










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.60 n.2 San José Jun. 2012
Ecología trófica en larvas de Rhinella arenarum (Anura: Bufonidae) en agroecosistemas y sus posibles implicaciones para la conservación
Clarisa Bionda1*,4*, Noemi Gari2*, Elisa Luque2, Nancy Salas1, Rafael Lajmanovich3* & Adolfo Martino1
*Dirección para correspondencia:
Abstract
The progress of the agriculture border has led an important loss of natural habitats, with significant consequences for biodiversity. In this sense, the studies in anuran amphibian tadpoles inhabiting these environments are relevant, because the larval stage is a phase of population regulation. The aim of this study was to analyze the body condition and diet in Rhinella arenarum, tadpoles, an anuran species widely distributed in South America and that inhabit agroecosystems. Three sites were sampled, two agroecosystems with different alteration degrees (C1 and C2) and an uncultured (SM) third place. The captured tadpoles were anesthetized, fixed and preserved in formaldehyde (10%). Subsequently, body measurements were made and the complete intestine was removed and analyzed for food items under a binocular microscope. The diet in R. arenarum tadpoles has a dominance of algae Bacillariophyceae, followed by Cyanophyceae. In particular, the class Bacillariophyceae, due to the presence of the genus Navicula, Nitzschia, Gomphonema and Hantzschia, was important in the diet of the anurans in those agroecosystems. Class Cyanophyceae, mainly represented by genus Oscillatoria and Euglenophyceae represented by Euglena and Strombomonas, were predominant in the diet of the anurans in SM. Some differences in the total items consumed by tadpole were observed between the studied sites. Tadpoles that inhabit the modified sites (C1 and C2) recorded a significantly smaller amount of food. Moreover, the tadpoles that inhabit these sites showed a lower body condition. The presence of certain algae associated with eutrophic environments, could indicate some pollution in agroecosystems (C1 and C2). Food resources would be lesser in places with strong agricultural activity, possibly with a greater degree of eutrophication. A smaller food amount could have consequences at population level for the short and long time terms, because of its impact on individual growth. Larval diet is suggested as a potential bioindicator of environmental health for these areas. Rev. Biol. Trop. 60 (2): 771-779. Epub 2012 June 01.
Key words: larval diet, food availability, body condition, agroecosystems, Rhinella arenarum.
Resumen
El crecimiento de la agricultura produce perdida del habitat natural, con consecuencias para la biodiversidad de los anfibios. Se analizo la dieta y condición corporal de larvas de anuros de Rhinella arenarum que habitan agroecosistemas. Los muestreos fueron realizados en dos agroecosistemas y en un tercer sitio, no afectado por cultivos. Las larvas capturadas fueron anestesiadas, fijadas y preservadas en formaldehido, se realizaron medidas corporales y se analizo la dieta de las larvas. Se registro diversidad en la oferta alimenticia proporcionada por los distintos ambientes. La dieta tuvo una predominancia de algas Bacillarophyceae, seguidas por Cyanophyceae. Particularmente, los generos Navicula, Nitzschia, Hantzschia y Gomphonema (clase Bacillarophyceae), fueron importantes en los agroecosistemas. Los generos Osillatoria, Euglena y Strombomonas (clases Cyanophyceae y Euglenophyceae), predominaron en la dieta de las larvas de anuros en el sitio menos alterado. Las larvas de los sitios mas alterados consumen menor cantidad de alimento y registran una menor condicion corporal. La presencia de determinadas algas, indicarían un mayor grado de contaminación en los agroecosistemas. La eutrofizacion de lagunas podría alterar la disponibilidad de alimento para larvas de anuros que podría tener consecuencias poblacionales negativas. Se sugiere el análisis de dietas larvarias como un potencial bioindicador de salubridad ambiental.
Palabras clave: dieta larval, disponibilidad de alimento, condición corporal, agroecosistemas, Rhinella arenarum.
La proporción de bosques transformados en tierras agrícolas esta ocurriendo rápidamente en algunas areas Neotropicales. Particularmente, la región central de Argentina ha sido afectada por el desarrollo agricola (Rossi2006), con una clara tendencia de reemplazo de la agricultura tradicional por una producción mas especializada, principalmente en la obtención de soya (Glycine max L.). Este progreso de la agricultura ha convertido a la región en un importante productor, procesador y exportador de granos, aceite y harina de soya (Rossi 2006). Sin embargo, la agricultura se ha convertido en la principal causa de reducción, fragmentación y deterioro del habitat natural, y en consecuencia, es considerada un factor relevante en la declinación registrada en los anfibios en las ultimas décadas (di Tada et al. 1996, Wake 1998, Davidson et al. 2002, GAA 2004, Stuart et al. 2004, Beebee & Griffiths 2005, Collins & Crump 2009).
En este contexto, resulta valioso el conocimiento sobre la ecologia de larvas de anfibios, ya que los estadios larvales presentan un papel fundamental en la regulación de la fase adulta (Heyer 1974, Wilbur 1980). Las alteraciones de los ambientes acuáticos o el deterioro en la calidad de los mismos, puede influir en las larvas de anfibios tanto en el tiempo y tamaño alcanzado en la metamorfosis, como en su desarrollo normal, aumentando la mortalidad o la depredación (Carey & Bryant 1995, Taylor et al. 2005, Alitg et al. 2007). Sin embargo, los trabajos desarrollados acerca de la alimentación en anfibios que habitan los agroecosistemas, han sido realizados principalmente en estadios terrestres (Attademo et al. 2005,2007) y poco se conoce sobre la alimentación en larvas. Algunos autores (Heyer 1974, Lajmanovich 2000, Rossa Feres et al. 2004) han demostrado que las larvas utilizan como recurso todo aquello que se encuentra disponible en su ambiente, ya que no necesariamente la alimentación constituye un factor para la segregación de las comunidades larvales. Asimismo, la alteración del habitat, puede modificar la disponibilidad de alimento en un ambiente dado, lo cual puede afectar el crecimiento y el desarrollo de los organismos que se sustentan en dichos ambientes. De este modo, los estudios sobre la dieta en larvas de anfibios pueden representar una medida general de la oferta y disponibilidad trófica de los distintos habitats y resultar una herramienta para la caracterización y monitoreo como bioindicador de la calidad ambiental (Jüttner et al. 1996, Blanco et al. 2004, Díaz Quirós & Rivera Rondón 2004).
En este trabajo nos proponemos explorar los posibles efectos de las modificaciones derivadas del establecimiento de agroecosistemas sobre la dieta y la condicion corporal de larvas de Rhinella arenarum Hensel 1867, una especie de anuro Neotropical con amplia distribución en Sudamerica.
Materiales y métodos
Los sitios de muestreo se encuentran en los alrededores de la localidad de Río Cuarto (33°07’55.18’’ S - 64°21’08.96’’ W), provincia de Córdoba, Argentina. La región se caracteriza por tener clima semiseco, con tendencia a semihumedo de las planicies. Las lluvias máximas se encuentran durante el periodo octubre-marzo, con una media anual de 800mm (Capitanelli 1979). La fisiografia del area de estudio corresponde a llanuras suavemente onduladas. La escasa pendiente favorece el estancamiento de las aguas, sumado al aporte subterráneo que junto con las precipitaciones, ayuda a mantener cursos de agua permanentes (Bridarolli & di Tada 1996).
La recolección de los especímenes fue realizada en tres sitios: dos de ellos se encuentran en zonas de cultivos de soya: Sitio Cultivo 1(C1, 33°05’54.38’’ S - 64°26’04.76’’ W) y Sitio Cultivo 2 (C2, 33°05’39.29’’ S - 64°25’57.72’’W). Ambos sitios presentan distintos grados de alteración en relación a la distancia de los cultivos a los cuerpos de agua. En C1, los cultivos estuvieron a escasos metros de los cuerpos de agua, mientras que en C2 se encontraban más alejados (200m aproximadamente). El tercer sitio de muestreo, llamado Sitio Semi-modificado (SM, 33°06’41.95’’ S - 64°18’14.71’’W), correspondió a una zona periurbana no afectada por cultivos, cubierta por pastizales y formaciones boscosas de arboles autoctonos e introducidos (Doffo 1989).
Las larvas de R. arenarum presentan habitos bentonicos, es decir, viven en el fondo de charcas y lagunas (Cei 1980, Lajmanovich 2000). La captura se realizo en diciembre de 2007, a lo largo de una transecta desde el margen del cuerpo de agua hasta 2m dentro del mismo. Luego de identificadas, las larvas fueron anestesiadas (MS 222), fijadas y preservadas en formaldehido al 10%. Se midió la longitud desde el rostro a la cloaca (LRC) con un calibre manual Vernier Somet Inox Extra (0.01mm precisión) y se obtuvo la masa con una balanza Mettler P11N 0-1000g. A partir de la LRC y la masa de las larvas, fue calculada la condición corporal (CC) de cada individuo. Para el calculo de la CC, fueron utilizados índices residuales de acuerdo a lo sugerido por Jakob et al. (1996) y Luddecke (2002).
Para un total de 10-12 larvas por sitio, entre los estadios 36-39 de Gosner (1960), se extrajeron los intestinos completos y el contenido del primer tercio fue analizado (Echeverria et al. 2007). Los recuentos de las algas se realizaron bajo microscopio optico Zeiss a 400X, utilizando camara porta y cubreobjeto de 24 x 50mm, se uso un volumen de 0.3mL de muestra para cada camara; se recorrieron tres transectas, una central y dos equidistantes a la central, cada una con una longitud igual a la del cubreobjeto y una altura correspondiente al diámetro del campo visual. Se considero como unidad de recuento al talo (unicelular, colonia, cenobio). Para la cuantificación de los filamentos, el equivalente a una longitud de 10μm fue considerado como una célula única (Hill et al. 2000). Para el calculo de la abundancia de organismos se utilizaron las ecuaciones según Villafane & Reid (1995). Con estos datos se obtuvieron los porcentajes de frecuencia de ocurrencia (FO%, porcentaje de individuos que consumieron determinada categoría de presa), se estimaron los totales y las frecuencias numéricas (N%) de los distintos items alimentarios cuantificables. Para describir la diversidad en la dieta entre las distintas poblaciones, se calculo el indice de diversidad (H) con la formula Shannon & Weaver (1949). Ademas, se midio la equitatividad (E) en la dieta para cada población usando la ecuacion de Magurran (1987). Para contrastar la diversidad media en la dieta de las poblaciones con distintos tamanos muestrales tambien se utilizo el metodo de Rarefaccion (Magurran 1987).
Los resultados para las medidas corporales y la cantidad promedio de items consumidos por larva fueron comparados estadísticamente entre sitios utilizando un analisis de la varianza (ANOVA) de una via y la prueba a posteriori de comparaciones multiples de Tukey. Los cálculos de rarefaccion fueron realizados con el programa EstimateS version 8.2.0 (Colwell 1997).
Resultados
En el cuadro 1 se muestran los resultados correspondientes a las medidas corporales realizadas en larvas de R. arenarum. Las larvas del sitio C1 registraron la menor LRC, masa y CC (Cuadro 1). La comparacion estadística de las medidas corporales, indico que la LRC y la masa de las larvas se diferencian significativamente entre los sitios (Tukey, p<0.05), con excepcion de LRC entre C1 y SM (Tukey, p>0.05) (Cuadro 1). Por otra parte, la CC presento diferencias significativas entre el C1 y el resto de los sitios (Tukey, p<0.05).
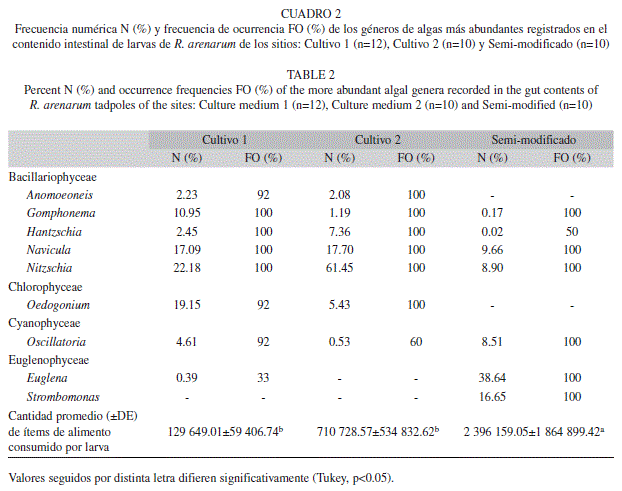
La dieta de las larvas de R. arenarum consistio principalmente en microalgas, particularmente diatomeas. En el cuadro 2 se muestran las clases y generos de algas mas abundantes registrados en la dieta de R. arenarum. En los sitios C1 y C2, las algas mas consumidas fueron las de la clase Bacillarophyceae, representada por 29 y 18 generos, y la clase Chlorophyceae con ocho y seis generos, respectivamente. En el sitio SM, además de la clase Bacillarophyceae, con 13 generos, otras como Cyanophyceae, con dos generos y Euglenophyceae, con cinco generos, fueron componentes principales en la dieta (Cuadro 2). En general, los generos Navicula y Nitzschia fueron predominantes en todos los sitios estudiados (Cuadro 2). Por otra parte, Gomphonema sp. y Hantzschia sp. fueron dominantes solamente en la dieta de las larvas de los ecosistemas agricolas. Mientras que en el SM, los generos mas consumidos fueron: Oscillatoria, Euglena y Strombomonas (Cuadro 2).
La cantidad de alimento consumido por las larvas del sitio SM difiere significativamente del resto de los sitios (Tukey, p<0.05) (Cuadro 2). Las larvas de los ecosistemas agrícolas consumen menor cantidad de alimento. La abundancia total registrada para los sitios C1 y C2 representa tan solo el 5% y 29% respectivamente, del total consumido por las larvas del SM (Cuadro 2). Sin embargo, la dieta en el C1 es mas diversa (C1: H=2.39; C2: H=1.55 y SM: H=2.00) y mas equitativa (C1: E=0.64; C2: E=0.43 y SM: E=0.63). La diversidad media en las distintas dietas calculadas según el método de rarefacción (C1=2.30±0.08; C2=1.53±0.03 y SM=2±0.12) también indica que la dieta del C1 es la mas diversa.
Discusión
Numerosos estudios utilizan a las algas diatomeas como bioindicadoras en diversos tipos de ambientes (Round et al. 1993, Jüttner et al. 1996, Blanco et al. 2004, Díaz Quiros & Rivera Rondón 2004). La dominancia en los cuerpos de agua de determinadas diatomeas, han sido asociadas a la contaminación por nitrógeno o fosforo (Blanco et al. 2004), componentes principales de los agroquímicos utilizados en los cultivos. En general, para todos los sitios estudiados, pero particularmente en los sitios agrícolas, las algas mas consumidas fueron Navicula y Nitzschia. Estos mismos géneros son considerados indicadores de las condiciones ambientales en distintos cuerpos de agua (Díaz Quiros & Rivera Rondón 2004) y son particularmente resistentes a ambientes de conductividad alta y posible contaminación orgánica (Ramírez & Plata Díaz 2008, Yucra & Tapia 2008). La abundancia significativa registrada de Nitzschia en el C2, podría estar asociada a niveles altos de salinidad para este ambiente (Nather Khan 1990). Por otra parte, Gomphonema y Hantzschia, presentes en ambos sitios de cultivos, son tipos de algas asociadas a ambientes con contaminación, pero particularmente Gomphonema, es encontrada en ambientes con niveles elevados de fosforo (Nather Khan 1990, Díaz Quiros & Rivera Rondón 2004, Kelly et al. 2005). En el sitio SM, los géneros que predominaron fueron: Oscillatoria, Euglena y Strombomonas. La predominancia de los géneros Oscillatoria y Euglena en la dieta de las larvas del SM, podría estar asociado a la presencia de mayor cantidad de materia orgánica en la laguna de este sitio (Fabrizi 2010). Si bien no se registran cultivos en los alrededores del sitio SM y por tal motivo resulta un ambiente de referencia para ser comparado con los agroecosistemas, este ambiente presenta cierto grado de alteración ya que se encuentra rodeado por una matriz periurbana. La presencia de residencias aledañas a este ambiente podría ser la razón de un incremento en la cantidad de materia orgánica y por ello la predominancia de estos géneros. De este modo, la presencia de ciertas algas registradas en la dieta de las larvas procedentes de distintos sitios podría estar corroborando sus niveles de alteración, lo que indica además, el estado de los sitios agrícolas estudiados.
La agricultura es uno de los factores principales de eutrofizacion de las aguas superficiales (USEPA 1994, Ongley 1997, Perdomo et al. 2001) y se conoce que los ambientes eutrofizados presentan menor calidad de alimento (Smol 2002). Las larvas de anuros normalmente muestran una alimentación poco especializada, raspando sobre superficies de plantas, por lo que sus dietas son indicadoras del tipo y abundancia del recurso alimenticio en sus ambientes (Heyer 1974, Lajmanovich 2000, Rossa Feres et al. 2004). En este sentido, los resultados obtenidos, sugieren que existe diversidad en la oferta alimenticia proporcionada por los distintos ambientes estudiados. Asimismo, podemos mencionar que de acuerdo a nuestros resultados la disponibilidad de alimento, en términos de cantidad, seria menor en aquellos sitios que registran un mayor grado de alteración por la actividad agrícola. La disponibilidad de alimento puede tener consecuencias poblacionales a corto y largo plazo. Una cantidad y calidad de alimento por debajo del valor optimo, puede influir de manera significativa en el tiempo de metamorfosis y en el tamaño de las larvas (Carey & Bryant 1995, Altig et al. 2007), afectando de este modo las principales interacciones biológicas, como la competencia y la depredación (Kupferberg 1997). En este sentido, nuestros resultados han mostrado que las larvas de R. arenarum procedentes de ambientes mas alterados, presentan una condición corporal mas baja. La condición corporal es un indicador de la calidad o estado fisiológico de un organismo (Bagenal & Tesch 1978, Jakob et al. 1996), lo cual puede estar directamente relacionado a su alimentación. La dieta analizada para las larvas de estos sitios indico que las mismas consumen una cantidad mas reducida de alimento. Esto podría ser considerado la causa por la cual dichas larvas registran una condición corporal mas baja. Estos posibles efectos de los ambientes modificados sobre la disponibilidad de alimento y condición corporal de las larvas, pueden comprometer la persistencia de la población, debido a que el tamaño y el estado fisiológico alcanzado por los individuos, influye en su supervivencia, reproducción y reclutamiento (Wilbur 1980, Semlitsch et al. 1988, Gray & Smith 2005). Un estudio previo realizado con estas mismas poblaciones ha indicado que la condición corporal alcanzada por las hembras influye de manera significativa en su output reproductivo (Bionda et al. 2011). Es necesario de este modo un mayor entendimiento de aquellos factores que afectan las poblaciones y comunidades “ensambles” de anuros, ya que además se ha demostrado que la eutrofización de los cuerpos de agua por escorrentias desde los agroecosistemas, puede llevar a un incremento en la mortalidad, a una variación en el comportamiento y a una alteración de las características de las historias de vida de las larvas de anfibios (Carey & Bryant 1995, Boone & Semlicht 2001, Carr et al. 2003, Coady et al. 2005, Taylor et al. 2005, Peltzer et al. 2008). Este tipo de eventos, puede distorsionar de forma significativa los procesos de regulación natural de poblaciones y comunidades lo que puede dar lugar a una disminución local en el tiempo (Boone et al. 2004).
Sin embargo, teniendo en cuenta que la dieta registrada en uno de los cultivos, fue la de mayor diversidad y equitatividad, y que las larvas consumen escaso alimento, cabria preguntarse si puede existir algún motivo por el que las larvas se alimentan poco, independientemente de la disponibilidad trófica. Existe escasa documentación sobre el tema, pero el retardo en el crecimiento de algunas larvas de anuros en presencia de contaminantes, ha llevado a preguntarse si estos pueden intervenir en la adquisición del alimento (Carey & Bryant 1995). Algunos estudios experimentales han demostrado que los fertilizantes nitrogenados, reducen la actividad de alimentación en larvas de algunas especies (Marco et al. 1999, Hatch & Blaustein 2000). Sin embargo, son necesarios más estudios para esclarecer este interrogante.
Este trabajo representa el primer aporte al conocimiento de la dieta de larvas de anuros en agroecosistemas del centro de Argentina. Asimismo, proponemos una nueva herramienta para el monitoreo de ambientes alterados, mediante el análisis de las dieta de estos organismos y los posibles efectos en las poblacionales en función de la disponibilidad de alimento. Por otra parte, la especie de anuro utilizada en este estudio presenta una amplia distribución en Sudamerica, seria interesante realizar estos estudios en otras regiones, para reconocer si el patrón de dieta registrado, es coincidente con otras poblaciones y/o ambientes.
Agradecimientos
Este trabajo fue financiado por el Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET) y SCyT UNRC, PIPCONICET N° 112-201001-00009. Los autores desean agradecer a dos revisores anónimos, cuyos comentarios permitieron mejorar este trabajo. Nuestro estudio fue autorizado por la Agencia Córdoba Ambiente (A.C.A.S.E.).
Referencias
Altig, R., M.R. Whiles & C.L. Taylor. 2007. What do tadpoles really eat? Assessing the trophic status of an understudied and imperiled group of consumers in freshwater habitats. Freshwater Biol. 52: 386-395. [ Links ]
Attademo, A.M., P. Peltzer & R. Lajmanovich. 2005. Amphibians occurring in soybean and implications for biological control in Argentina. Agr. Ecosyst.
Environ. 106: 389-394. [ Links ]
Attademo, A.M., P. Peltzer & R. Lajmanovich. 2007. Feeding habits of Physalaemus biligonigerus (Anura, Leptodactylidae) from soybean field of Cordoba province, Argentina. Russ. J. Herpetol. 14: 1-6. [ Links ]
Bagenal, T.B. & F.W. Tesch. 1978. Methods for assessment of a fish production in fresh waters, p. 101-136. In T.B. Bagenal (ed.). Age and growth. Blackwell, Oxford, Massachusetts, EEUU. [ Links ]
Beebee, T.J.C. & R.A. Griffiths. 2005. The amphibian decline crisis: A watershed for conservation biology? Biol. Conserv. 125: 271-285. [ Links ]
Bionda, C.L., R.C. Lajmanovich, N.E. Salas, A.L. Martino & I.E. di Tada. 2011. Reproductive Ecology of the Common South American Toad Rhinella arenarum (Anura: Bufonidae): Reproductive Effort, Clutch Size, Fecundity, and Mate Selection. J. Herp. 45: 261-264. [ Links ]
Blanco, S., L. Ector & E. Becares. 2004. Epiphytic Diatoms as water quality indicators in Spanish shallow lakes. Vie Milieu 54: 71-79. [ Links ]
Boone, M.D. & R.D. Semlitsch. 2001. Interactions of insecticide with larval density and predation in experimental amphibian communities. Conserv. Biol. 15: 228-238. [ Links ]
Boone, M.D., R.D. Semlitsch, J.F. Fairchild & B.B. Rothermel. 2004. Effects of an insecticide on amphibians in large-scale experimental ponds. Ecol. Appl. 14: 685-691. [ Links ]
Bridarolli, M.E. & I.E. di Tada. 1996. Algunos aspectos de la geografia fisica de la provincia de Cordoba, p. 15-38. In I.E. di Tada & E.H. Bucher (eds.). Biodiversidad de la provincia de Cordoba. Fauna. Vol. 1. Universidad Nacional de Rio Cuarto, Cordoba, Argentina. [ Links ]
Capitanelli, R.G. 1979. III. Clima, p. 45-138. In R.A.Vazquez, R.A. Miatello & M.E. Roque (eds.). Geografia Fisica de la provincia de Cordoba. Boldt, Buenos Aires, Argentina. [ Links ]
Carey, C. & C.J. Bryant. 1995. Possible interrelations among environmental toxicants, Amphibian development, and decline of Amphibian populations. Environ. Health Persp. 103: 13-17. [ Links ]
Carr, J., A. Gentles, E.E. Smith, W. Goleman, L. Urquidi, K. Thuett, J.R. Kendall, J.P. Giesy, T.S. Gross, K.R. Solomon & G. Van Der Kraak. 2003. Response of larval Xenopus laevis to atrazine: assessment of growth, metamorphosis, and gonadal and laryngeal morphology. Environ. Toxicol. Chem. 22: 396-405. [ Links ]
Cei, J.M. 1980. Amphibians of Argentina. Monit. Zool. Ital. Monogr. 2, Nuova Serie, Firenze, Italia. [ Links ]
Coady, K.K., M.B. Murphy, D.L. Villeneuve, M. Hecker, P.D. Jones, J.A. Carr, K.R. Solomon, E.E. Smith, G. Van Der Kraak, R.J. Kendall & J.P. Giesy. 2005. Effects of atrazine on metamorphosis, growth, laryngeal and gonadal development, aromatase activity, and sex steroid concentrations in Xenopus laevis. Ecotox. Environ. Safe. 62: 160-173. [ Links ]
Collins, J.P. & M. Crump. 2009. Extinction in Our Time. Oxford University, Nueva York, Nueva York, EEUU. [ Links ]
Colwell, R.K. 1997. EstimateS: Statistical estimation of species richness and shared species from samples. Version 5. User’s Guide and application published. (Consultado: 22 agosto 2011, http://viceroy.eeb.uconn.edu/estimates). [ Links ]
Davidson, C., H.B. Shaffer & M.R. Jennings. 2002. Spatial tests of the pesticide drift, habitat destruction, UV-B, and climate-change hypotheses for California amphibian declines. Conserv. Biol. 16: 1588-1601. [ Links ]
di Tada, I.E., M.V. Zavattieri, M.E. Bridarolli, N.E. Salas & A.L. Martino. 1996. Anfibios Anuros de la provincia de Cordoba, p. 191-215. In I.E. di Tada & E.H. Bucher (eds.). Biodiversidad de la provincia de Cordoba. Fauna. Vol. 1. Universidad Nacional de Rio Cuarto, Cordoba, Argentina. [ Links ]
Diaz Quiros, C. & C.A. Rivera Rondón. 2004. Diatomeas de pequenos rios andinos y su utilizacion como indicadoras de condiciones ambientales. Caldasia 26: 381-394. [ Links ]
Doffo, N. 1989. Geomorfologia del area urbana de la ciudad de Rio Cuarto y de la cuenca del arroyo El Banado, algunas consideraciones aplicadas al manejo del medio natural. Tesis de grado, Universidad Nacional de Rio Cuarto, Cordoba, Argentina. [ Links ]
Echeverria, D.D., A.V. Volpedo & V.I. Mascitti. 2007. Diet of tadpoles from a pond in Iguazu National Park, Argentina. Gayana 71: 8-14. [ Links ]
Fabrizi, L. 2010. Eutrophication of water bodies introduction. Lenntech. Water treatment solutions. (Consultado: 12 agosto 2010, http://www.lenntech.com/espanol/Eutroficacion-de-las-aguas/introduccion.htm). [ Links ]
GAA (Global Amphibian Assessment). 2004. Summary of key findings. (Consultado: 10 abril 2007, http://www.globalamphibians.org). [ Links ]
Gosner, K.L. 1960. A simplified table for stading anuran embryos and larvae with notes on identification. Herpetologica 16: 183-190. [ Links ]
Gray, M.J. & L.M. Smith. 2005. Influence of land use on postmetamorphic body size of playa lake amphibians. J. Wildlife Manage. 69: 515-524. [ Links ]
Hatch, A.C. & A.R. Blaustein. 2000. Combined effects of UV-B, nitrate, and low pH reduce the survival and activity level of larval Cascades frogs (Rana cascadae). Arch. Environ. Con. Tox. 39: 494-499. [ Links ]
Heyer, W.R. 1974. Niche measurements of frog larvae from a seasonal tropical location in Thailand. Ecology Tempe 55: 651-656. [ Links ]
Hill, B.H., A.T. Herlihy, P.R. Kaufmann, R.J. Stevenson, F.H. Mccormick & C.B. Johnson. 2000. Use of periphyton assemblage data as an index of biotic integrity.
JNABS 19: 50-67. [ Links ]
Jakob, E.M., S.D. Marshall & G.W. Uetz. 1996. Estimating fitness: a comparison of body condition indices. Oikos 77: 61-67. [ Links ]
Juttner, I., H. Rothfritz & S.J. Ormerod. 1996. Diatoms as indicators of river quality in the Nepalese Middle Hills with consideration of the effects of habitatspecific sampling. Freshwater Biol. 36: 475-486. [ Links ]
Kelly, M.G., H. Bennion, E.J. Cox, B. Goldsmith, J. Jamieson, S. Juggins D.G. Mann & R.J. Telford. 2005. Common freshwater diatoms of Britain and Ireland: an interactive key. Environment Agency, Bristol, Inglaterra. (Consultado: 20 enero 2011, http://craticula.ncl.ac.uk/EADiatomKey/html/index.html). [ Links ]
Kupferberg, S.J. 1997. The role of larval diet in anuran metamorphosis. Am. Zool. 37: 146-159. [ Links ]
Lajmanovich, R.C. 2000. Interpretacion ecologica de una comunidad larvaria de anfibios anuros. Interciencia 25: 71-79. [ Links ]
Lüddecke, H. 2002. Variation and trade-off in reproductive output of the Andean frog Hyla labialis. Oecologia 130: 403-410. [ Links ]
Magurran, A.E. 1987. Ecological diversity and its measurements. Vedra, Princeton, Nueva Jersey, EEUU. [ Links ]
Marco, A., C. Quilchano & A.R. Blaustein. 1999. Sensitivity to nitrate and nitrite in pond-breeding amphibians from the Pacific Northwest. Environ. Toxicol. Chem. 18: 2836-2839. [ Links ]
Nather Khan, I.S.A. 1990. Assessment of water pollution using Diatom community structure and species distribution. A case study in a tropical river basin. Int. Revue ges. Hydrobiol. 75: 317-338. [ Links ]
Ongley, E.D. 1997. Lucha contra la contaminacion agrícola de los recursos hidricos. Cuadernos de Riego y Drenaje N 55. FAO, Roma, Italia. (Consultado: 15 septiembre 2010, http://www.fao.org/docrep/W2598S/W2598S00.htm). [ Links ]
Peltzer, P.M., R.C. Lajmanovich, J.C. Sanchez Hernandez, M. Cabagna, A.M. Attademo ; A. Basso. 2008. Effects of agricultural pond eutrophication on survival and health status of Scinax nasicus tadpoles. Ecotox. Environ. Safe. 70: 185-197. [ Links ]
Perdomo, C.H., O.N. Casanova & V.S. Ciganda. 2001. Contaminacion de aguas subterraneas con nitratos y coliformes en el litoral sudoeste del Uruguay. Agrociencia V: 10-22. [ Links ]
Ramirez, C.A.M. & Y. Plata Diaz. 2008. Diatomeas perifiticas en diferentes tramos de dos sistemas loticos de alta montana (paramo de Santurban, norte de Santander, Colombia) y su relacion con las variables ambientales. Acta Biol. Colom. 13: 199-216. [ Links ]
Rossa Feres, D., J. Jim & M.G. Fonseca. 2004. Diets of tadpoles from a temporary pond in southeastern Brazil (Amphibia, Anura). Rev. Bras. Zool. 21: 745-754. [ Links ]
Rossi, R.L. 2006. Impactos recientes de la soja en la Argentina. Panorama productivo del cultivo. Agromercado 129: 4-7. [ Links ]
Round, F.E., R.M. Crawford & D.G. Mann. 1993. The diatoms. Biology & Morfology of the genera. Cambridge University, Cambridge, Massacchusetts, EEUU. [ Links ]
Semlitsch, R.D., D.E. Scott & H.K. Pechmann. 1988. Time and size at metamorphosis related to adult fitness in Ambystoma talpoideum. Ecology 69: 184-192. [ Links ]
Shannon, C.E. & W. Weaver. 1949. The mathematical theory of communications. Universidad de Illinois, Urbana, Illinois, EEUU. [ Links ]
Smol, J.P. 2002. Pollution of lakes and rivers: A paleoenvironmental perspective. Arnold, Londres, Inglaterra; copublicado por Oxford University, Nueva York,
Nueva York, EEUU. [ Links ]
Stuart, S.N., J.S. Chanson, N.A. Cox, B.E. Young, A.S.L. Rodrigues, D.L. Fischman & R.W. Waller. 2004. Status and trends of amphibian declines and extinctions worldwide. Science 306: 1783-1786. [ Links ]
Taylor, B., D. Skelly, L.K. Demarchis, M.D. Slade, D. Galusha & P.M. Rabinowitz. 2005. Proximity to pollution sources and risk of amphibian limb malformation.
Environ. Health Persp. 113: 1497-1501. [ Links ]
USEPA, United States Environmental Protection Agency. 1994. National Water Quality Inventory. 1992 Report to Congress. EPA-841-R-94-001. Office of Water,
Washington D.C., EEUU. [ Links ]
Villafañe, V.E. & F.M.H. Reid. 1995. Métodos de microscopia para la cuantificación del fitoplancton, p. 169-185. In K. Alveal, M.E. Ferrario, E.C. Oliveira & E. Sar (eds.). Manual de Métodos Ficológicos. Universidad de Concepción, Concepción, Chile. [ Links ]
Wake, D.B. 1998. Action on amphibians. Trends Ecol. Evol. 13: 379-380. [ Links ]
Wilbur, H.M. 1980. Complex life cycle. Annu. Rev. Ecol. Syst. 11: 67-93. [ Links ]
Yucra, H.A. & P.M. Tapia. 2008. El uso de microalgas como bioindicadoras de polución orgánica en Brisas de Oquendo, Callao, Perú. Biologist (Lima) 6: 41-47. [ Links ]
*Correspondencia a: Clarisa Bionda, Nancy Salas & Adolfo Martino: Ecología, Departamento de Ciencias Naturales, Facultad de Ciencias Exactas, Físico-Químicas y Naturales, UNRC, ruta 36km 601, Río Cuarto, Córdoba, Argentina; cbionda@exa.unrc.edu.ar, nsalas@exa.unrc.edu.ar, amartino@exa.unrc.edu.ar
Noemi Gari & Elisa Luque: Botánica Sistemática, Departamento de Ciencias Naturales, Facultad de Ciencias Exactas, Fisico-Quimicas y Naturales, UNRC; ngari@exa.unrc.edu.ar, mluque@exa.unrc.edu.ar
Rafael Lajmanovich: Ecotoxicologia, Escuela Superior de Sanidad “Dr. Ramón Carrillo”, Facultad de Bioquímica y Ciencias Biológicas, Paraje “El Pozo” s/n, Santa Fe, Argentina; lajmanovich@hotmail.com
Clarisa Bionda: Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET), Argentina
1. Ecología, Departamento de Ciencias Naturales, Facultad de Ciencias Exactas, Fisico-Quimicas y Naturales, UNRC, ruta 36km 601, Rio Cuarto, Córdoba, Argentina; cbionda@exa.unrc.edu.ar, nsalas@exa.unrc.edu.ar, amartino@exa.unrc.edu.ar
2. Botánica Sistemática, Departamento de Ciencias Naturales, Facultad de Ciencias Exactas, Fisico-Quimicas y Naturales, UNRC; ngari@exa.unrc.edu.ar, mluque@exa.unrc.edu.ar
3. Ecotoxicologia, Escuela Superior de Sanidad “Dr. Ramón Carrillo”, Facultad de Bioquímica y Ciencias Biológicas, Paraje “El Pozo” s/n, Santa Fe, Argentina; lajmanovich@hotmail.com
4. Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET), Argentina
Recibido 01-VI-2011. Corregido 13-X-2011. Aceptado 15-XI-2011.


{kind=link}
{kind=link}