










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.60 n.3 San José Sep. 2012
Differential effects of land use on ant and herbivore insect communities associated with Caryocar brasiliense (Caryocaraceae)
Frederico S. Neves1*,2*, Rodrigo F. Braga1, Lucimar S. Araújo1, Ricardo I. Campos3* & Marcílio Fagundes1
*Dirección para correspondencia
Abstract
Simplification of natural habitats leads to a modification of the community associated with a host plant. Pequi trees (Caryocar brasiliense) are common to find in central Brazil, especially in the middle of monocultures, such as soy, corn, pasturelands or Eucalyptus plantations. On this scenario we hypothesized that habitat modification differentially affects the diversity of ants and herbivore insects associated with this species. The aim of the work was to test if C. brasiliense trees located in human modified habitats, support a lower species richness and abundance of ants, and a greater species richness and abundance of insect herbivores, compared to preserved cerrado habitats. The study was conducted in a Cerrado area located in Northern Minas Gerais State, Brazil. Ants and herbivore insects were collected monthly during 2005 using beating technique. The results showed that ant species richness was higher in pequi trees located in preserved Cerrado, followed by trees in pastureland and Eucalyptus plantation, respectively. The ant abundance was lower in the Eucalyptus plantation but no difference in ant abundance was observed between trees in pastureland and the preserved Cerrado. Moreover, herbivore insects exhibited lower number of species and individuals in trees located in the preserved Cerrado than in the pastureland and Eucalyptus plantation. We concluded that habitats simplified by human activities may result in diversity loss and may change species interactions.
Key words: environmental complexity, habitat management, insect diversity, pequi trees, tri-trophic interactions.
Resumen
La simplificación de los hábitats naturales conduce a una modificación de la comunidad asociada con una planta huésped. En la región central de Brasil es muy común encontrar árboles aislados de “pequi” (Caryocar brasiliense) en medio de monocultivos de soya, maíz, pastos o plantaciones de eucalipto. Bajo este escenario, nosotros predecimos que la modificación del hábitat afecta diferencialmente la diversidad de hormigas y herbívoros asociados a árboles de “pequi”. El objetivo de este artículo fue determinar que árboles de “pequi” localizados en hábitats modificados por actividades humanas (i.e. pastizales y plantaciones de Eucalyptus), soportan una menor abundancia y riqueza de especies de hormigas. Por el contrario, se espera una mayor abundancia y riqueza de especies de herbívoros en hábitats modificados. Este estudio fue realizado en un área de Cerrado (sabana brasileña) localizado en el Norte del estado de Minas Gerais, Brasil. Las hormigas y los herbívoros fueron muestreados usando técnicas de golpe. La riqueza de especies de hormigas fue mayor en árboles de “pequi” localizados en áreas preservadas de Cerrado, seguida por árboles de pastizales y finalmente por árboles de plantaciones de Eucalyptus. La abundancia de hormigas fue menor en plantaciones de Eucalyptus, pero no hubo diferencias en la abundancia de hormigas entre árboles de pastizales y áreas preservadas de cerrado. Por otra parte, el número de especies de insectos herbívoros fue menor en árboles de áreas preservadas de Cerrado que en pastizales y plantaciones de Eucalyptus. Concluimos que las actividades humanas reducen la complejidad del hábitat resultando en la pérdida de la diversidad de insectos y procesos ecológicos.
Palabras clave: complejidad ambiental, manejo de hábitat, diversidad de insectos, árboles de “pequi”, interacciones tri-tróficas.
Understanding natural patterns of species distribution and mechanisms, which determine these patterns, constitute the basic information for biodiversity conservation (Basset et al. 2003, Samways 2007). In Brazil, studies that describe the patterns of species richness and distribution have primarily focused on the Amazon and Atlantic Forests (Myers et al. 2000). However, studies of this nature in the Brazilian cerrado (savanna) deserve special attention because it is considered a conservation priority biome (Silva & Bates 2002). In fact, the cerrado represent around 1.5 to 2.0 million km2 (approximately 20% of the country’s surface) and less than 50% of cerrado’s original area remains preserved, mainly because of agricultural expansion in central Brazil (Klink & Machado 2005).
Caryocar brasiliense Camb. (Caryocaraceae) is a typical tree of the Brazilian Cerrado and it can be found from the Southern of Pará State, along the edge of the Amazon region, until the South of Paraná State and also in Paraguay (Gribel & Hay 1993). The extraction of pequi fruits is considered an important economic activity in different regions of Brazil, since these fruits are used in preparation of dishes, condiments, oils and sweetened beverages (Almeida & Silva 1994). During the summer, in Northern Minas Gerais State, the pequi fruit collection engages approximately 50% of the population in rural areas, representing 54.7% of the annual income of these people (Fagundes et al. 2007).
The pequi tree is legally protected in Minas Gerais State (Law 10,883 of 02-20-1992), but their natural habitat has been constantly altered by different human land uses such as cattle-raising pasturelands and agroforestry systems. Thus these trees are commonly found isolated in these structurally simpler habitats (Fernandes et al. 2004). However, the pequi phenology change among different habitats (Fagundes et al. 2007) and recently, several herbivorous insects have been observed causing damage to the trees located in pasture (M.F. personal observation). In this system, ants are an important component controlling insect herbivores. In fact, extrafloral nectarines located on the pequi leaves serve as attractors to several ant species which can decrease herbivore abundance (Oliveira 1997).
Besides the direct negative effect of ants on herbivores, habitat structure can modify ant diversity, causing an indirect effect on herbivores (Hoffmann & Andersen 2003, Lassau & Hochuli 2004, Rosumek et al. 2009). Few studies have focused on insect diversity in human modified habitats, including both, Eucalyptus forests and agricultural systems (Szinicz et al. 2005, Tylianakis et al. 2007). In studies about moth species from forest fragments and from different surrounding agricultural habitats, the species richness decreased with increasing distance to the forest fragments. It was also found that the majority of moth species frequently moved between forest and agricultural habitats (Ricketts et al. 2001). Thus, habitat simplification around the pequi trees could decrease ant diversity and, consequently, decreases the pressure of this predator on the insect herbivore fauna associated with the same tree. Based on that, the aim of the present study was to test the hypothesis that pequi trees located in human modified habitats, which are structurally simpler, support a lower species richness and abundance of ants, and a greater species richness and abundance of insect herbivores, compared to more complex preserved cerrado habitats.
Materials and methods
Study area: The study was conducted in the Cerrado biome, in Morro Alto (16°53’17” S - 44°00’06’’ W), a district that belongs to Montes Claros municipality, Northern of Minas Gerais State (Brazil). The climate of the region is considered as tropical semi-arid (Peel et al. 2007), characterized by the existence of a severe dry season during the winter. The average temperature is 21°C and the annual rainfall is approximately 1 100mm, concentrated from November to January. To test the hypothesis, three adjacent habitats (500m), with similar soil characteristics and density of pequi trees, were chosen in the study area. Two habitats have been modified by human activities: cattle grazing pastureland and Eucalyptus plantation; and one preserved area (Cerrado strictu sensu physiognomy). In each habitat 15 trees of similar sizes (5-8m heights and diameter at breast height above 5cm) were marked in order to characterize the herbivore insects, and ant richness and abundance.
Sampling of ants and herbivore insects: The effects of human impact on insect fauna associated with pequi trees were evaluated monthly throughout 2005. Ants and herbivore insects were sampled (08:00-11:00am) using the beating technique (Neves et al. 2010): for each selected plant, a branch was chosen at random (selected branches in small and large trees had approximately the same amount of leaves), and the branch was hit ten times with a beating stick in order to dislodge the associated arthropods on to a tray placed beneath. The insects collected were taken to the laboratory and sorted to the lowest taxonomic level possible, and deposited at the Laboratório de Biologia da Conservação, Universidade Estadual de Montes Claros (Unimontes).
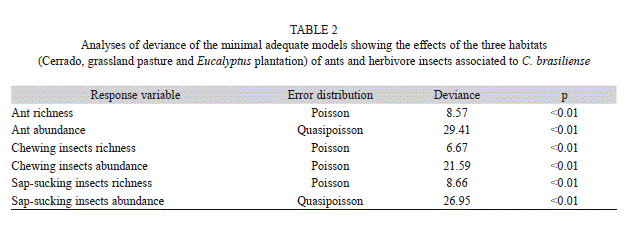
A generalized linear model was used to verify the habitat effect on ant and insect herbivore richness and abundance. The mean number of species or individuals per tree over the 12 months of sampling was used as response variables in order to remove the temporal pseudoreplication. The habitats (preserved Cerrado, pastureland and Eucalyptus plantation) were used as explanatory variables. The minimal model was constructed from the junction of qualitative factors (habitat), which were statistically different from the full model (analysis of contrast) (Crawley 2007). Next, a residual analysis was conducted to check data adequacy for the assumed probability distribution (Crawley 2007). All analysis was performed using the statistical program R (R Development Core Team 2010).
Results
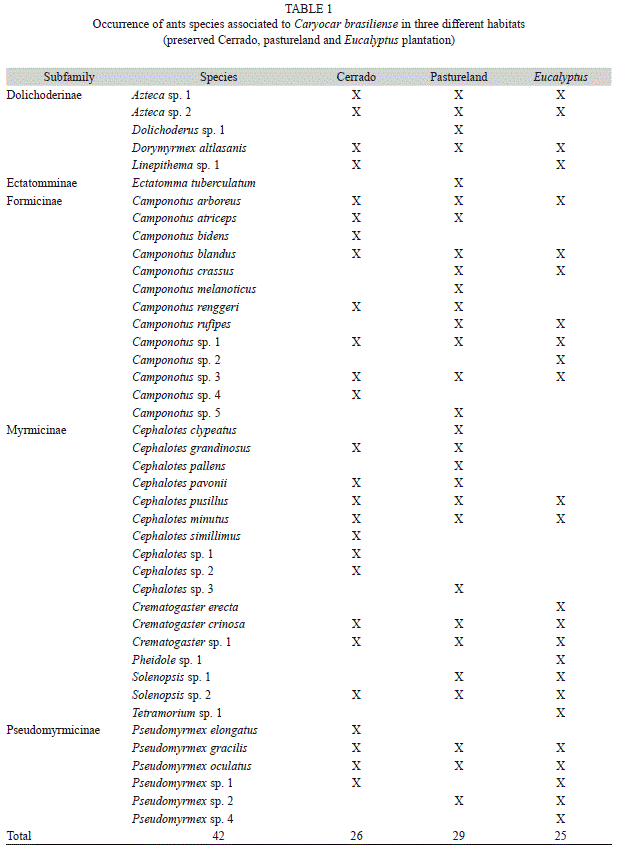
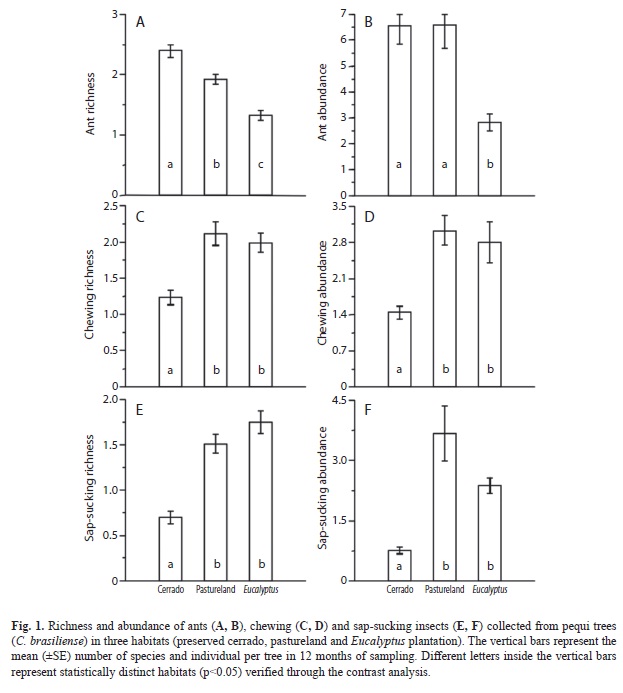
A total of 2 827 ants belonging to 42 species, 12 genera and four subfamilies were collected from C. brasiliense trees (Table 1). The two richest ant genera were Camponotus and Cephalotes, with 13 and 10 species, respectively. The ant species richness associated with pequi trees varied among habitats (Table 2). The preserved Cerrado trees had a higher ant richness (p<0.05), followed by those in pastureland and Eucalyptus plantation (Fig. 1A). Higher ant abundance was found in trees of the preserved Cerrado and pastureland, with no significant difference between them. While trees from the Eucalyptus plantation observed the lowest ant abundance (Fig. 1B).
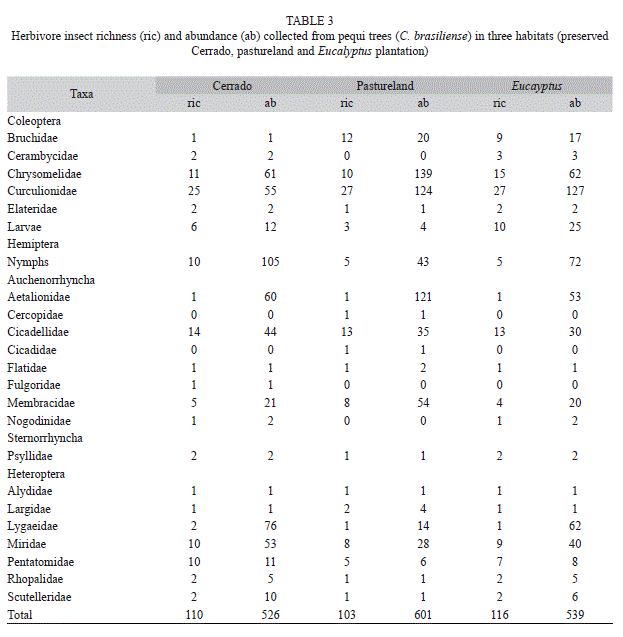
The herbivore insects sampled on C. brasiliense trees were composed by 1 666 individuals (877 chewing and 789 sap-sucking). All chewing herbivores belonged to the order Coleoptera (14 families). The families Curculionidae (50 morphospecies and 306 individuals) and Chrysomelidae (24 morphospecies and 262 individuals) resulted with the highest richness and abundance (Table 3). The richness and abundance of chewing insects associated with these trees differed significantly among habitats (Table 2): these were lower in pequi trees located in the preserved area (p<0.05) than those from either pastureland or Eucalyptus plantation (Fig. 1C, D).
The guild of sap-sucking insects sampled was grouped into 15 Hemiptera families. Aetalionidae (234 individuals) and Cicadellidae (21 morphospecies) were the most abundant and rich sap-sucking families (Table 3). Sap-sucking insects showed the same pattern observed for chewing insects, with the richness and abundance being lower in pequi trees from the preserved Cerrado (p<0.05) than those from pastureland and Eucalyptus plantation (Table 2, Fig. 1 E, F).
Discussion
Many studies have shown a positive relationship between diversity of terrestrial arthropods and habitat heterogeneity (Lassau & Hochuli 2004, Sperber et al. 2004). Positive relations between ant diversity and habitat heterogeneity can be found in different Brazilian biomes, e.g: Amazon (Vasconcelos et al. 2000, Cerrado: Ribas et al. 2003), Pantanal (Ribas & Schoereder 2007) and Atlantic Subtropical forest (Silva et al. 2007). In our study, the ant richness was greater in trees located in preserved cerrado, which has more complex vegetation than trees found in a grassland pasture or Eucalyptus plantation. This result indicates that variations in habitats due human activity negatively affected the ant fauna associated with pequi trees. Thus, the removal of native vegetation to establish Eucalyptus plantation or pastureland may have caused the habitat simplification and, indirectly, a decrease on ant richness, probably due to a decrease in resource availability (e.g. food and nesting sites) in simplified habitats. This pattern supports the hypothesis that ant diversity is positively related to vegetation structural complexity due the classical niche diversification hypothesis.
We found that ant richness and abundance were smaller in Eucalyptus plantation than pasturelands. On the other hand, we did not find differences in ant abundance between preserved Cerrado and pasturelands. Probably this result may be influenced by two main factors. First, structural complexity of the preserved Cerrado seems to be much more similar to pastureland than Eucalyptus plantation. Second, disturbance intensity due human activities is certainty bigger and more frequent in Eucalyptus plantation than in pastureland, which could decrease ant species richness and abundance in Eucalyptus area. This is the first study that shows the negative effect of pastureland and Eucalyptus plantation on ant richness.
In contrast, pequi trees located in preserved Cerrado had lower richness and abundance of insect herbivores than those in pastureland or Eucalyptus plantation. A similar result was found for herbivores from distinct feeding guilds between managed and conserved areas (Speight et al. 2003). Moreover, some studies indicate that insect herbivore’s natural enemies are more effective in pristine areas (Altieri & Letourneau 1982). Normally, predators and parasitoid insects species show high diversity in forest than herbivore insects, that can be pest in agricultural areas (Szinicz et al. 2005). Predators and parasites have a stronger negative effects on herbivores associated with Caryocar brasiliense in Cerrado area. In this way, the decrease in ant diversity on Eucalyptus plantation and pastureland had positively affected the insect herbivore richness and abundance in these habitats. In contrast, tropical agroecosystems in Ecuador had a higher ratio of parasitoid to host species and increased parasitism rates (Tylianakis et al. 2007). In fact, many studies have reported high predation pressure by ants on arthropods (Heil & McKey 2003) while others showed that ant presence could reduce the herbivore insects abundance on their host plants (Oliveira 1997, Fagundes et al. 2005, Rosumek et al. 2009, Neves et al. 2011).
In addition to ants effects (Top-down effects), herbivore insects performance is also affected by host plant quality (Bottom-up effects) (Coley et al. 1985, Coley & Barone 1996). The resource availability hypothesis (Coley et al. 1985) predicts that plants with readily available light but low nutrient availability have elevated rates of carbon fixation and low investment in chemical defenses. In this study, isolated pequi trees in modified habitats should provide a high density of herbivores for the lack of chemical defense. Some authors have argued that higher selection pressure imposed by herbivores had resulted in plants evolving stronger chemical resistance to leaf herbivory in these environments (Coley & Barone 1996, Dirzo & Boege 2008).
The present study shows that the number of ant species on pequi trees decreases with habitat simplification. On the other hand, insect herbivore richness and abundance follows a reverse trend. From conservation point of view we could point out that natural habitat preservation in Cerrado could only benefit ants and at the same time the land use modification could benefit insect herbivore biodiversity. However this conclusion is wrong and must be carefully discussed. In fact, recent studies showed that some herbivore insects species have shown eruptive populations on trees located in disturbed habitats, affecting productivity and often plant survival (Fernandes et al. 2004, Fagundes et al. 2007). Moreover, networks structure can be altered by environmental change and these alterations may have important ecosystem level consequences (Tylianakis et al. 2010). At this point is important to empathize the danger of use just a unique species in conservation programs, forgetting the role of species interaction to community organization and maintenance in space and time (Fagundes et al. 2005).
We suggest a decrease in the transformation of natural habitats in pastureland and Eucalyptus plantations, and in this way the ant top-down effects on herbivore insects. The natural habitat preservation of the Brazilian Cerrado is very important for biodiversity conservation and ecosystem optimum function. Experimental studies on tri-trophic interaction and food webs could be essential to elucidate the patterns of insect distribution and the effect of habitat modification on insect biodiversity and ecological processes.
Acknowledgments
We thanks Ronaldo Reis Júnior and Patrícia de Abreu Moreira for insightful comments on earlier drafts of the manuscript and two other anonymous referees who made valuable contributions to this paper. Financial support by the Fapemig (the Minas Gerais Research Foundation) and CNPq (the National Research Council of Brazil).
References
Almeida, S.P. & J.A. Silva. 1994. Piqui e Buriti: importancia alimentar a população dos cerrados. Embrapa-CPAC, Planaltina, Brasil. [ Links ]
Altieri, M.A. & D.K. Letourneau. 1982. Vegetation management and biological control in agroecosystems. Crop Prot. 1: 405-430. [ Links ]
Basset, Y., V. Novotny, S.E. Miller & R.L. Kitching. 2003. Arthropods of tropical forests-spatio-temporal dynamics and resource use in the canopy. Cambridge University, Cambridge, United Kingdom. [ Links ]
Coley, P.D., J.P. Bryant & F.S. Chapin III. 1985. Resource availability and plant antiherbivore defense. Science 230: 895-899. [ Links ]
Coley, P.D. & J.A. Barone. 1996. Herbivory and plant defenses in tropical forests. Annu. Rev. Ecol. Syst. 27: 305-335. [ Links ]
Crawley, M.J. 2007. Statistical computing-an introduction to data analysis using s-plus. John Wiley & Sons, London, United Kingdom. [ Links ]
Dirzo, R. & K. Boege. 2008. Patterns of herbivory and defense in tropical dry and rain forests, p. 63-78. In W. Carson & S.A. Schnitzer (eds.). Tropical forest community ecology. Blackwell Science, West Sussex, United Kingdom. [ Links ]
Fagundes, M., F.S. Neves & G.W. Fernandes. 2005. Direct and indirect interactions involving ants, insect herbivores, parasitoids, and the host plant Baccharis dracunculifolia (Asteraceae). Ecol. Entomol. 30: 28-35. [ Links ]
Fagundes, M., L.S. Araujo & Y.R.F. Nunes. 2007. Efeito do estágio sucessional do habitat na fenologia do pequizeiro (Caryocar brasiliense: Caryocaraceae). Rev. Bras. Bioc. 5: 144-146. [ Links ]
Fernandes, L.C., M. Fagundes, G.A. Santos & G.M. Silva. 2004. Abundância de insetos herbívoros asociados ao pequizeiro (Caryocar brasiliense Cambess.). Rev. Árvore 28: 919-924. [ Links ]
Gribel, R. & J.D. Hay. 1993. Pollination ecology of Caryocar brasiliense (Caryocaraceae) in central Brazil Cerrado. J. Trop. Ecol. 9: 199-211. [ Links ]
Heil, M. & D. McKey. 2003. Protective ant-plant interactions as model systems in ecological and evolutionary research. Annu. Rev. Ecol. Syst. 34: 425-453. [ Links ]
Hoffmann, B.D. & A.N. Andersen. 2003. Responses of ants to disturbance in Australia, with particular reference to functional groups. Austral Ecol. 28: 444-464. [ Links ]
Klink, C.A. & R.B. Machado. 2005. Conservation of the Brazilian Cerrado. Conserv. Biol. 19: 707-713. [ Links ]
Lassau, S.A. & D.F. Hochuli. 2004. Effects of habitat complexity on ant assemblages. Ecography 27: 157-164. [ Links ]
Myers, N., R.A. Mittermeier, C.G. Mittermeier, G.A.B. Fonseca & J. Kent. 2000. Biodiversity hotspots for conservation priorities. Nature 403: 853-858. [ Links ]
Neves, F.S., L.S. Araújo, M.M. Espírito-Santo, M. Fagundes, G.W. Fernandes, G.A. Sanchez-Azofeifa & M. Quesada. 2010. Canopy herbivory and insect herbivore diversity in a dry forest-savana transition in Brazil. Biotropica 42: 112-118. [ Links ]
Neves, F.S., M. Fagundes, C.F. Sperber & G.W. Wilson. 2011. Tri-trophic level interactions affect host plant development and abundance of insect herbivores. Arthropod-Plant Interac. 5: 351-357. [ Links ]
Oliveira, P.S. 1997. The ecological function of extrafloral nectaries: herbivore deterrence by visiting ants and reproductive output in Caryocar brasiliense (Caryocaraceae). Funct. Ecol 11: 323-330. [ Links ]
Peel, M.C., B.L. Finlayson & T.A. McMahon. 2007. Updated world map of the Koeppen-Geiger climate classification. Hydrol. and Earth Syst. Sciences. 11: 1633-1644. [ Links ]
R Development Core Team, 2010. R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. [ Links ]
Ribas, C.R., J.H. Schoereder, M. Pic & S.M. Soares. 2003. Tree heterogeneity, resource availability, and larger scale processes regulating arboreal ant species richness. Austral Ecol. 28: 305-314. [ Links ]
Ribas, C.R. & J.H. Schoereder. 2007. Ant communities, environmental characteristics and their implications for conservation in the Brazilian Pantanal. Biodivers. Conserv. 16: 1511-1520. [ Links ]
Ricketts, T.H., G. Daily, P.R. Ehrlich & J.P. Fay. 2001. Countryside biogeography of moths in a fragmented landscape: biodiversity in native and agricultural habitats. Conserv. Biol. 15: 378-388. [ Links ]
Rosumek, F.B., F.A.O. Silveira, F.S. Neves, N.P.U. Barbosa, L. Diniz, Y. Oki, F. Pezzini, G.W. Fernandes & T. Cornelissen. 2009. Ants on plants: a meta-analysis of the role of ants as plant biotic defenses. Oecologia 160: 537-549. [ Links ]
Samways, M.J. 2007. Insect Conservation: A synthetic management approach. Annu. Rev. Entomol. 52: 465-487. [ Links ]
Silva, J.M.C. & J.M. Bates. 2002. Biogeographic patterns and conservation in the south American Cerrado: a tropical savanna hotspot. Bioscience 52: 225-233. [ Links ]
Silva, R.R., R.S.M. Feitosa & F. Eberhardt. 2007. Reduced ant diversity along a habitat regeneration gradient in the southern Brazilian Atlantic Forest. For. Ecol. Manag. 240: 61-69. [ Links ]
Speight, M.R., J. Intacha, C.Y. Khen & A.Y.C. Chung. 2003. Influences of forest management on insects, p. 380-393. In Y. Basset, V. Novotny, S. Miller & R. Kitching. (eds.). Arthropods of tropical forests: spatiotemporal dynamics and resource use in the canopy. Cambridge University, Cambridge, Unied Kingdom. [ Links ]
Sperber, C.F., K. Nakayama, M.J. Valverde & F.S. Neves. 2004. Tree species richness and density affect parasitoid diversity in cacao agroforestry. Basic Appl. Ecol. 5: 241-251. [ Links ]
Szinicz, G., K. Martin & J. Sauerborn. 2005. Abundance of selected insect species in natural and agricultural habitats of a tropical upland (Leyte, Philippines). Agric. Ecos. Env. 111: 104-110. [ Links ]
Tylianakis, J.M., T. Tscharntke & O.T. Lewis. 2007. Habitat modification alters the structure of tropical hostparasitoid food webs. Nature 455: 202-205. [ Links ]
Tylianakis, J.M., E. Laliberté, A. Nielsen & J. Bascompte. 2010. Conservation of species interaction networks. Biol. Conserv. 143: 2270-2279. [ Links ]
Vasconcelos, H.L., J.M.S. Vilhena & G.J.A. Caliri. 2000. Responses of ants to selective logging of a central Amazonian forest. J. Appl. Ecol. 37: 508-514. [ Links ]
*Correspondencia:
Frederico S. Neves: Laboratório de Biologia da Conservação, Departamento de Biologia Geral, Universidade Estadual de Montes Claros, Montes claros, MG, Brasil/ Laboratório de Ecologia de Insetos, Departamento de Biologia Geral, Universidade Federal de Minas Gerais, Belo Horizonte, MG, Brasil. fred.neves@gmail.com.
Rodrigo F. Braga: Laboratório de Biologia da Conservação, Departamento de Biologia Geral, Universidade Estadual de Montes Claros, Montes claros, MG, Brasil. rodrigo.fagundes@yahoo.com.br.
Lucimar S. Araújo: Laboratório de Biologia da Conservação, Departamento de Biologia Geral, Universidade Estadual de Montes Claros, Montes claros, MG, Brasil. lucimar.araujo@gmail.com.
Ricardo I. Campos: Laboratório de Ecologia de Formigas,Universidade Federal de Viçosa, 36570-000, Viçosa, MG, Brasil. ricardoicampos@gmail.com
Marcílio Fagundes: Laboratório de Biologia da Conservação, Departamento de Biologia Geral, Universidade Estadual de Montes Claros, Montes claros, MG, Brasil. marcilio.fagundes@unimontes.br.
1. Laboratório de Biologia da Conservação, Departamento de Biologia Geral, Universidade Estadual de Montes Claros, Montes claros, MG, Brasil; rodrigo.fagundes@yahoo.com.br, lucimar.araujo@gmail.com, marcilio.fagundes@unimontes.br
2. Laboratório de Ecologia de Insetos, Departamento de Biologia Geral, Universidade Federal de Minas Gerais, Belo Horizonte, MG, Brasil; fred.neves@gmail.com
3. Laboratório de Ecologia de Formigas,Universidade Federal de Viçosa, 36570-000, Viçosa, MG, Brasil; ricardoicampos@gmail.com
Received 03-VIII-2011. Corrected 08-II-2012. Accepted 09-III-2012.


{kind=link}
{kind=link}
{kind=link}
{kind=link}