crustacea: Decapoda) in the eastern Pacific. Proc. Biol. Soc. Wash. 102: 644-645. [ Links ]
Williams, B.A. 1984. Shrimps, lobsters and crabs of the Atlantic coast of the eastern United States, Maine to Florida. Smithsonian Institution, Washington, D.C., USA. [ Links ]
Zieman, J.C., R. Orth, R.C. Phillips, G. Thayer & A. Thorhaug. 1984. The effects of oil on seagrass ecosystems. In J. Cairns & A.L. Buikema (eds.). Restoration of habitats impacted by oil spills. Butterworth, Boston, Massachusetts, USA. [ Links ]
Zupo, V. & W.G. Nelson. 1999. Factors influencing the association patterns of Hippolyte zostericola and Palaemonetes intermedius (Decapoda: Natantia) with seagrasses of the Indian River Lagoon, Florida. Mar. Biol. 134: 181-190. [ Links ]
*Correspondencia:
Everardo Barba Macías: El Colegio de la Frontera Sur, Unidad Villahermosa, Depto. de Aprovechamiento y Manejo de Recursos Acuáticos. Carretera Villahermosa-Reforma km. 15.5, Ranchería Guineo 2ª sección C.P. 86280 Villahermosa, Tabasco, Mexico. ebarba@ecosur.mx
1. El Colegio de la Frontera Sur, Unidad Villahermosa, Depto. de Aprovechamiento y Manejo de Recursos Acuáticos. Carretera Villahermosa-Reforma km. 15.5, Ranchería Guineo 2ª sección C.P. 86280 Villahermosa, Tabasco, Mexico; ebarba@ecosur.mx
Received 10-VI-2011. Corrected 12-XII-2011. Accepted 01-II-2012.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}