










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.60 n.3 San José Sep. 2012
Relaciones entre los hongos filamentosos y solubilizadores de fosfatos con algunas variables edáficas y el manejo de cafetales
Raúl Hernando Posada1*, Marina Sánchez de Prager2*, Ewald Sieverding3*, Karla Aguilar Dorantes1 & Gabriela Patricia Heredia-Abarca1
*Dirección para correspondencia
Abstract
Soil properties and the environment have multiple outcomes on fungal communities. Although, the interaction effects between management intensity, pH, available phosphorus, organic carbon, soil texture and different fractions of water stable macro-aggregates on the communities of microscopic filamentous fungi (MFF), iron phosphate solubilizing fungi (PSF-Fe), and iron and calcium phosphate solubilizing fungi (PSF-(Fe+Ca)), have been previously evaluated in field conditions, this has never been performed in terms of their combined effects, neither with phosphate solubilizing fungi. To assess this, we collected 40 composite soil samples from eight Mexican and Colombian coffee plantations, with different management intensities and physico-chemical edaphic parameters, during 2008-2009. We isolated different communities of MFF, PSFFe and PSF-(Fe+Ca), by wet sieving and soil particles culture in Potato-Dextrose-Agar from soil samples, and we classified isolates in terms of their phosphate solubilizing ability. Following the principal component analysis results, we decided to analyze fungal communities and abiotic factors interactions for each country separately. Structural Equation Models revealed that organic carbon was positively associated to MFF richness and number of isolates (λ>0.58), but its relationship with PSF-Fe and PSF-(Fe+Ca) were variable; while the available phosphorus, pH and water stable macro-aggregate fractions did not show a clear pattern. Management intensity was negatively related to PSF-Fe (λ≤-0.21) morphotype richness and the number of isolates in Colombian coffee plantations. We found that the relationships of clay and organic carbon content, and available phosphorus and soil pH, with the species richness and number of isolates of MFF, PSF-Fe and PSF-(Fe+Ca) were highly variable; this made impossible to generalize the responses between saprotrophic fungal groups and geographic zones. The management intensity was not related to species richness and number of isolates of MFF in any coffee areas, while for PSF the relationship could not be defined. The different water stable macro-aggregates fractions did not show a defined pattern in relation to the species richness and the number of isolates of saprophytic and phosphate solubilizing fungi (PSF). This study highlights the need to take into account edaphic and geographic context in order to reach a better understanding of the intensity management effects on MFF and PSF function in agroecosystems.
Key words: tricalcium phosphate, iron phosphate, water-stable aggregates, soil, fungal richness, fungal abundance,structural equation models.
Resumen
El suelo y sus propiedades tienen múltiples relaciones con las comunidades fúngicas. El efecto conjunto de la intensidad de manejo y las variables edáficas, incluida la estabilidad de agregados sobre las comunidades de hongos microscópicos filamentosos (HMF), solubilizadores de fosfato de hierro (HSP-Fe) y solubilizadores tanto de fosfato de hierro como de calcio (HSP-(Fe+Ca)) no han sido evaluadas en campo. A partir de 40 muestras edáficas de ocho plantaciones de café de Colombia y México, con diferentes intensidades de manejo (IMPC) y con diferencias en sus variables edáficas, se aislaron y evaluaron las comunidades de HMF, HSP-Fe y HSP-(Fe+Ca) durante 2008-2009. Empleando modelos basados en ecuaciones estructurales se encontró que el carbono orgánico se relacionó positivamente con la riqueza y abundancia de HMF (λ>0.58) y fue variable en su relación con HSP-Fe y HSP-(Fe+Ca). Las relaciones del fósforo disponible, pH y las fracciones de macro-agregados fueron altamente variables. El IMPC se relacionó negativamente con HSP-Fe (λ≤-0.21) en cafetales colombianos. Se discuten las interacciones para cada conjunto de variables (químicas, estabilidad de macro-agregados y de manejo de plantación) y se explican las relaciones resultantes. Las relaciones de cada variable son inseparables del contexto edáfico y geográfico, los cuales imprimen marcadas diferencias.
Palabras clave: trifosfato de calcio, fosfato de hierro, agregados estables al agua, suelo, riqueza fúngica, abundancia fúngica, modelos basados en ecuaciones estructurales.
El café (Coffea arabica L.) es cultivado en más de 70 países (Bhattacharya & Bagyaraj 2002) y es uno de los productos de más intenso comercio del mundo moderno; representa una importante fuente de ingresos para diversos países en África, Asia y Latinoamérica, generando directa e indirectamente millones de empleos (Andrade et al. 2009). Según la Organización Internacional del Café (ICO 2011), Colombia y México se encuentran actualmente entre los 10 mayores productores de café del mundo.
Las zonas productoras de café en estos dos países se encuentran mayoritariamente en suelos de origen volcánico, en los cuales el bosque ha sido reemplazado por plantaciones cafetaleras. El origen volcánico del suelo implica la presencia de fósforo, pero este se encuentra entre un 54-90% inmovilizado por el aluminio y hierro no cristalinos, lo cual repercute en una escasa disponibilidad de este nutriente, constituyendo un serio limitante para el crecimiento de las plantas naturales y los cultivos (Geissert & Ibañez 2008, Codazzi 2009). La escasa movilidad y alta retención edáfica de este nutriente, acompañados de la condición ácida de la mayoría de los suelos han llevado al uso de diferentes estrategias para abastecer las demandas del cultivo, principalmente mediante el suministro de enmiendas orgánicas y/o químicas (Arcila et al. 2007, Hernández-Martínez 2008) y la aplicación de hongos que forman micorriza arbuscular (Rodríguez 2001, Cardoso & Kuyper 2006, Herrera-Peraza et al. 2011), pero en este cultivo no se ha implementado el uso de hongos solubilizadores de fosfatos.
La mayoría de los hongos del suelo están conformados por microscópicos filamentos hifales o micelio y se les denomina hongos microscópicos filamentosos (HMF), y por su capacidad saprofítica también son conocidos como micromicetos saprobios filamentosos. Participan en el equilibrio edáfico mediante la descomposición de los residuos vegetales y animales, el ciclaje de nutrientes y la descontaminación de suelos, entre otros procesos (Dighton 2003). Dentro de este grupo de hongos es común encontrar hongos solubilizadores de fosfatos (HSP), que mediante acción enzimática específica liberan los fosfatos capturados en la fracción mineral y lo hacen disponible para las plantas. Debido a la abundante presencia de los HMF en todos los sistemas naturales y agrícolas, se esperaría la presencia de HSP en los sistemas cafetaleros, sin embargo hasta la fecha no hay reportes de su presencia.
El suelo además de proveer nutrientes a los microorganismos, constituye el ambiente químico y físico donde se asocian con el entorno. Existe evidencia de la influencia positiva de la materia orgánica en el crecimiento y la riqueza de especies fúngicas (Dix & Webster 1995, Girvan et al. 2004); la materia orgánica también ha mostrado una relación negativa con la abundancia de hongos solubilizadores de fosfatos (Narsian & Patel 2009), y aún se desconoce su relación con la riqueza de especies.
En general el crecimiento fúngico es favorecido por las condiciones ácidas del suelo (Griffin 1996), y en particular los HSP han mostrado en condiciones in vitro (Narsian & Patel 2009, Aciego & Brookes 2009, Wu et al. 2009) que crecen en condiciones ácidas y además utilizan la acidificación como mecanismo para hacer disponible el fósforo; por lo tanto, reducciones de pH deberían ser favorables para su abundancia en condiciones de campo. Por su parte, el fósforo disponible (P) suele ser limitante del crecimiento en sistemas edáficos ácidos, debido a su inmovilización mineral con el hierro y aluminio contenidos en la fracción arcillosa (Hinsinger 2001); la presencia de P permite el crecimiento fúngico indiferenciado, pero en condiciones de escasez de P, los HSP deberían ser privilegiadamente favorecidos sobre otros hongos saprobios debido a su capacidad solubilizadora de fósforo a partir de diferentes fuentes inorgánicas (Lauber et al. 2008, Narsian & Patel 2009). Adicionalmente, ni para el pH ni para el P, existen estudios en donde se muestre su relación con la riqueza de especies fúngicas.
La actividad fúngica predomina en los macro-agregados de suelo (≥0.25mm de diámetro), donde contribuye a su estabilización, mientras la presencia bacteriana predomina en los micro-agregados (Ashman et al. 2003, Six et al. 2004). En cuanto a la agregación del suelo, se conoce que la ausencia de arado conduce al incremento en la agregación del suelo por la reducción de la descomposición de la materia orgánica contenida en los macro-agregados (Bossuyt et al. 2002), y que los macro-agregados de mayor tamaño están correlacionados positivamente con la riqueza y diversidad de especies fúngicas (Wang et al. 2010); a pesar de eso, se desconoce la relación de las diferentes fracciones estables y la textura del suelo con la riqueza de especies y la abundancia de aislamientos de grupos fúngicos específicos.
El interior de los microporos (0.2-2.5μm, Tisdall 1994) es poco accesible a las hifas fúngicas (3-8μm diámetro), motivo por el cual los hongos habitan preferencialmente los espacios entre los agregados y el interior de los macroporos (>2.5μm diámetro) predominantes en los macro-agregados (Six et al. 2004). Los HSP son indistinguibles del resto de HMF en el grosor de sus hifas, por lo tanto los HMF y los HSP pueden ocupar todas las fracciones de macro-agregados. Por otra parte una extensa red hifal se asocia con una mayor estabilidad de los agregados (Oades & Waters 1991, Six et al. 2004) y por lo tanto con la presencia de macro-agregados de mayor tamaño, motivo por el cual a pesar de la presencia fúngica en las diferentes fracciones, una mayor abundancia puede estar asociada con las fracciones de macro-agregados de mayor tamaño.
Diversos trabajos con especies vegetales de ciclo corto confirman la influencia negativa del manejo agronómico sobre las comunidades y la actividad de la microbiota edáfica, incluidos los HMF y HSP (Girvan et al. 2004, Oliveira et al. 2009, Suzuki et al. 2009). En el caso del café, como otros cultivos de carácter permanente, carece de arado y las condiciones edáficas son el resultado de varios años de permanente manejo; por este motivo los diferentes grupos fúngicos podrían haberse adaptado a las características edáficas y por lo tanto su riqueza y abundancia pueden ser menos influenciables por el manejo agronómico.
Todo el conjunto de incógnitas asociadas a las posibles relaciones de las variables edáficas (carbono orgánico, pH, P, agregados estables al agua y la proporción de arena, limo y arcilla), y la intensidad de manejo con la riqueza de especies y la abundancia de comunidades de HSP-(Fe+Ca), como subgrupo de los HSP-Fe y éstos a su vez como parte de los HMF, en sistemas agrícolas de carácter permanente y sin arado, hace interesante al sistema cafetalero para su estudio.
Con el objetivo de relacionar la riqueza de especies (S) y el número de aislamientos (Nai) de HMF, de HSP-Fe y HSP-(Fe+Ca), con las diferentes fracciones de macro-agregados estables, con las variables físico-químicas edáficas (pH, fósforo disponible, carbono orgánico y la proporción de arena, limo y arcilla) y con la intensidad de manejo de los cafetales; se realizaron análisis de regresión múltiple a partir de datos obtenidos del muestreo edáfico de ocho agroecosistemas cafetaleros localizados en Colombia y México. Postulamos las siguientes hipótesis:
a) El carbono orgánico estará relacionado de forma positiva con la S y el Nai de los tres grupos fúngicos estudiados, debido al suministro de carbono, necesario para el crecimiento fúngico.
b) El pH estará más asociado con la S y el Nai de ambos grupos de HSP en comparación con la S y el Nai de los HMF.
c) Por su capacidad para solubilizar fosfatos de fuentes minerales, la S y el Nai de los HSP estarán menos relacionados con los contenidos edáficos de P disponible en comparación a los HMF.
d) La S y el Nai de HMF, HSP-Fe y HSP-(Fe+Ca), se relacionarán positivamente con las fracciones de agregados estables de mayor tamaño.
e) La S y el Nai de los diferentes grupos fúngicos estudiados estará asociada negativamente con la intensidad de manejo de las plantaciones de café (IMPC).
Materiales y métodos
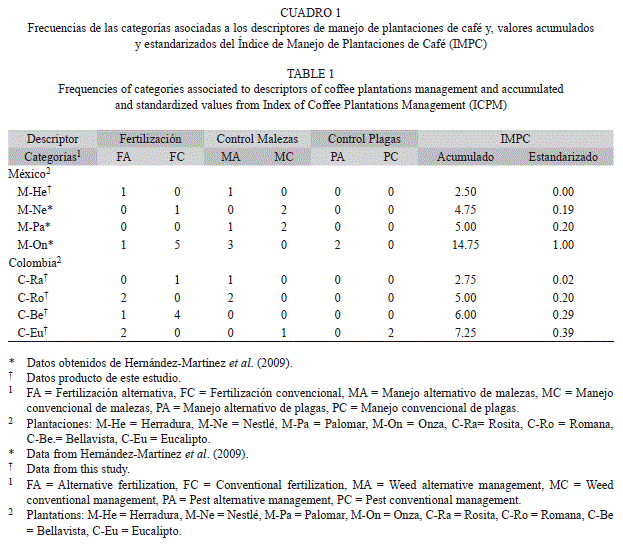
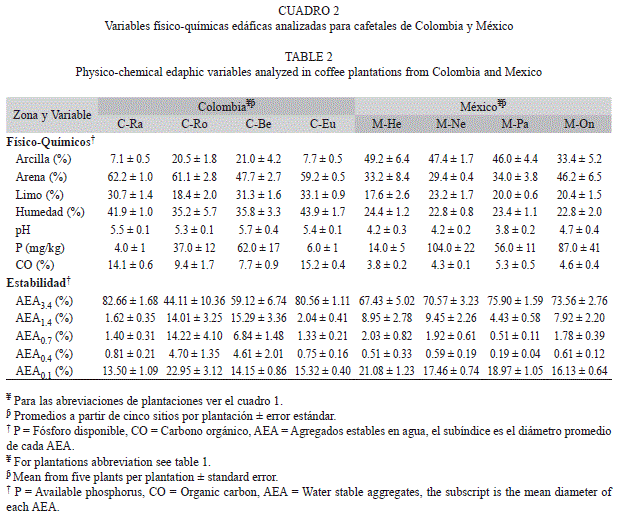
Sitios de muestreo: Este estudio se realizó a partir de muestras edáficas tomadas en ocho plantaciones de dos zonas productoras de café en Colombia y México. En Colombia las plantaciones seleccionadas fueron: Roma (C-Ro), Rosita (C-Ra), Eucalipto (C-Eu) y Bellavista (C-Be), localizadas en el municipio de Apia - Risaralda, con intervalo de latitud entre 5°7’55.04’’ N - 5°5’38.55’’ N y longitud 75°59’41.41’’ W - 75°57’41.32’’ W y altitud de aproximadamente 1 600m.s.n.m. En México las plantaciones estudiadas fueron: Herradura (M-He), Onza (M-On), Nestlé (M-Ne) y Palomar (M-Pa), localizadas la primera en el municipio de Xalapa, la segunda y tercera en Coatepec y Palomar en Teocelo en el estado de Veracruz, con intervalo de latitud entre 19°30’52.44’’ N - 19°22’53.62’’ N y longitud 96°59’20.04’’ W - 96°56’3.12’’ W y altitud aproximada de 1 200m.s.n.m.
Para las zonas cafetaleras de ambos países los suelos estudiados son andisoles: Typic dystrandept para Colombia y Andic Hapludalf para México, cuyo origen volcánico hace que presenten una moderada retención de agua, buen drenaje, baja capacidad de intercambio catiónico (CIC), pobre saturación de bases y generalmente fósforo disponible bajo (Geissert & Ibáñez 2008, Codazzi 2009).
En la región de estudio en Colombia se presentan dos periodos de lluvias, marzo-mayo y septiembre-noviembre, y dos periodos secos, diciembre-febrero y junio-agosto (Guzmán & Jaramillo 1989). En México el periodo seco comprende desde febrero hasta junio, y el de lluvias de junio a octubre; adicionalmente, hay presencia de un fenómeno repetitivo de corta duración denominado “nortes” entre octubre y febrero, caracterizado por vientos fuertes con temperaturas inferiores al promedio mensual de los meses de invierno. Además de los periodos de lluvias y secas, presenta estaciones climáticas bien definidas (Montiel & Robledo 1998).
En la selección de las plantaciones se buscó diferencias contrastantes en el manejo, desde químico hasta orgánico y para cuantificar la intensidad de manejo, se empleó un índice específico para cafetales denominado “Índice de Manejo de Plantaciones de Café o IMPC” (Hernández-Martínez 2008). El IMPC se calculó a partir de encuestas a los agricultores, donde se obtuvo la información de los productos, frecuencia de aplicación y prácticas de manejo usadas para la fertilización y para el control de plagas y malezas en un periodo anual; sin embargo, el IMPC no considera las cantidades y concentraciones de los productos aplicados, ni toma en cuenta información específica como los ingredientes activos, las formulaciones o los tiempos de aplicación de los productos antes de la evaluación.
Cada práctica agronómica fue categorizada como alternativa o convencional, resultando en seis categorías: fertilización alternativa (FA) o convencional (FC), manejo de malezas alternativo (MA) o convencional (MC), manejo de plagas alternativo (PA) o convencional (PC). Producto de esta categorización se calculó el IMPC como sigue: IMPC= [(FA*1)+(FC*1.25)]+[(MA*1.5)+(MC*1.75)]+[(PA*1.5)+(PC*1.75)]. De esta forma se pudo caracterizar cada una de las plantaciones por su intensidad de manejo (Cuadro 1). Los datos de las plantaciones Nestlé, Onza y Palomar en México fueron producto de las evaluaciones de Hernández-Martinez et al. (2009), la información de las demás plantaciones fue producto de este trabajo. Finalmente, los valores de IMPC se estandarizaron a valores entre “cero” y “uno”, donde uno “1” correspondió a mayor intensidad de manejo y/o manejo agrícola convencional, sometido a insumos de síntesis industrial, y la calificación cercana a “cero” significó menor intensidad de manejo y/o un manejo alternativo, cercano a lo orgánico.
Toma de muestras: Todas las muestras se tomaron en el periodo seco, la precipitación en los sitios de estudio en los cinco días previos al muestreo alcanzó 12.3mm en Colombia (diciembre de 2008) y 6.9mm en México (abril de 2008). Las plantas de cada plantación se seleccionaron con la misma altura y cobertura y los sitios de muestreo tenían similar inclinación topográfica, de tal manera que el desarrollo de las plantas y las condiciones de crecimiento en cada plantación fueron similares. En cada finca se tomaron cinco plantas seleccionadas al azar y con una distancia mínima de cuatro metros entre ellas, bajo el dosel de cada una de las plantas se tomaron tres muestras de suelo equidistantes a 20cm del tallo y, entre 5-10cm de profundidad. Las muestras se almacenaron en refrigeración para su posterior análisis. Antes de ser procesadas se agitaron, mezclaron y homogenizaron en una muestra compuesta por planta, lo que representa 20 muestras por región geográfica.
Variables físicas y químicas del suelo: Se analizaron: el pH (1:1) en agua por medio de potenciómetro, el fósforo disponible por el método de Bray II (Fixen & Grove 1990) y los contenidos de arena, limo y arcilla por medio de la determinación de la textura por el método hidrométrico de Bouyoucos (Bouyoucos 1962). La determinación del contenido de carbono orgánico del suelo (CO), se realizó siguiendo el procedimiento descrito en Manns et al. (2007) de pérdida por ignición, el cual es equivalente en materia orgánica (MO) a: MO=CO*1.724.
La estabilidad de los agregados de suelo se valoró siguiendo el método de Yoder (1936), modificado y empleado por el Centro Internacional de Agricultura Tropical (CIAT), Colombia, 2009. El método consistió en tamizar el suelo con mallas de 4.8 y 2.36mm de apertura, posteriormente 20 a 25g del suelo remanente sobre el tamiz de 2.36mm se secaron al aire durante 24hr; simultáneamente una submuestra de 10g se secó a 105oC durante 24hr para determinar el peso seco inicial (PS1). El suelo secado al aire fue colocado en la cima de una serie de tamices (2, 0.84, 0.5 y 0.25mm de apertura de malla), y fue cuidadosamente depositado en un recipiente con agua, de tal manera que el suelo del tamiz superior estuviera marginalmente en contacto con agua para pre-humedecimiento por capilaridad, a temperatura ambiente por 10min antes del tamizaje. La separación de los agregados, se realizó por agitación vertical con continuo ascenso y descenso de los tamices de una distancia de aproximadamente 3.5cm, a razón de 30 oscilaciones por minuto por 30min. A continuación los agregados estables de cada tamiz se secaron por separado a 105oC y se obtuvo el peso seco (PS2). El peso del material grueso de los agregados (PS3), se determinó en cada fracción por remoción de la materia orgánica mediante aplicación de hexametafosfato de sodio (50g/L) por 20min. Finalizado este tiempo a cada fracción se le realizó tamizaje húmedo sobre la malla de donde fue obtenida y el remanente sobre el tamiz fue secado a 105oC durante 24hr.
Con los datos obtenidos, se hizo la corrección del peso del material grueso y humedad de las muestras para cada fracción. El porcentaje de Agregados Estables en Agua (AEA) fue definido en cinco categorías, donde el subíndice de cada AEA corresponde al diámetro promedio entre las dos aperturas de malla: AEA3.4 (4.8-2mm), AEA1.4 (2-0.84mm), AEA0.7 (0.84-0.5mm), AEA0.4 (0.5-0.25mm), AEA0.1 (0.25-0mm). Los AEA se calcularon como la relación entre los pesos obtenidos previamente: AEA(x) = (PS2 - PS3 / PS1 - PS3) * 100 (Márquez et al. 2004, Daraghmeh et al. 2009, Su et al. 2009). En el presente trabajo sólo se analizó el efecto de las fracciones superiores a 0.5mm (AEA3.4, AEA1.4 y AEA0.7) debido a que en ellas se desarrolla la mayor actividad fúngica y donde hay mayor disponibilidad de oxígeno (Ashman et al. 2003, Six et al. 2004, Six et al. 2006).
Obtención de aislamientos fúngicos: Se empleó el método de filtrado de partículas mediante tamizado húmedo descrito en Bills et al. (2004) hasta obtener gránulos de suelo con micelio vivo adherido, sin esporas o propágulos libres. Los gránulos se secaron a temperatura ambiente y sembraron en diez cajas con Papa-Dextrosa-Agar (PDA) de preparación comercial (BIOXON, 39g/l), incluyendo una mezcla de antibióticos para evitar bacterias y hongos de rápido crecimiento (Cloranfenicol-0.1g/L, Gentamicina-50mg/L, Rifampicina-50mg/L y Tetraciclina-50mg/L). Cinco gránulos fueron sembrados por caja de Petri, para un total de 50 gránulos por sitio y 250 gránulos por plantación. A partir de los cuatro días de incubación a temperatura ambiente (22±4ºC) y durante diez días, se extrajeron todas las colonias que emergían de las partículas de suelo, y se sembraron en tubos con PDA inclinado y sin antibióticos. En los casos donde dos o más colonias crecieron, fueron realizados sub-cultivos hasta obtener cultivos puros, los cuales se agruparon en morfoespecies por similitud en caracteres como color, apariencia y forma de las colonias. La S fue considerada como el número de morfoespecies aisladas y, el Nai fue cuantificado como el número de colonias emergentes en el medio PDA.
Pruebas de solubilización: Los cultivos puros fueron mantenidos en PDA inclinado sin antibióticos en refrigeración (3±1ºC) y siete días antes de las pruebas fueron resembrados en cajas Petri con el mismo medio e incubados a temperatura ambiente (22±4ºC). Cada uno de los aislamientos obtenidos se sometió a una prueba de solubilización de fosfato de hierro (FePO4.H2O, PFe) empleando el medio de Sundara & Sinha (1963) consistente en: (NH4)2SO4, 0.5g; KCl, 0.2g; MgSO4.7H2O, 0.3g; MnSO4.H2O, 0.004g; FeSO4.7H2O, 0.002g; NaCl, 0.2g; D-Glucosa, 10g; extracto de levadura, 0.5g; cloranfenicol, 0.1g; agar, 18g; agua, 0.9L + solución de fosfato (goma arábiga, 0.5g; FePO4.H2O o Ca3(PO4)2, 0.5g; agua, 0.1L). El pH del medio se ajustó a 6.8 con una solución de NaOH 1M. El medio se esterilizó en autoclave durante 15min a 22psi y 121ºC. A los aislamientos que presentaron actividad solubilizadora de PFe, también se les realizó la misma prueba de solubilización empleando fosfato tricálcico (PCa) como fuente de P.
A partir de los cultivos puros se realizó la inoculación por punción central en cajas Petri, en los medios mencionados anteriormente. Después de seis días de incubación a 25ºC, se registró para el medio con PCa, la aparición de halos de solubilización alrededor de las colonias. Para el medio con PFe se evaluó cuidadosamente por inspección al estereoscopio la desaparición de los gránulos de PFe por debajo y/o alrededor de las colonias, lo cual fue considerado como resultado positivo de solubilización. Para poner a prueba las hipótesis del presente trabajo se consideró solamente los resultados positivos o negativos en las pruebas de solubilización de PCa y PFe, los detalles cuantitativos pueden ser consultados en Posada et al. (2012). Solamente los aislamientos que mostraron actividad solubilizadora de PCa y/o PFe fueron identificados hasta género siguiendo las claves de Domsch et al. (1980); los aislamientos no identificados debido a la ausencia de estructuras reproductivas, fueron considerados como “otros aislados”.
Un análisis de componentes principales (ACP) basado en una matriz de correlación de 40 puntos por 12 variables edáficas y de manejo, fue realizado en búsqueda de variables de asociación que pudieran resumir la variabilidad obtenida y para observar la presencia de patrones de agrupación de los 40 puntos de muestreo evaluados. Todos los cálculos se efectuaron con el programa Statistica 6 (Programa de Stat Soft, Inc., Tulsa, OK, EEUU).
Para relacionar las variables edáficas yel manejo de las plantaciones de café con la S y el Nai de HMF, HSP-Fe y HSP-(Fe+Ca), se usaron Modelos basados en Ecuaciones Estructurales (MEE, Grace & Keeley 2006). El bajo número de datos para cada variable (20 por país), redujo la cantidad de parámetros – varianzas, covarianzas y correlaciones – que se podían determinar en los modelos (Arbuckle 2010); por lo tanto, los efectos de las variables físico-químicas, de estabilidad de agregados y del manejo de las plantaciones se evaluaron en modelos separados.
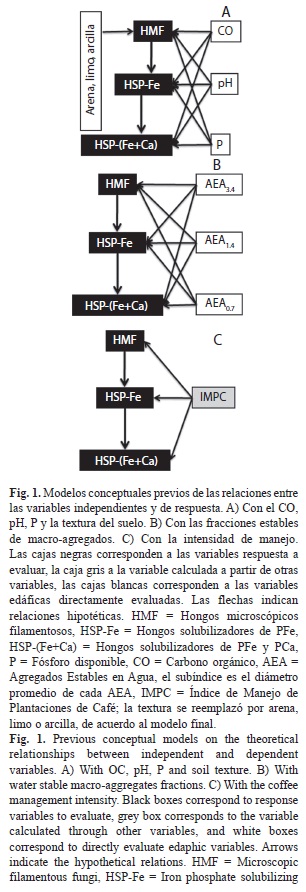
Se crearon tres modelos conceptuales previos (Fig. 1) de las relaciones hipotéticas entre las variables independientes (físico-químicas, de estabilidad y el manejo agronómico) y la S y el Nai de los grupos fúngicos evaluados como variables respuesta (Bollen 1989, Shipley 2000, McCune & Grace 2002). En el modelo A (Fig. 1), se consideró los porcentajes de arena, limo y arcilla del suelo como variables que afectan en cada región geográfica por igual a todos los HMF, incluidos los HSP, por lo tanto no se estableció relación hipotética directa de esta variable con los HSP-Fe o HSP-(Fe+Ca).
A partir de los modelos previos se establecieron las relaciones necesarias para reducir el chi cuadrado (χ2) y obtener un alto valor de P. En este test de χ2, valores de P bajos indican carencia de ajuste, contrario a muchas pruebas estadísticas, altos valores de P son indicadores de un buen ajuste. Las interacciones cuya presencia no contribuyó o dañó el ajuste del modelo (Cuando los coeficientes de ruta (λ) fueron ≤0.05) fueron removidos para conservar las variables necesarias y mantener el poder estadístico. Basados en las interacciones resultantes, se realizó una búsqueda heurística entre los subgrupos de interacciones que generaran posibles variaciones del modelo, buscando una óptima combinación de simplicidad y ajuste. El modelo de mejor ajuste se seleccionó siguiendo la máxima P y mínima c/df (discrepancia/grados de libertad) como medidas determinantes; de no existir coincidencia se seleccionó el modelo con c/df más cercano a 1 (Byrne 1989, Arbuckle 2010).
Las relaciones del carbono orgánico, el pH y el fósforo disponible con la S y el Nai de los grupos fúngicos estudiados, fueron representadas como coeficientes estandarizados (estandarizados por la desviación estándar de las variables), son una medida de la fuerza de la relación causal y matemáticamente son iguales a los coeficientes de regresión parcial o coeficientes de correlación (Antoninka et al. 2009). Los valores de r2 representan la proporción de la varianza explicada por cada conjunto de variables vinculadas a los HMF, HSP-Fe o HSP-(Fe+Ca).
Los Modelos basados en Ecuaciones Estructurales (MEE) son una herramienta idónea para estos propósitos, porque permiten el análisis de los efectos de las variables entrelazadas sobre las variables respuesta y también facilitan el examen de las interacciones entre las variables respuesta. Los MEE derivan de una síntesis de análisis de relaciones y análisis de factores, usualmente con un test de bondad de ajuste basado en máxima verosimilitud (Antoninka et al. 2009). Por medio de la comparación de la covarianza de la estructura de datos que está implícita en el modelo, con la estructura de covarianza actualizada con los datos, se probó si el modelo se ajustaba a los datos (Grace & Pugesek 1998). Los coeficientes de la relación indirecta fueron matemáticamente obtenidos usando las reglas de relaciones por medio de la cuantificación de los efectos de todas las rutas indirectas a las variables de interés (Grace 2006); de esta manera, los efectos totales fueron calculados por la suma de los efectos directos e indirectos. Todos los análisis fueron realizados usando el programa AMOS 18 (Programa de SPSS-2010).
Resultados
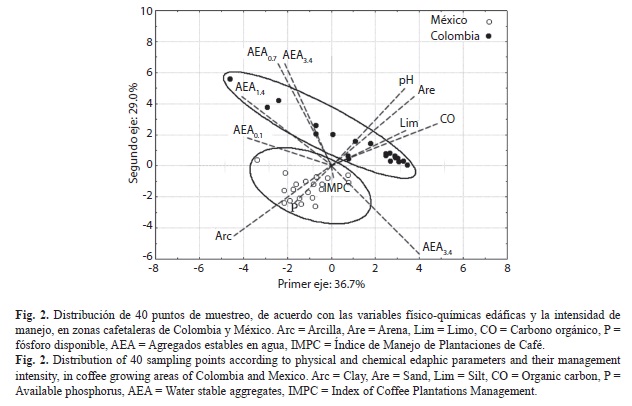
En general la variación entre puntos de muestreo fue mayor en cafetales colombianos que en mexicanos (Fig. 2). Al agrupar los puntos de muestreo por plantaciones (promedio de cinco puntos de muestreo ± error estándar) se observó que el contenido de arcilla y el AEA0.1 fueron mayores en México, mientras la arena, pH, CO, AEA0.7, y AEA0.4 fueron más altos en Colombia (Cuadro 2). A diferencia del pH; el CO y las fracciones AEA0.7 y AEA0.1 fueron más variables entre plantaciones que entre sitios de muestreo en ambos países. Los contenidos de arena, limo, arcilla, P y las fracciones AEA3.4, AEA1.4 y AEA0.4 fueron más variables entre plantaciones que entre puntos de muestreo en Colombia y al contrario en México. Los contenidos de CO fueron altos a muy altos para suelos volcánicos de los cafetales de Colombia y medios a altos para los de México. El fósforo disponible fue de muy bajo a muy alto en cafetales colombianos (Codazzi 1998) y de medios a muy altos para los mexicanos (Portilla 2001).
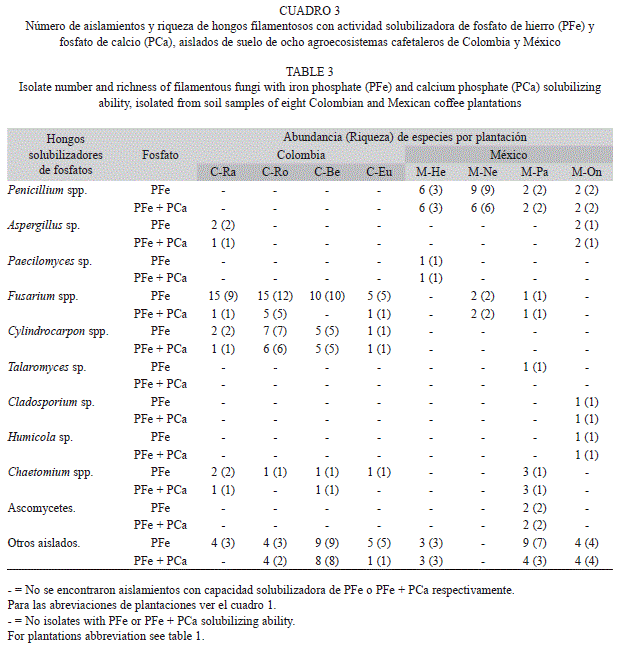
HMF y HSP aislados en las zonas cafetaleras: Se aisló un total de 849 hongos filamentosos (438 para Colombia y 411 para México), de éstos, 137 (89 para Colombia y 48 para México) solubilizaron PFe y 76 (36 para Colombia y 40 para México) solubilizaron PFe y PCa. Aislamientos de los géneros Chaetomium,Aspergillus y Fusarium mostraron solubilización de PFe y PCa en ambas regiones geográficas. Entre los aislamientos, Fusarium spp. y Cylindrocarpon spp. en Colombia y Penicillium spp. en México, fueron los géneros que más contribuyeron a la S y el Nai de HSPFe y HSP-(Fe+Ca) (Cuadro 3).
Distribución de los puntos de muestreo de acuerdo a sus características físicoquímicas y al IMPC: Los tres principales componentes (ejes 1 a 3) explicaron el 77.4% de la varianza total de los datos. El primer eje explicó el 36.7% de esta variabilidad y las variables que más contribuyeron en orden descendente fueron el carbono orgánico (CO, 0.41)>arcilla (-0.37)>AEA1.4 (-0.35)>AEA3.4 (0.33)>AEA0.1 (-0.33)>arena (0.32). Por su parte el segundo eje explicó el 29.0% de la variabilidad y las variables que más contribuyeron en orden descendente fueron AEA0.7 (0.44)>AEA0.4 (0.44)>AEA3.4 (-0.38)>pH (0.33). El tercer eje explicó el 11.7% de la variabilidad y las variables que más contribuyeron en orden descendente fueron IMPC (-0.68)>P (-0.58)>AEA0.1 (0.31)>CO (0.15)>pH (-0.14). Solamente el tercer eje fue explicado mayoritariamente por las variables físico-químicas y la intensidad de manejo. Adicionalmente la proyección cartesiana de los puntos (Fig. 2) mostró dos grupos separados y correspondientes a los países de procedencia de las muestras: México con alta influencia de la arcilla y AEA3.4, mientras Colombia con alta influencia de CO, arena, pH y AEA3.4. Por lo tanto se decidió analizar estos grupos por cada país.
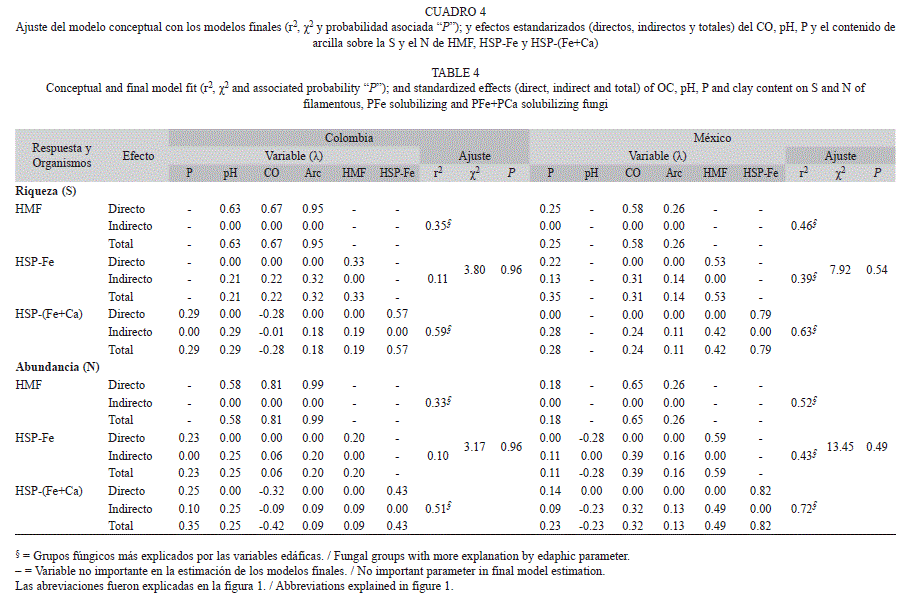
De las variables químicas evaluadas, las que se asociaron con la S y el Nai de HMF en el caso de Colombia fueron el CO y el pH, este último no se relacionó directamente con los HSP-Fe ni HSP-(Fe+Ca); la S y el Nai de HSP-(Fe+Ca) se relacionaron directamente con el CO y el P y, solo indirectamente con el pH. En el caso de México el CO y el P se asociaron directamente con la S y el Nai de HMF; el Nai de HSP-Fe se relacionó directamente con el pH y el Nai de HSP-(Fe+Ca) con el P. La S de HSP-Fe se asoció directamente con el P (Cuadro 4).
Indistinto de la ubicación geográfica del cafetal, el fósforo disponible (P) mostró una relación positiva con la S y el Nai de HSP-(Fe+Ca); sólo en las condiciones más ácidas de los suelos de cafetales mexicanos, se asoció por igual a todos los grupos fúngicos evaluados (r2=0.25-0.35) para la S y (r2=0.11-0.23) para el Nai (Cuadro 4). En agroecosistemas cafetaleros colombianos el pH correlacionó positivamente con la S y el Nai de los todos los grupos fúngicos evaluados, mientras en los mexicanos solo se relacionó y negativamente con el Nai de HSP. El CO se asoció en forma positiva con la S y el Nai de todos los grupos fúngicos, excepto en cafetales colombianos donde la relación fue negativa con a los HSP-(Fe+Ca) (Cuadro 4).
La arcilla se relacionó positivamente con la S y el Nai de los grupos fúngicos evaluados, fue menor su asociación con los HSP, lo cual se observó en la magnitud decreciente de relación total (Cuadro 4). En cafetales colombianos la relación de la arcilla fue mayor (λ). En los casos donde el modelo tuvo mayor poder explicativo (r2>0.2), la S y el N de las poblaciones de proveniencia (HMF para HSPFe y HSP-Fe para HSP-(Fe+Ca)), tuvieron una importante contribución a la explicación del modelo (Cuadro 4).
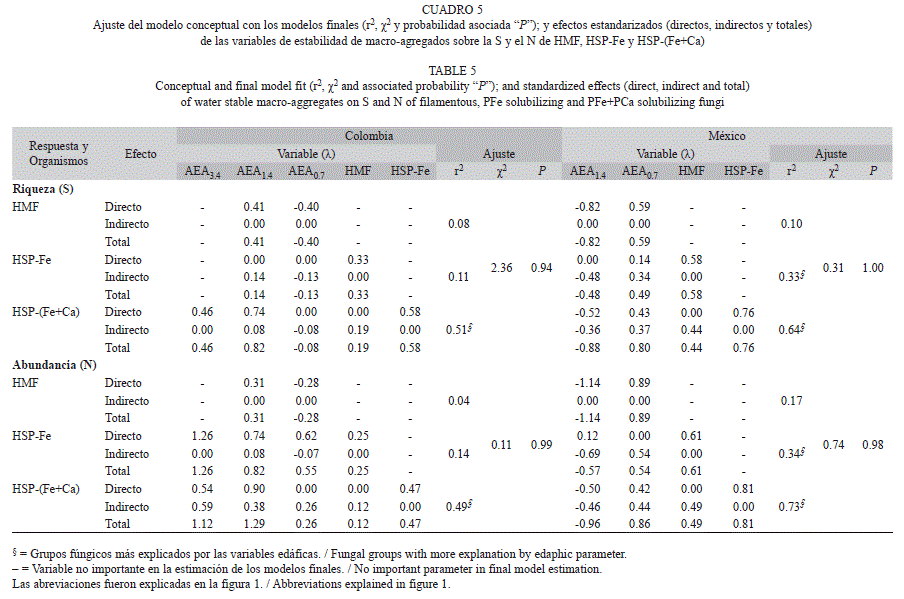
Relaciones de los macro-agregados estables: De las variables de estabilidad de agregados evaluados, en cafetales colombianos las que se asociaron con la S y el Nai de HMF fueron las fracciones AEA1.4 y AEA0.7, las que se relacionaron directamente con la S y el Nai de HSP-(Fe+Ca) fueron AEA3.4 y AEA0.7, mientras que el Nai de HSP-Fe se asoció con las tres fracciones evaluadas. En el caso de los cafetales mexicanos la fracción AEA3.4 no se relacionó con la S y el Nai de los grupos fúngicos evaluados, mientras AEA1.4 y AEA0.7 se asociaron directamente con la S y el Nai de los tres grupos fúngicos evaluados, excepto por AEA0.7, el cual no se asoció directamente con la S de HSP-Fe (Cuadro 5).
Los macro-agregados se asociaron con la S y el Nai de los HSP-(Fe+Ca) (r2>0.4, λ>0.1) en cafetales de ambos países, y con los HSPFe (r2>0.3, λ>0.4) en cafetales mexicanos. En cafetales colombianos la relación de los macroagregados fue positiva; mientras en cafetales mexicanos, la fracción AEA1.4 tuvo una relación negativa y la fracción AEA0.7 positiva sobre los HSP (Cuadro 5). En los casos donde el modelo tiene mayor poder explicativo (r2>0.2), la S y el Nai de las poblaciones de proveniencia (HMF para HSP-Fe y HSP-Fe para HSP-(Fe+Ca)) tuvieron una relación positiva.
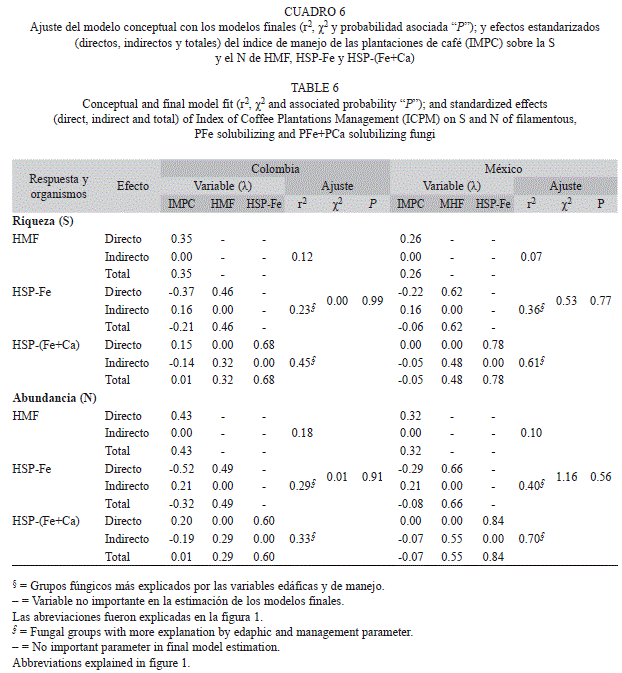
Relaciones de la intensidad de manejo: Todas las relaciones propuestas entre la S y el Nai de HMF, HSP-Fe, HSP-(Fe+Ca) y la intensidad de manejo en cafetales, se confirmaron en ambas regiones geográficas, excepto con los HSP-(Fe+Ca) en condiciones de cafetales mexicanos. La intensidad de manejo (IMPC), tuvo una relación negativa con la S y el Nai de HSP-Fe solo en condiciones de cafetales colombianos. Los modelos mostraron una fuerte relación entre los grupos fúngicos estudiados, mientras la relación de éstos con el IMPC fue menor (Cuadro 6).
Discusión
Dos sistemas cafetaleros edáficamente diferentes: Colombia con pH (5.3-5.7), P disponible (4-62mg/kg), CO (7.7-15.2%) y arena (47.7-62.2%); México con pH (3.8-4.7), P disponible (14-104mg/kg), CO (3.8-5.3%), arcilla (33.4-49.2%) y con estaciones climáticas definidas, ejemplifican cambios en la S y el Nai de HMF y HSP ante cambios del entorno. Se corroboró que en cada agroecosistema, distintas variables físicas, químicas y biológicas se relacionaron directa e indirectamente con la micobiota, lo que hizo poco generalizable la relación con un solo factor edáfico.
Variables que han sido registradas en distintas investigaciones como determinantes de las poblaciones microbianas, apoyan este trabajo. Por ejemplo, los contenidos de arena y arcilla (Zhang et al. 2007), el pH (Bååth 1996, Aciego & Brookes 2009) y el carbono orgánico (Huang et al. 2005, Vaidya et al. 2008), han mostrado una fuerte influencia en las poblaciones microbianas y, en los cafetales evaluados se encontraron como variables edáficas relacionadas con las comunidades fúngicas analizadas. El hecho que los grupos formados en el ACP correspondieran a plantaciones geográficamente cercanas y que a su vez presentaran ambientes edáficos diferentes, indicó la necesidad de discriminar el análisis de las comunidades de HMF, HSP-Fe y HSP-(Fe+Ca) por país.
El carbono orgánico se relacionó positivamente con los HMF en los cafetales analizados, posiblemente debido a su efecto estimulante del crecimiento microbiano (Vaidya et al. 2008) y al suministro de carbono necesario para el crecimiento. Ambientes nutricionalmente no limitados por fuentes de carbono como los cafetales evaluados, permiten el crecimiento de los microorganismos sin competencia inter o intra-específica (Bonilla 2004, May & McLean 2007) por este recurso, hecho que permite la expresión de un mayor número de especies fúngicas (S) y posibilita la presencia de mayores poblaciones funcionalmente activas (Nai).
La evaluación simultánea de diferentes comunidades fúngicas en diversas condiciones edáficas, permitió encontrar similitudes y diferencias en las relaciones de los HSP-(Fe+Ca). En cafetales colombianos, los HSP-(Fe+Ca) tuvieron una relación negativa con el CO, Narsian & Patel (2009) también reportan una relación negativa de los HSP con el CO, explicada por un bajo potencial de los HSP para solubilizar los complejos fosfatados orgánicos. En el caso de los cafetales colombianos, Fusarium spp. y Cylindrocarpon spp. estuvieron presentes en el suelo, son solubilizadores de PCa y pueden aprovechar diversas fuentes de P (Domsch et al. 1980); es posible que los aislamientos tuvieran una desventaja competitiva y fueran excluidos por otros aislamientos con mayor velocidad de crecimiento, en especial en suelos con altos contenidos de P disponible proveniente de fuentes orgánicas y pH moderadamente ácido. Esta relación fue menos notoria en los suelos de cafetales mexicanos posiblemente debido a las menores concentraciones de CO, pH más bajo y a que las bajas temperaturas del invierno hacen más lento el proceso de descomposición y sucesión (Henriksen & Breland 1999, Pietikäinen et al. 2005).
El pH además de influir en la selección de las especies fúngicas predominantes (Dix & Webster 1995, Hujslova et al. 2010); junto a la materia orgánica y las formas de fosfato en el suelo, se relacionó de diferente forma con la S y el Nai de los hongos. La mayoría de los cafetales colombianos, incluidos los analizados, se asientan en suelos con pH>4.9, alta disponibilidad de materia orgánica y alto contenido de arena, en ellos el fósforo predominante proviene del PCa (Fassbender & Bornemisza 1987) y de la materia orgánica (Muleta 2007). El pH edáfico se asoció positivamente con la S y el Nai de HMF, mientras las relaciones fueron menos pronunciadas con la S de HSP. Resultados similares fueron encontrados con hongos saprobios por Aliasgharzad et al. (2010) en pasturas con alto contenido de arena y pH entre 5-8, sin considerar el contenido de P en el suelo. También con HSP-Ca en suelos básicos, con un pH óptimo entre 7.4-8.4 (Narsian & Patel 2009).
En suelos con pH<4.9, contenidos medios de materia orgánica y con alto contenido de arcilla como los caracterizados en los cafetales mexicanos estudiados, predominan los fosfatos de hierro y aluminio (Hemwall 1957), solamente solubilizables por algunas especies fúngicas. Esto hace que reducciones en el pH puedan favorecer el Nai de HSP-Fe y HSP-(Fe+Ca).
Es ampliamente aceptado que la disponibilidad de fósforo en el suelo, depende de las fuentes orgánicas e inorgánicas existentes, producto de la descomposición de la materia orgánica (enmienda o natural), la fertilización fosfatada o la solubilización biológica a partir de formas minerales. Todos los microorganismos y las plantas utilizan el fósforo disponible en el suelo y, en condiciones de alta disponibilidad – como los cafetales mexicanos evaluados – se incrementa su inmovilización biológica a través del desarrollo de poblaciones microbianas, incluidas las fúngicas (Rooney & Clipson 2009). Al superarse la limitante de disponibilidad de fósforo, las especies pueden crecer sin competir por este recurso, permitiendo el incremento de la S fúngica.
Los suelos de los agroecosistemas cafetaleros colombianos presentaron contenidos de carbono orgánico (7.7-15.2%), pH (5.3-5.7) y contenidos de P disponible (4-62mg/kg), que en algunos casos llegan a ser deficientes: <5mg/kg (Codazzi 1998, Portilla 2001). Bajo estas condiciones se espera que la materia orgánica provea de P a través de la mineralización y que la fracción mineral, compleja con iones Ca2+, Fe2+ y Al2+ también aporte P, aunque en menor medida, a través de la solubilización (Alexander 1978, Fassbender & Bornemisza 1987). Los HSP-Ca pueden solubilizar permanentemente el PCa contenido en suelos como los caracterizados, motivo por el cual las poblaciones fúngicas que trabajan sobre esta fracción mineral pueden estar asociadas positivamente con la presencia de P. Estos resultados amplían y complementan los alcances del trabajo con HSP-Ca de Narsian & Patel (2009), ya que ellos encontraron resultados similares, realizando sus evaluaciones en suelos básicos.
En relación con los agregados estables al agua, aunque los macro-agregados se encuentran vinculados con todos los grupos fúngicos evaluados, sólo se asociaron ampliamente (r2>0.3 y λ>0.1) a la S y el Nai de los HSP-Fe y HSP-(Fe+Ca). Según el trabajo de Lupwayi (2001) con suelos limosos y franco limosos cultivados con cebada, y de Wang et al. (2010) con comunidades fúngicas, se esperaría una mayor S de micromicetos saprobios con el aumento de agregados de mayor tamaño. En el presente trabajo al parecer la ausencia de arado, acompañado de un alto porcentaje de agregados superiores a 0.84mm en las muestras, modifica este patrón. En las condiciones de los cafetales colombianos evaluados, los valores de materia orgánica además de asegurar el abastecimiento de P orgánico, también garantizan la solubilización permanente a partir de PCa. La relación positiva entre los macro-agregados y los HSP-(Fe+Ca), puede estar asociada con el Ca2+ presente en todas las fracciones; ya que el catión Ca2+ se ha referenciado como participante en la estabilidad estructural permanente de los suelos (Tisdall 1991, Torres et al. 2007). En los cafetales mexicanos, la relación de los HSP-Fe y HSP-(Fe+Ca) con los agregados entre 0.8- 2mm fue negativa y con los agregados entre 0.5-0.8mm fue positiva; este hecho puede estar relacionado con variables no evaluadas, relacionadas directa o indirectamente con cada una de las fracciones como: los tipos de arcilla presentes, los cuales junto con los contenidos de arcilla pueden regular los contenidos de P (Fontes & Weed 1996, Hinsinger 2001); las comunidades bacterianas potencialmente solubilizadoras de fosfatos; la estructura del carbono orgánico contenido en los agregados (Six et al. 2001, 2002); la oxigenación y porosidad, los cuales pueden afectar las propiedades físicas y químicas de las diferentes fracciones de agregados (Schjonning et al. 2002, Six et al. 2004).
Los hongos filamentosos solubilizadores de fosfatos aislados: Fusarium spp., Penicillium spp., Cylindrocarpon spp. y Chaetomium spp. se encuentran ampliamente distribuidos en los suelos del mundo (Domsch et al. 1980), y aunque la presencia de hifas se ha documentado como elemento temporal importante en la formación y mantenimiento estructural del suelo (Tisdall 1991, 1994); los datos del presente estudio no permitieron hacer afirmaciones específicas con respecto a la participación de los géneros y especies fúngicas encontradas en relación con la agregación del suelo. La información obtenida en el presente trabajo permite confirmar las observaciones de Dix & Webster (1995) respecto a la relación del pH edáfico y el predominio de géneros o especies fúngicas así: en cafetales colombianos con pH>4.9 y la presencia de Fusarium spp. y Cylindrocarpon spp., y en cafetales mexicanos con pH<4.9 y la presencia de Penicillium spp.
Contrario a lo que otras investigaciones han señalado (Simmons & Coleman 2008, Suzuki et al. 2009, Beauregard et al. 2010), respecto a la influencia de los sistemas de fertilización, tipos de suelo y sistemas de labranza sobre las comunidades fúngicas, nosotros solo encontramos relación entre la intensidad de manejo de los cafetales y la S y el Nai de HSPFe, pero no con la S y el Nai de HSP-(Fe+Ca) o HMF. Entre las razones que pueden explicar que el IMPC no contribuyera ampliamente a la S y el Nai fúngicos en la presente investigación, se encuentran: a) el uso de enmiendas orgánicas solas (Girvan et al. 2004, Vaidya et al. 2008) o en combinaciones específicas con enmiendas de síntesis industrial (He et al. 2007, Wenhui et al. 2010), son prácticas que han mostrado efecto positivo sobre las comunidades microbianas, pero en la estimación del IMPC no se incluye la información de productos específicos, dosis ni tiempo de aplicación. b) el carácter aditivo del IMPC no diferencia enmiendas de síntesis industrial u orgánica en el resultado final (Hernández-Martínez et al. 2009), de tal manera que valores cercanos de IMPC pueden corresponder tanto a manejo convencional como alternativo. c) el control de plagas (PC y PA) no siempre va dirigido al suelo y principalmente las prácticas asociadas al suelo son las que afectan directamente a las comunidades edáficas de HMF (Beauregard et al. 2010, Wang et al. 2010) y, d) las condiciones edáficas bajo los primeros 5cm de un suelo sin arado, donde fueron tomadas las muestras, suponen condiciones más estables que en la parte superior del mismo, por lo tanto un menorefecto de la actividad agrícola superficial.
Los resultados mencionados son la base para continuar con esta línea de investigación ya que el empleo de HSF en suelos ácidos como los cafetales estudiados, es una mejor opción que el uso de bacterias, por su afinidad para desarrollarse bajo condiciones de acidez en contraposición a las bacterias que prefieren ambientes alcalinos. Queda por investigar en detalle si los cambios en la S de los grupos fúngicos estudiados (HMF, HSP-Fe y HSP- (Fe+Ca)) son importantes para la productividad del cultivo de café a nivel regional. Así mismo, se requiere de la evaluación de las interacciones entre aislamientos de HSP y de hongos de micorriza arbuscular (HMA), las cuales en algunas investigaciones han mostrado una respuesta sinérgica, promoviendo el crecimiento de las plantas gracias al mejor aprovechamiento del P (Osorio & Habte 2001, Zhang et al. 2011). No obstante, en otros trabajos no se han encontrado respuestas positivas (Cabello et al. 2005), lo que muestra la importancia de continuar con investigaciones básicas que profundicen en forma integral con los factores bióticos y abióticos involucrados con la diferentes comunidades fúngicas del suelo.
Las relaciones obtenidas entre las variables edáficas y la intensidad de manejo con los grupos fúngicos evaluados deben ser evaluadas también en época de lluvias para confirmar su permanencia y estabilidad. Las relaciones pueden cambiar radicalmente en cuanto la humedad propicia el crecimiento fúngico (aumentando el número de aislamientos) en comparación a la época de secas, hecho que también puede propiciar la expresión de un mayor número de especies fúngicas, afectando las relaciones edáficas con la S.
Las relaciones del carbono orgánico, el fósforo disponible y el pH con la S y el Nai de los grupos fúngicos estudiados fueron altamente variables, lo cual impide realizar cualquier tipo de generalizaciones de asociación, bien sea al evaluar grupos fúngicos funcionalmente diferentes o regiones geográficas.
La intensidad del manejo agrícola no afectó ni la S ni el Nai de los HMF en ninguna de las regiones cafetaleras estudiadas; para los HSP no se encontraron patrones en la relación entre la intensidad de manejo de los cafetales, la S y el Nai de HSP.
En los agroecosistemas cafetaleros evaluados, las diferentes fracciones de macroagregados no muestran un patrón definido en relación con la S ni con el Nai de HMF y HSP.
El efecto de todos y cada uno de las variables edáficas sobre la micobiota no puede ser explicado alejado de su contexto, lo que hace indispensable que se tenga en cuenta las particularidades de las propiedades edáficas en cada zona.
Agradecimientos
Los autores expresan sus agradecimientos a Nubia Rodríguez Vargas, Fernando Ramos Naranjo y Felipe Vergara por su colaboración en actividades de laboratorio en Colombia. Agradecemos a la empresa Química Barquim S.A. por la donación del fosfato de hierro. Asimismo agradecemos a la OEA y al CONACyT por el apoyo económico que soportaron la beca de sostenimiento del primer autor y al Instituto de Ecología A.C. por su apoyo logístico y económico. Un especial agradecimiento a los lectores anónimos, por sus aportes en el mejoramiento del documento.
Referencias
Aciego, J.C. & P.C. Brookes. 2009. Substrate inputs and pH as factors controlling microbial biomass, activity and community structure in an arable soil. Soil Biol. Biochem. 41: 1396-1405. [ Links ]
Alexander, M. 1978. Introduction to soil microbiology. Wiley, New York, EEUU. [ Links ]
Aliasgharzad, N., L.M. Martensson & P.A. Olsson. 2010. Acidification of a sandy grassland favours bacteria and disfavours fungal saprotrophs as estimated by fatty acid profiling. Soil Biol. Biochem. 42: 1058-1064. [ Links ]
Andrade, S.A.L., P. Mazzafera, M.A. Schiavinato & A.P.D. Silveira. 2009. Arbuscular mycorrhizal association in coffee. J. Agric. Sci. 147: 105-115. [ Links ]
Antoninka, A., J.E. Wolf, M. Bowker, A.T. Classen & N.C. Johnson. 2009. Linking above- and belowground responses to global change at community and ecosystem scales. Glob. Chang. Biol. 15: 914-929. [ Links ]
Arbuckle, J.L. 2010. Amos 18 User’s Guide. SPSS, Chicago, EEUU. [ Links ]
Arcila, P.J., F.P. Farfán, A.B. Moreno, L.F. Salazar & E. Hincapie. 2007. Sistemas de producción de café en Colombia. Cenicafé y Federación Nacional de Cafeteros de Colombia, Chinchiná, Colombia. [ Links ]
Ashman, M., P.D. Hallett & P.C. Brookes. 2003. Are the links between soil aggregate size class, soil organic matter and respiration rate artifacts of the fractionationprocedure? Soil Biol. Biochem. 35: 435-444. [ Links ]
Bååth, E. 1996. Adaptation of soil bacterial communities to prevailing pH in different soils. FEMS Microbiol. Ecol. 19: 227-237. [ Links ]
Beauregard, M.S., C. Hamel, A. Nayyar & M. St-Arnaud. 2010. Long-term phosphorus fertilization impacts soil fungal and bacterial diversity but not AM fungal community in Alfalfa. Microb. Ecol. 59: 379-389. [ Links ]
Bhattacharya, S. & D.J. Babyaraj. 2002. Effectiveness of arbuscular mycorrhizal fungal isolates on arabica coffee (Coffea arabica L.). Biol. Agric. Hortic. 20: 125-131. [ Links ]
Bills, G.F., M. Christensen, M. Powell & G. Thorn. 2004. Saprobic soil fungi, p. 271-302. In G.M. Mueller, M.S. Foster & G.F (eds.). Bills. Biodiversity of fungi: Inventory and monitoring methods. Elsevier, California, EEUU. [ Links ]
Bollen, K.A. 1989. Structural equations with latent variables. Wiley, New York, EEUU. [ Links ]
Bonilla, M.A. 2004. Ecología de poblaciones. Notas de clase. Universidad Nacional de Colombia, UNIBIBLOS, Bogotá, Colombia. [ Links ]
Bossuyt, H., J. Six & P.F. Hendrix. 2002. Aggregate-protected carbon in no-tillage and conventional tillage agroecosystems using carbon-14 labeled plant residue. Soil Sci. Soc. Am. J. 66: 1965-1973. [ Links ]
Bouyoucos, G.J. 1962. Hydrometer method improved for making particle size analyses of soils. Agron. J. 54: 464-465. [ Links ]
Byrne, B.M. 1989. A primer of LISREL: Basic applications and programming for confirmatory factor analytic models. Springer-Verlag, New York, EEUU. [ Links ]
Cabello, M., G. Irrazabal, A.M. Bucsinszky, M. Saparrat & S. Schlamuk. 2005. Effect of an arbuscular mycorrhizal fungus, Glomus mosseae, and a rockphosphate-solubilizing fungus, Penicillium thomii, on Mentha piperita growth in a soilless medium. J. Basic Microbiol. 45: 182-189. [ Links ]
Cardoso, I.M. & T.W. Kuyper. 2006. Mycorrhizas and tropical soil fertility. Agric. Ecosyst. Environ. 116: 72-84. [ Links ]
Codazzi, A. 1998. Consideraciones generales para interpretar análisis de suelos. Instituto Geográfico Agustín Codazzi, Bogotá, Colombia. [ Links ]
Codazzi, A. 2009. Estudio general de suelos y zonificación de tierras. Departamento de Risaralda. Instituto Geográfico Agustín Codazzi, Bogotá, Colombia. [ Links ]
Daraghmeh, O.A., J.R. Jensen & C.T. Petersen. 2009. Soil structure stability under conventional and reduced tillage in a sandy loam. Geoderma 150: 64-71. [ Links ]
Dighton, J. 2003. Fungi in Ecosystem Processes. Marcel Dekker, New York, EEUU. [ Links ]
Dix, N.J. & J. Webster. 1995. Fungal Ecology. Chapman and Hall, Londres, Inglaterra. [ Links ]
Domsch, K.H., W. Gams & T.H. Anderson. 1980. Compendium of soil fungi. Academic, San Francisco, EEUU. [ Links ]
Fassbender, H.W. & E. Bornemisza. 1987. Química de suelos con énfasis en América Latina. Instituto Iberoamericano de Cooperación para la Agricultura, San José, Costa Rica. [ Links ]
Fixen, P.E. & J.H. Grove. 1990. Soil testing and plantanalysis, p. 141-180. In R.L. Westerman (ed.). Testing soils for phosphorus. Soil Science Society America Book Number 3, Madison, Wisconsin, EEUU. [ Links ]
Fontes, M.P.F. & S.B. Weed. 1996. Phosphate adsorption by clays from brazilian oxisols: relationships with specific surface area and mineralogy. Geoderma 72: 37-51. [ Links ]
Geissert, D. & A. Ibáñez. 2008. Calidad y ambiente físicoquímico de los suelos, p. 213-222. In R. Manson, V. Hernández-Ortiz, S. Gallina & K. Mehltreter (eds.). Agroecosistemas Cafetaleros de Veracruz: Biodiversidad, manejo y conservación. Instituto de Ecología A.C., Xalapa, México. [ Links ]
Girvan, M.S., J. Bullimore, A.S. Ball, J.N. Pretty & A.M. Osborn. 2004. Responses of active bacterial and fungal communities in soils under winter wheat to different fertilizer and pesticide regimens. Appl. Env. Microbiol. 70: 2692-2701. [ Links ]
Grace, J.B. & B.H. Pugesek. 1998. On the use of path analysis and related procedures for the investigation of ecological problem. Am. Nat. 152: 151-159. [ Links ]
Grace, J.B. 2006. Structural equation modeling and natural systems. Universidad de Cambridge, New York, EEUU. [ Links ]
Grace, J.B. & J.E. Keeley. 2006. A structural equation model analysis of postfire plant diversity in California Shrublands. Ecol. Appl. 16: 503-514. [ Links ]
Griffin, D.H. 1996. Fungal physiology. Wiley, New York, EEUU. [ Links ]
Guzmán, O. & A. Jaramillo. 1989. Estudio climático de Risaralda y Quindío. Bol. Téc. Cenicafé 15: 1-64. [ Links ]
He, J.Z., J.P. Shen, L.M. Zhang, Y.G. Zhu, Y.M. Zheng, M.G. Xu & H. Di. 2007. Quantitative analyses of the abundance and composition of ammonia-oxidizing bacteria and ammonia-oxidizing archaea of a Chinese upland red soil under long-term fertilization practices. Environ. Microbiol. 9: 2364-2374. [ Links ]
Hemwall, J.B. 1957. The fixation of phosphorus by soils, p. 95-113. In A.G. Norman (ed.). Advances in Agronomy IX. Academic, New York, EEUU. [ Links ]
Henriksen, T.M. & T.A. Breland. 1999. Decomposition of crop residues in the field: evaluation of a simulation model developed from microcosm studies. Soil Biol. Biochem. 31: 1423-1434. [ Links ]
Hernández-Martínez, G. 2008. Clasificación Agroecológica, p. 15-34. In R.H. Manson, V. Hernández-Ortiz, S. Gallina & K. Mehltreter (eds.). Agroecosistemas cafetaleros de Veracruz: Biodiversidad, manejo y conservación. Instituto de Ecología A.C., Xalapa, México. [ Links ]
Hernández-Martínez, G., R.H. Manson & A. Contreras. 2009. Quantitative classification of coffee agroecosystems spanning a range of production intensities in central Veracruz, México. Agric. Ecosyst. Environ. 134: 89-98. [ Links ]
Herrera-Peraza, R.A., C. Hamel, F. Fernández, R.L. Ferrer & E. Furrazola. 2011. Soil-strain compatibility: the key to effective use of arbuscular mycorrhizal inoculants? Mycorrhiza 21: 183-93. [ Links ]
Hinsinger, P. 2001. Bioavailability of soil inorganic P in the rhizosphere as affected by root-induced chemical changes: a review. Plant Soil 237: 173-195. [ Links ]
Huang, P., M. Wang & C. Chiu. 2005. Soil mineral-organic matter-microbe interactions: Impacts on biogeochemical processes and biodiversity in soils. Pedobiology 49: 609-635. [ Links ]
Hujslova, M., A. Kubatova, M. Chudickova & M. Kolarik. 2010. Diversity of fungal communities in saline and acidic soils in the Soos National Natural Reserve, Czech Republic. Micol. Prog. 9: 1-15. [ Links ]
ICO. 2011. Total production of exporting countries: crop years commencing 2005 to 2010. International Coffee Organization, Londres, Inglaterra. [ Links ]
Lauber, C.L., M.S. Strickland, M.A. Bradford & N. Fierer. 2008. The influence of soil properties on the structure of bacterial and fungal communities across land-use types. Soil Biol. Biochem. 40: 2407-2415. [ Links ]
Lupwayi, N. 2001. Bacterial diversity in water-stableaggregates of soils under conventional and zero tillage management. Appl. Soil Ecol. 16: 251-261. [ Links ]
Manns, H.R., C.D. Maxwell & R.J.N. Emery. 2007. The effect of ground cover or initial organic carbon on soil fungi, aggregation, moisture and organic carbon in one season with oat (Avena sativa) plots. Soil Tillage Res. 96: 83-94. [ Links ]
May, R. & A. McLean. 2007. Theoretical ecology: principles and applications. Oxford University, New York, EEUU. [ Links ]
McCune, B. & J.B. Grace. 2002. Analysis of Ecological Communities. MjM Software Design, Gleneden, EEUU. [ Links ]
Montiel, R.B. & M.J.D. Robledo. 1998. Caracterización, descripción y evaluación de especies frutales en el banco genético del CRUO. Universidad Veracruzana, Córdoba, Veracruz, México. [ Links ]
Muleta, D. 2007. Microbial Inputs in coffee (Coffea Arabica L .) Production Systems, Southwestern Ethiopia. Production. Tesis de Doctorado, Universidad Sueca de Ciencias Agrícolas, Upsala, Suecia. [ Links ]
Márquez, C.O., V.J. García, C.A. Cambardella, R.C. Schultz & T.M. Isenhart. 2004. Aggregate size stability distribution and soil stability. Soil Sci. Soc. Am. J. 68: 725-735. [ Links ]
Narsian, V.T. & H.H. Patel. 2009. Relationship of physicochemical properties of rhizosphere soils with native population of mineral phosphate solubilizing fungi. Indian J. Microbiol. 49: 60-67. [ Links ]
Oades, J.M. & A.G. Waters. 1991. Aggregate hierarchy in soils. Aust. J. Soil Res. 29: 815-828. [ Links ]
Oliveira, C.A., V.M.C. Alves, I.E. Marriel, E.A. Gomes, M.R. Scotti, N.P. Carneiro, C.T. Guimarães, R.E. Schaffert & N.M.H. Sá. 2009. Phosphate solubilizing microorganisms isolated from rhizosphere of maize cultivated in an oxisol of the Brazilian Cerrado Biome. Soil Biol. Biochem. 41: 1782-1787. [ Links ]
Osorio, N.W. & M. Habte. 2001. Synergistic influence of an arbuscular mycorrhizal fungus and a P solubilizing fungus on growth and P uptake of Leucaena leucocephala in an Oxisol. Arid Land Res. Manag. 15: 263-274. [ Links ]
Pietikäinen, J., M. Pettersson & E. Bååth. 2005. Comparison of temperature effects on soil respiration and bacterial and fungal growth rates. FEMS Microbiol. Ecol. 52: 49-58. [ Links ]
Portilla, N. 2001. Cuadernillo de procedimientos analíticos de suelos. Departamento de investigación y diagnóstico regional. Laboratorio de suelos, aguas y plantas. Texcoco, México. [ Links ]
Posada, R.H., G. Heredia-Abarca, E. Sieverding & M. Sánchez de Prager. 2012. Solubilization of iron and calcium phosphates by soil fungi isolated from coffee plantations. Arch. Agron. Soil Sci. (in press). DOI: 10.1080/03650340.2011.610030. [ Links ]
Rodríguez, J.L. 2001. Efecto del biofertilizante Mycoral® (micorriza arbuscular) en el desarrollo del café (Coffea arabica L), en vivero de Zamorano, Honduras. Tesis de Licenciatura, Universidad de Zamorano, Tegucigalpa, Honduras. [ Links ]
Rooney, D.C. & N.J.W. Clipson. 2009. Phosphate addition and plant species alters microbial community structure in acidic upland grassland soil. Microb. Ecol. 57: 4-13. [ Links ]
Schjonning, P., L.J. Munkholm, P. Moldrup & O.H. Jacobsen. 2002. Modeling soil pore characteristics from measurements of air exchange: the long-term effects of fertilization and crop rotation. Eur. J. Soil Sci. 53: 331-339. [ Links ]
Shipley, B. 2000. Cause and correlation in biology: A user’s guide to path analysis, structural equations and causal inference. Cambridge University, Londres, Inglaterra. [ Links ]
Simmons, B. & D.C. Coleman. 2008. Microbial community response to transition from conventional to conservation tillage in cotton fields. Appl. Soil Ecol. 40: 518-528. [ Links ]
Six, J., G. Guggenberger, K. Paustian, L. Haumaier, E.T. Elliot & W. Zech. 2001. Sources and composition of soil organic matter fractions between and within soil aggregates. Eur. J. Soil Sci. 52: 607-618. [ Links ]
Six, J., R.T. Conant, E.A. Paul & K. Paustian. 2002. Stabilization mechanisms of soil organic matter: Implications for C-saturation of soils. Plant Soil 241: 155-176. [ Links ]
Six, J., H. Bossuyt, S. Degryze & K. Denef. 2004. A history of research on the link between (micro) aggregates, soil biota, and soil organic matter dynamics. Soil Tillage Res. 79: 7-31. [ Links ]
Six, J., S.D. Frey, R.K. Thiet & K.M. Batten. 2006. Bacterial and fungal contributions to carbon sequestration in agroecosystems. Soil Sci. Soc. Am. J. 70: 555-569. [ Links ]
Su, Y.Z., W.J. Liu, R. Yang & X.X. Chang. 2009. Changes in soil aggregate, carbon, and nitrogen storages following the conversion of cropland to alfalfa forage land in the marginal oasis of Northwest China. Environ. Manag. 43: 1061-1070. [ Links ]
Sundara, R. & M. Sinha. 1963. Organisms phosphate solubilizers in soil. Indian J. Agric. Sci. 33: 272-278. [ Links ]
Suzuki, C., K. Nagaoka, A. Shimada & M. Takenaka. 2009. Bacterial communities are more dependent on soil type than fertilizer type, but the reverse is true for fungal communities. Soil Sci. Plant Nutr. 55: 80-90. [ Links ]
Tisdall, J.M. 1991. Fungal hyphae and structural stability of soil. Aust. J. Soil Res. 29: 72-743. [ Links ]
Tisdall, J.M. 1994. Possible role of soil microorganisms in aggregation in soils. Plant Soil 159: 115-121. [ Links ]
Torres, R., E. Barrios & M. Sánchez de Prager. 2007. El papel del micelio externo de HMA asociado a barbechos mejorados en suelos degradados de pescador - Cauca, p. 251-261. In M. Sánchez de Prager (ed.). Las endomicorrizas: Expresión bioedáfica de importancia en el trópico. Universidad Nacional de Colombia, Palmira, Colombia. [ Links ]
Vaidya, G.S., K. Shrestha, B.R. Khadge, N.C. Johnson & H. Wallander. 2008. Organic matter stimulates bacteria and arbuscular mycorrhizal fungi in Bauhinia purpurea and Leucaena diversifolia plantations on eroded slopes in Nepal. Restor. Ecol. 16: 79-87. [ Links ]
Wang, Y., J. Xu, J. Shen, Y. Luo, S. Scheu & X. Ke. 2010. Tillage, residue burning and crop rotation alter soil fungal community and water-stable aggregation in arable fields. Soil Tillage Res. 107: 71-79. [ Links ]
Wenhui, Z., T. Gu, W. Wang, B. Zhang, X. Lin, Q. Huang & W. Shen. 2010. The effects of mineral fertilizer and organic manure on soil microbial community and diversity. Plant Soil 326: 511-522. [ Links ]
Wu, Y., B. Ma, L. Zhou, H. Wang, J. Xu, S. Kemmitt & P.C. Brookes. 2009. Changes in the soil microbial community structure with latitude in eastern China, based on phospholipid fatty acid analysis. Appl. Soil Ecol. 43: 234-240. [ Links ]
Yoder, R.E. 1936. A direct method of aggregate analysis and study of physical nature of erosion losses. J. Am. Soc. Agron. 28: 337-351. [ Links ]
Zhang, P., J. Zheng, G. Pan, X. Zhang, L. Li & T. Rolf. 2007. Changes in microbial community structure and function within particle size fractions of a paddy previous termsoil under different long-term fertilization treatments from the Tai Lake region, China. Colloid. Surf. B: Biointerfaces 58: 264-270. [ Links ]
Zhang, H., X. Wu, G. Li & P. Qin. 2011. Interactions between arbuscular mycorrhizal fungi and phosphatesolubilizing fungus (Mortierella sp.) and their effects on Kostelelzkya virginica growth and enzyme activities of rhizosphere and bulk soils at different salinities. Biol. Fert. Soils 47: 543-554. [ Links ]
*Correspondencia:
Raúl Hernando Posada: Instituto de Ecología, A. C. Carretera antigua a Coatepec 351, El Haya, Xalapa 91070, Veracruz, México. raulposada@hotmail.com.
Marina Sánchez de Prager: Universidad Nacional de Colombia, sede Palmira. Carrera 32 # 12 - 00 Chapinero, Vía Candelaria, Palmira, Valle del Cauca, Colombia. msanchezpr@unal.edu.co.
Ewald Sieverding: University of Hohenheim, Institute of Plant Production and Agroecology in the Tropics and Subtropics, Garbenstr. 13, Stuttgart Hohenheim, Germany. sieverdinge@aol.com.
Karla Aguilar Dorantes: Instituto de Ecología, A. C. Carretera antigua a Coatepec 351, El Haya, Xalapa 91070, Veracruz, México. karlachic@gmail.com.
Gabriela Patricia Heredia-Abarca: Instituto de Ecología, A. C. Carretera antigua a Coatepec 351, El Haya, Xalapa 91070, Veracruz, México. gabriela.heredia@inecol.edu.mx.
1. Instituto de Ecología, A. C. Carretera antigua a Coatepec 351, El Haya, Xalapa 91070, Veracruz, México; raulposada@hotmail.com, karlachic@gmail.com, gabriela.heredia@inecol.edu.mx.
2. Universidad Nacional de Colombia, sede Palmira. Carrera 32 # 12 - 00 Chapinero, Vía Candelaria, Palmira, Valle del Cauca, Colombia; msanchezpr@unal.edu.co.
3. University of Hohenheim, Institute of Plant Production and Agroecology in the Tropics and Subtropics, Garbenstr. 13, Stuttgart Hohenheim, Germany; sieverdinge@aol.com.
Recibido 26-VII-2011. Corregido 30-II-2012. Aceptado 30-III-2012.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}