










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.60 n.3 San José Sep. 2012
Dynamics of leaf litter humidity, depth and quantity: two restoration strategies failed to mimic ground microhabitat conditions of a low montane and premontane forest in Costa Rica
Zaidett Barrientos1*
*Dirección para correspondencia
Abstract
Little is known about how restoration strategies affect aspects like leaf litter’s quantity, depth and humidity. I analyzed leaf litter’s quantity, depth and humidity yearly patterns in a primary tropical lower montane wet forest and two restored areas: a 15 year old secondary forest (unassisted restoration) and a 40 year old Cupressus lusitanica plantation (natural understory). The three habitats are located in the Río Macho Forest Reserve, Costa Rica. Twenty litter samples were taken every three months (April 2009-April 2010) in each habitat; humidity was measured in 439g samples (average), depth and quantity were measured in five points inside 50x50cm plots. None of the restoration strategies reproduced the primary forest leaf litter humidity, depth and quantity yearly patterns. Primary forest leaf litter humidity was higher and more stable (x=73.2), followed by secondary forest (x=63.3) and cypress plantation (x=52.9) (Kruskall-Wallis=77.93, n=232, p=0.00). In the primary (Kruskal-Wallis=31.63, n=78, p<0.001) and secondary (Kruskal-Wallis=11.79, n=75, p=0.008) forest litter accumulation was higher during April due to strong winds. In the primary forest (Kruskal-wallis=21.83, n=78, p<0.001) and the cypress plantation (Kruskal-wallis=39.99, n=80, p<0.001) leaf litter depth was shallow in October because heavy rains compacted it. Depth patterns were different from quantity patterns and described the leaf litter’s structure in different ecosystems though the year.
Key words: restoration strategies evaluation, leaf litter humidity, leaf litter quantity, leaf litter depth, leaf litterstructural complexity.
Resumen
Poco se sabe acerca de cómo las estrategias de restauración afectan aspectos como la cantidad, profundidad y humedad de la hojarasca. Se analizaron estas variables en un bosque tropical húmedo montano bajo, considerado bosque primario y dos áreas restauradas: un bosque secundario de 15 años (restauración natural) y una plantación de Cupressus lusitanica de 40 años con sotobosque restaurado naturalmente. Los sitios estudiados se ubican en la reserva forestal Río Macho, Costa Rica. Los muestreos se realizaron cada tres meses (abril 2009-abril 2010). En cada ocasión se escogieron al azar 20 cuadrículas de 50x50cm de las que se recogió 439g en promedio de hojarasca para medir la humedad por diferencia entre peso seco y húmedo. En cada cuadrícula se midió la profundidad y cantidad de hojarasca haciendo un promedio de cinco puntos. La cantidad se midió con el número de hojas ensartadas en un picahielos. La profundidad se midió con una regla metálica. Ninguna de las técnicas de restauración reproduce los patrones anuales del bosque primario en cuanto a la humedad, profundidad y cantidad de hojarasca. La humedad de la hojarasca del bosque primario es mas alta y estable ( =73.2), seguida por el bosque secundario ( =63.3) y el cipresal ( =52.9) (Kruskall-Wallis=77.93, n=232, p=0.00). La cantidad de hojarasca acumulada en el bosque primario (Kruskal-Wallis=31.63, n=78, p<0.001) y el secundario (Kruskal-Wallis=11.79, n=75 p=0.008) es mayor en abril debido a los fuertes vientos. La profundidad de la hojarasca del bosque primario (Kruskal-wallis=21.83, n=78, p<0.001) y en el cipresal (Kruskal-wallis=39.99, n=80, p<0.001) es menor durante octubre debido a que los fuertes aguaceros la compactan. Los patrones de cantidad y profundidad de la hojarasca presentan diferencias, que describen la estructura de la hojarasca en diferentes ecosistemas a lo largo del año.
Palabras clave: evaluación de estrategias de restauración, humedad de hojarasca, cantidad de hojarasca, profundidad de hojarasca, complejidad estructural de hojarasca.
Tropical forests have undergone extensive deforestation throughout the world (Geist & Lambin 2002, Quesada et al. 2009, FAO 2010), increasing the need to develop scientific restoration efforts. The selected restoration strategy will impact soil quality, biodiversity, aquifer recharge and forest environmental services. However, the effects of restoration strategies on different ecosystem components are not well known and the importance of deterministic and stochastic factors needs more discussion (Chadzon 2008).
In Costa Rica all restoration strategies leave understory to natural ecological succession (or “unassisted restoration”), but the canopy establishment follows three major types: natural ecological succession (“do nothing restoration”), native plant species plantation, and introduced plant species plantation (like Cuppresus lusitanica) (Cusack & Montagnini 2004, Jiménez 2005, Murillo 2005, Bonilla et al. 2008, Sampaio et al. 2008, Quesada et al. 2009, Barrientos & Monge 2010, Cole et al. 2010, Castellanos-Barliza & León 2011). More complex restoration options like direct transfer of intact habitat islands or seed addition (Watts et al. 2008) are not common in Costa Rica.
Restoration strategy selection in Costa Rica is a consequence of political and economic national strategies (Jiménez 2005, Murillo 2005). Nevertheless, the application of a restoration strategy should take into account technical assessments (De Camino 2005), or management plans that include all the complex elements of an ecological restoration program (Windhager 1999, Fernández 2006, Clewell & Aronson 2008).
Selection of a restoration strategy impacts micro and macro-scale elements, such as soil temperature, litter quality, soil respiration rates, nitrogen availability, microbial biomass, faunal community composition, among others, that would influence the functioning of the restored forest or ecosystem (Ayres et al. 2009a). Therefore, technical analysis of restoration strategies require multi-disciplinary and ecosystem level studies. However, to achieve such knowledge it is important to understand the dynamics of several phenomena: understory composition; forest temperature and moisture; litter production, structure, humidity, and decomposition rates; soil erosion, plant dispersion, etc. In Costa Rica the study of these phenomena was started by L.A. Fournier in the 20th century (Fournier & Camacho de Castro 1973, Fournier & Herrera de Fournier 1978).
One of the basic components of a tropical forest is the litter that accumulates on the ground, it constitutes an essential part of nutrient cycling (Wardle 2002, Álvarez-Sánchez & Harmon 2003, Ayres et al. 2009a, Castellanos-Barliza & León 2011). The vegetation that is chosen in a restoration program will define temperature and soil humidity, mainly due to canopy density, stratus structure, sunlight that can reach understory and soil, leaf litter production and decomposition rate, and tree falling rate, among others (Mosquera et al. 2007). It also defines: soil physical and chemical properties (Ayres et al. 2009a); understory plant species composition; litter composition coming from the canopy and understory; leaf litter nutrients, production and decaying rate (Mosquera et al. 2007, Scherer-Lorenzen et al. 2007, Hättenschwiler et al. 2008, Vivanco & Austin 2008); organism diversity (Vasconcelos 1999, Naranjo-García 2003, Doblas 2007, Sánchez et al. 2007, Bonilla et al. 2008, Castro-Díez et al. 2008, Ayres et al. 2009c); and forest regeneration and recovery rates and patterns (Letcher & Chadzon 2009). It has been shown, that the replacement of native by exotic species affects the litter production rate, nitrogen content and nutrient release during decomposition (Bonilla et al. 2008, Castro-Díez et al. 2008).
Many studies have been performed on leaf litter production (Bonilla et al. 2008), decomposition rates (Álvarez-Sánchez & Harmon 2003, Castro-Díez et al. 2008, Ayres et al. 2009b, c), nutrient release (Ayres et al. 2009a, Castellanos-Barliza & León 2011) and on litter organism diversity and its impact on decomposition rates (Fournier & Herrera de Fournier 1978, Barrientos 2000, Palacios-Vargas et al. 2007, Ayres et al. 2009a). Despite the large number of species that inhabit the leaf litter, few studies have been done on its structural properties, dynamics and relation with organisms.
A high diversity of angiosperms is characteristic of tropical forests and allows the establishment of a structurally complex and diverse leaf litter layer, even if there is an accumulation of leaves belonging to the nearest plant, a phenomenon known as “home field” (Gholz et al. 2000, Ayres et al. 2009a, Ayres et al. 2009b). To my knowledge, no leaf litter structural complexity hypotheses or indexes have been built. Future ecologic work on this matter should consider plant species diversity, litter quantity, vertical space covered (depth), accumulation and decomposition rate, hyphal density (as hypha may be attaching litter layers) and diversity of type texture and shape of material added to the litter.
A more complex litter layer has more species and organisms, probably because it provides more area to hide from predators, feed and lay eggs (Barrientos 2000, Sabo et al. 2005, Palacios-Vargas et al. 2007, Sabu et al. 2008). In addition, the amount of litter defines the amount and rate of the interactions in the different trophic levels (Sabo et al. 2005). But contrary to what could be expected, Ayres et al. (2009c) found that litter decomposes more rapidly near the plant that produces it. This is probably the result of specialization by decomposers.
Litter also retains soil humidity longer than bare soil (Anderson 1990), allowing water to percolate instead of rapidly evaporating. Litter makes forest humidity more stable by keeping water (Díaz-Fernández et al. 2006, Ruiz et al. 2009), and prevents rain’s direct impact on the soil, reducing erosion (Di Stefano & Fournier 2005).
Rainfall and litter humidity are key factors in a complex interplay of processes. There is a negative relationship between litterfall and rainfall (Mosquera et al. 2007), witch at least in some tropical forests can be attributed to the presence of deciduous plants (Fournier & Camacho de Castro 1973). However, rainfall is crucial for litter decay (Cornejo et al. 1994, Castellanos-Barliza & León 2011) and correlates with microbial biomass (Schimel et al. 1999) and abundance of other organisms (Bonilla et al. 2008). Litter humidity affects the community living under, in and on the litter, because in many cases species migrate vertically in order to achieve optimal environmental conditions (Barrientos 2000, Naranjo-García 2003, Doblas 2007). Another important finding is that extreme drought and occasional rewetting cause water stress, which significantly reduces biomass and microbial biodiversity and causes changes in the structure of the decomposers community (Schimel et al. 1999).
Litter humidity is affected by rainfall, litter composition and canopy cover, as well as by type, thickness and permeability of the soil (Álvarez-Sánchez & Harmon 2003, Díaz Fernández et al. 2006, Sampaio et al. 2008). All these factors are modified with deforestation and establishment of a different flora community (Vasconcelos & Laurance 2005, Bonilla et al. 2008); therefore, any restoration process should consider these factors. However, litter humidity has been studied almost exclusively in relation to forest fires in temperate regions and lowland tropical dry forests (Odiwe & Muoghalu 2003, Dezzeo & Chacón 2006, Ruiz et al. 2009).
The effect of different restoration strategies on humidity, structure, temperature, species composition and nutrient release of forest litter has not been analysed. This study analysed three variables that are important to understand leaf litter complexity and general patterns that affect biodiversity in the forests (leaf litter humidity, depth and quantity) in a primary forest, a secondary forest and a plantation.
Materials and methods
Research area: The study was carried out in Orosi Valley, Costa Rica, at Reserva Forestal Río Macho. This reserve limits with the Tapantí-Macizo Cerro de la Muerte National Park, which has more than 58 000ha of primary forest. The dry season goes from December through April, whereas the rainy season runs from May through November.
Three habitats were selected: a primary forest (or “old growth forest” according to Clark (1996)) near the “El llano” water dam (9°45’56.07” N - 83°51’47.11” W, 1 640msm), in a tropical lower montane wet forest area with ultisol humult soil; a secondary forest (or “young secondary forest” according to Clark (1996)) left to natural succession for about 15 years (9°45’29.52” N - 83°51’23.27” W, 1 684 msm), in a tropical lower montane wet forest area with ultisol humult soil, and a Cupressus lusitanica plantation that has been without management for nearly 40 years and therefore has a poorly developed understory dominated by hardwood leaves species (9°47’52” N - 83°51’51” W, 1 309msm). The C. lusitanica plantation belongs to a tropical humid premontane forest area and the soil is inceptison dystrandept. The three places have similar slopes, all of over 40°.
Sampling: Samples were collected every three months in each habitat: January 2010, April 2009, July and October 2009. Due to logistic problems the cypress plantation was sampled in April 2010 instead of April 2009.
In each sampling date a 200m randomly selected transect was set at least 10m away from any trail to avoid border effect and away of tree gaps to avoid the effect of direct sun evaporation on litter (Camargo & Kapos 1995). Along each line, 20 sampling plots (50x50cm) were chosen with a digital random number generator. Samples were collected between 9:00am and 2:00pm, to standardize the daily humidity fluctuation (Ruiz et al. 2009) and also because this is the time of the day with less rain and where re-humidification by atmospheric water vapour is less important (Pyne et al.1996, Dirks et al. 2010).
One leaf litter sample (mean 439g, SD=188, min=89, max=1 470) was collected in each 50x50cm plot. In each plot all litter was collected including small branches less than 5mm in diameter, fragmented litter and humus (representing successive decaying stages), only bare soil, living plants, stones and branches bigger than 5mm in diameter were left. Litter samples were kept in a plant oven (60oC) for several days until constant dry weight was reached to apply the formula: Humidity percentage=(wet weight–dry weight)/wet weight*100.
Litter depth was measured in each plot with a standard millimetric ruler in five independent places of a 50x50cm subplot. Average leaf-litter depth for each plot was calculated. Litter quantity was assessed by the number of hardwood leaves that could be threaded with an ice pick (10cm long, 3.5mm diameter) (five samples were taken for each plot) in a 50x50cm subplot. Cypresses needles were not considered in this methodology as they do not form layers. Leaf layers were analysed because they are useful structural elements for litter tropical dwellers (Naranjo-García 2003, Doblas 2007, Palacios-Vargas et al. 2007, Eaton et al. 2011).
All statistical analyses where performed with Statgraphics Centurion XV. Leaf-litter humidity comparisons were made between habitats and between sampling dates (Kruskal-Wallis ANOVA). Litter abundance and depth were analyzed in relation to each other (Spearman correlation), and to humidity, habitat and sampling date (Kruskal-Wallis ANOVA).
Results
Leaf litter humidity comparison between habitats
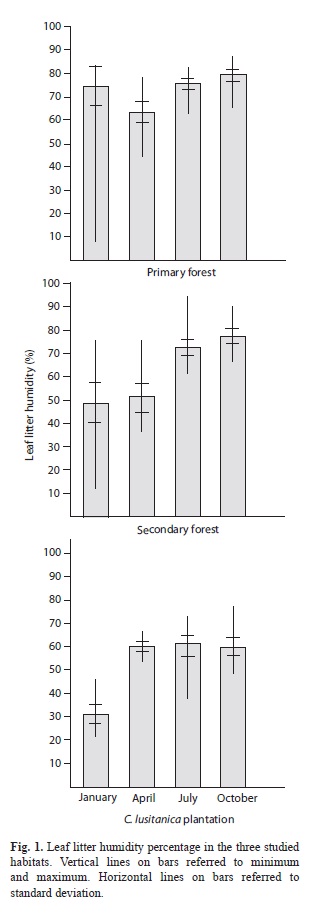
Primary forest had the wettest litter (x=73.2, n=77, SD=11.6, min=8, max=87), followed by the secondary forest (x=63.3, n=75, SD=16.8, min=12, max=94) and cypress plantation (x=52.9, n=80, SD=14.6, min=21.3, max=77.6) (Kruskall-Wallis=77.93, n=232, p=0.00).
Leaf litter humidity according to season
Dry season: During January the driest place was the plantation (x=31.1%) followed by the secondary forest (x=48.9%) and the primary forest (x=74.7%) (Kruskall-Wallis=35.75, n=59, p<0.001) (Fig.1). During the April samplings the litter humidity was similar in the primary forest and the cypress plantation, but the secondary forest was dryer (Kruskall-Wallis=12.99, n=55, p=0.0015) (Fig. 1).
Leaf litter humidity yearly pattern according to habitat
Primary forest: Litter was drier during the April sampling (Kruskall-Wallis=33.28, n=77, p<0.001), but the difference between the driest and the wettest sampling was of only 15.8% (x max=79.4%, x min=63.6%) (x max=meanmaximum, x min=mean minimum) (Fig. 1).
Secondary forest: Litter humidity pattern shows a longer period of low litter humidity than in the primary forest and the C. lussitanica plantation. In this habitat the litter was dryer during the January and April samplings (Kruskall-Wallis=49.16, n=75, p<0.001), and the difference between the driest and the wettest sampling was of 28.7% (x max=77.6%, x min=48.9%) (Fig. 1).
Cupressus lusitanica plantation: Litter was driest during the January sampling (Kruskall-Wallis=44.1617, n=80, p<0.001) and the difference between the driest and the wettest sampling was 29.5% (x max=60.6%, x min=31.1%) (Fig.1). The wettest sampling mean (60.6%) in this habitat was even drier than the driest sampling mean in the primary forest (63.6%).
Leaf litter depth and quantity in the three studied habitats in a year
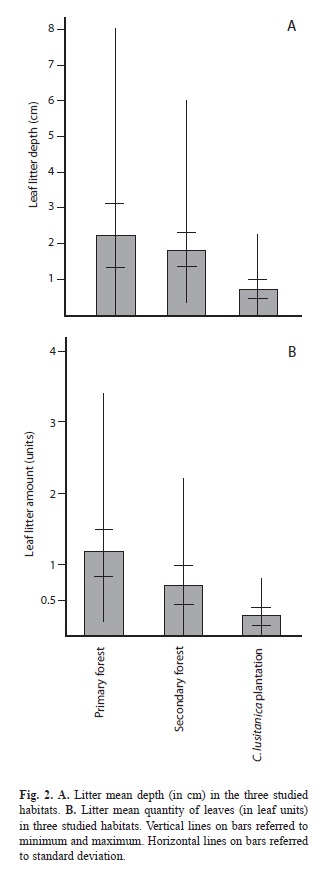
Thicker litter layer and greater quantity of leaves were associated with greater levels of litter humidity (Litter quantity, Spearman correlation r=0.3, n=232, p=0.000) (r=correlation, n=sample size), (Litter depth, Spearman correlation r=0.27, n=232, p=0.000). The litter depth (Kruskall-Wallis=78.95, n=233, p=0.0) (Fig. 2A) and quantity (Kruskall-Wallis=92.47, n=233, p=0.0) (Fig. 2B) were higher in the primary forest followed by the secondary forest and the cypress plantation.
Leaf litter depth and quantity yearly patterns according to habitat
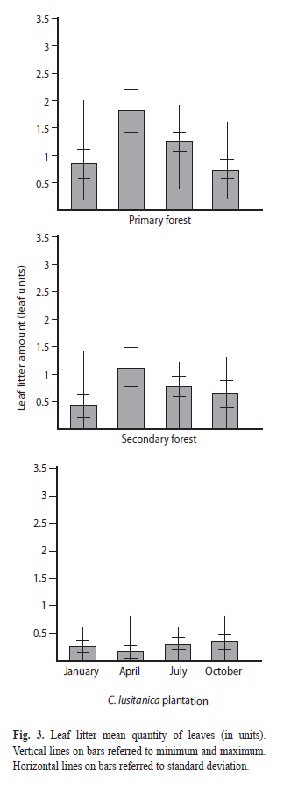
Litter quantity pattern in primary (Kruskal-Wallis=31.63, n=78, p<0.001) (Fig. 3) and secondary forest (Kruskal-Wallis=11.79, n=75 p=0.008) (Fig. 3) show that leaf is more abundant in April and decreases until January. A completely different pattern was found in the cypress plantation where October is the sampling with more leaf abundance while April had the fewest (Kruskal-Wallis=7.77, n=80, p=0.0509, marginally significant).
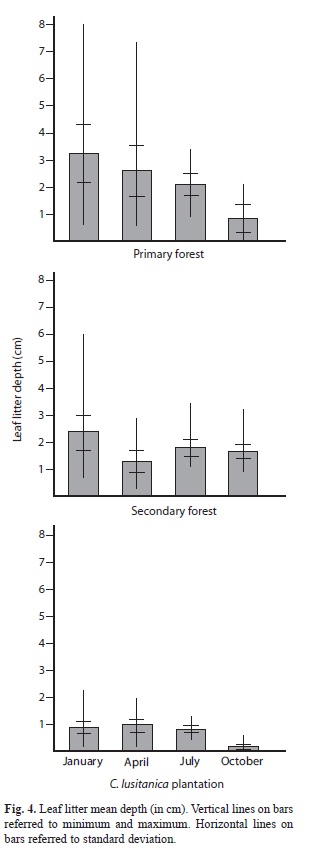
Litter depth in primary forest (Kruskal-Wallis=21.83, n=78, p<0.001) (Fig. 4) and C. lusitanica plantation (Kruskal-Wallis=39.99, n=80, p<0.001) (Fig. 4) had their lowest value during the October sampling, while in the secondary forest all the samples had about the same litter depth values, just slightly lower during April (Kruskal-Wallis=10.68, n=75, p=0.014) (Fig. 4).
Discussion
The findings in this research show that leaf litter humidity in primary forest is higher and more stable around the year than in these restoration habitats. Wind, evaporation, solar radiation and UV-B radiation, soil`s capacity to keep humidity, drainage and temperature are some factors that affect litter humidity (Ruiz et al. 2009, Dirks et al. 2010, Smith et al. 2010). In this case, probably the taller and more abundant canopy and understory dicotiledonean foliage cover of the primary forest (personal observation) help keeping the litter`s humidity higher and more stable the year around. The structure (quantity and depth) and species composition of the litter may also help keeping high humidity levels (Álvarez-Sánchez & Harmon 2003).
The C. lusitanica plantation`s litter humidity is lower than in the primary and secondary forest almost the year around. The only exception is in April when the secondary forest is the driest. Nevertheless, the highest litter’s humidity in the cypress plantation was 60.6% which is lower than the driest sampling mean in the primary forest (63.6%). This can be attributed to lower values of litter quantity and depth (Ruiz et al. 2009), litter decomposition stage (cypress secondary compounds may delay decomposition rates) (Ruiz et al. 2009), litter composition (mainly cypress needles), a poorly developed understory (personal observation), a more regular canopy structure and a more homogenous foliage cover (Rodriguez & Cordero, unpublished data).
Secondary forest’s and cypress plantation’s litter gain and lose more humidity than the primary forest’s litter, but during the rainy season, the secondary forest can be almost as humid as the primary forest. The only study known to me that compares angiosperm and gymnosperm litter humidity was made by Díaz-Fernández et al. (2006). This study shows that both kinds are able to keep the same humidity, the only exception are grasses, which are able to keep twice as much. Therefore, the wider humidity range of these habitats may be a result of the lower amount of litter, shallower litter and less developed canopy in the secondary forest, and less developed understory in the cypress plantation (personal observation). This allows the sun to increase litter’s temperature and evaporation rate. In the secondary forest, another possible explanation is the abundant grass of the secondary forest, because grass is able to keep more moisture (Díaz-Fernández et al. 2006). The grass abundance in the secondary forest may be a result of occasional cattle activity in the area. In future studies the wind speed should also be considered.
It is interesting that litter quantity and litter depth have the same general pattern: primary forest has the highest values, followed by the secondary forest; the cypress plantation has the smallest litter quantity and the shallower litter. Nevertheless, the patterns around the year in the three habitats are different.
In the primary and secondary forest litter quantity had its highest values during the April samplings. This pattern matches the inverse relation between litter productivity and rainfall found by other researchers (Di Stefano & Fournier 2005, Mosquera et al. 2007, Sánchez et al. 2007). The three studied habitats are within tropical lower montane wet forest and tropical humid premontane forest areas (now altered by human activity), therefore, some deciduous trees occur. Nevertheless, most are evergreen species, and hence these results cannot be attributed only to the leaf fall found in tropical dry forest (Fournier & Camacho de Castro 1973). The litter quantity in Rio Macho primary and secondary forest is probably also a consequence of the strong winds that blow from the beginning of December-March or April. This may have caused a litter accumulation in the April sample. After that, litter decomposes slowly until December when another cycle begins. In addition, it could be that the low humidity of the dry season reduces the decomposition rate. Nevertheless, this last item should be considered carefully as field studies show contradictions about the relation between litter decaying rate, temperature, rainfall, ultraviolet-B radiation and evapotranspiration (Cornejo et al. 1994, Monedero & González 1995, Aerts 1997, Powers et al. 2009, Smith et al. 2010, Eaton et al. 2011, Salinas et al. 2011); experimental studies on this topic are needed. This pattern could not be detected in the cypress plantation, maybe as a result of the absence of hardwood trees.
The primary forest and cypress plantation showed a litter’s depth pattern that has the lowest values in October, while April and January have the highest values. As the studied areas have steep slopes, it would be reasonable to think that some litter is flown downhill by heavy rains, but it is not the case because the amount of leaves in the litter was similar to January. September and October are the wettest months of the year in this area (Herrera 1986, UNESCO 2007). This may produce a compaction effect, leaving less space between leaves. This effect may also help keeping high litter moisture during that time of the year, but it also means that less interleaf space is left for micro and macro organisms. Besides this, another aspect that may strongly affect biota and decaying rates is the oxygen concentration. If litter is more compact and wet it may be also more anaerobic. On the contrary, during January, although there are less leaves in the litter they are more lose and occupy more vertical space. In the secondary forest the lowest litter depth values were obtained in April; further studies are needed to understand this pattern.
Litter depth patterns found in this study are different to litter quantity patterns, because describe different aspects of the litter’s structure and they should not be considered equivalents.
It is reasonable to expect that less litter humidity correlated with less litter quantity and depth, but as shown in this research the relation between them is much more complex. Therefore litter humidity, depth and quantity must be analyzed in relation with understory and canopy foliage cover and composition, and with soil features. In fact litter humidity presents a continuum with soil and understory, therefore many litter dwellers migrate between these strata to achieve better humidity conditions (Naranjo-García 2003, Doblas 2007, Ayres et al. 2009a). Complementary research is being conducted on terrestrial molluscs in these plots, showing that their abundance and size distribution is strongly correlated with these factors (Barrientos, unpublished data). It would also be interesting to analyse regeneration patterns in relation to litter humidity and structure.
The selection of a given restoration strategy affects ecosystem features like the litter humidity and structure, that, in return, will determine the biodiversity that can get established in it. Here it is shown that natural succession and reforestation with cypress present litter humidity, depth and quantity patterns that do not resemble conditions in a primary forest floor. Future studies should also analyze if plantations with native species where the understory is left to natural succession, produce patterns of litter humidity, quantity and depth more similar to primary forest. Other aspect not considered in this study is the effect of fragment sizes on these variables (Didham 1998).
Acknowledgment
Andrés Monge, Danyi Prieto and FabianAraya gave field assistance and Junior Pérez helped with literature search. I am especially grateful to Esteban Acuña, Ligia Bermudez, Gabriela Pérez and Maribel Zúñiga for field and technical assistance. Julián Monge-Nájera, Roberto Cordero, Edna Naranjo and five anonymous reviewers helped improving this paper. This paper was partially financied by the project “FEES-CONARE Análisis ecosistémico para la evaluación de la restauración forestal y sus implicaciones en el secuestro de carbono en un bosque nublado” and by the UNED’s Research Vicepresidency. Special acknowledgment to Katya Calderón for her administrative support. This paper is part of the Ph.D. requirements of the Doctorado en Ciencias Naturales para el Desarrollo at the Instituto Tecnológico de Costa Rica (ITCR), Universidad Nacional (UNA) and Universidad Estatal a Distancia (UNED).
References
Aerts, R. 1997. Climate, leaf litter chemistry and leaf litter decomposition in terrestrial ecosystems: a triangular relationship. Oikos 79: 439-449. [ Links ]
Álvarez-Sánchez, J. & M.E. Harmon. 2003. Descomposición de hojarasca: hojas y madera, p. 108-122. In J. Álvarez-Sánchez & E. Naranjo-García (eds.). Ecología del suelo en la selva tropical húmeda de México. Instituto de Ecología, A.C., Instituto de Biología y Facultad de Ciencias, UNAM, Xalapa, México. [ Links ]
Anderson, H.E. 1990. Moisture diffusivity and response time in fine forest fuels. Can. J. For. Res. 20: 315-325. [ Links ]
Ayres, E., H. Steltzer, S. Berg, M.D. Wallenstein, B.L. Simmons & D.H. Wall. 2009a. Tree species traits influence soil physical, chemical, and biological properties in high elevation forests. Plos One 4: 1-11 (también disponible en línea: http://www.plosone.org/article/info%3Adoi%2F10.1371%2Fjournal.pone.0005964). [ Links ]
Ayres, E., H. Steltzer, B.L. Simmons, R.T. Simpson, J.M. Steinweg, M.D. Wallenstein, N. Mellor, W.J. Parton, J.C. Moore & D.H. Wall. 2009b. Homefield advantage accelerates leaf litter decomposition in forests. Soil Biol. Biochem. 41: 606-610 (también disponible en línea: http://warnercnr.colostate.edu/~edayres/Pubs/2009%20Ayres%20et%20al%20Soil%20Biol%20Biochem.pdf). [ Links ]
Ayres, E., H. Steltzer, S. Berg & D.H Wall. 2009c. Soil biota accelerate decomposition in high-elevation forests by specializing in the breakdown of litter produced by the plant species above them. J. Ecol. 97: 901-912 (también disponible en línea: http://warnercnr.colostate.edu/~edayres/Pubs/2009%20Ayres%20et%20al%20J%20Ecol.pdf). [ Links ]
Barrientos, Z. & J. Monge-Nájera. 2010. Restauración ecológica en la meseta central de Costa Rica. Biocenosis 23: 20-25 (también disponible en línea: http://web.uned.ac.cr/biocenosis/images/stories/Vol232/05-barrientos-restauracion.pdf). [ Links ]
Barrientos, Z. 2000. Population dynamics and spatial distribution of the terrestrial snail Ovachlamys fulgens (Stylommatophora: Helicarionidae) in a tropical environment. Rev. Biol. Trop. 48: 71-87 (también disponible en línea: http://www.scielo.sa.cr/scielo.php ? script=sci_arttext & pid=S0034-77442000000100009 & lng=es & nrm=iso & tlng=en). [ Links ]
Bonilla, R., B. Roncall, J. Jimeno & T. García. 2008. Producción y descomposición de la hojarasca en bosques nativos y de Leucaena sp., en Codazzi, Cesar. Revista Corpoica – Ciencia y Tecnología Agropecuaria 9: 5-11 (también disponible en línea: http://www.corpoica.gov.co/SitioWeb/Archivos/Revista/Produccinydescomposicindelahojarasca.pdf). [ Links ]
Camargo, J.L.C. & V. Kapos. 1995. Complex edge effects on soil moisture and microclimate in central Amazonian forest. J. Trop. Ecol. 11: 205-221. [ Links ]
Castellanos-Barliza, J. & J.D. León. 2011. Descomposición de hojarasca y liberación de nutrientes en plantaciones de Acacia mangium (Mimosaceae) establecidas en suelos degradados de Colombia. Rev. Biol. Trop. 59: 113-128. [ Links ]
Castro-Díez, P., N. González & A. Fernández. 2008. Los árboles exóticos invasores alteran la tasa de descomposición de la hojarasca. Cuad. Soc. Esp. Cienc. For. 25: 99-104. [ Links ]
Chadzon, R.L. 2008. Chance and determinisms in tropical forest sucession, p. 384-408. In W.P. Carson & S.A. Schinitzer (eds.). Tropical forest community ecology. Wiley-Blackwell, Oxford (también disponible en línea: http://books.google.es/books ? hl=es & lr= & id=GqJrYsYlw-kC & oi=fnd & pg=PA384 & dq=Chance+and+determinism+in+tropical+forest+succession & ots=N7v5zb8fEQ & sig=qYDHlTS7ywX0E2Eqx9kAjRf09a8#v=onepage & q=Chance%20and%20determinism%20in%20tropical%20forest%20succession&f=false). [ Links ]
Clark, D.B. 1996. Abolishing virginity. J. Trop. Ecol. 12: 735-739 (también disponible en linea: http://www.fsl.orst.edu/Oldgrowthworkshop/download/Clarke_Abolishing_Virginity.pdf). [ Links ]
Clewell, A. & J. Aronson. 2008. Ecological restoration: principles, values, and structure of an emerging profession. Island, Washington, D.C., USA. [ Links ]
Cole, R.J., K.D. Holl & R.A. Zahawi. 2010. Seed rain under tree islands planted to restore degraded lands in a tropical agricultural landscape. Ecol. Appl. 20: 1255-1269. [ Links ]
Cornejo, F.J., A. Varela & S.J. Wright. 1994 Tropical forest litter decomposition under seasonal drought: nutrient release, fungi and bacteria. Oikos 70: 183-190. [ Links ]
Cusack, D. & F. Montagnini. 2004. The role of native species plantations in recovery of understory wood diversity in degraded pasturelands of Costa Rica. For. Ecol. Manage. 188: 1-15. [ Links ]
De Camino, R. 2005. ¿Especies nativas o exóticas? Ese es el dilema. Ambientico 141: 7-8 (también disponible en línea: http://www.ambientico.una.ac.cr/141.pdf). [ Links ]
Dezzeo, N. & N. Chacón. 2006. Litterfall and nutrient input in undisturbed and adjacent fire disturbed forests of the Gran Sabana, Southern Venezuela. Interciencia 31: 894-899. [ Links ]
Di Stefano, J.F. & L.A. Fournier. 2005. Caída de hojarasca y tasas de descomposición de las hojas de Vochysia guatemalesis en una plantación de 10 años, Tabarcia de Mora, Costa Rica. Agron. Costarr. 29: 9-16 (también disponible en línea: http://www.mag.go.cr/rev_agr/v29n01_009.pdf). [ Links ]
Díaz-Fernández, E., A.F. Gómez Tagle Rojas, Y. Chávez Huerta, H. Zepeda Castro & A. Gómez Tagle Chávez. 2006. Análisis comparativo sobre la interceptación pluvial entre hojarasca de pino, encino y pastizal utilizando un simulador de lluvia en la cuenca hidrográfica de Cointzio, Michoacán. V Congreso Internacional & XI Nacional de Ciencias Ambientales, Morelos, México. [ Links ]
Didham, R.K. 1998. Altered leaf-litter decomposition rates in tropical forest fragments. Oecologia 116: 397-406. [ Links ]
Dirks, I., Y. Navon, D. Kanas, R. Dumbur & J.M. Grünzweig. 2010. Atmospheric water vapor as driver of litter decomposition in Mediterranean shrub land and grassland during rainless seasons. Glob. Change Biol. 16: 2799-2812. [ Links ]
Doblas, E. 2007. Ecología de los macroinvertebrados edáficos en un ecosistema árido mediterráneo. Tesis de Doctorado, Universidad de Granada, España. (Consultado: 26 setiembre 2011, http://digibug.ugr.es/bitstream/10481/1537/1/16713187.pdf). [ Links ]
Eaton, W.D., S. Mcdonald, M. Roed, M. Vandecar, L. Karen, J.B. Hauge & D. Barry. 2011. A comparison of nutrient dynamics and microbial community characteristics across seasons and soil types in two different old growth forests in Costa Rica. Trop. Ecol. 52: 35-48. [ Links ]
FAO. 2010. Evaluación de los recursos forestales mundiales. 2010. FAO, Roma, Italia. (Consultado: 23 setiembre 2011, http://www.fao.org/docrep/013/i1757s/i1757s.pdf). [ Links ]
Fernández, A. 2006. Restauración ecológica. (Consultado: 25 mayo 2010, http://www.consumer.es/web/es/medio_ambiente/urbano/2006/05/05/151595.php). [ Links ]
Fournier, L.A. & M.E. Herrera de Fournier. 1978. Cambios de la microflora del suelo en varias etapas de la sucesión en Ciudad Colón, Costa Rica. Rev. Biol. Trop. 26: 103-112. [ Links ]
Fournier, L.A. & L. Camacho de Castro. 1973. Producción y descomposición del mantillo en un bosque secundario húmedo de premontano. Rev. Biol. Trop. 21: 59-67. [ Links ]
Geist, H.J. & E.F. Lambin. 2002. Proximate causes and underlying driving forces of tropical deforestation. BioScience 52: 143-150 (también disponible en línea: http://www.puce.edu.ec/zoologia/vertebrados/personal/sburneo/cursos/EcologiaII/Bibliografia/2-5%20Deforestacion%20tropical.pdf). [ Links ]
Gholz, H.L., D.A. Wedin, S.M. Smitherma, M.E. Harmon & W.J. Parton. 2000. Long term dynamics of pine and hardwood litter in contrasting environments: toward a global model of decomposition. Glob. Change Biol. 6: 751-765. [ Links ]
Hättenschwiler, S., B. Aeschlimann, M.M. Couteaux, J. Roy & D. Bonal. 2008. High variation in foliage and leaf litter chemistry among 45 tree species of a neotropical rainforest community. New Phytol. 179: 165-175. [ Links ]
Herrera, W. 1986. Vegetación y clima de Costa Rica. EUNED, San José, Costa Rica. [ Links ]
Jiménez, Q. 2005. Cultivo de árboles en Costa Rica: ¡nativos y exóticos! Ambientico 141: 9-11 (tambiéndisponible en línea: http://www.ambientico.una.ac.cr/141.pdf). [ Links ]
Letcher, S. & R. Chadzon. 2009. Rapid recovery of biomass, species richness, and species composition in a forest chronosequence in northeastern Costa Rica. Biotropica 41: 608-617. [ Links ]
Monedero, C. & V. González. 1995. Producción de hojarasca y descomposición en una selva nublada del ramal interior de la cordillera de la costa, Venezuela. Ecotropicos 8: 1-14 (también disponible en línea: http://ecotropicos.saber.ula.ve/db/ecotropicos/edocs/vol8_n1-2/articulo1.pdf). [ Links ]
Mosquera, H., Y. Ramos & D. Abadía. 2007. Cuantificación de la caída de hojarasca como medida de la productividad primaria neta en un bosque pluvial tropical en Salero, Chocó, Colombia. Revista Institucional Universidad Tecnológica del Chocó 26: 28-41. [ Links ]
Murillo, O. 2005. Desmitificación del debate entre especies exóticas y nativas. Ambientico 141: 4-6 (también disponible en línea: http://www.una.ac.cr/ambi/Ambien-Tico/141.pdf#page=9). [ Links ]
Naranjo-García, E. 2003. Malacofauna de la hojarasca, p. 141-161. In J. Álvarez-Sánchez & E. Naranjo-García (eds.). Ecología del suelo en la selva tropical húmeda de México. Instituto de Ecología, A.C., Instituto de Biología y Facultad de Ciencias, UNAM, Xalapa, México. [ Links ]
Odiwe, A.I. & J.I. Muoghalu. 2003. Litterfall dynamics and forest floor litter as influenced by fire in a secondary lowland rain forest in Nigeria. Trop. Ecol. 44:243-251. [ Links ]
Palacios-Vargas, J.G., G. Castaño-Meneses, J.A. Gómez-Anaya, A. Martínez-Yrizar, B.E. Mejía-Recamier & J. Martínez-Sánchez. 2007. Litter and soil arthropods diversity and density in a tropical dry forest ecosystem in Western Mexico. Biodivers. Conserv. 16: 3703-3717 (también disponible en línea: http://www.springerlink.com/content/vwwv752468337uj7/). [ Links ]
Powers, J.S., R.A. Montgomery, E.C. Adair, F.Q. Brearley, S.J. DeWalt, C.T. Castanho, J. Chave, E. Deinert, J.U. Ganzhorn, M.E. Gilbert, J.A. González-Iturbe, S. Bunyavejchewin, H.R. Grau, K.E. Harms, A. Hiremath, S. Iriarte-Vivar, E. Manzane, A.A. de Oliveira, L. Poorter, J-B. Ramanamanjato, C. Salk, A. Varela, G.D. Weiblen & M.T. Lerdau. 2009. Decomposition in tropical forests: a pan-tropical study of the effects of litter type, litter placement and mesofaunal exclusion across a precipitation gradient. J. Ecol. 97: 801-811. [ Links ]
Pyne, S.J., P.L. Andrews & R.D. Laven. 1996. Introduction to wildland fire. John Wiley & Sons, New York, USA. (Consultado: 4 abril 2011, http://books.google.es/books ? hl=es & lr= & id=yT6bzpUyFIwC & oi=fnd & pg=PR20 & dq=Introduction+to+wildland+fire & ots=AgeQVU9NTv & sig=WzXwUEhBUZy0FNhLhWNT9UNdmcs#v=onepage & q & f=false). [ Links ]
Quesada, M., G.A. Sanchez-Azofeifa, M. Alvarez-Anorve, K.E. Stoner, L. Avila-Cabadilla, J. Calvo-Alvarado, A. Castillo, M.M. Espirito-Santo, M. Fagundes, G.W. Fernandes, J. Gamon, M. Lopezaraiza-Mikel, D. Lawrence, P. Morellato, J. Powers, F.S. Neves, V. Rosas-Guerrero, R. Sayago & G. Sanchez-Montoya. 2009. Succession and management of tropical dry forests in the Americas: Review and new perspectives. For. Ecol. Manage. 258: 1014-1024. [ Links ]
Ruiz, A.D., J.A. Vega & J.G. Álvarez. 2009. Modelización de la variabilidad horaria de los contenidos de humedad en hojarasca de Eucalyptus globulus. Invest. Agrar: Sist. Recur. For. 18: 247-263. [ Links ]
Sabo, J.L., C.U. Soykan & A. Keller. 2005. Functional roles of leaf litter detritus in terrestrial food webs, p. 211- 223. In P.C. Ruiters, V. Wolters & J.C. Moore (eds.).Dynamic food webs: Multispecies assemblages, ecosystem development, and environmental change. Academic, Massachusetts, USA (también disponible en línea: http://www.public.asu.edu/~jlsabo/pubs/Sabo_et_al_IN_PRESS_Litter_Chapter.pdf). [ Links ]
Sabu, T.K., P.J. Vineesh & K.V. Vinod. 2008. Diversity of forest litter-inhabiting ants along elevations in the Wayanad region of the Western Ghats. J. Insect Sci. 8: 1-14 (también disponible en línea: http://www.insectscience.org/8.69/i1536-2442-8-69.pdf). [ Links ]
Salinas, N., Y. Malhi, P. Meir, M. Silman, R. Roman Cuesta, J. Huaman, D. Salinas, V. Huaman, A. Gibaja, M. Mamani & F. Farfan. 2011. The sensitivity of tropical leaf litter decomposition to temperature: results from a large-scale leaf translocation experiment along an elevation gradient in Peruvian forests. New Phytol. 189: 967-977 (también disponible en línea: http://onlinelibrary.wiley.com/doi/10.1111/j.1469-8137.2010.03521.x/pdf). [ Links ]
Sampaio, A., P. Rodríguez-González, S. Varandas, R.M. Cortes & M.T. Ferreira. 2008. Leaf litter decomposition in western Iberian forested wetlands: lentic versus lotic response. Limnetica 27: 93-106. [ Links ]
Sánchez, S., G. Crespo & M. Hernández. 2007. Acumulación de hojarasca en un pastizal de Panicum máximum y en un sistema silvopastoril de Panicum maximum y Leucaena leucocephala. Pastos y Forrajes 30: 357-371 (también disponible en línea: http://scielo.sld.cu/pdf/pyf/v30n3/pyf06307.pdf). [ Links ]
Scherer-Lorenzen, M., J.L. Bonilla & C. Potvin. 2007. Tree species richness affects litter production and decomposition rates in a tropical biodiversity experiment. Oikos 116: 2108-2124. [ Links ]
Schimel, J.P., J.M. Gulledge, J.S. Clein-Curley, J.E. Lindstrom & J.F. Braddock. 1999. Moisture effects on microbial activity and community structure in decomposing birch litter in the Alaskan taiga. Soil Biol. Biochem. 31: 831-838 (también disponible en línea: http://www.lter.uaf.edu/dev2009/pdf/584_schimel_gulledge_1999.pdf). [ Links ]
Smith, W.K., W. Gao, H. Steltzer, M.D. Wallenstein & R. Tree. 2010. Moisture availability influences the effect of ultraviolet-B radiation on leaf litter decomposition. Glob. Change Biol. 16: 484-495. [ Links ]
UNESCO. 2007. Balance hídrico superficial de Costa Rica: Periodo 1070-2002. Documentos técnicos del PHI-LAC, No. 10. Ed. Programa hidrológico internacional (PHI) de la Oficina Regional de la Ciencia para América Latina y el Caribe de la Organización de las Naciones Unidas para la Educación, la Ciencia y la Cultura (UNESCO), Montevideo, Uruguay. [ Links ]
Vasconcelos, H.L. 1999. Effects of forest disturbance on the structure of ground-foraging ant communities in central Amazonia. Biodivers. Conserv. 8: 409-420 (también disponible en línea: http://www.springerlink.com/content/p231p37106021848/). [ Links ]
Vasconcelos, H.L. & W.F. Laurence. 2005. Influence of habitat, litter type, and soil invertebrates on leaf-litter decomposition in a fragmented Amazonian landscape. Oecologia 144: 456-462. [ Links ]
Vivanco, L. & A.T. Austin. 2008. Tree species identity alters forest litterdecomposition through long-term plant and soil interactions in Patagonia, Argentina. J. Ecol. 96: 727-736. [ Links ]
Wardle, D.A. 2002. Communities and ecosystems: linking the aboveground and belowground components. Princeton University, New Jersey, USA. [ Links ]
Watts, C.H., M. Vojvodic-Vukovic, G.C. Arnold & R.K. Didham. 2008. A comparison of restoration techniques to accelerate recovery of litter decomposition and microbial activity in an experimental peat bog restoration trial. Wetlands Ecol. Manage. 16: 199-217. [ Links ]
Windhager, S. 1999. Restauración ecológica y evolutiva. Ciencia al día internacional 2: 1-9. (Consultado: 26 Marzo 2011, http://www.ciencia.cl/CienciaAlDia/volumen2/numero4/articulos/v2n4a7v1.PDF). [ Links ]
*Correspondencia:
Zaidett Barrientos: Laboratorio de Ecología Urbana, UNED, 2050 San José, Costa Rica. zbarrientos@uned.ac.cr
1. Laboratorio de Ecología Urbana, UNED, 2050 San José, Costa Rica; zbarrientos@uned.ac.cr
Received 02-VI-2011. Corrected 14-I-2012. Accepted 17-II-2012.


{kind=link}
{kind=link}
{kind=link}
{kind=link}