










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.60 n.3 San José Sep. 2012
Efecto de la asociación alga-bacteria Bostrychia calliptera (Rhodomelaceae) en el porcentaje de remoción de cromo en laboratorio
*Dirección para correspondencia
Abstract
Water pollution is one of the most important environmental problems worldwide. Recently, biotechnology studies have oriented efforts to study algae-bacterium consortia with the aim to understand the mechanisms to find a possible solution in environmental sciences. This study determined the percentage of chromium removal by the alga-bacterium association exposed to a set of different chromium concentrations under controlled in vitro conditions. Wild plants of Bostrychia calliptera associated with bacterial populations were collected from Dagua River, Pacific coast of Colombia, and were monitored in the laboratory. The trial was conducted with synthetic seawater in bioreactors at two chromium levels: 5 and 10mg/L, and four different experimental treatments: i) algae-bacteria (AB), ii) algae with antibiotic (AA), iii) algal surface sediment, Natural Bacterial Consortium (CBN), and iv) the control without algae or bacteria. The experimental design followed a model of two factors (chromium concentration x combination types) with repeated measures using one factor. The microbial population behavior and the chromium concentration percentage were monitored by using atomic absorption spectroscopy (AAS). According to the data, Algae-bacteria (AB) treatment was the most efficient combination at 10mg/L (87%), whereas the bacterial consortia (CBN) was the most efficient at 5mg/L (62.85%). The results showed significant differences of chromium uptake between algae-bacteria (AB) and natural bacterial consortia (CBN), meaning the importance of those treatments in the chromium removal from coastal waters.
Key words: Bostrychia calliptera, chromium-reducing bacteria, biotransformation, water pollution, chromium contamination.
Resumen
Para determinar el porcentaje de remoción de cromo en la asociación alga-bacteria, se tomaron ejemplares del alga Bostrychia calliptera de pneumatóforos de Avicennia germinans y Rizophora mangle, en la desembocadura del Río Dagua, Pacífico colombiano. El ensayo se realizó in vitro en agua marina sintética a dos concentraciones de cromo 5 y 10mg/L, empleando biorreactores con cuatro tratamientos; i) material Alga-Bacteria (AB), ii) material algal con antibiótico Alga-antibiótico (AA), iii) consorcio bacteriano natural (CBN) y iv) control sin presencia de B. calliptera ni bacteria. Se monitoreó el comportamiento de poblaciones bacterianas y el porcentaje de disminución de cromo mediante AAS (Espectroscopía de absorción atómica). Se obtuvo diferencias significativas entre los tratamientos Alga-bacteria (AB) y el consorcio bacteriano natural (CBN), siendo Alga-bacteria más eficiente a 10mg/L (87%). El consorcio bacteriano natural CBN obtuvo mayor porcentaje de remoción a bajas concentraciones de cromo a 5ppm (62.85%). Los resultados mostraron una interacción positiva entre las bacterias asociadas (CBN) a la superficie del alga roja B. calliptera en su proceso acumulativo de cromo.
Palabras clave: cromo, Bostrychia calliptera, bacterias reductoras de cromo, biotransformación de metales pesados, contaminación por cromo.
La contaminación de las aguas por metales pesados constituye actualmente un grave problema ambiental en los ecosistemas acuáticos continentales y marinos. Particularmente, en la Bahía de Buenaventura (Pacífico colombiano), se han registrado niveles de contaminación por metales pesados como el caso del cromo (Garay et al. 2001, Garay et al. 2004), a concentraciones entre 0.0078mg/L y 25.9mg/L para agua y sedimentos, respectivamente (Calero & Casanova 1997). Según Garay (2004), su concentración va en aumento, donde se han encontrado valores de 2.1 a 5.1mg/L para sedimentos, sobrepasando los 2mg/L reportados como ambientalmente tolerables. Esto como consecuencia al verter de forma directa aguas residuales al mar, y a la influencia del puerto industrial como fuente de contaminación (Acuario et al. 1998, Garay et al. 2001, Garay 2004).
Se ha demostrado que el cromo, es poco biodisponible en condiciones naturales, sin embargo, se ha registrado su bioacumulación por el alga roja B. calliptera (Montagne 1842) en su estado de oxidación Cr (III) (Mostaert et al. 1996, Peña et al. 2008). Actualmente, se desconoce la presencia de mecanismos biológicos, tales como, la interacción alga-bacteria que permitan la incorporación de este metal al protoplasma del alga. Sin embargo, diversos autores (Wang et al. 1989, Cervantes 1991, Lovley & Phillips 1994, Nejidat et al. 2005, Liao et al. 2007, Kidd et al. 2008) reportan la existencia de diferentes géneros bacterianos capaces de realizar cambios en el estado de oxidación del cromo que podrían generar esta situación (Wang et al. 1989, Cervantes 1991, Lovley & Phillips 1994). En Colombia, son pocos los estudios que involucran el papel de la biota microbiana en la bioacumulación de metales pesados. Recientemente, se ha reportado la dinámica de la acumulación del cromo en el alga roja B. calliptera y la participación de los consorcios bacterianos en los procesos de asimilación y detoxificación de metales en el alga (Peña et al. 2008, Peña et al. 2011).
El objetivo de esta investigación fue determinar la participación de las poblaciones bacterianas asociadas a B. calliptera en su proceso de acumulación de cromo y evaluar el porcentaje de remoción del metal a dos concentraciones, 5 y 10mg/L, en condiciones de laboratorio.
Materiales y métodos
Área de estudio: Se realizaron muestreos, entre enero y julio 2009 en la desembocadura del Río Dagua, frente a la Bahía de Buenaventura, en el Departamento del Valle del Cauca (3°51’39.3’’ - 3°51’50’’ N y 77°03’56.7’’ - 77°04’07.9’’ W). Este manglar, plano lodoso, pertenece al estuario poco profundo de la Bahía de Buenaventura (3°54’ N - 77°05’ W), localizada en la costa Pacífica de Colombia. Esta cuenta con un área de 70km2, con un canal central de 16km, un ancho promedio de 4km y una profundidad promedio de 10m. La boca de la bahía se conecta directamente con el océano Pacífico y posee un promedio de precipitación anual de 4 700mm, con un régimen mareal semidiurno y un ámbito promedio de 3.2m. El manglar se encuentra rodeado por una extensa franja de manglares de ribera, especialmente en la parte sur de la bahía (Peña-Salamanca 2008).
Se recolectaron muestras de la biomasa del alga adherida a las raíces y neumatóforos de Avicennia germinans (Linnaeus 1764) y Rhizophora mangle (Linnaeus 1753), empleando un cuadrante de 25×25cm, siguiendo los protocolos de asepsia y bioseguridad para muestras de origen microbiológico (ICONTEC 1994, Eaton et al. 2005). El material vegetal fue recolectado en bolsas con cierre hermético Ziploc® y refrigerado inmediatamente a 5°C, con hielo seco en una nevera portátil. Las muestras fueron transportadas en un lapso máximo de cinco horas a los laboratorios de la Universidad del Valle en la ciudad de Cali, Colombia. Se determinaron las concentraciones de cromo mediante espectroscopía (espectrofotómetro Perkin Elmer Aanalisust 100) de absorción atómica (ASS).
Organismo de estudio: El alga roja Bostrychia calliptera (Rhodomelaceae) se caracteriza por talos de hasta 7cm de largo, de color púrpura a pardo muy ramificado formando cojinetes, con ejes principales corticados y ramitas de crecimiento limitado, parcialmente corticadas. Sus valores de cobertura son relativamente altos en varios lugares tales como Bahía Málaga, Bahía de Buenaventura y la Ensenada de Tumaco (costa Pacífica colombiana), sitios donde se han reportado niveles de mercurio, cobre y plomo que sobrepasan los niveles normales considerados para aguas marinas (Peña et al 2003). La abundancia y amplia distribución de esta especie hacen que tenga una posible utilidad como organismo bioacumulador de metales. De esta alga hay información reportada acerca de su composición química la cual sería útil desde el punto de vista científico y práctico, pues contribuye a dilucidar e interpretar el aporte de las comunidades bénticas vegetales al funcionamiento de los ecosistemas regionales y ofrece información de gran ayuda en la planificación y potencial utilización del recurso (Lenis et al 2007). Estudios ecofisiológicos en estas algas epífitas del manglar, han demostrado su amplio ámbito de tolerancia a fluctuaciones en factores como la salinidad, irradiancia y desecación, debido a los ciclos de emersión-inmersión generados por la marea, al igual que a los niveles de contaminación por metales (Peña et al. 1999, Ospina-Alvarez et al. 2006, Peña-Salamanca 2008).
B. calliptera es la especie dominante en la biomasa algal del estero, con 32% de la biomasa total, seguida por otras especies del complejo “Bostrichetum”, presentando diferencias significativas dependiendo de los periodos climáticos presentes en el área de estudio (periodo lluvioso y seco). Basándose en la distribución vertical de la biomasa en la raíz de los especímenes arbóreos examinados, se detectaron tres zonas (superior, media y baja), encontrando una zona superior estrecha dominada por B. calliptera, y una participación compartida en la zona media con poblaciones mezcladas Peña-Salamanca (2008). El nivel de inundación mareal y la distribución vertical en la raíz parecen ser los factores de mayor influencia en las variaciones de la biomasa algal encontradas.
Los bosques de manglar son uno de los ecosistemas más importantes en el Pacífico colombiano, dominando las zonas costeras (Peña 1998). La comunidad de algas epífitas está dominada principalmente por algas rojas, entre estas el género Bostrychia. Su importancia ecológica recae en su importante aporte de carbono orgánico al ecosistema, al ser uno de los ambientes más productivos de la tierra (Peña-Salamanca 2008).
Estudios desde su fisiología bioacumulativa, han encontrado que B. calliptera posee capacidad de acumulación de metales pesados como el cromo, objeto de este estudio. Sin embargo, se desconoce la naturaleza de la interacción entre el metal y el proceso de acumulación del alga a partir del cambio en su estado de oxidación, lo que permite justificar la evaluación del proceso de remoción del metal, a partir de la exploración del proceso en interacción con microbios tales como las bacterias marinas asociadas a su superficie (Rengifo 2010).
Evaluación del porcentaje de remoción de cromo en la asociación alga-bacteria (B. calliptera) a dos concentraciones del metal: Se utilizó el medio de cultivo Provasoli modificado o ES (Andersen 2005) a 5 y 10mg/L de K2Cr2O7 (Dicromato de Potasio), esterilizado por filtración mediante una membrana de 0.25μm de porosidad, y se distribuyó en porciones de 190mL por erlenmeyer (Rengifo 2010). Las condiciones experimentales fueron las siguientes: Para la primera condición denominada Alga-Bacteria (AB), se tomaron 2g del alga en su estado natural, sin ningún tratamiento y se colocaron en erlenmeyers previamente preparados con el medio de cultivo descrito a las dos concentraciones de cromo. Para la segunda condición se tomaron 2g del alga, pero se les adicionó antibiótico (ciprofloxacina® 2mg/mL, con aplicación cada tres días, según Rengifo 2010), se le denominó Alga-Antibiótico (AA); la tercera condición correspondió a un raspado de la biopelícula presente en la superficie del alga, denominada Consorcio Bacteriano Natural (CBN) y la cuarta condición fue el medio de cultivo sin inocular, o control de degradación química; con dos réplicas cada una. A todos los biorreactores se les adicionó cuatro gotas de dióxido de germanio (GeO2) por litro de agua de mar sintética (West & Calumpong 1988), con el fin de evitar la proliferación de diatomeas (Andersen 2005) y se sometió a bombeo continuo de aire por medio de aireadores de flujo.
El experimento se desarrolló en un periodo de siete días, con muestreos los días uno, tres y siete (que fueron determinados con ensayos preliminares), registrando el valor de pH, contenido bacteriano y concentración de cromo total, mediante ASS. Se sostuvo un fotoperiodo 12h de luz/12h de oscuridad en un cuarto con características ambientales a 70% de humedad relativa y una temperatura de 30°C. Para el último análisis se extrajo 1mL de medio por biorreactor y se filtró con una membrana de 0.25μm previamente enjuagada con HNO3 (1%), el filtrado se almacenó en frascos de vidrio anteriormente enjuagados con una solución de HNO3 (10%), se diluyó a 10mL y se acidificó con HNO3 concentrado hasta obtener un pH menor a dos. Las muestras fueron almacenadas a 4°C, para su posterior lectura mediante espectroscopía de absorción atómica con horno de grafito, empleando un espectrofotómetro Perkin Elmer Aanalisust 100.
Análisis microbiológico: Los recuentos bacterianos se realizaron a las muestras de los sistemas AB y CBN, mediante la técnica de recuento estándar en placa (ICONTEC 1994, Eaton et al. 2005). En el caso del material vegetal (sistema AB) se extrajeron 0.2g de cada biorreactor, que fue macerado y diluido en agua destilada estéril. Posteriormente, se sembró en profundidad 1mL de las diluciones 10-2 a 10-4 en el medio mínimo Agar-agar-K2Cr2O7 a las dos concentraciones de cromo y con dos repeticiones. Las cajas de petri fueron incubadas a 28°C por siete días (Rengifo 2010). Para CBN, el material extraído (1mL) fue diluido según elprocedimiento descrito para el material vegetal. Estos procedimientos se llevaron a cabo en el laboratorio de Microbiología Industrial y Ambiental del Departamento de Biología de la Universidad del Valle.
Caracterización bioquímica: Para ambas muestras se realizó aislamiento de colonias hasta obtener cultivos axénicos. Se sometió a caracterización morfológica (macro y microscópica), tinción Gram, tinción de cápsula y prueba de oxidasa para las cepas Gram negativas. La caracterización bioquímica de las cepas aisladas se realizó mediante el kit EnteroPluri-Test (LIOFILCHEM Bacteriology Products) por un periodo de trece días (ICONTEC 1994), pero no fue posible llegar una identificación más fina.
El diseño experimental correspondió a un modelo de dos factores (Concentración de cromo vs. Tipo de combinación), con medidas repetidas en un factor (Concentración de cromo) y dos réplicas. Se corrió en el programa STATITISCA v7 (Statsoft 2004), mediante una ANOVApr, para este efecto se corrieron pruebas de normalidad de Kolmogorov-Smirnov, homogeneidad de Levene y esfericidad de Mauncly-, pruebas post Hoc (Tukey), parámetros descriptivos y análisis gráfico resultante de los datos para las repeticiones.
Resultados
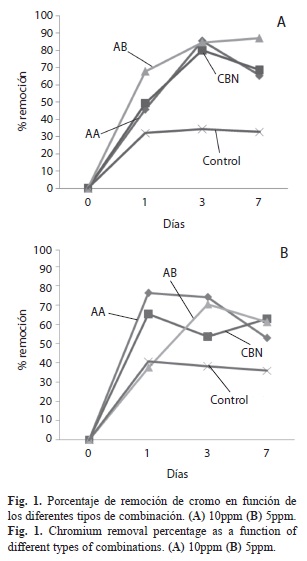
Remoción de cromo: Los resultados mostraron que el mayor porcentaje de remoción de cromo se registró a 10mg/L (Fig. 1), en el tratamiento Alga-Bacteria (AB) con un porcentaje global de remoción del 87%, seguido por el consorcio bacteriano natural (CBN) con un valor de 68.5%. En el análisis de Tukey (p=0.5), la asociación AB presentó mejor rendimiento que el alga tratada con antibiótico (AA), y que el consorcio bacteriano (CBN) separados, a dicha concentración. Mientras que a 5mg/L, se observó la mayor eficiencia, 62.85% en el consorcio bacteriano, (CBN, p=0.004), seguido por AB con un 61.18% (p=0.007693) y finalmente AA con una eficiencia de 52.87% (p=0.013473). Igualmente, se observó que las diferencias en la remoción fueron dependientes de la concentración de cromo total y del tipo de combinación (AA, AB, CBN y control: p=0.035) (Fig. 1). Sin embargo, no se detectó interacción significativa entre ellos.
Crecimiento bacteriano: Los resultados mostraron que el crecimiento bacteriano registró mayores valores a la concentración de 10mg/L y el menor crecimiento a concentraciones de 5mg/L. El consorcio bacteriano natural – aislado– registró el mayor crecimiento (Fig. 2).
De acuerdo con los resultados, se observó que existe una correlación entre el tipo de asociación y su eficiencia en la remoción del cromo. El tratamiento alga-bacteria (AB), mostró mayor eficiencia de remoción a la mayor concentración (10mg/L), mientras que el consorcio bacteriano natural (CBN) presentó mayor eficiencia a 5mg/L (p=0.008). El análisis de varianza mostró que hubo diferencias significativas entre las concentraciones de cromo (p=0.037) y entre los tratamientos (p=0.008).
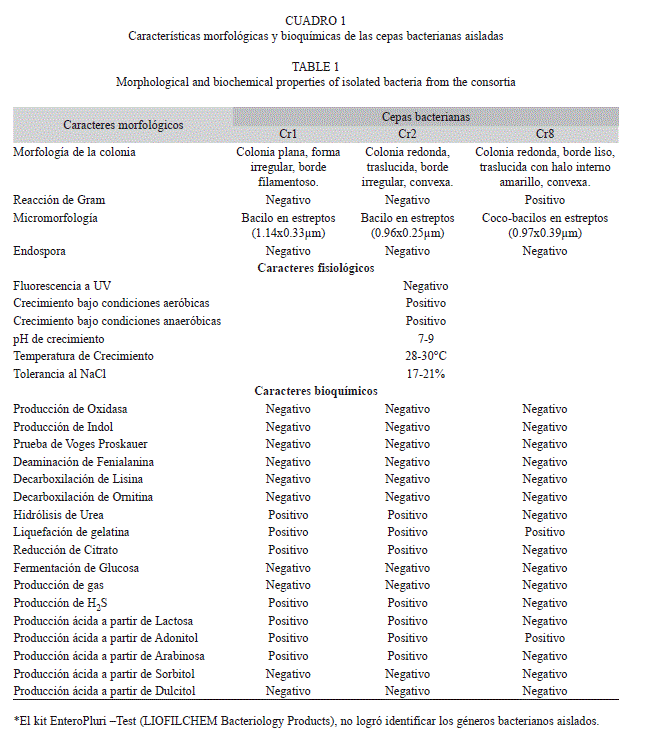
Características morfológicas y bioquímicas de las cepas aisladas: Los resultados registraron tres cepas bacterianas, dos Gram negativas y una Gram positiva de forma bacilar, sin cápsula ni espora, con producción de H2S y fermentación positiva para azúcares, a excepción de la glucosa, ninguna mostró fluorescencia ante la luz ultravioleta, y mostraron una tolerancia importante a la sal en el rango de 17 a 21% y con excelente crecimiento a pH neutro (Cuadro 1).
Discusión
Los resultados permitieron observar una mayor eficiencia de la combinación Alga-Bacteria, respecto a la de Alga-antibiótico. Esto sugiere que las bacterias asociadas a la superficie de B. calliptera poseen participación en el proceso bioacumulativo del alga, dado que el porcentaje de reducción de cromo es mayor en la combinación Alga-bacteria (87% a 10mg/L) que en los sistemas libres de bacteria (Algaantibiótico 65.3% a 10mg/L).
El comportamiento de la remoción de cromo en ensayos de laboratorio, fue reportado por Acosta et al. (2005), quienes señalaron que en el caso de los hongos Cryptococcus neoformans (Vuill 1901) y Helminthosporium sp. (Raabe et al.1981), el porcentaje de remoción del metal en su estado hexavalente no depende de la concentración total en el medio sino de una capacidad innata de la especie. Se sugiere por lo tanto, que el alga B. calliptera como el Consorcio Bacteriano Natural poseen una limitada capacidad de absorción de hasta 4.5mg/Len medio líquido para 168h. Estudios realizados con B. calliptera, han encontrado ámbitos de acumulación para cobre de 2.2 a 3.7mg/L, mercurio hasta 0.9mg/L y plomo hasta 1.3mg/L (Duque et al. 2005). En contraste a Ospina & Peña (2003) quienes observaron valores de acumulación en la misma especie que oscilan entre 8.333 y 29.048mg/L para cobre, 0.050 y 0.198 para mercurio y 1.095 y 8.548mg/L para plomo. Ambos casos sugieren la importancia del alga roja B. calliptera como un bioindicador de contaminación ambiental al presentar el comportamiento de bioacumulador de los metales pesados examinados. Adicionalmente y en relación a los valores de adsorción del alga y el Consorcio Bacteriano Natural, se considera que es posible, la existencia de una interacción positiva entre el consorcio bacteriano encontrado en la superficie de B. calliptera y el proceso de transformación de cromo observado por varios autores, tales como: Arias & Tebo 2003, Cervantes 1991, Kwak et al. 2003 y Paul 2004.
El Consorcio bacteriano natural (CBN), presentó mayor eficiencia en comparación a la combinación Alga-bacteria a la concentración menor (5mg/L). Sin embargo, a una concentración superior (10mg/L) el sistema Algabacteria presenta mayor eficiencia que CBN, lo que permite identificar la importancia de la presencia bacteriana sobre la superficie del alga, al compararlos con los niveles de transformación que presenta el alga y el consorcio bacteriano aislado. Los resultados coinciden con lo planteado por Cervantes (1991), Arias & Tebo (2003) y Kwak et al. (2003), los cuales han demostrado la capacidad de ciertas bacterias de reducir cromo (VI) a cromo (III) e igualmente de bio-adsorberlo (Rabbania et al. 2005, Cabrera et al. 2007). Los experimentos en este estudio, permiten confirmar ambas condiciones, ya que supone que para el sistema Alga-bacteria, las bacterias (o CBN) pueden funcionar como mediadoras en el proceso reductor, al cambiar el estado de oxidación del Cr (VI) a Cr (III), facilitando la bio-adsorción por parte del alga (Acosta et al. 2005). Adicionalmente, la combinación consorcio bacteriano natural (CBN) demostró ser capaz de bio-adsorber cromo, confirmando las observaciones aportadas por Lee et al. (2006). Zhu et al. (2008) igualmente reportan la reducción de cromo por parte de diversos grupos bacterianos, como es el caso del consorcio bacteriano chino SRB III (Shen & Wang 1993), generando una posible explicación a los fenómenos de transformación que presentó el consorcio examinado, sustentándolos en función al co-metabolismo con azufre.
De acuerdo con los resultados obtenidos por Fude et al. (1994), existen al menos dos posibles rutas para la inmovilización de Cr. La primera involucra a Pseudomonas ambigua G-1 (Suzuki 1992), mecanismo descartado ya que las cepas empleadas resultaron ser oxidasa negativas, y la segunda explicación, de aplicación necesaria, sugiere que las bacterias son productoras de H2S (Arias & Tebo 2003), y que este actúa como agente reductor para el Cr (VI). Esto ha sido observado en ambientes marinos (Smillie et al. 1981) y es consistente con las bacterias de origen marino extraídas en este estudio. En este caso, el H2S se re-oxida al ser producido por las bacterias y el potencial redox del mismo no baja hasta que la mayoría o la totalidad del Cr (VI) se reduce. Fude et al. (1994) indicaron este posible mecanismo para el consorcio SRB III, por cuanto la mayoría de las bacterias que forman dicho complejo son productoras de H2S, similares a las observaciones comprobadas para los consorcios bacterianos analizados en este trabajo.
Las variaciones observadas en las concentraciones de cromo a lo largo del experimento, podrían corresponder a lo que Puzon et al. (2008) denominaron como parte del régimen continuo o de ciclaje del metal en su medio natural; mediante la producción de Cr (III) soluble en el sobrenadante, lo que ecológicamente correspondería al cromo biodisponible y asimilable por demás algas, plantas, bacterias y/o organismos circundantes (Shen & Wang 1993, Campos et al. 1995, McLean & Beveridge 2001, Bencheikh-Latmani et al. 2007, Megharaj et al. 2003, VanEngelen et al. 2008). Igualmente, la formación de complejos órgano-Cr (III) en la reducción enzimática de Cr (VI) señaladas por Puzon et al. (2008) y Kwak et al. (2003), en donde el Cr (III) está ligado a la forma oxidada de los reductores, tales como NAD+ señalarían el siguiente paso de asimilación bacteriana del contaminante.
En este trabajo se señala por primera vez la presencia del consorcio alga-bacteria y su papel en el porcentaje de remoción del cromo en aguas costeras, reiterando la importancia de la presencia bacteriana en la remoción del metal en su estado de oxidación (VI) en aguas marinas costeras con algún grado de contaminación por metales. Las perspectivas de este estudio están orientadas a ampliar las observaciones sobre los mecanismos biológicos que emplea el alga B. calliptera y la identificación de sus bacterias asociadas en los procesos de biorremediación de aguas marinas con presencia de cromo. Igualmente, generar aplicaciones de carácter ecológico para solucionar problemáticas de contaminación presentes en cuerpos de agua, a través de biopelículas, filtros, o celdas que aporten procesos descontaminantes con efectos adicionales que permitan desarrollo en poblaciones aisladas o de baja influencia tecnológica.
Agradecimientos
El presente estudio se realizó en el marco del proyecto de convocatoria interna de la Universidad del Valle # CI 57748 de 2007 “Efectos de la concentración de metales pesados sobre la biología reproductiva y características ultraestructurales en algas rojas y sus implicaciones en biorremediación”. Los autores agradecen al personal del laboratorio de microbiología industrial y ambiental del departamento de Biología y del laboratorio de análisis industrial del departamento de Química, por su apoyo en el análisis y tratamiento de las muestras. Al personal de la sección de botánica de la Universidad del Valle, Colombia, por su apoyo en el desarrollo del proyecto.
Referencias
Acosta, M., G. Moctezuma-Zárate, C. Gutiérrez & X. Rodríguez. 2005. Bioadsorción de Cromo (VI) en Solución Acuosa por la Biomasa Celular de Cryptococcus neoformans y Helminthosporium sp. Inf. tecnol. 16: 11-15. [ Links ]
Acuario, G., A. Rossin, P.F. Teixeira & F. Zepeda. 1998. Diagnóstico de la situación del manejo de residuos sólidos municipales en América Latina y el Caribe. Organización Panamericana de la Salud. Organización Mundial de la Salud. Banco Interamericano de Desarrollo y la Organización Panamericana de la Salud. Serie ambiental 18. OPS. América Latina. [ Links ]
Andersen, R.A. (ed.) 2005. Algal Culturing Techniques: A Book for All Phycologists. Elsevier, West Boothbay Harbor, EEUU. [ Links ]
Arias, Y & B. Tebo. 2003. Cr (VI) Reduction by Sulfidogenic and Nonsulfidogenic Microbial Consortia. Appl. Microbiol. 69: 1847-1853. [ Links ]
Assfalg, M., I. Bertini, M. Bruschi, C. Michel & P. Turano. 2002. The metal reductase activity of some multiheme cytochromes c: NMR structural characterization of the reduction of chromium (VI) to chromium (III) by cytochrome c7. PNAS 99: 9750-9754. [ Links ]
Bencheikh-latmani, R., S. Middleton-william, L. Haucke, C. Criddle, L. Wu, L. Zhou & B. Tebo. 2005. Global Transcriptional Profiling of Shewanella oneidensis MR-1 during Cr (VI) and U (VI) Reduction. Appl. Microbiol. 71: 7453-7460. [ Links ]
Buerge, I. & S. Hug. 1999. Influence of Mineral Surfaces on Chromium(VI) Reduction by Iron(II). Environ. Sci. Technol. 33: 4285-4291. [ Links ]
Cabrera, G., M. Viera, J. Gómez, D. Cantero & E. Donati. 2007. Bacterial removal of chromium (VI) and (III) in a continuous system. Biodegradation 18: 505-513. [ Links ]
Campos, J., M. Martinez-Pacheco & C. Cervantes. 1995. Hexavalent-chromium reduction by a chromateresistant Bacillus sp. strain. Antonie Leeuwenhoek 68: 203-208. [ Links ]
Cervantes, C. 1991. Bacterial interactions with chromate. Antonie Leeuwenhoek 59:229-233. [ Links ]
Duque, Y, E. Peña, J. Cantera & R. Benitez. 2003. Macroalgas acumulando metales pesados en la Bahía de Buenaventura - Pacifico Colombiano. Programa y resúmenes congreso Latinoamericano de Ciencias del Mar COLACMAR. Colombia. [ Links ]
Eaton, A., L. Clesceri., E. Rice & A. Greenberg. 2005. Standard methods for the examination of water and wastewater. AWWA, Nueva York, EEUU. [ Links ]
Fude, L., B. Harris, M. Urrutia & T. Beveridge. 1994. American Society for Microbiology Reduction of Cr (VI) by a Consortium of Sulfate-Reducing Bacteria (SRB III). Appl. Microbiol. 60: 1525-1531. [ Links ]
Garay, J., B. Marín & A. Vélez. 2001. Contaminación marino-costera en Colombia. Informe del Estado de los Ambientes Marinos y Costeros en Colombia. Santa Martha, Instituto de Investigaciones Marinas y Costeras José Benito Vives de Andréis - INVEMAR, San Andrés, Colombia. [ Links ]
Garay, J. 2004. Programa Nacional de Investigación, Evaluación, Prevención, Reducción y Control de Fuentes Terrestres y Marinas de Contaminación al Mar PNICM. Instituto de Investigaciones Marinas y Costeras José Benito Vives de Andréis - INVEMAR, San Andrés, Colombia. [ Links ]
Hadbock, B. & C. Jones. 1997. Bacterial Respiration. Bacteriological review 41: 47-99. [ Links ]
Icontec.1994. Norma técnica colombiana NTC 4092. Microbiología de alimentos y de alimentos para animales. Reglas generales para el análisis microbiológico. Instituto Colombiano de normas técnicas y certificación - ICONTEC, Santafé de Bogotá [ Links ].
Kwak, Y., D. Lee & H. Kim. 2003. Vibrio harveyi Nitroreductase Is Also a Chromate Reductase. Appl. Microbiol. 69: 4390-4395. [ Links ]
Kidd, P., A. Prieto-fernández, C. Monterroso & M.J. Acea. 2008. Rhizosphere microbial community and hexachlorocyclohexane degradative potential in contrasting plant species. Plant Soil 302: 233-247. [ Links ]
Lee, S., J. Lee, J. Lee & H. Chon. 2006. Effects of indigenous bacteria on Cr (VI) reduction in Cr-contaminated sediment with industrial wastes. J. Geochem. Explor. 88: 41-44. [ Links ]
Lenis, L., E. Peña-Salamanca & D. Chito-Trujillo. 2007. Extracción, Separación y Elucidación estructural de dos metabolitos secundarios de alga marina Bostrychia callipetera. Scientia et Technica. 13: 97-102. [ Links ]
Lovley, D.W. 1991. Dissimilatory Fe (III) and Mn (IV) reduction. Microbiol. Rev. 55:259-287. [ Links ]
Lovley, D. & E. Phillips. 1994. Reduction of Chromate by Desulfovibrio vulgaris and Its C3 Cytochrome. Appl. Microbiol. 60: 726-728. [ Links ]
Mclean, J. & T. Beveridge. 2001. Chromate Reduction by a Pseudomonad Isolated from a Site Contaminated with Chromated Copper Arsenate. Appl. Microbiol. 67: 1076-1084. [ Links ]
Megharaj, M., S. Avudainayagam & R. Naidu. 2003. Toxicity of Hexavalent Chromium and Its Reduction by Bacteria Isolated from Soil Contaminated withTannery Waste. Curr. Microbiol. 47: 51-54. [ Links ]
Mostaert, A., D. Orlovich & R. King. 1996. Ion Compartmentation in the Red Alga Caloglossa leprieurii in Response to Salinity Changes: Freeze-Substitution and X-ray Microanalysis. New Phytol. 132: 513-519. [ Links ]
Nejidat, A., I. Saadi & Z. Ronen. 2005. Degradation of 2,4,6-tribromophenol by bacterial cells attached to chalk collected from a contaminated aquifer. J. Ind. Microbiol. Biotechnol. 32: 502-513. [ Links ]
Paul, A. 2004. Aerobic chromate reduction by chromiumresistant bacteria isolated from serpentine soil Arundhati Pal. Microbiol. Res. 159:347-354. [ Links ]
Ospina-Alvarez, N., E. Peña-Salamanca & R. Benitez. 2003. Uso de algas bénticas con indicadores de contaminación por metales pesados en la Bahía de Buenaventura – Pacifico Colombiano. Conferencia Internacional Usos múltiples del Agua: Para la Vida y el Desarrollo Sostenible. Instituto Cinara, Universidad del Valle, Colombia. [ Links ]
Ospina-Alvarez, N., E. Peña-Salamanca & R. Benitez. 2006. Efecto de la salinidad en la capacidad de bioacumulación de plomo en el alga verde Rhizoclonium riparium (Roth) Harvey (Chlorophyceae, Cladophorales). Actual Biol. 28: 17-25. [ Links ]
Peña, E.J. 1998. Physiological ecology of mangrove associated macroalgae in a tropical estuary. Ph.D. Thesis, University of South Carolina, EEUU. [ Links ]
Peña, E.J., R. Zingmark & C. Nietch. 1999. Comparative photosynthesis of two species of intertidal epiphytic macroalgae on mangrove roots during submersion and emersion. J. Phycol. 35: 1206-1214. [ Links ]
Peña-Salamanca, E., Lozano-Duque & R. Benitez. 2003. Dinámica de la Bioacumulación de Metales Pesadosen dos Especies de Algas en la Bahía de Buenaventura, Pacifico Colombiano. Universidad del Valle. Departamento de Biología. Universidad del Cauca. Departamento de Química. Cali, Colombia. [ Links ]
Peña-Salamanca, E. 2008. Dinámica espacial y temporal de la biomasa algal asociada a las raíces de mangle en la bahía de Buenaventura, Costa pacífica de Colombia. Bol. Invemar 37: 55-70. [ Links ]
Peña-Salamanca, E., A. Rengifo-Gallego & N. Benitez-Campo. 2011. Detoxification mechanisms of heavy metals by algal-bacteria consortia, p. 441-453. In K. Se-Kwon (ed.). Handbook of Marine Macroalgae: Biotechnology and Applied Phycology. JohnWiley & Sons, EEUU. [ Links ]
Puzon, G.J., J.N. Petersen, A.G. Roberts, D.M. Kramer & L. Xun. 2002. A bacterial flavin reductase system reduces chromate to a soluble chromium (III)-NAD+ complex (2002). Biochem. Biophys. Res. Commun. 294: 76-81. [ Links ]
Rabbania, M., H. Ghafouriana, S. Sadeghib & Y. Nazerib. 2005. Biosorption of chromium (III) by new bacterial strain (NRC-BT-2). Int. Congr. Ser. 1276: 268-269. [ Links ]
Rengifo, A. 2010. Caracterización bacteriana y evaluación del efecto de la asociación alga-bacteria (alga roja Bostrychia calliptera Rhodomelaceae) en el porcentaje de remoción de cromo. Tesis de pregrado, Universidad del Valle, Cali, Colombia. [ Links ]
Shen, H. & Y. Wang. 1993. Characterization of Enzymatic Reduction of Hexavalent Chromium by Escherichia coli ATCC 33456. Appl. Microbiol. 59: 3771-3777. [ Links ]
Smillie, R.H., K. Hunter & M. Loutit. 1981. Reduction of Chromium (VI) by Bacterially Produced Hydrogen Sulphide in a Marine Environment. Water Res. 15: 1351-1354. [ Links ]
Vanengelen, M., B. Peyton, M. Mormile & M. Pinkart. 2008. Fe (III), Cr (VI), and Fe (III) mediated Cr (VI) reduction in alkaline media using a Halomonas isolate from Soap Lake, Washington. Biodegradation 19:841-850. [ Links ]
Wang, P., T. Mori, K. Komori, M. Sasatsu, K. Toda & H. Ohtake. 1989. Isolation and Characterization of an Enterobacter cloacae Strain That Reduces Hexavalent Chromium under Anaerobic Conditions. Appl. Microbiol. 55: 1665-1669. [ Links ]
West, J.A. & H.P. Calumpong. 1988. Mixed-phase reproduction of Bostrychia (Ceramiales: Rhodophyta) in cultura. I. B. tenella (Lamouroux) J. Agardh. Jpn. J. Phycol. 36: 292-310. [ Links ]
Zhu, W., L. Chai, Z. Yang & Z. Ma. 2008. Reduction of high concentrations of chromate by Leucobacter sp. CRB1 isolated from Changsha, China. World J. Microbiol. Biotechnol. 24: 991-996. [ Links ]
*Correspondencia:
Ana Lucía Rengifo-Gallego: Grupo de Biología Vegetal Aplicada. Facultad de Ciencias Naturales y Exactas. Universidad del Valle. Calle 13 N°100-00 Cali, Colombia. anarengifog@gmail.com
Enrique Peña-Salamanca: Grupo de Biología Vegetal Aplicada. Facultad de Ciencias Naturales y Exactas. Universidad del Valle. Calle 13 N°100-00 Cali, Colombia. enrique.pena@correounivalle.edu.co
Neyla Benitez-Campo: Grupo de Microbiología y Biotecnología Ambiental. Facultad de Ciencias Naturales y Exactas. Universidad del Valle. Calle 13 N° 100-00 Cali, Colombia. neyla.benitez@correounivalle.edu.co
1. Grupo de Biología Vegetal Aplicada. Facultad de Ciencias Naturales y Exactas. Universidad del Valle. Calle 13 N°100-00 Cali, Colombia; anarengifog@gmail.com, enrique.pena@correounivalle.edu.co
2. Grupo de Microbiología y Biotecnología Ambiental. Facultad de Ciencias Naturales y Exactas. Universidad del Valle. Calle 13 N° 100-00 Cali, Colombia; neyla.benitez@correounivalle.edu.co
Recibido 02-VI-2011. Corregido 09-III-2012. Aceptado 12-IV-2012.


{kind=link}
{kind=link}
{kind=link}