










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.60 n.3 San José Sep. 2012
Limnology in El Dorado: some surprising aspects of the regulation of phytoplankton productive capacity in a high-altitude Andean lake (Laguna de Guatavita, Colombia)
*Dirección para correspondencia
Abstract
High-altitude mountain lakes remain understudied, mostly because of their relative inaccessibility. Laguna de Guatavita, a small, equatorial, high-altitude crater lake in the Eastern Range of the Colombian Andes, was once of high cultural importance to pre-Columban inhabitants, the original location of the legendary El Dorado. We investigated the factors regulating the primary production in Laguna de Guatavita (4°58’50” N - 73°46’43” W, alt. 2 935m.a.s.l., area: 0.11km2, maximum depth: 30m), during a series of three intensive field campaigns, which were conducted over a year-long period in 2003-2004. In each, standard profiles of temperature, oxygen concentration and light intensity were determined on each of 16-18 consecutive days. Samples were collected and analysed for chlorophyll and for biologically-significant solutes in GF/F-filtered water (NH4+, NO3- , NO2-; soluble reactive phosphorus). Primary production was also determined, by oxygen generation, on each day of the campaign. Our results showed that the productive potential of the lake was typically modest (campaign averages of 45-90mg C/m2.h) but that many of the regulating factors were not those anticipated intuitively. The lake is demonstrably meromictic, reminiscent of karstic dolines in higher latitudes, its stratification being maintained by solute- concentration gradients. Light penetration is poor, attributable to the turbidity owing to fine calcite and other particulates in suspension. Net primary production in the mixolimnion of Laguna de Guavita is sensitive to day-to-day variations in solar irradiance at the surface. However, deficiencies in nutrient availability, especially nitrogen, also constrain the capacity of the lake to support a phytoplankton. We deduced that Laguna de Guatavita is something of a limnological enigma, atypical of the common anticipation of a “mountain lake”. While doubtlessly not unique, comparable descriptions of similar sites elsewhere are sufficiently rare to justify the presentation of the data from Laguna de Guatavita that our studies have revealed so far.
Key words: chlorophyll a, photosynthetic rate, stratification, meromictic structure, sacred lakes.
Resumen
Los factores que regulan la producción primaria en un lago remoto, pequeño, ecuatorial y de elevada altitud en la región Oriental de los Andes Colombianos (4°58’50” N - 73°46’43” W, altura 2 935m.s.n.m., área 0.11km2 y profundidad máxima 30m), que también tuvo importancia cultural para los indígenas precolombinos y los orígenes de la leyenda de El Dorado, fueron investigados. La relativa lejanía del lugar requirió una serie de tres campañas intensivas de estudio, en un periodo de un año (2003-2004). Durante cada campaña se realizaron perfiles temperatura, concentración de oxígeno e intensidad de luz. Se recolectaron muestras para medir la concentración de clorofila y de solutos de significado biológico en agua filtrada GF/F. Cada día de la campaña se llevaron a cabo mediciones de la producción primaria, como se indica por la producción de oxígeno. Estas demostraron que el potencial productivo es particularmente modesto (promedio de campañas de 45-90mgC/m2.h, pero casi todos los aspectos de su regulación fueron sorprendentes y contrarios al sentido común. El lago es meromíctico, reminiscencia de dolinas kársticas de latitudes más altas y la estratificación se mantiene por los solutos. La penetración de luz es pobre, atribuible a la alta turbidez debida a la calcita fina y a otras partículas en suspensión. La producción primaria neta del mixolimnion de la Laguna de Guatavita es sensible a las variaciones del día a día de la radiación superficial. Deficiencias en la disponibilidad de nutrientes, sobre todo, del nitrógeno, también limitaron la capacidad productiva del fitoplancton. La laguna de Guatavita es una especie de enigma limnológico: aunque sin duda, no es el único, las descripciones de sitios similares en otras partes son lo suficientemente raras para justificar la presentación de los datos revelados hasta hoy en nuestro estudio.
Palabras clave: clorofila a, tasa fotosintética, estratificación, estructura meromíctica, lagos sagrados.
High-altitude mountain lakes remain relatively under-researched, mostly because of their remoteness and relative inaccessibility render them logistically difficult locations for sustained research programmes. The popular assumption that they are usually highly transparent, oligotrophic and nutrient-poor has been scarcely challenged. When mountain lakes are simultaneously located at low latitudes, small annual fluctuations in day length and potentially narrow ranges of annual temperature variation seem likely to generate only modest seasonal effects on phytoplankton production, at least when compared to the cyclic, seasonal variability detected among temperate lakes (Lewis 1996). On the other hand, as at all other latitudes, short-term variability in insolation, in hydrological exchanges, in wind induced vertical mixing and in their various impacts on the underwater availability of light and nutrients, in the intensity of planktivores activity, is as potentially pervasive in influencing phytoplankton dynamics and periodicity (Talling 1969, Lewis 1996, Hubble & Harper 2001). Nevertheless, lakes situated in mountainous regions may well be subject to fluctuating local microclimates and weather conditions, introducing extremes of cloud and precipitation, including snow, that leave habitats as vulnerable to short-term variability in irradiance, nocturnal cooling, mixed depth and light availability, as they do in the trophogenic layers of lakes at higher latitudes.
Whereas the photosynthetic responses of phytoplankton to short-term variations in light intensity are now well documented (Marra 1978, Yoder & Bishop 1985, Kroon et al. 1992, Mallin & Paerl 1992), longer-term adaptive adjustments –in swimming behaviour, pigmentation, and growth rates– eventually affecting changes in species composition - are still being resolved (Reynolds 2006). The case for investigating phytoplankton assemblages, in the context of a wide range of well-characterized habitat conditions, remains strong. The desire to include information on the environmental conditions and dynamic responses of phytoplankton in high-altitude, low-latitude lakes requires no further justification. The choice of suitable locations in the high mountains of the Andean Cordillera and close to the equator seems, retrospectively, self-selecting: there are numerous small lakes, of various origin (orogenic, volcanic and glacial), located in the Cundinamarca and Boyacá departments of Colombia, and not excessively distant from the National University in Bogotá, that offer prima-facie sites for investigation of the influences of low latitude and high altitude on fundamental hydrobiological processes. Even in this particular region, there are inevitable logistic difficulties in organizing and sustaining relevant systematic field studies. In the end, the results of an earlier survey of one of these lakes (Zapata 2001, Rivera et al. 2005, Donato 2010) were decisive in selecting Laguna Guatavita as the site for the present study.
The first purpose of this paper is to convey the results of this investigation. It was necessarily undertaken in a series of intense 16-18 day field campaigns that were designed to obviate a large number of distant sampling expeditions. However, as we shall demonstrate, almost every aspect of the limnology of Laguna de Guatavita was substantially outside our expectations. Our findings showed that the environmental factors that substantially control the biological productivity of the lake were sufficiently counterintuitive to persuade us to share our observations with a wider audience of limnologists.
It is relevant to mention that there was another factor influencing our selection of Laguna de Guatavita as a study site and it has less to do with limnology and rather more to its significance in the long history of the pre-Columbian settlement of the central Andes by indigenous people. At least one tribe of Indians, the Muiscas, flourished here until the 16th Century. They belonged to the Chibcha linguistic family that settled mainly in the basins of the former Pleistocene lakes, between Bogotá and Tunja (Reichel-Dolmatoff 1997). Indeed, lakes figured importantly in Muiscan culture: life and death were regarded as transitions between terrestrial activity and aquatic dormancy. The various high-altitude lagoons were treated as sacred places, dedicated to the gods and where they were worshiped and were paid tributes in the form of figurines, fashioned from wood, cotton, stone and, especially, gold, called “tunjos”, and emeralds. The famous legend of El Dorado originated from the Muiscan practice of anointing new chiefs with turpentine and gold dust, from forehead to feet, so that they resembled a shaft of radiant light. The new chief became purified by bathing in the limpid waters of the Guatavita lagoon, from which activity, he drew the strength and wisdom necessary to govern his people with due justice (Perdomo 1989). It was the ablutions that became of interest to the post-Columban conquistadores.
Materials and methods
Laguna de Guatavita is one of a series of heterogeneous lakes located in the Eastern Range of the Colombian Andes, supposed to have been formed towards the end of the Pleistocene glaciation, 10 000-12 000 years before present (Psenner & Catalan 1994). The lakes vary in their morphometry, hydrology, chemical composition and stratification, as well as the nature and abundance of the phytoplankton they support (Donato 2010). Laguna de Guatavita (4°58’50” N - 73°46’43” W) is a small, roughly elliptical (437×325m) lake, believed to be a karstic collapse crater (Rivera et al. 2005). Its area (0.114km2) and current altitude of its surface (2 935m.a.s.l) are regulated by an artificial channel, which was cut in the year 1 580 as part of an attempt (largely successful) to recover Muiscan treasures from the lake and which dropped the water level by about 20m. The maximum depth of the present lake is 30m (mean depth: 13.1m, relative depth: 7.9%) and it has a capacity of 1.45×106m3. It has a small, wooded topographic catchment (area: 0.67km2). Rivera et al. (2005) also revealed the presence of a steep chemical gradient in the lake, separating the upper and lower water masses and confining photosynthetic production to the insolated part of the water column (Zapata 2001); carbon fixation rates in the order 24-165mgC/m2.d were consistent with those said to typify oligo productive lakes.
Our field studies were necessarily carried out in a series of three intensive campaigns, as the relative remoteness of Laguna de Guatavita from Bogotá made frequent, routine visits quite impractical. The campaigns, each lasting 16-18 days, were initiated during April 2003, August 2003 and April 2004. On each day of each campaign, net primary production (NPP) was estimated from the change in oxygen concentration in isolates of plankton exposed in light- and darkened- bottles, suspended in situ in the water column (basically the method of Gaarder & Gran (1927), as later modified by Vollenweider (1974). All oxygen concentration measurements were determined titrometrically by the standard Winkler method. Exposures commenced between 10.00-11.00 on each occasion and were run for 4-6 hours, i.e., until 14.00-16.00.
Vertical profiles of dissolved oxygen concentration, temperature, conductivity and light at the deepest point of the lake were also constructed each day. Temperature and dissolved oxygen were measured using a YSI Model 57 probe (Yellow Springs, Ohio, USA); a separate, YSI Model 33 probe was used to measure conductivity. Light intensity was routinely measured using a digital light meter (FB70427, Fisher Scientific, Leicestershire, UK) that integrates the intensity (areal flux density) of photosynthetically - active radiation (PAR, 400-700nm: Wetzel & Likens 2001). Profiles of light intensity were also measured every 15min throughout each experimental NPP determination. Mean irradiances were calculated from integrated PAR readings, using the Microcall Origin 3.5 program; from mean irradiance measurements at each depth (Iz) and supposing conformity to the typical exponential series Iz=Io e-kz (where I0 is the intensity of light immediately beneath the water surface); the coefficient of vertical light extinction kz=(ln Iz- ln I0)/z was solved on each occasion. The depth of the optical disappearance of a Secchi disk was also noted on each occasion.
Water samples were collected in the field for immediate measurement of nutrient and chlorophyll-a content. The concentrations of combined inorganic nitrogen (NH4+, NO3- , NO2-) were determined separately, following standard APHA (1998) methods. Soluble reactive phosphorus (SRP) was measured in GF/Ffiltered samples, using molybdenum-blue method (details also in APHA 1998).
Chlorophyll-a concentrations were determined in samples collected at selected depths (0.1, 2, 4 and 8m). Hot extracts in 90% acetone were cooled and read in a Hewlett-Packard model 8453E; the formula of Jeffrey & Humphrey (1975), in APHA 1998) was applied to determine the concentration of chlorophyll-a in the original sample. Biomass-specific rates of net photosynthetic production were calculated as the quotient of NPP measurements (assuming molecular equivalence of carbon fixed and oxygen generated, in mgC/m3.h) and the corresponding chlorophyll concentration (mgchla/m3).
The program Winsurfer 6.0 (Goleen Software 1995) was used to generate the graphical representations of vertical gradients in environmental factors and in NPP. Depth-specific physical, chemical and biological variables were first normalized (as ln(X+1), square-root, cosine or arcsine equivalents) before plotting. The Shapiro-Will Test of normality was applied. The coefficients of variation (CV) were calculated to determine the strength of daily and vertical differences. Pearson correlation coefficients were used to establish relationships among the variables. For the spatial and temporal variables that were not normalized by transformation, non-parametric Kruskal-Wallis tests were applied. Spearman rank correlations were applied to define relations among the variables, using the Statistic 2000 program.
Results
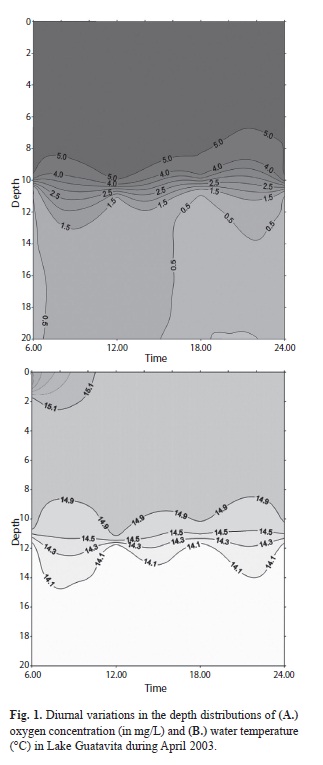
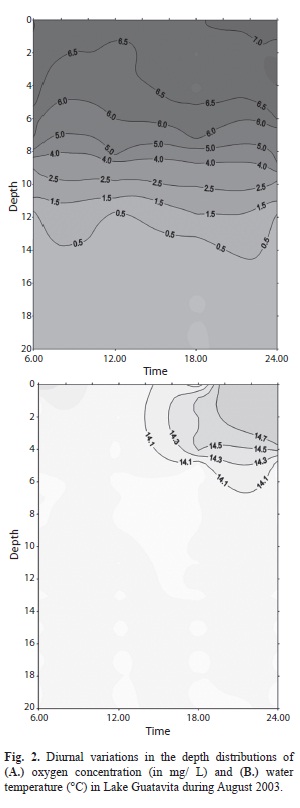
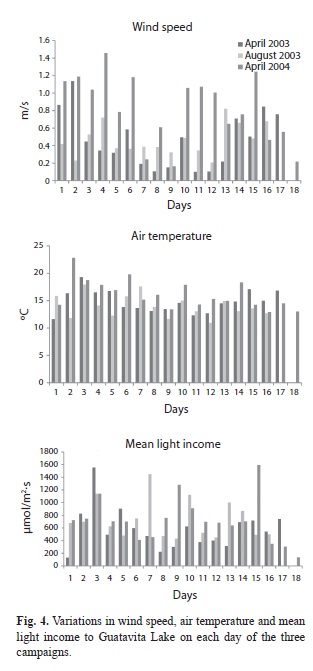
Stratification: The vertical distribution of oxygen isotherms and temperature through the period of each 16-18-day campaigns (April 2003, August 2003, April 2004) are shown in figures 1-3, respectively. It is clear that, to various extents, the distribution of oxygen isopleths (Figs. 1A, 2A, 3A) reveals an ongoing stratification lake stably stratified throughout the year, with the 2mg/L isopleth consistently located at a depth close to 10m below the surface. At the same time, the corresponding distribution of isotherms (Figs. 1B, 2B, 3B) is only sometimes well matched to the oxygen. Moreover, the temperature differences between the superficial layers and the deep water, perhaps as little as 0.5oC, do seem unlikely to be solely responsible for maintaining the evidently strong stratification. In figure 4, we summarize the daily information related to the average air temperature, light income and wind stress on the lake surface. We noted that in no case do the day-to-day variations in any of these components have any disruptive impact on the stability of the observed differentiation of the water column.
In contrast, the vertical gradient of specific electrical conductance was pronounced and recurrent throughout the sampling campaigns, separating the remarkably solute-dilute waters (~10μS/cm) of the upper part of the water column (≤11m depth) from deeper waters of ≥70μS/cm at depths of >20m. We deduce the observed density stratification in the Guatavita Lake and its apparent year-long persistence is a quasi-permanent feature. Without further evidence to the contrary, it is reasonable to regard the lake as being conventionally meromictic. We may thus justify reference henceforth to the upper mixolimnion and lower monimolimnion of Laguna de Guatavita.
Physico-chemical factors of biological relevance: Light income to the lake was found to be variable from day to day (Table 1). However, there is no evidence of significant seasonality in the light incomes recorded (fluctuations in duration and maximum intensity are not expected at this latitude!): the variations likely owe principally to day-to-day differences in the extent and duration of cloud cover. On the other hand, gradients of underwater light penetration have been found to be typically steep, with coefficients of extinction consistently falling between 1.0/m (in August 2003) and reaching values of 1.48/m in April, 2003 and 1.6/m in April, 2004 (Table 1). The implied levels of turbidity, largely attributable to suspended fine particles of clay and calcite, seem to be a persistent feature of the underwater light climate. Using conventional approximations (from Reynolds 2006), the depth of the euphotic zone (wherein net photosynthesis is supportable) is estimated rarely to have exceeded 3.5-4.5m; the Secchi disk measurements recorded during the three campaigns (Table 1) point to a similar extrapolation.
Investigations of nutrient availability failed to detect nitrate in any of the samples for all campaigns, while nitrite was found only once (April, 2004). Inorganic combined nitrogen was represented by ammonium concentrations between 3.17-5.55 μMN/L. Average values of SRP were measured in April up to a maximum of 0.95μMP/L in April 2003; in April 2004, values ≤0.54 μMP/L were observed. The modest concentrations of both nitrogen and phosphorus are not atypical of oligotrophic lakes; we deduce that availability of nitrogen is more likely than that of phosphorus to regulate phytoplankton production and the biomass supportive capacity of Laguna Guatavita.
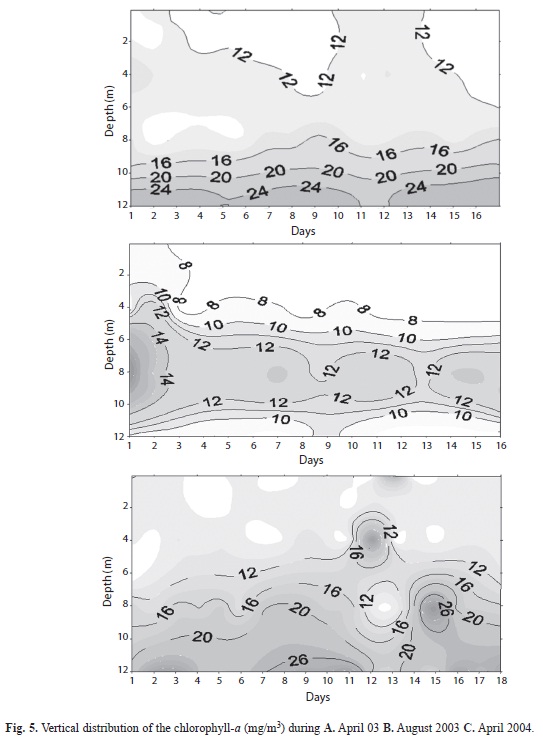
Distribution of chlorophyll and biomass:Isopleths of chlorophyll-a concentration in the vertical direction during each of the three campaigns are respectively illustrated in figures 5 A, B, C. Concentrations in the upper mixolimnion were generally lower than 12mg chla/m3 but, to a greater or lesser extent, were steeply graded to maxima of 20-28mg chla/m3 located in the pycnocline region. In each case, the main photosynthetic algae comprised mainly Cryptophytes, whose dominance of the phytoplankton of Laguna de Guatavita had been noted previously by Orjuela (2004). To the extent that the chlorophyll is healthy and functional, the algae are required to be substantially photoadapted to be capable of sustaining net photosynthesis at depths where residual light levels are usually less than <10mM/m.
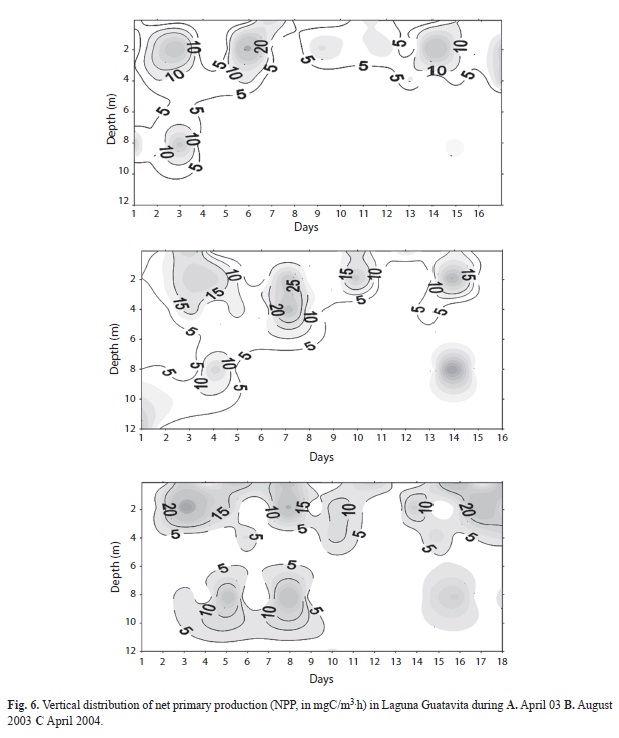
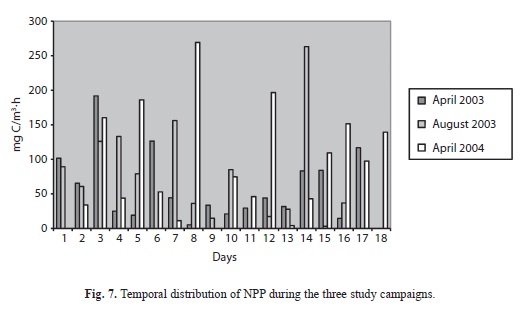
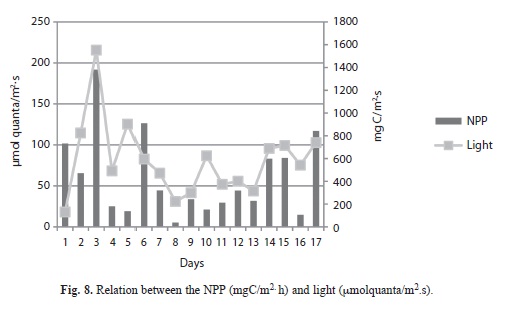
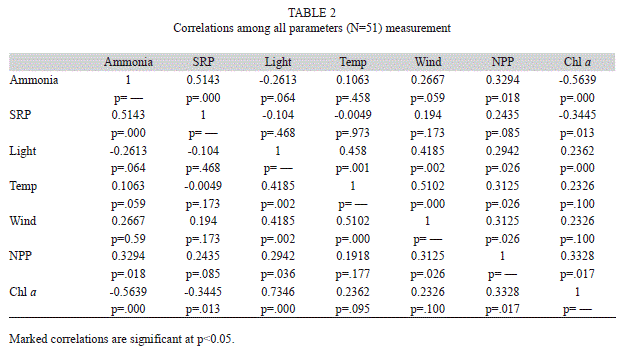
Primary production: Measured rates of net primary production (NPP) varied considerably from day-to-day (Figs. 6. A, B, C) but campaign-to-campaign differences in daily areal integrals (Fig. 7) were not significant (p=0.59, n=51). Solutions to the campaign averages expressed per unit of lake area ranged from 45.4-89.96mgC/m.h. However, daily variations in the areal integrals were evidently related to the surface irradiance, as represented by the data for the first campaign (Fig. 8). However, the matrix of correlations shown in table 2, relating NPP to variations in the physical (surface light income, wind speed, temperature), chemical (ammonium, soluble reactive phosphorus concentrations) and biological (chlorophyll concentrations), suggest that, in addition to the amount of chlorophyll present, temperature, windspeed and ammonia concentration are of greater significance (p<0.05) in influencing primary production in Laguna de Guatavita.
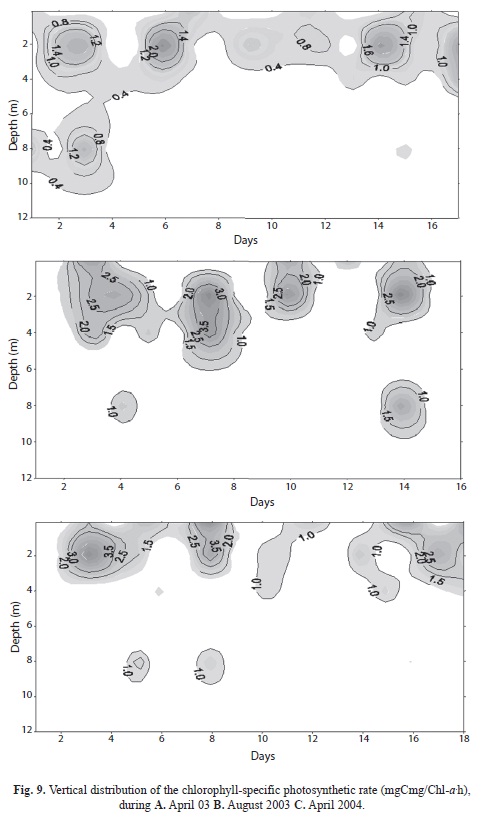
Day-to-day variations in dachlorophyll-specific photosynthetic rates in the mixolimnion of the lake were observed in all three campaigns (with maxima occasionally as high as 4.0mg Cmg/Chla.h, figures 9 A, B, C). Despite the presence of high concentrations of chlorophyll at depths >8m, the chlorophyll-specific rates of photosynthesis there were small. Discussion
The investigations presented here confirm several features of Laguna Guatavita that are not at all typical of those generally expected in high mountain lakes: low nutrient availability (especially of phosphorus), high clarity and supportive of only low plankton biomass, with low rates of recruitment by growth. Whereas the maximum chlorophyll-specific rates of photosynthesis observed, in the order of 3-4mgC mg/chla.h, are fairly typical (Table 3 of Reynolds 2006), their occurrence is constrained in vertical distribution and is evidently very sensitive to day-to-day fluctuations in surface irradiance. Plainly, slower rates of chlorophyll-specific production by rather greater concentrations of chlorophyll located at depths between 8-10m contribute to the higher absolute rates of photosynthetic production found.
These features of the primary production in Laguna Guatavita are directly related to the curious structural properties of the limnology. Despite the indications detected in previous field studies, the present confirmation of a stable, meromictic water column coupled with a severe restriction of light penetration through a turbid mixolimnion, simultaneously more deficient in nitrogen than phosphorus, reveals an unusual and strikingly unexpected environment. Of course, similar conditions have been described before: our findings are strongly reminiscent of those of the Laguna de la Cruz, a karstic doline, near Cuenca, in Spain (as described by Vicente & Miracle 1988). For instance, the solute-supported meromixis and the tendency to precipitate carbonate, even the dominance of cryptophytes in the stable part of the water column, and are similar to the features now recognized in Laguna de Guatavita. On the other hand, our knowledge of the extent of local carbonate deposits is insufficient to enable us to draw large-scale regional comparisons between the Andean and Spanish localities. Other locations of carbonate-bearing sedimentary deposits in the Andes are known in Northern Cordillera (Rios et al. 2008). Also known is the presence of small, stablystratified, likely meromictic, high-altitude lakes in Colombia, having distinctive metalimnetic layers dominated by purple sulphur bacteria (Nuñez-Cardona et al. 2008).
The relatively high turbidity is related to the precipitation of calcium carbonate. It is unclear how much of this material is dependent upon current rates of precipitation, as opposed to delayed sedimentation and frequent re-suspension. In either case, the truncation of downwelling light is nevertheless a persistent component regulating the environmental conditions experienced and accommodated by photosynthetic organisms in Laguna Guatavita. With relatively little variation in the daily light income being attributable to seasonal variation in day length, it is interesting to detect that the rates of community photosynthesis are so relatively sensitive to day-to-day variations in surface irradiance. Again, this may be an unusual feature among high-altitude lakes.
Deficiencies in nutrient supply are commonly encountered in high-altitude lakes having small, infertile hydrological catchments, and Laguna de Guatavita seems unexceptional in this context. The meromictic stratification and the substantially hypoxic condition of the monimolimnion surely benefit ammonification, leading to relatively high values of ammonia and phosphates in deep water. The N:P ratios in the upper waters nevertheless point to a likely, ongoing limiting role of nitrogen on the productive and biomass-supportive capacities in the lake. Although this might not be considered general among high-altitude lakes, it does agree with the findings of many other Andean lakes of Peru-Bolivia (Vincent et al. 1984), Colombia (Donato et al. 1996, Donato 2010) and Argentina (Carignan et al. 1998, Diaz et al. 2007).
The infrequency of disturbance of the stratification in meromictic lakes often allows the prolongation of limnetic conditions allowing the maturation of processes selecting for localized, low-diversity dominant assemblages of phytoplankton. Culmination of such series in, variously, filamentous, buoyancy-regulating Cyanobacteia or even by gas-vacuolate purple and green sulphur bacteria (Chromotiaceae, Chlorobiaceae), is recognized at all latitudes where the opportunities arise (Reynolds 1992). Alternatively (and possibly where the physical stratification is less stable), Cryptophyte species may fulfill this role, where they are able to adjust vertical position more readily than buoyancy-regulating organisms and to balance changes in light reception through photo-adaptive adjustment.
To conclude, we recognize that primary production in this oligotrophic high-altitude tropical lake, is regulated first by its meromictic structure. The stability is determined chemically, as a consequence of its situation in karstic formations and not primarily as a result of the regional climate. However, day-to-day variability in net primary production in the mixolimnion of Laguna de Guatavita is consequential on local factors affecting light income and availability. Deficiencies in nutrient availability, especially, of nitrogen, also constrain the phytoplankton supportive capacity. Despite the significance of the lake in the traditions of the Muiscan people, we have no evidence that the supply of gold or other precious minerals plays any significant part in the ecology of Laguna de Guatavita!
Acknowledgments
The authors thank Colciencias, Corporación Autónoma Regional de los Valles de Ubaté y Cundinamarca, CAR, Pontificia Universidad Javeriana and Universidad Nacional de Colombia for their support given during the time of this study.
References
American Public Health Association (APHA) 1998. Standard methods for the examination of water and wastewater. American Public Health Association, Washington D.C., USA. [ Links ]
Carignan, R., A.M. Blais & C. Vis. 1998. Measurement of primary production and community respiration in oligotrophic lakes using the Winkler method. Can. J. Fish. Aquat. Sci. 55: 1078-1084. [ Links ]
Diaz, M., F. Pedrozo, C. Reynolds & P. Temporetti. 2007. Chemical composition and the nitrogen-regulated trophic state of Patagonian lakes. Limnologica 37: 17-24. [ Links ]
Donato, J., L. González & C. Rodríguez. 1996. Ecología de dos sistemas acuáticos de páramo. Academia Colombiana de Ciencias Exactas Físicas y Naturales, Bogotá, Colombia. [ Links ]
Donato, J. 2010. Phytoplankton of Andean Lakes in Northern South America (Colombia). Diatom Monograph, Koenigstein, Germany. [ Links ]
Gaarder, T. & H.H. Gran. 1927. Investigations of the production of plankton in the Oslo Fjord. Rapp. Cons. Explor. Mer. 40: 1-48. [ Links ]
Hubble, D. & D. Harper. 2001. Impact of light regimen and self shading by algal cells on primary productivity in the water column of a shallow tropical lake (Lake Naivasha. Kenya). Lakes Reserv. Manage. 6: 2-143. [ Links ]
Jeffrey, S.W. & G.F. Humphrey. 1975. New spectrophotometric equations for determining chlorophylls a, b, c1 and c2 in higher plants, algae and natural phytoplankton. Biochem. Physiol. Pfl. 167: 191-194. [ Links ]
Kroon, B.M.A., M. Latasa, B.W. Ibelings & L.R. Mur. 1992. The effect of dynamic light regimes on Chlorella. I. Pigments and cross section. Hydrobiologia 238: 71-78. [ Links ]
Lewis, W.M. 1996. Tropical lakes: how latitude makes a difference, p. 43-64. In F. Schiemer & K.T. Boland (eds.). Perspectives in Tropical Limnology. SPB, The Hague, Holland. [ Links ]
Mallin, M.A. & H.W. Paerl. 1992. Effects of variable irradiance on phytoplankton productivity in Shallow estuaries. Limnol. Oceanogr. 37: 54-62. [ Links ]
Marra, J. 1978. Phytoplankton photosynthetic response to vertical movement in mixed layer. Mar. Biol. 46: 203-208. [ Links ]
Nuñez-Cardona, M.T., J. Donato-Rondon, C.S. Reynolds & J. Mas. 2008. A purple sulfur bacterium from a high-altitude lake in the Colombian Andes. J. Biol. Res. Thessalon. 9: 17-24. [ Links ]
Orjuela, R. 2004. Relaciones entre diversidad y producción primaria fitoplanctónica en el lago de Guatavita. Tesis Pregrado Ecología, Pontificia Universidad Javeriana, Bogotá, Colombia. [ Links ]
Perdomo, L. 1989. Manual de Arqueología colombiana. Carlos Valencia, Bogotá, Colombia. [ Links ]
Psenner, R. & J. Catalan. 1994. Chemical composition of lakes in crystalline basins: a combination of atmospheric deposition geologic backgrounds, biological activity and human action, p. 255-314. In R. Margalef (ed.). Limnology now: a paradigm of planetary problems. Elsevier, Amsterdam, Holland. [ Links ]
Reichel-Dolmatoff, G. 1997. Arqueología de Colombia. Imprenta Nacional de Colombia, Bogotá, Colombia. [ Links ]
Reynolds, C.S. 1992. Dynamics, selection and composition of phytoplankton in relation to vertical structure in lakes. Arch. Hydrobiol. Beih. Ergebn. Limnol. 35: 13-31. [ Links ]
Reynolds, C.S. 2006. Ecology of phytoplankton. Cambridge, Cambridge, England. [ Links ]
Ríos, C.A., O.M. Castellanos, S.I. Gómez & G.A. Ávila. 2008. Petrogenesis of the metacarbonate and related rocks of the Silgará formation, Central Santander Massif, Colombian Andes: An overview of a “reaction calcic exoscarn”. Earth Sci. Res. J. 12: 72-106. [ Links ]
Rivera, C., D. Solano, A. Zapata & J. Donato. 2005. Phytoplankton diversity in a tropical high mountain lake. Verh. Int. Ver. Theor. Angew. Limnol. 29: 418-421. [ Links ]
Talling, J.F. 1969. The incidence of vertical mixing, and some biological and chemical consequences in tropical Afircan lakes. Verh. Int. Ver. Theor. Angew. Limnol. 17: 998-1012. [ Links ]
Vicente, E. & M.R. Miracle. 1988. Physicochemical and microbial stratification in a mermictic karstic lake of Spain. Verh. Int. Ver. Theor. Angew. Limnol. 23: 522-529. [ Links ]
Vincent, W., W. Wurtsbaugh, C. Vincent & P. Richerson. 1984. Seasonal dynamics of nutrient limitation in a tropical high-altitude lake (Lake Titicaca, Peru-Bolivia): Application of physiological bioassays. Limnol. Oceanogr. 29: 540-552. [ Links ]
Vollenweider, R.A. 1974. A manual of methods for measuring primary production in aquatic environments. Blackwell, London, England. [ Links ]
Wetzel, R. 2001. Limnology. Lake and river ecosystems. Academic, San Diego, California, USA. [ Links ]
Wetzel, R & G.E. Likens. 2000. Limnological Analyses. Springer-Verlag, New York, USA. [ Links ]
Yoder, J.A. & S.S. Bishop. 1985. Effects of mixing - induced irradiance fluctuations on photosynthesis of natural assemblages of coastal phytoplankton. Mar. Biol. 90: 87-93. [ Links ]
Zapata, A. 2001. Variaciones diarias y mensuales de la productividad primaria en un lago andino (Laguna de Guatavita. Cundinamarca). Tesis Maestría, Pontificia Universidad Javeriana, Bogotá, Colombia. [ Links ]
*Correspondencia:
Jhon Donato: Departamento de Biología, Universidad Nacional de Colombia, Av (Cra.) 30 No. 45-03, Bogotá, Colombia. jcdonator@unal.edu.co.
Paola Jimenez: Departamento de Biología, Universidad Nacional de Colombia, Av (Cra.) 30 No. 45-03, Bogotá, Colombia. paolajimenezmedina@yahoo.com.ar
Colin Reynolds: CEH Lancaster, Library Avenue, Bailrigg, GB-LA1 4AP Lancaster UK. csr@ceh.ac.uk
1. Departamento de Biología, Universidad Nacional de Colombia, Av (Cra.) 30 No. 45-03, Bogotá, Colombia; jcdonator@unal.edu.co
2. Departamento de Biología, Universidad Nacional de Colombia, Av (Cra.) 30 No. 45-03, Bogotá, Colombia; paolajimenezmedina@yahoo.com.ar
3. CEH Lancaster, Library Avenue, Bailrigg, GB-LA1 4AP Lancaster UK; csr@ceh.ac.uk
Received 29-VII-2011. Corrected 30-I-2012. Accepted 29-II-2012.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}