










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.60 n.3 San José Sep. 2012
Reclutamiento de corales pétreos en arrecifes coralinos a diferentes distancias de fuentes de contaminación en La Habana, Cuba
Pedro Alcolado-Prieto1*, Hansel Caballero Aragón2*, Pedro M. Alcolado1 & Alexander Lopeztegui Castillo3*
*Dirección para correspondencia
Abstract
The effect of pollution on coral recruitment has been insufficiently studied. This research deals with coral recruitment in coastal areas and aimed to determine the variations of density and dominant species of corals recruits in sites at different distances from pollution sources. The composition and structure of stony coral (scleractinian and milleporids) recruit associations were characterized in the fringing reef of Western Havana, Cuba. This reef is influenced by urban pollution from the Almendares River and a sewage outlet located at its mouth. Four sites were sampled on the upper fore reef escarpment at 10m deep every three months between July 2007 and May 2008. A 25cm side quadrat was used to determine the density and taxonomic composition of recruits smaller than 3cm in diameter. Sampling units were placed following a random-systematic pattern. The mean density of recruits was determined both at assemble and species level. Bifactoral ANOVA tests were applied to compare mean densities at both sampling sites and dates. Student-Newman-Keuls test was applied to compare pairs of means. Cluster and nMDS analyses were applied to evaluate between site similarities. The predominant species was Siderastrea siderea followed by S. radians and Porites astreoides. Recruit densities were similar among La Puntilla, Calle 16 and Acuario sites. Lower densities were always found in Malecón. Significant differences in mean stony recruit densities were found both between sites and sampling dates. The statistic analysis did not show significant spatial-temporal interactions. Malecón, the most polluted site, showed the lowest recruit density and the greater presence of species considered as indicators of organic pollution, sedimentation and abrasion. The density of recruit species was higher in October 2007 and lower in May 2008, and it was apparently influenced by reproduction and cold front seasons, respectively. The higher dominance and abundance of S. siderea, S. radians and P. astreoides recruits were apparently due to their high reproductive potential and resistance to disturbances. The identity of dominant species was apparently related to distance from major pollution sources. We highly recommended the replication of this research in other areas of the greater Caribbean region to test the generality of present results and to compare among areas. Future research should take into account the influence of other environmental factors, along with an appraisal of recruit species tolerance to these factors, to better ponder the effect of urban pollution on recruitment. Areas with well assessed pollution regimes are recommended for research.
Key words: stony corals, pollution, dominant species, density, recruitment.
Resumen
El efecto de la contaminación sobre el reclutamiento ha sido insuficientemente estudiado. Este trabajo trata el tema del reclutamiento en sitios ubicados a diferentes distancias de dos importantes fuentes de contaminación. Se caracterizó la composición y estructura de las asociaciones de reclutas de corales pétreos (escleractinios y milepóridos) al oeste de la Ciudad de La Habana, Cuba. Esta área está influenciada por la contaminación urbana del río Almendares y del emisario submarino construido en su desembocadura. El objetivo de esta investigación fue determinar las variaciones en la densidad y predominio de las especies de reclutas, en sitios ubicados a diferentes distancias de fuentes de contaminación, en distintas épocas de un año. Se muestreó cada tres meses entre julio 2007- mayo 2008. Se escogieron cuatro sitios de muestreo cerca del borde superior del escarpe del arrecife frontal a 10m de profundidad. Se utilizó un marco cuadrado de 25cm de lado de forma aleatoria-sistémica para determinar la densidad y composición taxonómica de reclutas de corales pétreos (colonias menores de 3cm). Para comparar las densidades medias entre sitios y fechas de muestreo se aplicó un análisis de varianza bifactorial. Para comparar los pares de densidades medias se empleó la prueba de Student-Newman-Keuls. La aplicación de un análisis Clasificación Numérica Jerárquica Aglomerativa (Cluster Analysis) y uno de ordenamiento por Escalado Multidimensional no Métrico (nMDS) mostró una distribución en la que los sitios quedaron agrupados en función de sus distancias de las fuentes contaminantes. Predominó Siderastrea sidérea seguida por Siderastrea radians y Porites asteroides. Durante el año de muestreo, las densidades de reclutas fueron similares entre La Puntilla, Calle 16 y Acuario. Las menores densidades se observaron siempre en Malecón. Se encontraron diferencias significativas entre las densidades medias de los sitios, así como entre de las distintas fechas de muestreo. El análisis de variancia bifactorial no mostró interacción significativa espacio–tiemporal. Malecón, sitio más contaminado, presentó menor reclutamiento y mayor presencia de especies consideradas indicadoras de contaminación orgánica, sedimentación y abrasión. Las densidades fueron mayores en Octubre 2007 y menores en Mayo 2008, al parecer influenciados por las épocas de reproducción y la acción de los frentes fríos respectivamente. El mayor predominio y abundancia de reclutas de S. siderea, S. radians y P. astreoides parece responder a sus elevados potenciales reproductivos y altas resistencias a disturbios. La identidad de las especies dominantes estuvo relacionada aparentemente con las distancias de las principales fuentes de contaminación. Se recomienda replicar esta investigación en otros lugares de la región del Caribe para probar la generalidad de estos resultados. Estudios futuros debieran tener en cuenta la influencia de otros factores ambientales, junto con una valoración de la tolerancia de las especies de reclutas a estos factores, para ponderar mejor el efecto de la contaminación urbana sobre el reclutamiento.
Palabras clave: corales pétreos, contaminación, especies dominantes, densidad, reclutamiento.
Los arrecifes coralinos constituyen el ecosistema marino de mayor diversidad biológica del mundo. Ellos poseen una incuestionable y reconocida importancia económica, social, ecológica y conservacionista (Spalding et al. 2001, Conservation International 2008). En años recientes se ha llegado a considerar que estos ecosistemas a nivel mundial viven bajo amenaza de extinción como consecuencia del calentamiento global y la acidificación de los océanos (Veron et al. 2009, Pandolfi et al. 2011).
Los corales pueden morir como resultado de incidentes naturales y de las actividades humanas. Si estos no son reemplazados a través de procesos de reproducción y reclutamiento, el arrecife puede eventualmente degenerar (Richmond 1997). Desafortunadamente, la mortalidad a gran escala en un arrecife de coral reduce su capacidad de autogenerarse, por lo que es importante que los corales con buena salud produzcan larvas abundantes y saludables que lleguen a los arrecifes degradados, se fijen al fondo y crezcan (Grimsditch & Salm 2006). Es por ello importante también identificar y proteger los arrecifes que funcionan como fuentes de larvas, así como conocer las corrientes marinas que los conectan con los arrecifes receptores (Nyström et al. 2000). Las poblaciones grandes y bien conectadas tienen más capacidad de evolucionar ante los cambios bruscos que sufre el ambiente, que las pequeñas y con poca conexión (Pandolfi et al. 2011).
La cobertura de coral vivo no constituye un reflejo de la salud del arrecife. Este valor solo describe el estado de este ecosistema en ese momento preciso. Por eso, es importante el conocimiento de los patrones de reclutamiento para la predicción del futuro del arrecife. Los corales adultos pueden sobrevivir en áreas donde la reproducción está fallando y las larvas son incapaces de fijarse (Richmond 1997).
La contaminación por aguas residuales es un problema creciente en los ambientes marinos tropicales. Las sustancias tóxicas acarreadas por ellas pueden inducir cambios metabólicos en los corales, disminuir sus tasas de crecimiento y reproductivas o reducir su viabilidad (Pastorok & Bilyard 1985).
El reclutamiento exitoso de nuevas colonias es muy importante para la recuperación de arrecifes degradados después de eventos de mortalidad. Las larvas de corales requieren sustratos relativamente no contaminados para su metamorfosis hacia pólipos sésiles y por consiguiente la creciente contaminación de las aguas costeras constituye un riesgo adicional para los arrecifes resilientes (Negri & Hoogenboom 2011).
Los corales pétreos, han sido utilizados como bio-monitores para la evaluación de la calidad ambiental, a través del estudio de la estructura de sus comunidades, debido a que como organismos bentónicos sésiles, no pueden emigrar de las áreas contaminadas y reflejan, por lo tanto, de modo natural, las características ambientales que han tenido lugar durante su desarrollo (Herrera-Moreno 1991). Por ello, estudiando sus características ecológicas puede evaluarse comparativamente la magnitud y extensión de los efectos de la contaminación. Connell & Miller (1984) plantean que los bioensayos y pruebas biológicas con organismos sensibles como los corales, son las herramientas apropiadas para determinar si la contaminación ambiental alcanza niveles que puedan interferir con procesos biológicos críticos como la reproducción y el reclutamiento.
En Cuba no se han realizado investigaciones sobre el comportamiento de la composición y estructura de las asociaciones de reclutas de corales pétreos a lo largo de gradientes de contaminación urbana. El objetivo de este trabajo fue determinar la existencia de variaciones en la densidad y predominio de las especies en el reclutamiento de corales pétreos en sitios ubicados a diferentes distancias de fuentes de contaminación, en distintas épocas de un año.
Materiales y métodos
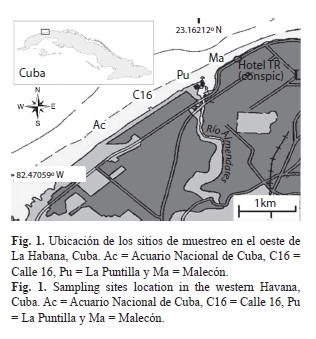
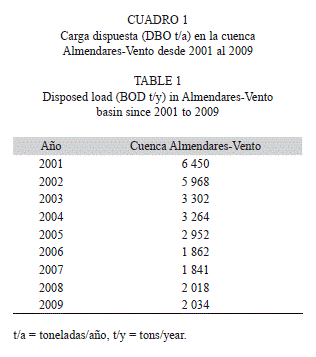
Descripción de la zona de estudio: El área de trabajo abarca un sector de la costa oeste del litoral de Ciudad de La Habana (Fig. 1). A lo largo de la costa, dentro de la cual se encuentra el área de estudio, se extiende un estrecho arrecife de tipo costero clasificado como tal por Zlatarski & Martínez-Estalella (1980). En el supralitoral se desarrollan actividades turísticas y recreativas. La mayor parte de esta área carece de sistemas de alcantarillado, por lo que las aguas de drenaje y los lixiviados de los desechos sólidos se descargan directamente sobre el sistema cárstico y son transportados hacia el mar o hacia el Río Almendares (Delgado & Miravet 2009). Según datos del Instituto de Geofísica y Astronomía-Delegación del CITMA, y el Instituto Nacional de Recursos Hidráulicos (INRH) a este último tributan 89 fuentes de contaminantes. En el cuadro 1 se presenta la carga dispuesta (DBO= Demanda Biológica de Oxígeno) desde el 2001 hasta el 2009 (datos del Centro de Información y Gestión Ambiental (CIGEA).
Se muestrearon cuatro sitios cerca del borde superior del escarpe del arrecife frontal. Se trabajó a 10m de profundidad siguiendo el criterio de Herrera-Moreno (1990), quien plantea que es el más adecuado para comparar el efecto de la contaminación. Este autor refiere que a menos de 10m de profundidad intervienen otros factores como las disminuciones superficiales de la salinidad por los drenajes urbanos, el oleaje, entre otros, y a mayores profundidades tiende a disminuir el efecto de la contaminación debido a un aumento en la dilución de los contaminantes. A continuación se describe cada sitio de muestreo.
Acuario (23.125480” N - 082.432830” O): Este sitio se considera como el más limpio entre los cuatro porque se encuentra más alejado del Río Almendares (principal fuente de contaminantes), y porque los residuales urbanos de la zona no se vierten directamente al mar sino que van al colector principal de aguas residuales que desemboca en el emisario submarino ubicado en La Puntilla, aproximadamente a 3km al este (Delgado & Miravet 2009). En este sitio se aprecia bastante transparencia en el agua, se observa turbidez sólo después de la presencia de fuertes lluvias o en períodos de oleaje intenso, fundamentalmente, durante la temporada de los fuertes vientos que acompañan a los eventos de “frentes fríos” que tienen lugar entre los meses de octubre y febrero, conocidos también en Cuba como “nortes”.
La comunidad coralina adulta está constituida fundamentalmente por colonias de pequeño y mediano tamaño de las especies Siderastrea siderea, Porites astreoides, Montastraea cavernosa y Agaricia agaricites.
Calle 16 (23.131820” N - 082.423670” W): Sitio con características parecidas y hacia el este del anterior. Se encuentra más cercano al Río Almendares, donde se vierten grandes cantidades de desechos y del emisario submarino construido en su desembocadura. Recibe de forma muy esporádica la influencia de dicho río, por lo que se puede considerar como relativamente limpio (Guardia & Gonzáles-Sansón 2000), aunque menos que el anterior. La comunidad de corales adultos es similar a la del sitio Acuario.
La Puntilla (23.135690” N - 082.416370” W): Recibe influencia del Río Almendares y del emisario submarino antes mencionado, aunque no de forma tan directa e intensa debido a su posición con respecto a la dirección de salida del agua del río. Se apreció turbidez en el agua durante todos los muestreos. La comunidad de corales adultos está formada fundamentalmente por S. siderea y S. radians.
Malecón (23.140290” N - 082.408300” W): Se localiza inmediatamente al este de la desembocadura del Río Almendares y del emisario submarino que recibe los residuales que transporta el colector principal del alcantarillado del Municipio Playa, y cuya descarga ocurre en este lugar, a una distancia aproximada de 600m de la costa (Delgado & Miravet 2009). Este emisario submarino sufrió una fractura total a sólo 12m de profundidad en la misma desembocadura del río, además presenta varias fugas de desechos por las uniones de los tubos.
La gran cercanía y el hecho de que se encuentre directamente en la misma dirección de la salida del agua del Río Almendares, sugiere que este sitio recibe mayor influencia de las aguas provenientes de ambas fuentes contaminantes. Herrera-Moreno & Martínez–Estalella (1987), Alcolado & Herrera-Moreno (1987) y Alcolado (2007), basados en estudios sobre comunidades de corales, octocorales y esponjas consideran a Malecón como el sitio más contaminado entre los muestreados. Durante todos los muestreos se observó una gran turbidez en el agua. La comunidad de corales adultos es similar a la de La Puntilla.
Métodos de muestreo: Los muestreos serealizaron mediante buceo autónomo (SCUBA) cada tres meses a partir de mediados del 2007 (julio y octubre 2007 y febrero y mayo 2008), para determinar las variaciones en la densidad y predominio de las especies de reclutas de corales. La estructura y composición de las asociaciones de reclutas de corales deben variar a lo largo del año en parte debido a que en esa etapa temprana de la vida son más susceptibles de morir por eventos climatológicos (huracanes y frentes fríos muy comunes en Cuba), pulsos de descargas de contaminantes a través de los ríos en las épocas de lluvias, entre otros y por otro lado los eventos reproductivos actúan aumentando sus densidades.
Se utilizó el marco cuadrado de 25cm de lado como unidad de muestreo para determinar la densidad y el predominio de las especies de reclutas (corales menores de 3cm) para cada sitio y durante cada muestreo. Se trabajó precisamente con reclutas que pudieron ser identificados hasta los niveles de género o especie, asentados en meses o años anteriores así como los que durante el tiempo de muestreo llegaron a adquirir un tamaño suficientemente grande como para ser visibles a simple vista y provocar variaciones en las densidades, estos últimos probablemente asentados antes y durante el período (un año) de muestreo. Para la ubicación aleatoria-sistemática de las unidades de muestreo se extendió una cinta métrica a lo largo de 160m paralelamente a la costa. En cada decena de metros se escogieron cinco puntos al azar donde se colocó el marco (80 marcos por sitio en total). Todas las unidades de muestreo se colocaron sobre sustrato rocoso coralino. Los corales fueron identificados in situ hasta la categoría taxonómica de especie según los criterios taxonómicos de Beltrán-Torres & Carricart-Ganivet (1999).
Procesamiento de datos: Kramer (2003), sugiere que 40 marcos de 25cm de lado es suficientemente representativo para estimar la densidad de reclutamiento. Teniendo en cuenta que en lugares contaminados son de esperar densidades muy bajas, se tomó el doble de esa cantidad de muestras para poder representar mejor la estructura de las asociaciones de especies de reclutas.
Para evaluar la similitud entre los sitios se realizó un análisis de Clasificación Numérica Jerárquica Aglomerativa (Cluster Analysis) y uno de ordenamiento de Escalado Multidimencional no Métrico (nMDS) mediante el coeficiente de Similitud de Bray Curtis.
Se le aplicó a la matriz de datos (transformada a raíz cuadrada) de predominio de reclutas. Los cálculos y gráficos se realizaron mediante el programa PRIMER, versión 5 (Clarke & Warwick 2001).
A los datos de densidad de reclutas se les comprobó la normalidad y homogeneidad de varianza. En los casos en que cumplieron estas premisas se realizó un análisis de varianza bifactorial (ANOVA) para conocer la existencia de diferencias significativas entre sitios en correspondencia con la fecha de muestreo. En los casos en que no se cumplieron, los datos se transformaron según fue necesario a partir de la relación propuesta por Taylor (1961) según la metodología de Elliot (1971). Para determinar las diferencias entre los pares de media se utilizó la prueba de Student- Newman-Keuls (SNK), con índice de significación de 0.05. Para estos cálculos se utilizó el programa Statistica 6.0 (StatSoft Inc. 2002).
Resultados
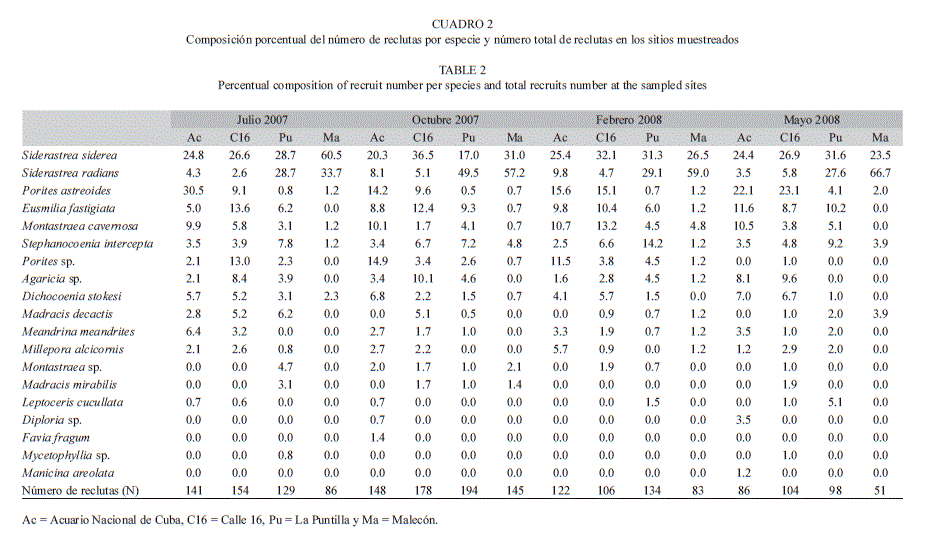
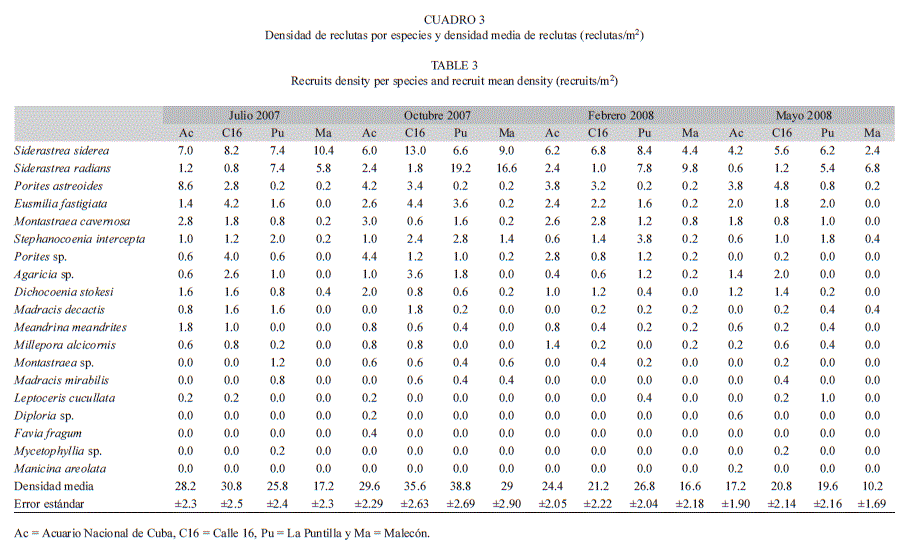
Predominio de especies de reclutas: La especie predominante fue S. siderea seguida por S. radians. Esta última ocupó el primer lugar en dominancia en todos los muestreos, solamente en La Puntilla y Malecón (sitios más cercanos a las fuentes de contaminación). P. asteroides se destacó en los sitios Acuario y Calle 16 en todos los muestreos (sitios más alejados de las fuentes de contaminantes). M. cavernosa mostró mayor predominio que S. radians en Acuario en todos los muestreos (Cuadro 2).
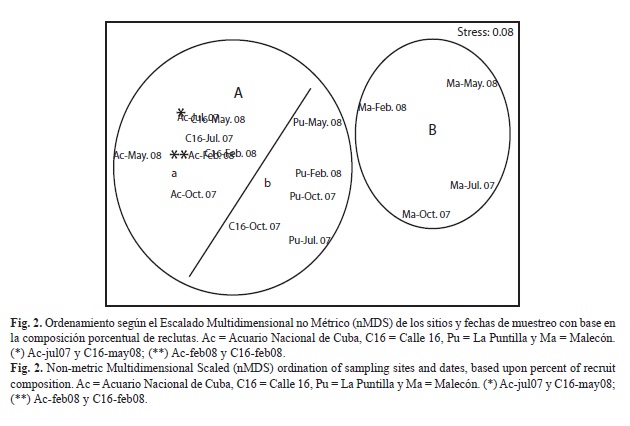
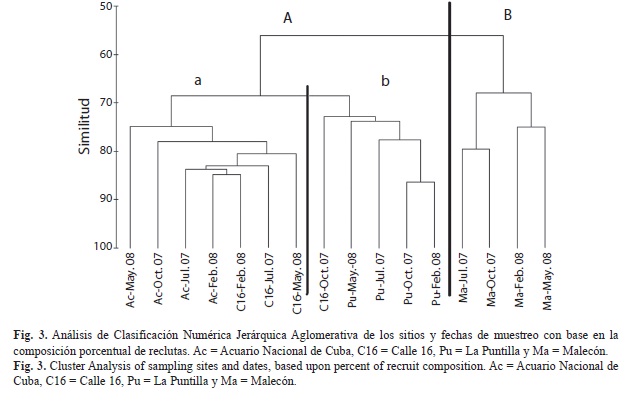
Clasificación y ordenamiento: Tanto el análisis de Clasificación Numérica Jerárquica Aglomerativa, como el de ordenamiento por Escalado Multidimensional no Métrico (nMDS) mostraron distribuciones en la que los sitios quedaron agrupados en función con sus distancias de las fuentes contaminantes. Quedaron definidos dos grupos: uno formado por los cuatro muestreos de Malecón (B, sitio más cercano a las fuentes de contaminantes), y otro, por el resto de los sitios (más alejados del efecto de las fuentes de contaminación). Dentro de este segundo grupo se formaron dos subgrupos. En el primero (a) se observó mayor afinidad entre los sitios del Acuario y Calle 16 en todas las fechas de muestreo excepto en octubre 2007, en que Calle 16 mostró más similitud con el grupo de La Puntilla (b) (Fig. 2 y 3).
Densidad de reclutas: Durante el año de muestreo, las densidades de reclutas fueron similares entre La Puntilla, Calle 16 y Acuario. Las menores densidades se observaron siempre en Malecón. Las especies más abundantes en el área fueron: S. siderea, seguido de S. radians y P. asteroides. S. radians mostró valores muy superiores en Malecón y La Puntilla en comparación con los observados en el resto de los sitios menos influenciados por la contaminación proveniente del río Almendares y el emisario submarino. Por su parte, P. astreoides fue más abundante hacia los sitios Calle 16 y Acuario, más alejados de estas fuentes de contaminación (Cuadro 3).
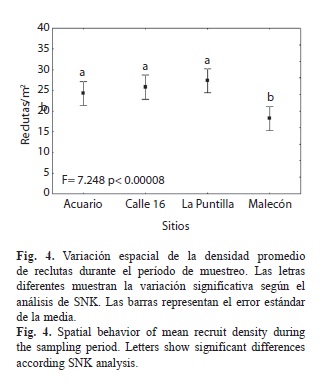
Se encontraron diferencias significativas entre las densidades medias de los sitios, así como entre de las distintas fechas de muestreo. Entre sitios, la prueba de SNK no mostró diferencias significativas entre Acuario, Calle 16 y La Puntilla (valores entre 25-27reclutas/m2). Sin embargo, estos registros fueron significativamente inferiores en Malecón, sitio más impactado (Fig. 4).
El mayor predominio y abundancia de reclutas de S. siderea, S. radians y P. astreoides en todos los sitios del área de estudio y fundamentalmente las dos primeras especies en Malecón y La Puntilla, parece estar determinado en parte porque estas son especies oportunistas de rápido crecimiento y resistentes al oleaje y la sedimentación (Hubbard & Pocock 1972, Kolehmainen 1973, Torres & Morelock 2002).
El mayor predominio de reclutas de P. astreoides en Acuario en julio 2007 pudo estar determinado por un fuerte evento reproductivo de esta especie en los primeros meses del año, época en la cual según Birkeland (1997) se reproduce esta especie. Según Szmant (1986) esta especie se reproduce entre los meses de enero y septiembre. La disminución en su dominancia en octubre 2007 (que se mantuvo hasta mayo 2008) pudiera estar relacionada con eventos de mortalidad asociados al efecto del oleaje provocado por la llegada de los primeros frentes fríos.
Particularmente, P. astreoides ha sido señalada como una especie con gran potencial reproductivo y comparativamente poco dañada por los huracanes (Glynn et al. 1964, Lirman & Fong, 1996, 1997). Según Szmant (1986) la estrategia reproductiva de esta especie es la anidación o “brooding” en lugar de la diseminación de gametos y el desarrollo externo de las larvas. Las plánulas liberadas son capaces de asentarse enseguida que son liberadas y pueden colonizar áreas cercanas a las de los progenitores. Esta estrategia es característica de especies de colonias pequeñas, con múltiples ciclos reproductivos en el año y altas tasas de reproducción; y generalmente habitan en ambientes inestables. Por su parte, Martínez-Estalella & Herrera-Moreno (1989) plantean que esta especie está mejor representada en zonas de embate y en terrazas rocosas abrasivas donde el oleaje es constante y que es una especie oportunista (estratega r) apoyándose en Duyl (1985), quien refiere que gasta más energía en su reproducción sexual que en su supervivencia y mantenimiento. Alcolado et al. (2009) observó en crestas del sur y este del Golfo de Batabanó que P. astreoides aumentó su predominio a expensas de Acropora palmata después de la mortalidad de esta última especie causada por los huracanes en el período 2001-2006. Lo mismo fue observado por González-Ferrer et al. (2007) en el Archipiélago Sabana-Camagüey, donde las crestas se han convertido en bajos rocosos y han pasado a predominar otras especies como P. astreoides.
Herrera-Moreno (1990) refiere que en las zonas de descarga de aguas residuales y al oeste de la Bahía de La Habana las condiciones de sedimentación y turbidez exceden la tolerancia de la mayor parte de las especies, razón por la cual sólo es posible la existencia de S. radians por su resistencia extrema. Al igual que P. astreoides también se reproduce mediante “brooding” (Szmant 1986, 1991). Es considerada como muy resistente a la contaminación (Herrera-Moreno & Martínez–Estalella 1987) y a la sedimentación (Cortés & Risk 1985). Lewis (1989) plantea que es una de las especies más tolerantes en el Caribe, encontrada a menudo en localidades de mucho disturbio. Según Lirman (2002), posee una alta resistencia a salinidades bajas y enterramientos por sedimentos, además de una gran capacidad de recuperación; condiciones que deben ser frecuentes cerca del Río Almendares, sobre todo después de lluvias fuertes.
Hay que tener en cuenta que Herrera-Moreno & Martínez-Estalella (1987), siguiendo los criterios de Zlatarski & Martínez-Estalella (1980), consideraron a S. radians como la especie indicadora de contaminación, concibiendo a S. siderea como una forma de ésta. El predominio de reclutas de S. radians y S. sidérea en los sitios más cercanos al río Almendares en el presente trabajo refuerza, desde la óptica de una etapa más temprana del desarrollo de los corales, el criterio de Herrera-Moreno & Martínez-Estalella (1987) acerca de la resistencia a la contaminación urbana de ambas especies incluidas en S. radians (sensu latu).
Estudios anteriores de Alcolado et al. (2003, 2010), Caballero et al. (2004) y González-Ferrer (2004) en diferentes arrecifes de Cuba alejados de asentamientos y fuentes de contaminación urbana coinciden con el presente en que S. siderea y P. astreoides están entre las especies de reclutas más predominantes, independiente de la calidad del agua. Por otra parte, a diferencia de lo observado en el oeste de Ciudad de La Habana, en estas regiones muy alejadas de zonas urbanas codominaron, junto con ambas especies, otras consideradas por Alcolado et al. (2010), Alcolado et al. (2003) y Caballero et al. (2004), como poco tolerantes a los sedimentos y a la contaminación.
Las densidades de reclutas observadas durante el período de muestreo en toda el área de estudio se pueden considerar como altas. Estos valores fueron mayores que los reportados por otros autores (Alcolado et al. 2010, Espinosa et al. 2002, Alcolado et al. 2003, Caballero et al. 2005,2007, González-Ferrer et al. 2003) en otras localidades de Cuba alejadas de zonas urbanas. Esto podría deberse al predominio de especies de corales que según Gonzáles-Ferrer (2004) tienen una alta tasa de reclutamiento (S. siderea, S. radians y P. astreoides), además de que dos de ellas son incubadoras (las dos últimas). En efecto, Szmant (1986) plantea que la fuerza que radica detrás de la evolución de la incubación por parte de especies que habitan en ambientes inestables y que sufren grandes tasas de mortalidad es lograr una alta tasa de reclutamiento local. Moulding (2005) reporta valores similares a los de este estudio en algunos arrecifes de los Cayos de la Florida. Por su parte Glassom & Chadwick (2006) reportan densidades muy superiores (42-171 reclutas/m2) en arrecifes del Mar Rojo localizados a diferentes distancias de fuentes de contaminación.
Las densidades de reclutas, siempre significativamente menores en Malecón, probablemente estén determinadas por el efecto restrictivo impuesto por la cercanía a la desembocadura del río Almendares. Su ubicación justamente en la misma dirección de la salida del agua del río, implicaría una mayor influencia de sus aguas contaminadas. Por otra parte, el emisario submarino presenta una fractura total a pocos metros de este sitio. Guardia & González-Sansón (2000) encontraron resultados similares al comparar sitios ubicados en la Caleta de San Lázaro (muy cercanos a la muy contaminada Bahía de La Habana y orientados a lo largo de la dirección de la salida del agua de ésta) con otros lejos al oeste y menos contaminados.
El hecho de que el predominio de reclutas de P. astreoides haya sido menor en los sitios más cercanos a las fuentes contaminantes posiblemente esté asociado a una aparente menor resistencia a la contaminación y a salinidades episódicamente bajas. También podría estar influyendo el alto cubrimiento del fondo por el alga Lobophora sp. observado en estos sitios, la que, según Kuffner et al. (2006), puede provocar tanto inhibición del reclutamiento como respuestas de rechazo por parte de las larvas de esta especie de coral. González-Ferrer (2004) plantea que además de esta macroalga parda, otras como Dictyota spp. y Sargassum spp. (que estuvieron entre las más representadas en el área de estudio) se consideran fuertes competidoras con los corales por el sustrato; impidiendo en muchos casos la fijación de los reclutas, ocasionando daños en los bordes de las colonias, o incluso, estableciéndose sobre éstas hasta asfixiarlas.
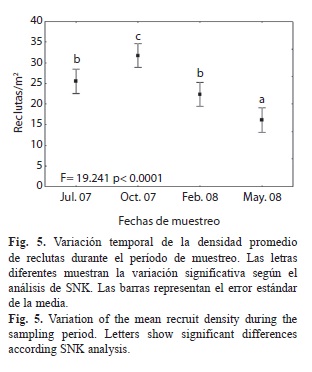
El análisis de la densidad de reclutas en el tiempo sugirió un evento de mayor reclutamiento de las especies más abundantes en el área antes de octubre 2007, lo que coincide con lo planteado por Birkeland (1997) acerca de que la mayoría se reproducen en las estaciones de primavera y verano. La caída de la densidad observada posteriormente sugiere una eliminación de reclutas por el oleaje generado por la temprana llegada de los frentes fríos, los que según González-Ferrer (2004) afectan el reclutamiento por el aumento del efecto de la sedimentación, aunque no puede descartarse el efecto abrasivo de sedimentos y rocas movidos por el oleaje.
A la luz del presente trabajo no se refleja claramente que S. siderea, tratada ya como una especie independiente, esté indicando mayores niveles de contaminación en las estaciones más cercanas al río Almendares y al emisario submarino. La explicacion más parsimoniosa podría ser que el reclutamiento en esta especie no se ve afectado por la influencia de las diferentes distancias de los sitios al río Almendares y el emisario submarino. Por otra parte, puede que haya disminuido la carga de contaminantes desde los muestreos de los autores antes citados producto de la construcción de una planta para depurar los residuales de zonas urbanas e industrias que históricamente han contaminado el río (Michel Alcolado-Silva, com. pers. Ingeniero hidráulico involucrado en el diseño del emisario submarino del río Almendares), al punto de no hacer gran diferencia en el predominio de esta especie euritópica en los cuatro sitios. También es posible que el paso cercano de los cinco huracanes que afectaron el área en 2004 (Charley) y 2005 (Dennis, Rita, Wilma y Katrina) hayan impuesto una dominancia más generalizada de esta especie en este sector litoral, y disminuido las posibles diferencias relacionadas con el efecto puntual del río y del emisario. Sin embargo, estas hipótesis merecen ser sometidas a prueba por separado o en conjunto.
Dada la cercana presencia del río Almendares, en eventos de fuertes lluvias pueden producirse pulsos agudos de baja salinidad en Malecón y en menor grado en La Puntilla, cuyo efecto sobre los reclutas no pueden evaluarse en el presente trabajo y que en cierta medida pudieran afectarlos, atendiendo a los criterios de Jokiel (1985) y Richmond (1997) sobre la sensibilidad de los gametos y las plánulas a ese factor. Teniendo en cuenta los trabajos de Jokiel (1985) y Tomascik & Sander (1987), esta consideración también sería válida para la posible influencia de la persistente turbidez del agua observada durante los muestreos. Estos son aspectos que merecen consideración en un estudio futuro en el área muestreada, junto con una valoración de la tolerancia de las especies de reclutas a estos factores.
Por otro lado, la forma de distribución del análisis de Clasificación Numérica Jerárquica Aglomerativa y el nMDS, aplicado a los datos de composición porcentual de reclutas, demuestra la utilidad de la estructura de las asociaciones de reclutas como indicadoras alternativas o complementarias en la discriminación de niveles de disturbio ambiental, aspecto que ha sido referido por Connell & Miller (1984) quienes plantean que mediante el conocimiento de la biología de la reproducción de los corales y del reclutamiento, es fácil ver cómo la calidad ambiental puede afectar estos dos procesos que son responsables de la persistencia del arrecife.
Se recomienda replicar esta investigación a otros lugares de la región del Caribe para probar la generalidad de estos resultados. Estudios futuros debieran tener en cuenta la influencia de otros factores ambientales, junto con una valoración de la tolerancia de las especies de reclutas a estos factores, para ponderar mejor el efecto de la contaminación urbana sobre el reclutamiento.
Agradecimientos
Agradecemos a los buzos del Centro de Investigaciones Marinas de la Universidad de La Habana, Cuba, por su apoyo incondicional, por estar siempre disponibles y dispuestos a ayudarnos con todo lo necesario para los muestreos.
Referencias
Alcolado, P.M. 2007. Reading the code of coral reef sponge community composition and structure for environmental biomonitoring: some experiences from Cuba, p. 3-10. In M.R. Custódio, G. Lobo-Hajdu, E. Hajdu & G. Muricy (eds.). Porifera Research: Biodiversity, Innovation and Sustaniability, Rio de Janeiro: Museu Nacional. [ Links ]
Alcolado, P.M. & A. Herrera-Moreno. 1987. Efectos de la contaminación sobre las comunidades de esponjas en el Litoral de La Habana, Cuba. Rep. Invest. Inst. Oceanol., Academia Ciencias de Cuba 68: 1-17. [ Links ]
Alcolado, P.M., I.E. Morgan, P.A. Kramer, R.N. Ginsburg, P. Blanchon, E. de la Guardia, V. Kosminin, S. González-Ferrer & M. Hernández. 2010. Condition of remote reefs off southwest Cuba. Ciencias Marinas 36: 179-197. [ Links ]
Alcolado, P.M., B. Martínez-Daranas, G. Menéndez-Macía, R. del Valle, M. Hernández & T. García. 2003. Rapid assessment of coral communities of María la Gorda, southeast Ensenada de Corrientes, Cuba (part 1: stony corals and algae), p. 268-277. In J.C. Lang (ed.). Status of Coral Reefs in the Western Atlantic: Results of initial surveys, Atlantic and Gulf Rapid Reef Assessment (AGRRA) Program. Atoll Res. Bull. [ Links ]
Alcolado, P.M., D. Hernández-Muñoz, H. Caballero, L. Busutil, S. Perera & G. Hidalgo. 2009. Efectos de un inusual período de alta frecuencia de huracanes sobre el bentos de arrecifes coralinos. REVMAR. 1:74-94. [ Links ]
Beltrán-Torres, A.U. & J.P. Carricart-Ganivet. 1999. Lista revisada y clave para los corales pétreos zooxantelados (Hidrozoa: Milleporina; Anthozoa: Scleractinia) del Atlántico mexicano. Rev. Biol. Trop. 47: 813-829. [ Links ]
Birkeland, C. 1997. Life and death of coral reefs. Chapman & Hall, Nueva York, Nueva York, EEUU. [ Links ]
Caballero, H., G. Varona & Y. García. 2004. Estructura ecológica de las comunidades de corales de la costa oriental de Bahía de Cochinos, Cuba. Rev. Invest. Mar. 25: 23-36. [ Links ]
Caballero, H., D. Rosales & A. Alcalá. 2005. Estudio diagnóstico del arrecife coralino del Rincón de Guanabo, Ciudad de La Habana, Cuba. 1. Corales, esponjas y gorgonáceos. Rev. Invest. Mar. 26: 207-217. [ Links ]
Caballero, H., S. González-Ferrer, D. Cobián, S. Álvarez & P. Alcolado-Prieto. 2007. Evaluación AGRRA del bentos de 10 sitios de buceo de María La Gorda, Bahía de Corrientes, Cuba. Rev. Invest. Mar. 28:131-138. [ Links ]
Clarke, K.R. & R.M. Warwick. 2001. Change in marine communities: an approach to statistical analysis and interpretation, 2nd edition. PRIMER-E: Plymouth. [ Links ]
Connell, D.W. & G.J. Miller. 1984. Chemistry and ecotoxicology of pollution. Wiley, Nueva York, Nueva York, EEUU. [ Links ]
Cortés, J. & M.J. Risk. 1985. A reef under siltation stress: Cahuita, Costa Rica. Bull. Mar. Sci. 36: 339-356. [ Links ]
Conservation International. 2008. Economic Values of Coral Reefs, Mangroves and Seagrasses. A Global Compilation. Center for Applied Biodiversity Science. Conservation International, Arlington, Virginia, USA. [ Links ]
Delgado, Y. & M.E. Miravet. 2009. Metodología para la evaluación de riesgos sanitarios ante la contaminación fecal. Serie Oceanológica 5: 74-87. Inst. Oceanol. Ciudad de La Habana, Cuba. (Consultado: Diciembre 12, 2009, http://www.oceanologia.redciencia.cu). [ Links ]
Duyl, F.C. 1985. Atlas of the living reefs of Curaçao and Bonaire (Netherland Antilles). Nature. Studiekring Suriname un der Nederlandse Antillen, Utrecht, Holland. [ Links ]
Elliot, J.M. 1971. Some methods of the statistical analysis of samples of benthic invertebrates. Freshwater Biol. Ass. Sci. 25: 144. [ Links ]
Espinosa-Sáez, J., F. Pina-Amargós, K. Cantelar-Ramos, S. González-Ferrer, P.M. Alcolado, M. Hernández-González & J.L. Hernández-López. 2002. Evaluación del estado de salud de los arrecifes coralinos del Archipiélago Sabana -Camagüey (2001). Informe del Proyecto PNUD/GEF Sabana-Camagüey (CUB/98/G32). [ Links ]
Glynn, P.W., L.R. Almodóvar & J.G. González. 1964. Effects of Hurricane Edith on marine life in La Parguera; Puerto Rico. Caribb. J. Sci. 4: 335-345. [ Links ]
Glassom D. & N.E. Chadwick. 2006. Recruitment, growth and mortality of juvenile corals at Eilat, Northern Red Sea. Mar. Ecol. Prog. Ser. 318: 111-122. [ Links ]
González-Ferrer, S., Z. Marcos, A. Jiménez, H. Caballero, P.M. Alcolado & M. Hernández. 2003. Evaluación del estado de salud del bentos de los arrecifes coralinos del Archipiélago Sabana-Camagüey, Cuba. Informe Preliminar de la Expedición CUBAGRRA 2003. [ Links ]
González-Ferrer, S. 2004. Catálogo de corales hermatípicos de aguas cubanas, p. 318. In S. González-Ferrer (ed.). Corales pétreos. Jardines sumergidos de Cuba. Inst. Oceanol. Editorial Academia, La Habana, Cuba. [ Links ]
González-Ferrer, S., K. Cantelar, F. Pina, A. Jiménez, J. Espinosa, M. Hernández & J.L. Hernández. 2007. Estado de los ecosistemasmarinos y costeros, y algunas características ambientales y tendencias, p. 38-45. In P.M. Alcolado, E.E. García & M. Arellano-Acosta (eds.). Ecosistema Sabana-Camagüey. Estado actual, avances y desafíos en la protección y uso sostenible de la biodiversidad. Proyecto PNUD/GEF Sabana-Camagüey. CUB/98/G32; CUB/99/G81. Editorial Academia, Cuba. [ Links ]
Grimsditch, G.D. & R.V. Salm. 2006. Coral Reef Resilience and Resistance to Bleaching. UICN, Gland, Switzerland. [ Links ]
Guardia, E. & G. Gonzáles-Sansón. 2000. Asociaciones de corales, gorgónias y esponjas del sublitoral habanero al oeste de la Bahía de la Habana, I. Gradiente ambiental. Rev. Invest. Mar. 21: 1-8. [ Links ]
Herrera-Moreno, A. 1990. Efectos de la contaminación sobre las caracteristicas de las comunidades bentónicas: el arrecife coralino. Tesis Doctorado, Instituto de Oceanología, Academia de Ciencias de Cuba, Cuba. [ Links ]
Herrera-Moreno, A. 1991. Efectos de la contaminación sobre la estructura ecológica de los arrecifes coralinos en el litoral habanero. Tesis Doctorado, Academia de Ciencias de Cuba, Cuba. [ Links ]
Herrera-Moreno, A. & N. Martínez-Estalella. 1987. Efectos de la contaminación sobre las comunidades de corales escleractíneos al Oeste de la Bahía de la Habana. Rep. Inv. Inst. Oceanol., Academia de Ciencias de Cuba 62: 1-29. [ Links ]
Hubbard, J.A. & Y.P. Pocock. 1972. Sediment Rejection by recent scleractinian corals: A key to paleoenvironmental reconstruction. Geol. Rundschuge 61: 598-626. [ Links ]
Jokiel, P.L. 1985. Lunar periodicity of planula release in the reef coral Pocillopora damicornis in relation to various environmental factors. Proc. 5th Internat. Coral Reef Symposium. Tahiti 4: 307-312. [ Links ]
Kolehmainen, S.E. 1973. Siltation experiments on corals in situ. Puerto Rico. Final Report Mission 8. PRINUL. [ Links ]
Kramer, P. 2003. Synthesis of coral reef health indicators for the western Atlantic: Results of the AGRRA Program (1997-2000). In J.C. Lang (ed.). Status of Coral Reefs in the Western Atlantic: Results of Initial Surveys, Atlantic and Gulf Rapid Reef Assessment (AGRRA) Program. Atoll Res. Bull. 496: 1-57. [ Links ]
Kuffner, I.B., L.J. Walters, M.A. Becerro, V.J. Paul, R. Ritson-Williams & K.S. Beach. 2006. Inhibition of coral recruitment by macroalgae and cyanobacteria. Mar. Ecol. Prog. Ser. 323: 107-117. [ Links ]
Lewis, J.B. 1989. Spherical growth in the Caribbean coral Siderastrea radians (Pallas) and its survival in disturbed habitats. Coral Reefs 7: 161-7. [ Links ]
Lirman, D. & P. Fong. 1996. Sequential storms cause zone specific damage on a reef in the northern Florida reef tract: evidence from Hurricane Andrew and the 1993 Storm of the Century. Fla. Sci. 59: 50-63. [ Links ]
Lirman, D. y P. Fong. 1997. Susceptibility of coral communities to storm intensity, duration and frequency. Proceedings of the 8th International Coral Reef Symposium 1: 561-566. [ Links ]
Lirman, D. 2002. Competition between macro-algae and corals: effects of herbivore exclusion and increased algal biomass on coral survivorship and growth. Coral Reefs 19: 392-399. [ Links ]
Martínez-Estalella, N. & A. Herrera-Moreno. 1989. Estudio de la comunidad coralina del arrecife del Rincón de Guanabo, Cuba. Rep. Invest. Inst. de Oceanol., Academia de Ciencias de Cuba 9: 24. [ Links ]
Moulding, A.L. 2005. Coral recruitment patterns in the Florida Keys. Rev. Biol. Trop. 53: 75-82. [ Links ]
Negri, A.P. & M.O. Hoogenboom. 2011. Water Contamination Reduces the Tolerance of Coral Larvae to Thermal Stress. PLoSONE 6(5): e19703. doi:10.1371/journal. pone.0019703. [ Links ]
Nyström, M., C. Folke & F. Moberg. 2000. Coral reef disturbance and resilience in a human-dominated environment. Trends Ecol. Evolut. 15: 413- 417. [ Links ]
Pandolfi, J.M., S.R. Connolly, D.J. Marshall & A.L. Cohen. 2011. Projecting Coral Reef Futures Under Global Warming and Ocean Acidification. Science 333: 418-422. [ Links ]
Pastorok, R.A. & G.R. Bilyard. 1985. Effects of sewage pollution on coral-reef communities. Mar. Ecol. Prog. Ser. 21:175-189. [ Links ]
Richmond, R.H. 1997. Reproducción and recruitment in corals: critical links in the persistence of reef. p. 175-197. In C. Birkeland (ed.). Life and death of coral reefs. Chapman & Hall, Nueva York, Nueva York, EEUU. [ Links ]
Spalding, M.D., C Ravilious & E.P Green. 2001. World Atlas of Coral Reefs. Prepared at the UNEP World Conservation Monitoring Centre. University of California, Berkeley, EEUU. [ Links ]
StatSoft, Inc. 2002. STATISTICA 6.0 for Windows (Computer Program Manual). StatSoft, Tulsa, Oklahoma, EEUU. [ Links ]
Szmant, A.M. 1986. Reproductive ecology of Caribbean reef corals. Corals reefs 5: 43-54. [ Links ]
Szmant, A.M. 1991. Sexual reproduction by the Caribbean reef corals Montastrea annularis and M. cavernosa. Mar. Ecol. Prog. Ser.,74:13-25. [ Links ]
Taylor, W.R. 1961. Aggregation variance and the mean. Nature 189: 732-735. [ Links ]
Tomascik, T. & F. Sander. 1987. Effects of eutrophication on reef-building corals III. Reproduction of the reefbuilding coral Porites porites. Mar. Biol. 94: 77-94. [ Links ]
Torres, J.L. & J. Morelock. 2002. Effect of terrigenous sediment influx on coral cover and linear extension rates of three Caribbean massive coral species. Caribb. J. Sci. 38: 222-229. [ Links ]
Veron, J.E.N., O. Hoegh-Guldberg, T.M. Lenton, J.M. Lough, D.O. Obura, P. Pearce-Kelly, C.R.C. Sheppard, M. Spalding, M.G. Stafford-Smith & A.D. Rogers. 2009. The coral reef crisis: The critical importance of <350 ppm CO2. Mar. Pollut. Bull. 58: 1428-1436. [ Links ]
Zlatarsky, V. & N. Martínez-Estalella. 1980. Scleractinians of Cuba, with data on associated organisms. Bulgarian Academy of Sciences, Sofia, Bulgaria. [ Links ]
*Correspondencia:
Pedro Alcolado-Prieto: Instituto de Oceanología. Calle 1ra No. 18406, Playa, CP 12100, Ciudad de La Habana, Cuba. palcolado@oceano.inf.cu
Hansel Caballero Aragón: Acuario Nacional de Cuba. Calle 1ra y 60. Playa, CP 11300, Ciudad de La Habana, Cuba; hanselc@acuarionacional.cu
Pedro M. Alcolado: Instituto de Oceanología. Calle 1ra No. 18406, Playa, CP 12100, Ciudad de La Habana, Cuba. alcolado@ama.cu
Alexander Lopeztegui Castillo: Centro de Investigaciones Pesqueras, Ministerio de la Industria Alimentaria. Ave 5ta y 246, edificio 2, Barlovento, Jaimanitas; sasha@cip.telemar.cu
1. Instituto de Oceanología. Calle 1ra No. 18406, Playa, CP 12100, Ciudad de La Habana, Cuba; palcolado@oceano.inf.cu, alcolado@ama.cu
2. Acuario Nacional de Cuba. Calle 1ra y 60. Playa, CP 11300, Ciudad de La Habana, Cuba; hanselc@acuarionacional.cu
3. Centro de Investigaciones Pesqueras, Ministerio de la Industria Alimentaria. Ave 5ta y 246, edificio 2, Barlovento, Jaimanitas; sasha@cip.telemar.cu
Recibido 11-VII-2011. Corregido 10-III-2012. Aceptado 17-IV-2012.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}