










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.60 n.3 San José Sep. 2012
Aerial organ anatomy of Smilax syphilitica (Smilacaceae)
*Dirección para correspondencia
Abstract
Smilax L. in Brazil is represented by 32 taxa and it is a taxonomically difficult genus because the plants are dioecious and show wide phenotypic variation. The analysis and use of leaf anatomy characters is recognized as a frequently successful taxonomic method to distinguish between individual taxon, when floral material is absent or minute differences in flowers and foliage exist such as in Smilax. The aim of this study was to characterize the anatomical features of the aerial organs in Smilax syphilitica collected from the Atlantic Rainforest, in Santa Teresa-ES and the Smilax aff. syphilitica from the Amazon Rainforest, in Manaus, Brazil. For this, a total of three samples of Smilax were collected per site. Sample leaves and stems were fixed with FAA 50, embedded in historesin, sectioned on a rotary microtome, stained and mounted in synthetic resin. Additionally, histochemical tests were performed and cuticle ornamentation was analyzed with standard scanning electron microscopy. S. syphilitica and S. aff. syphilitica differed in cuticle ornamentation, epidermal cell arrangement and wall thickness, stomata type and orientation, calcium oxalate crystal type, and position of stem thorns. Leaf blades of S. syphilitica from the Amazon Rainforest have a network of rounded ridges on both sides, while in S. aff. syphilitica, these ridges are parallel and the spaces between them are filled with numerous membranous platelets. Viewed from the front, the epidermal cells of S. syphilitica have sinuous walls (even more pronounced in samples from the Amazon); while in S. aff. syphilitica, these cells are also sinuous but elongated in the cross-section of the blade and arranged in parallel. Stomata of S. syphilitica are paracytic, whereas in S. aff. syphilitica, are both paracytic and anisocytic, and their polar axes are directed towards the mid-vein. Calcium oxalate crystals in S. syphilitica are prisms, whereas in S. aff. syphilitica, crystal sand. Thorns occur in nodes and internodes in S. syphilitica but only in internodes in S. aff. syphilitica. These features have proven to be of diagnostic value and may support a separation into two species, but future studies are needed to confirm that S. aff. syphilitica is indeed a new taxon.
Key words: aerial stems, crystals, cuticle, leaves, greenbrier, Smilacaceae.
Resumen
Smilax L. en Brasil está representado por 32 táxones y es un género difícil en lo que respecta a su taxonomía porque las plantas son diocas y presentan mucha variación fenotípica. El análisis y uso de los caracteres anatómicos de la hoja es reconocido frecuentemente como un exitoso método taxonómico para distinguir entre los táxones individuales cuando las flores están ausentes o hay diminutas diferencias. El objetivo de este estudio fue caracterizar los rasgos anatómicos de los órganos aéreos en Smilax syphilitica recolectados en el Bosque Atlántico en Santa Teresa- ES y de Smilax aff. syphilitica del Amazonas, Manaus, Brasil. Un total de tres muestras fueron recolectadas por sitio. Las muestras fueron fijadas en FAA 50, sumergidas en historesina, seccionadas en un micrótomo rotatorio, teñidas y montadas en resina sintética. Se hicieron pruebas histoquímicas y la ornamentación de la cutícula fue analizada con microscopía electrónica de barrido estándar. S. syphilitica y difirieron en la ornamentación de la cutícula, la disposición de células epidérmicas y el grosor de la pared, tipo y orientación de estomas, tipo de cristal de oxalato de calcio y la posición de las espinas del tallo. Estas características han probado ser de valor diagnóstico y pueden apoyar la separación de dos especies, pero se necesitan futuros estudios para confirmar que S. S. aff. syphiliticaaff. syphilitica es un nuevo taxon.
Palabras clave: tallos aéreos, cristales, cutículas, hojas, Smilacaceae.
The genus Smilax L. (Smilacaceae Vent) contains the largest number of species within the Smilacaceae family, of which 32 species occur in Brazil (Andreata 1997, 2009).
The medicinal importance of this genus has been globally recognized since antiquity, and extracts from its leaves and roots are used to treat diseases such as syphilis, gout, rheumatism, skin disorders, asthma, toothaches, wounds, and eye pain (Vandercolme 1947). In Brazil, medicinal use of the Smilax species popularly known as “sarsaparilla” dates back to the 16th century (Medeiros et al. 2007).
Sarsaparilla is often misidentified due to morphological similarities among Smilax species (Lorenzi & Matos 2002). Similarly, Martins & Appezzato-da-Glória (2006) showed that the descriptive features listed in the Brazilian Pharmacopoeia of 1929 are insufficient to differentiate the species.
Andreata (1997) recognized 100 bynonyms for 60 Smilax L. species. The confusion of Smilax species with some of the Herreria ones (Lorenzi & Matos 2002) reinforces the need for a conclusive taxonomic revision, as pointed out by Koyama (1960), Andreata (1980), and Guaglianone & Gattuso (1991).
According to Moore et al. (2010), identification of Smilax species from Thailand by floral features is complicated because the plants have similar flowers; the same problem occurs for Brazilian species (Andreata 1997). Anatomical studies in this group are important, since there are many external feature variations within populations and in the same individual of Smilax as verified by Mandarin-de-Lacerda et al. (1992) and Andreata (1997) in S. rufescens Grisebach and S. japecanga Grisebach respectively, among other species.
Caponetti & Quimby (1956) studied leaf, stem, and root anatomy and found differences between the epidermal and hypodermal cells in the following five species: S. auriculata, S. hispida, S. glauca, S. bona-nox, and S. herbacea. Guaglianone & Gattuso (1991) proposed a comparative framework for distinguishing between four Argentine species of the genus (S. fluminensis, S. cognata, S. pilcomayensis, S. assumptionis and S. campestris) on the basis of mesophyll tissue, parenchyma cells, stomata location, venation patterns, vascular bundles, and leaf blade edges. Marquete & Pontes (1994) compared the anatomy of S. spicata, S. rufescens, and S. fluminensis and observed a difference only in the leaves; S. fluminensis has leaves that are amphistomatic, and the other two species have leaves that are hypostomatic.
Moore et al. (2008) identified the species S. petiolatumidus from Thailand based only on epicuticular leaf architecture. Finally, Palhares et al. (2009) distinguished S. goyazana from other species of the genus based on the stomata; mesophyll; sclerenchyma sheaths shared by mid-vein vascular bundles; the occurrence of idioblasts and raphides; and the presence of starch and tannin in the leaf blade.
The current study presents anatomical characteristics of the aerial system in Smilax syphilitica Humboldt & Bonpland ex Willdenow and S. aff. syphilitica Humboldt & Bonpland ex Willdenow. The first species is found in seven Brazilian states, and its extracts are used in the treatment of syphilis, menstrual cramps, and as an abortifacient (Andreata 1997). Smilax aff. syphilitica has not been described until now. Our objective was to increase the diagnostic value of plant features to support correct identification.
Materials and methods
We analyzed three individual Smilax syphilitica Humboldt & Bonpland ex Willdenow collected from the Atlantic Rainforest in Santa Teresa-ES and the Amazon Rainforest in Manaus, Brazil. The three samples of Smilax aff. syphilitica Humboldt & Bonpland ex Willdenow were collected exclusively from the Amazon Rainforest in Manaus, Brazil. In this study we chose to identify the material as S. aff. syphilitica until future studies, morphological and anatomical, let clarify the true identity of the taxon. Sampling from the Atlantic Rainforest occurred at Santa Teresa-ES (19°56’21.0” S - 0°35’55.2” W) in March 2009. Sampling from the Amazon Rainforest occurred at the Adolpho Ducke Forest Reserve in the municipality of Manaus-AM (03°00’00” - 03°08’00” S - 59°52’40” - 5958’00” W) in January 2010. The collected material was identified by Dr. Regina Helena Potsch Andreata, an expert on the genus Smilax in Brazil, and specimens were incorporated into the ESA herbarium under the numbers 107665 (S. syphilitica-Atlantic Rainforest), 112606 (S. syphilitica-Amazon Rainforest), and 111412 (S. aff. syphilitica- Amazon Rainforest).
The leaf blades were cut at the mid-vein, main vein, interveinal area and at the leaf edge. The stems were analyzed at the third internode (from the apex), the internode closest to the ground, and the underground internode.
For the anatomical study, aerial systems of three adult plants were fixed in FAA 50 (one part formaldehyde: one part glacial acetic acid: 18 parts 50% ethanol, v/v) for 48h (Johansen 1940), dehydrated in a graded ethylic series and infiltrated in glycol methacrylate resin (Leica Historesin-LeicaTM-Wetzlar, Germany).
Serial sections (5-7μm thick) were performed on a rotary microtome and stained with toluidine blue (Sakai 1973), astra blue and basic fuchsin (Roeser 1972). The chemical nature of the cellular content was determined using the following histochemical tests: Ferric trichloride solution for phenolic compounds (Johansen 1940), Lugol’s iodine solution to identify starch (Berlyn & Miksche 1976), Methylene blue to identify pectins (Johansen 1940) and Aniline blue black to identify the total protein (Fisher 1968). For analysis of Calcofluor White MR2 for cellulose (Hughes & McCully 1975) the microscope was equipped for epiillumination with an HBO 50 mercury lamp and a Leica® D filter, providing excitation (Bandpass filter 355-425nm) and suppression (Long-pass filter 470nm).
The tissue dissociation technique was used for analysis of epidermal and lignified mesophyll cells. This technique involves treating the leaf with chromic and nitric acids both at a concentration of 5%, applying dye, and mounting the blade with glycerin (Johansen 1940).
For SEM analyses, leaf blades were fixed in Karnovsky (Karnovsky 1965) for 24h, dehydrated in a graded acetone series and critical point-dried with CO2 (Horridge & Tamm 1969). Samples were attached to aluminium stubs and coated with gold (30-40nm). Then, the samples were examined under a LEO VP 435 scanning electron microscope at 20kV. The ornamental patterns of epicuticular wax were identified following the classification of Metcalfe & Chalk (1979) and Barthlott et al. (1998).
The images were digitally captured with a Leica DMLB microscope (LeicaTM-Wetzlar, Germany) by using a video camera plugged to a computer utilizing the IM50 (LeicaTM-Wetzlar, Germany) software for image analysis.
Results
Leaf blades of Smilax syphilitica and S. aff. syphilitica are leathery, lanceolate- shaped, apex apiculate, obtuse at the base, and fully margined. The venation is acrodromous, with three principle and two inconspicuous veins.
The leaf is complete and features a blade and petiole with a pair of tendrils and a sheath.
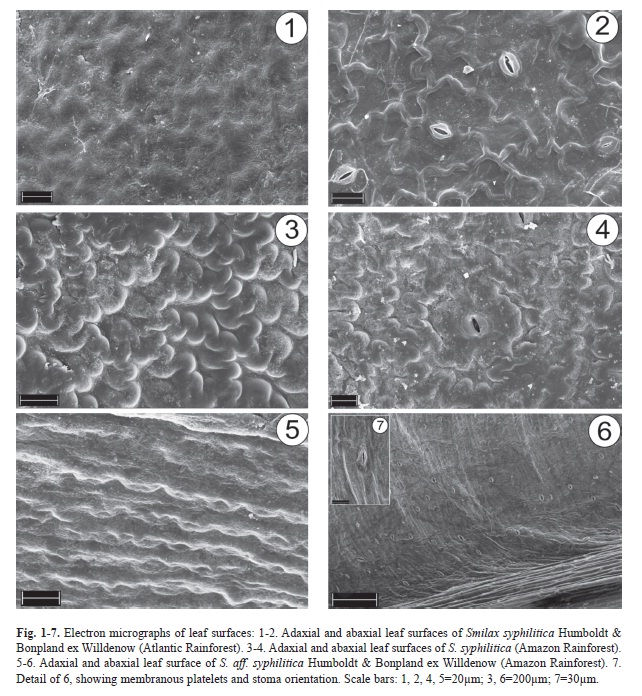
Epicuticular wax structure varies with leaf surface and species (Fig. 1-7). On the adaxial side, leaves of S. syphilitica from the Atlantic Rainforest have granules and platelets (Fig. 1), whereas on the abaxial surface, there is a reticulum with rounded ridges (Fig. 2). Leaf blades of S. syphilitica from the Amazon Rainforest have a network of rounded ridges on both sides (Fig. 3-4), and in S. aff. syphilitica, these ridges are parallel and the spaces between them are filled with numerous membranous platelets (Fig. 5-7).
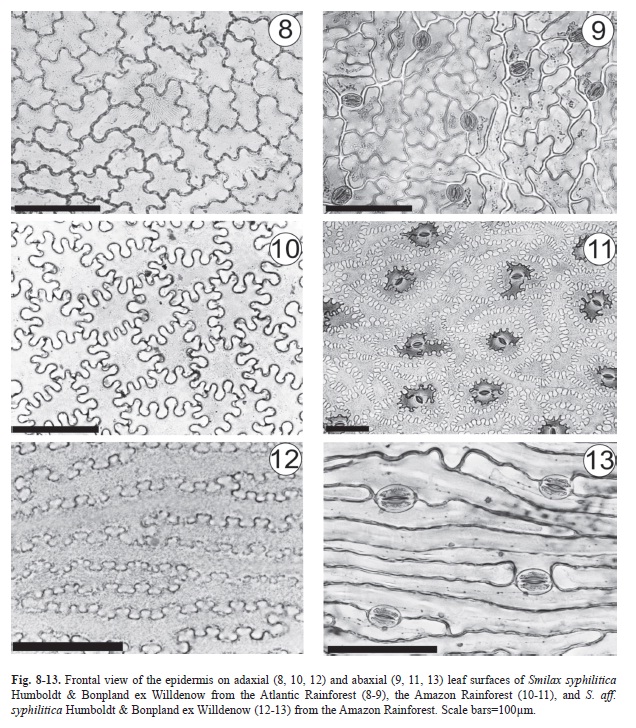
In Smilax syphilitica, the leaves are hypostomatic with randomly oriented stomata at the same level as the adjacent cells (Fig. 8-11); the polar axes are directed towards the mid-vein in S. aff. syphilitica (Fig. 6). However, stomata of S. syphilitica are paracytic (Fig. 9, 11), whereas in S. aff. syphilitica, the stomata are both are paracytic and anisocytic (Fig. 13). In all of the samples, a uniseriate epidermis covered by a thick cuticle, which forms flanges. Viewed from the front, the epidermal cells of S. syphilitica have sinuous walls (Fig. 8-9) that are even more pronounced in samples from the Amazon (Fig. 10-11). In S. aff. syphilitica, these cells are also sinuous but elongated in the cross-section of the blade and arranged in parallel (Fig. 12-13).
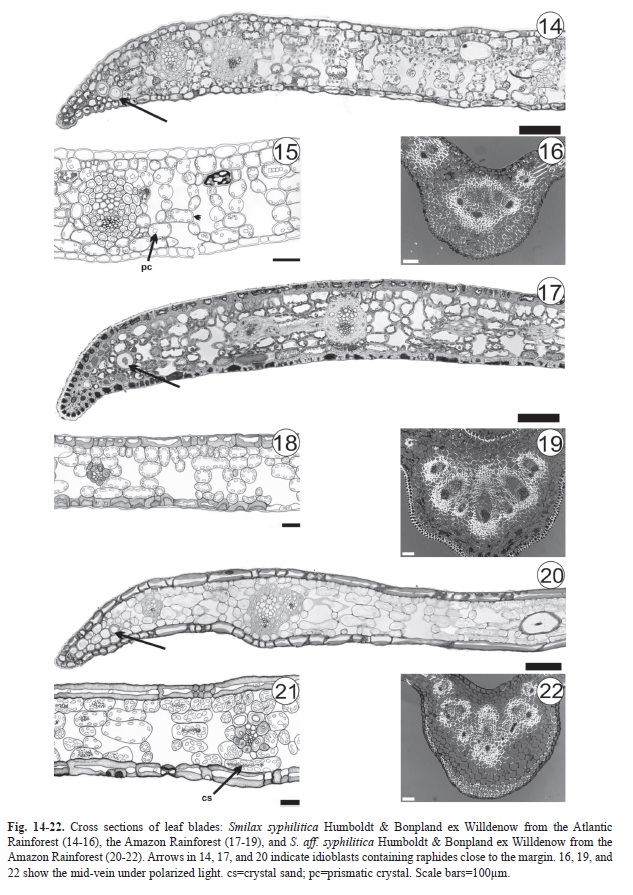
Epidermal cell walls are thickened in the inner periclinal and anticlinal walls, as seen in the cross-sections of S. syphilitica leaves from the Amazon (Fig. 17-18). In S. aff. syphilitica, this thickening occurs in all epidermal cell walls (Fig. 20-21). Staining with methylene blue and Calcofluor White MR2 has shown that the thickened wall is composed of pectin and cellulose.
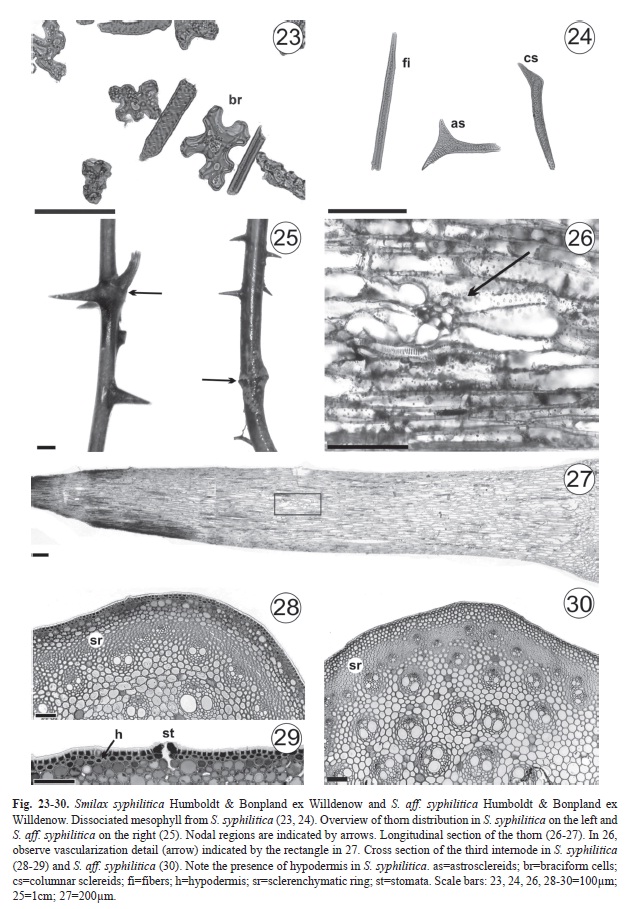
In all of the samples, the mesophyll is homogeneous (Fig. 14-15, 17-18, 20-21) and consists of five-seven layers of braciform cells (Fig. 23). Idioblasts containing raphides along the leaf margin region are common (Fig. 14, 17, 20). Calcium oxalate crystals in S. syphilitica are prisms (Fig. 15), whereas in S. aff. syphilitica, they are crystal sand (Fig. 21).
Fibers, columnar sclereids, and astrosclereids, the latter of which is not observed in S. aff. syphilitica, are the types of lignified cells that were found (Fig. 24). In S. syphilitica from the Atlantic Rainforest, the parenchyma cells have a conspicuous thickening of the cell wall (Fig. 15) not observed in other samples (Fig. 18, 21).
The mid-vein region has between threeeight collateral vascular bundles (Fig. 16, 19, 22), and sclerenchyma sheaths sharedby mid-vein vascular bundles were observed only in S. syphilitica from the Atlantic Rainforest (Fig. 16).
Phenolic, protein, and starch grain content (the latter two are only observed in the midvein) are present only in leaf blade cells of S. syphilitica from both sources.
Thorns (Fig. 25-27) occur in nodes and internodes of S. syphilitica but only in the internodes of S. aff. syphilitica.
In the three internode regions that were analyzed, the epidermis is uniseriate with stomata and covered by a thick cuticle. In S. syphilitica, the epidermis has cells with phenolic and protein content. We observed that the hypodermis, which underlies the epidermis (Fig. 28-29), has different degrees of lignifications and many protein and phenolic idioblasts. This tissue was not observed in plants that attach themselves to tree trunks; this characteristic is typical of vines, but only in those that remain small shrubs. In the third internode of Smilax syphilitica and S. aff. syphilitica, there are between three-five chlorenchyma layers and a continuous sclerenchymatic ring enveloping the vascular cylinder (Fig. 28, 30).
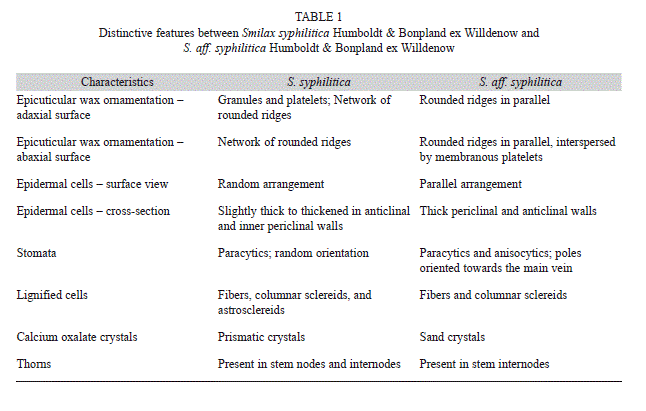
In the cortex of S. syphilitica, phenolic and protein idioblasts are common, and spherical and polyhedral starch grains are observed exclusively in the central parenchyma. Idioblasts containing raphides are restricted to the cortex in S. syphilitica and S. aff. syphilitica. Table 1 summarizes the distinguishing features between them.
Discussion
First, it is noteworthy that very few individuals of the species examined could be located in the field. The reasons behind this distribution are uncertain, although as stated by Lacy (2000), a small population of scattered individuals is more vulnerable to extinction.
Leaf morphology did not vary between S. syphilitica and S. aff. syphilitica or between sampling areas. Although there are differences in the length and width of the leaf blades, the shape and venation patterns are the same.Guaglianone & Gattuso (1991) observed differences in leaf shape in S. campestris Grisebach and recognized that this character is subject to phenotypic variation among species according to its distribution.
According to Jenks & Ashworth (1999), variations in the deposition patterns of epicuticular wax may be attributable to different stressors at various stages of plant development. However, Moore et al. (2008) only used these patterns to taxonomically differentiate the species for the first time (according to the authors). The ornamental patterns of epicuticular wax should not be used as the sole taxonomic tool for distinguishing species of Smilax; in S. syphilitica and S. aff. syphilitica, there are variations between individuals of S. syphilitica from the Atlantic and Amazon Rainforests. However, the patterns observed in S. syphilitica and S. aff. syphilitica differ from those described in S. spicata Vell., S. rufescens Griseb., S. fluminensis Steudel (Marquete & Pontes 1994), and S. polyantha (Martins & Appezzato-da-Glória 2006).
Laakso et al. (2000) found an increase in secondary wall thickness of Pinus L. leaf epidermis that had been subjected to high UVB radiation. Wall thickness is dependent on variations in gene regulation (Caño-Delgado et al. 2000, Kim et al. 2002), thickness, and composition, which involve differentiation and communication between neighboring cells (Roberts 2001). This becomes clearer when examining the DNA of Arabidopsis thaliana (L.) Heynh, in which approximately 1 000 genes are involved in cell wall formation (Roberts 2001). Therefore, this mechanism appears to be more related to genomic characteristics of the S. syphilitica and S. aff. syphilitica specimens we analyzed rather than to environmental influences, suggesting that cell wall formation may consequently be of taxonomic value.
Analysis of epidermal cell walls in surface views has been a widely used method of characterizing species of the genus Smilax L. (Guaglianone & Gattuso 1991, Marquete & Pontes 1994, Guimarães et al. 2010), and we also considered that these cell walls have taxonomic value; both S. syphilitica and have sinuous walls but only in S. S. aff. Syphiliticaaff. Syphilitica they are elongated in the cross-section of the blade and arranged in parallel. According to Haberlandt (1928) the cell wall sinuosity of the epidermis increases contact among adjacent cells, and may help to maintain leaf structure under mechanical stress. Variation in the degree of sinuosity in epidermal cell walls observed in S. syphilitica is related to stress during leaf differentiation (Avery 1933), the cuticle hardening process (Watson 1942), and variations in environmental conditions (Watson 1942, Omosun et al. 2008).
Classification of stomata and mesophyll has also been used for distinguishing species of Smilax (Guaglianone & Gattuso 1991, Marquete & Pontes 1994, Andreata 1997, Martins & Appezzato-da-Glória 2006, Palhares et al. 2009, Guimarães et al. 2010). Thus, leaves of S. syphilitica and S. aff. syphilitica can also be differentiated by this criterion. Among other criteria, Moore et al. (2010) used the orientation of the stomatal poles to differentiate six species (S. verruculosa, S. megacarpa, S. bracteata, S. pottingeri, S. micro-china, and S. corbularia). This analysis of stomatal poles also enhanced our ability to distinguish between S. syphilitica and S. aff. syphilitica.
The presence of idioblasts with raphides on the leaf blade margins of S. syphilitica and S. aff. syphilitica is common for the genus (Yates & Duncan 1970, Guaglianone & Gattuso 1991, Marquete & Pontes 1994, Martins & Appezzato-da-Glória 2006). The prismatic and sand crystals observed throughout the mesophyll of S. syphilitica and S. aff. syphilitica, respectively, are tiny structures that together with the styloids, raphides, and druse crystals make up the five main forms of calcium oxalate crystals (Metcalfe & Chalk 1950). Prychid & Rudall (1999) noted in a review that the absence, presence, and shape of calcium oxalate crystals are “useful taxonomic features” in monocotyledon classification. Crystal morphology can vary between different plant organs and tissues, whereas within the same species, this feature remains constant (Franceschi & Nakata 2005). Differences in crystal type were observed between S. syphilitica and S. aff. syphilitica.
The analysis of mesophyll type, lignified cell type, vascular bundle type, and sclerenchyma sheaths shared along the mid-vein are commonly used for identifying species of the genus (Yates & Duncan 1970, Guaglianone & Gattuso 1991, Marquete & Pontes 1994, Gattuso 1995, Martins & Appezzato-da-Glória 2006, Guimarães et al. 2010) and can also be used to differentiate S. syphilitica and S. aff. syphilitica.
The presence of hypodermis in the stem of S. syphilitica from the Atlantic Rainforest can be explained by the habitats in which individuals were collected; in other words, hypodermis was absent in epiphytes but present in shrubs. This is because in individuals from the Amazon Rainforest (all shrubs), the hypodermis is similar to Lycopordiella cernua (L.) Pic. Serm (Lycopodiaceae), a vine that keeps this tissue until it reaches a height of approximately one meter and needs mechanical support from adjacent plants to develop an epiphytic habit (Rowe et al. 2004). The presence of hypodermis is considered useful for differentiating between species of Indian palm (Arecaceae) (Mathew & Bhat 2008). Similarly, paleobotany studies use this tissue for taxonomic distinction between extinct vine species (Li & Taylor 1998, Dunn et al. 2003). Here, the presence or absence of hypodermis as it relates to species, environment of origin, habit, and stem portion is a major caveat regarding use of this character to differentiate between groups.
Based on anatomical differences between S. syphilitica and S. aff. syphilitica we suggest further morphological studies in all Smilax species occurring in the Amazon region in order to verify if S. aff. syphilitica is in fact a new taxon.
Acknowledgments
We thank to São Paulo Research Foundation– FAPESP-BIOTA Program (Project Proc. nº 05/54984-5 and Proc. nº 05/58964-9), for the financial support of this research and CNPq (National Council for Scientific and Technological Development) for research grants (Proc. nº 302776/2010-9 and 304147/2009-5). We also thank Marli Kasue Misaki Soares for technical laboratory support.
References
Andreata, R.H.P. 1980. Smilax Linnaeus (Smilacaceae). Ensaio para uma revisão das espécies brasileiras. Arq. Jard. Bot. Rio de Janeiro 24: 179-301. [ Links ]
Andreata, R.H.P. 1997. Revisão das espécies brasileiras do gênero Smilax Linnaeus (Smilacaceae). Pesqui. Bot. 47: 7-244. [ Links ]
Andreata, R.H.P. 2009. A new species of Smilax and a key to all species of Minas Gerais, Brazil. Syst. Bot. 34: 28-31. [ Links ]
Avery, G.S. 1933. Structure and development of the tobacco leaf. Am. J. Bot. 20: 565-592. [ Links ]
Barthlott, W., C. Neinhuis, D. Cutler, F. Ditsch, I. Meusel, I. Theisen & H. Wilhelmi. 1998. Classification and terminology of plant epicuticular waxes. Bot. J. Linn. Soc. 126: 237-260. [ Links ]
Berlyn, G.P. & J.P. Miksche. 1976. Botanical microtechnique and cytochemistry. Iwoa State University, Ames, Iwoa. [ Links ]
Caño-Delgado, A.I., K. Metzlaff & M.W. Bevan. 2000. The eli1 mutation reveals a link between cell expansion and secondary cell wall formation in Arabidopsis thaliana. Development 127: 3395-3405. [ Links ]
Caponetti, J.D. & M.W. Quimby. 1956. The comparative anatomy of certain species of Smilax. J. Am. Pharm. Assoc. 45: 691-696. [ Links ]
Dunn, M.T., M. Krings, G. Mapes, G.W. Rothwell, R.H. Mapes & S. Keqin. 2003. Medullosa steinii sp. nov., a seed fern vine from the Upper Mississippian. Rev. Palaeobot. Palyno. 124: 307-324. [ Links ]
Fisher, D.B. 1968. Protein staining of ribonned epon sections for light microscopy. Histochem. Cell Biol. 16: 92-96. [ Links ]
Franceschi, V.R. & P.A. Nakata. 2005. Calcium oxalate in plants: formation and function. Annu. Rev. Plant. Biol. 56: 41-71. [ Links ]
Gattuso, S. 1995. Exomorfologia y anatomia de Smilax campestris Griseb. (Smilacaceae). Acta Farm. Bonaerense 14: 181-190. [ Links ]
Guaglianone, E.S. & S. Gattuso. 1991. Estudios taxonómicos sobre el genero Smilax (Smilacaceae) I. Bol. Soc. Argent. Bot. 27: 105-129. [ Links ]
Guimarães, A.R., C.G. Costa & R.H.P. Andreata. 2010. Morfoanatomia do sistema subterrâneo de Smilax subsessiliflora (Smilacaceae). Rodriguésia 62: 181-194. [ Links ]
Haberlandt, G. 1928. Physiological plant anatomy. MacMillan Company, London, United Kingdom. [ Links ]
Horridge, G.A. & S.L. Tamm. 1969. Critical point drying for scanning electron microscopy study of ciliary motion. Science 163: 817-818. [ Links ]
Hughes, J. & M.E. McCully. 1975. The use of an optical brightener in the study of plant structure. Stain Technol. 50: 319-452. [ Links ]
Jenks, M.A. & E.N. Ashworth. 1999. Plant epicuticular waxes: function, production, and genetics, p. 1-68. In J. Janick (ed.). Horticultural reviews. John Wiley & Sons, New York, USA. [ Links ]
Johansen, D.A. 1940. Plant microtechnique. McGraw-Hill, New York, EEUU. [ Links ]
Karnovsky, M.J. 1965. A formaldehyde-glutaraldehyde fixative of high osmolality for use in electron microscopy. J. Cell Biol. 27: 137-138. [ Links ]
Kim, G., K. Shoda, T. Tsuge, K. Cho, H. Uchimia, R. Yokoyama, K. Nishitani & H. Tsukaya. 2002. The ANGUSTIFOLIA gene of Arabidopsis, a plant CtBP gene, regulates leaf-cell expansion, the arrangement of cortical microtubules in leaf cells and expression of a gene involved in cell-wall formation. EMBO J. 21: 1267-1279. [ Links ]
Koyama, T. 1960. Materials toward a monograph of the genus Smilax. Quart. J. Taiwan Mus. 13: 1-61. [ Links ]
Laakso, K., J.H. Sullivan & S. Huttunen. 2000. The effects of UV-B radiation on epidermal anatomy in loblolly pine (Pinus taeda L.) and Scots pine (Pinus sylvestris L.). Plant Cell Environ. 23: 461-472. [ Links ]
Lacy, R.C. 2000. Considering threats to the viability of small populations. Ecol. Bull. 48: 39-51. [ Links ]
Li, H. & D.W. Taylor. 1998. Aculeovinea yunguiensis gen. et sp. nov. (Gigantopteridales), a new taxón of Gigantopterid stem from the upper permian of Guizhou Province, China. Int. J. Plant Sci. 159: 1023-1033. [ Links ]
Lorenzi, H. & J.A. Matos. 2002. Plantas medicinais no Brasil: nativas e exóticas cultivadas. Instituto Plantarum, Nova Odessa, São Paulo, Brazil. [ Links ]
Mandarim-de-Lacerda, A.F., R.H.P. Andreata & P.R. Peres-Neto. 1992. Ensaio metodológico de avaliação biométrica em folhas adultas de Smilax rufescens (Smilacaceae). Pesqui. Bot. 43: 199-221. [ Links ]
Marquete, O. & R.G. Pontes. 1994. Estudo anatómico foliar comparativo de Smilax spicata Vell., Smilax rufescens Griseb. e Smilax fluminensis Steudel. Rev. Bras. Biol. 54: 413-426. [ Links ]
Martins, A.R. & B. Appezzato-da-Glória. 2006. Morfoanatomia dos órgãos vegetativos de Smilax polyantha Griseb. (Smilacaceae). Rev. Bras. Bot. 29: 555-567. [ Links ]
Mathew, A. & K.M. Bhat. 2008. Anatomical diversity of Indian rattan palms (Calamoideae) in relation to biogeography and systematic. Bot. J. Linn. Soc. 125: 71-86. [ Links ]
Metcalfe, C.R. & L. Chalk. 1950. Anatomy of the dicotyledons. Clarendon, Oxford, United Kingdom. [ Links ]
Metcalfe, C.R. & L. Chalk. 1979. Anatomy of the dicotyledons. Clarendon, Oxford, United Kingdom. [ Links ]
Medeiros, M.F.T., L. Senna-Valle & R.H.P. Andreata. 2007. Histórico e o uso da “salsaparrilha” (Smilax spp.) pelos boticários no Mosteiro de São Bento. Rev. Bras. Bioci. 5: 27-29. [ Links ]
Moore, B.R., N.A. Narkkong, T. Moore & P. Lutat. 2008. Epicuticular leaf architecture confirms a new Smilax species (Smilacaceae) from northeast Thailand. Sci. Asia 34: 103-106. [ Links ]
Moore, B.R., T. Moore & N.A. Narkkong. 2010. A preliminary systematic analysis of leaf epidermal characters for six Smilax species in Thailand. Sci. Asia 36: 175-179. [ Links ]
Omosun, G., A.A. Markson & O. Mbanasor. 2008. Growth and anatomy of Amaranthus hybridus affected by different crude oil concentrations. Am. Eurasian J. Sci. Res. 3: 70-74. [ Links ]
Palhares, D., C.E.S. Silveira, L.B.P. Zaidan & L.A.R. Pereira. 2009. Leaf anatomy of Smilax goyazana (Smilacaceae). Acta Bot. Hung. 51: 115-127. [ Links ]
Prychid, C.J. & P.J. Rudall. 1999. Calcium oxalate crystals in monocotyledons: a review of their structure and systematic. Ann. Bot. 84: 725-739. [ Links ]
Roberts, K. 2001. How the cell wall acquired a cellular context. Plant Physiol. 125: 127-130. [ Links ]
Roeser, K.R. 1972. Die nadel der schwarzkiefer-massenprodukt und kunstwerk der natur. Mikrokosmos 61: 33-36. [ Links ]
Rowe, N., S. Isnard & T. Speck. 2004. Diversity of mechanical architecture of climbing plants: evolutionary perspective. J. Plant Growth Regul. 23: 108-128. [ Links ]
Sakai, W.S. 1973. Simple method for differential staining of paraffin embedded plant material using toluidine blue. Stain Technol. 48: 247-248. [ Links ]
Watson, R.W. 1942. The effect of cuticular hardening on the form of epidermal cells. New Phytol. 41: 223-229. [ Links ]
Vandercolme, E. 1947. História botânica e terapêutica das salsaparrilhas. Rev. Flora Med. 7: 316-524. [ Links ]
Yates, I.E. & W.H. Duncan. 1970. Comparative studies of Smilax, section Smilax, of the southeastern United States. Rhodora 72: 289-312. [ Links ]
*Correspondencia:
João Marcelo Silva: Departmento de Ciências Biológicas, Escola Superior de Agricultura ‘Luiz de Queiroz’, Universidade de São Paulo, 13418-900, Piracicaba, SP, Brazil. joaomarcsilva@yahoo.com.br
Regina Helena Potsch Andreata: Instituto de Ciências Biológicas e Ambientais, Universidade Santa Úrsula, 22610-020, Rio de Janeiro; regina.andreata@gmail.com
Beatriz Appezzato-da-Glória: Departmento de Ciências Biológicas, Escola Superior de Agricultura ‘Luiz de Queiroz’, Universidade de São Paulo, 13418-900, Piracicaba, SP, Brazil. bagloria@usp.br
1. Departmento de Ciências Biológicas, Escola Superior de Agricultura ‘Luiz de Queiroz’, Universidade de São Paulo, 13418-900, Piracicaba, SP, Brazil; joaomarcsilva@yahoo.com.br, bagloria@usp.br
2. Instituto de Ciências Biológicas e Ambientais, Universidade Santa Úrsula, 22610-020, Rio de Janeiro; regina.andreata@gmail.com
Received 18-VIII-2011. Corrected 08-II-2012. Accepted 08-III-2012.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}